Poplar Short Rotation Coppice Plantations under Mediterranean Conditions: The Case of Spain

 , ,
, , 
Abstract
1. Introduction
2. Short Rotation Forest Crops for the Production of Biomass: The Populus Genus
3. Lines of Progress: State of the Art
3.1. Suitability of Genetic Material
Clonal Selection
3.2. Plantation Design
3.2.1. Densities
3.2.2. Rotation Lengths and Number of Coppices
3.2.3. Species Mixtures
3.3. Management: Cultural Treatments
3.3.1. Control of Competing Vegetation
3.3.2. Fertilization
3.3.3. Control of Pests and Diseases
3.3.4. Irrigation
3.4. Production
Modeling of Growth, Production, and Biometric Relationships in SRC Poplar Plantations
3.5. Biomass Characterization
3.6. Sustainability and Ecosystem Services
4. Current Status and Future Prospects in Spain
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Commission. The European Green Deal; European Commission: Brussels, Belgium, 2019; p. 24. [Google Scholar]
- Hetemäki, L.; Hanewinkel, M.; Muys, B.; Ollikainen, M.; Palahí, M.; Trasobares, A. Leading the Way to a European Circular Bioeconomy Strategy; Science to Policy 5; European Forest Institute: Joensuu, Finland, 2017; Volume 5. [Google Scholar]
- European Commission. Innovating for Sustainable Growth: A Bioeconomy for Europe; European Commision: Brussels, Belgium, 2012. [Google Scholar]
- European Commission. Review of the 2012 European Bioeconomy Strategy; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- MINECO. Estrategia Española de Bioeconomía: Horizonte 2030; Ministerio de Economía y Competitividad, Secretaría de Estado de Investigación, Desarrollo e Innovación: Madrid, Spain, 2015; p. 46. [Google Scholar]
- MITECO. Law on Climate Change and Energy Transition; Ministerio para la Transición Ecológica y el Reto Demográfico: Madrid, Spain, 2020. [Google Scholar]
- Bioplat. Agenda Estratégica de Investigación e Innovación. Biomasa y Bioeconomía; Ministerio de Ciencia e Innovación, Gobierno de España: Madrid, Spain, 2020. [Google Scholar]
- Sancho, A. Día de la Bioenergía en España: 28 días de Independencia Energética; Bioenergy International: Stockholm, Sweden, 2018. [Google Scholar]
- Gardiner, E.S.; Ghezehei, S.B.; Headlee, W.L.; Richardson, J.; Soolanayakanahally, R.Y.; Stanton, B.J.; Zalesny, R.S., Jr. The 2018 Woody Crops International Conference, Rhinelander, Wisconsin, USA, 22–27 July 2018. Forests 2018, 9, 693. [Google Scholar] [CrossRef]
- Broeckx, L.; Verlinden, M.; Vangronsveld, J.; Ceulemans, R. Importance of crown architecture for leaf area index of different Populus genotypes in a high-density plantation. Tree Physiol. 2012, 32, 1214–1226. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.A.; May, K.; Hülsbergen, K.-J. Allometric tree biomass models of various species grown in short-rotation agroforestry systems. Eur. J. For. Res. 2017, 136, 75–89. [Google Scholar] [CrossRef]
- Weih, M. Intensive short rotation forestry in boreal climates: Present and future perspectives. Can. J. For. Res. Rev. Can. Rech. For. 2004, 34, 1369–1378. [Google Scholar] [CrossRef]
- Aylott, M.J.; Casella, E.; Tubby, I.; Street, N.R.; Smith, P.; Taylor, G. Yield and spatial supply of bioenergy poplar and willow short-rotation coppice in the UK. New Phytol. 2008, 178, 358–370. [Google Scholar] [CrossRef]
- Berhongaray, G.; Verlinden, M.S.; Broeckx, L.S.; Janssens, I.A.; Ceulemans, R. Soil carbon and belowground carbon balance of a short-rotation coppice: Assessments from three different approaches. Glob. Chang. Biol. Bioenergy 2016, 9, 299–313. [Google Scholar] [CrossRef]
- European Commission. State of Play on the Sustainability of Solid and Gaseous Biomass Used for Electricity, Heating and Cooling in the EU (SWD 259 Final); European Commission: Brussels, Belgium, 2014. [Google Scholar]
- Dimitriou, I.; Berndes, G.; Englund, O.; Murphy, F. Lignocellulosic Crops in Agricultural Landscapes: Production Systems for Biomass and Other Environmental Benefits–Examples, Incentives, and Barriers; IEA Bioenergy: Paris, France, 2018. [Google Scholar]
- Dickmann, D.I. An overview of the genus Populus. In Poplar Culture in North America; Dickmann, D.I., Isebrands, J.G., Eckenwalder, J.E., Richardson, J., Eds.; NRC Research Press: Ottawa, ON, Canada, 2001; pp. 1–42. [Google Scholar]
- Al Afas, N.; Marron, N.; Van Dongen, S.; Laureysens, I.; Ceulemans, R. Dynamics of biomass production in a poplar coppice culture over three rotations (11 years). For. Ecol. Manag. 2008, 255, 1883–1891. [Google Scholar] [CrossRef]
- Laureysens, I.; Pellis, A.; Willems, J.; Ceulemans, R. Growth and production of a short rotation coppice culture of poplar. III. Second rotation results. Biomass Bioenergy 2005, 29, 10–21. [Google Scholar] [CrossRef]
- Ceulemans, R.; McDonald, A.; Pereira, J. A comparison among eucalypt, poplar and willow characteristics with particular reference to a coppice, growth-modelling approach. Biomass Bioenergy 1996, 11, 215–231. [Google Scholar] [CrossRef]
- Patel, B.; Gami, B. Biomass characterization and its use as solid fuel for combustion. Iran. J. Energy Environ. 2012, 3, 123–128. [Google Scholar] [CrossRef]
- Zhang, X.; Tu, M.; Paice, M. Routes to Potential Bioproducts from Lignocellulosic Biomass Lignin and Hemicelluloses. Bioenergy Res. 2011, 4, 246–257. [Google Scholar] [CrossRef]
- Surendran Nair, S.; Kang, S.; Zhang, X.; Miguez, F.E.; Izaurralde, R.C.; Post, W.M.; Dietze, M.C.; Lynd, L.R.; Wullschleger, S.D. Bioenergy crop models: Descriptions, data requirements, and future challenges. Glob. Chang. Biol. Bioenergy 2012, 4, 620–633. [Google Scholar] [CrossRef]
- Rodrigues, A.; Loureiro, L.; Nunes, L.J.R. Torrefaction of woody biomasses from poplar SRC and Portuguese roundwood: Properties of torrefied products. Biomass Bioenergy 2018, 108, 55–65. [Google Scholar] [CrossRef]
- Schultz, R.; Isenhart, T.; Simpkins, W.; Colletti, J. Riparian forest buffers in agroecosystems–lessons learned from the Bear Creek Watershed, central Iowa, USA. Agrofor. Syst. 2004, 61, 35–50. [Google Scholar] [CrossRef]
- Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. Fine root dynamics, coarse root biomass, root distribution, and soil respiration in a multispecies riparian buffer in Central Iowa, USA. Agrofor. Syst. 1998, 44, 163–174. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; Headlee, W.L.; Banuelos, G.S.; Hass, A. Ecosystem services of woody crop production systems. Bioenergy Res. 2016, 9, 465–491. [Google Scholar] [CrossRef]
- Coleman, M.D.; Friend, A.L.; Kern, C.C. Carbon allocation and nitrogen acquisition in a developing Populus deltoides plantation. Tree Physiol. 2004, 24, 1347–1357. [Google Scholar] [CrossRef]
- Grigal, D.F.; Berguson, W.E. Soil carbon changes associated with short-rotation systems. Biomass Bioenergy 1998, 14, 371–377. [Google Scholar] [CrossRef]
- Baum, S.; Bolte, A.; Weih, M. Short rotation coppice (SRC) plantations provide additional habitats for vascular plant species in agricultural moisac landscapes. Bioenerg. Res. 2012, 5, 573–583. [Google Scholar] [CrossRef][Green Version]
- Christian, D.P.; Niemi, G.J.; Hanowski, J.M.; Collins, P. Perspectives on biomass energy tree plantations and changes in habitat for biological organisms. Biomass Bioenergy 1994, 6, 31–39. [Google Scholar] [CrossRef]
- Vanbeveren, S.P.P.; Ceulemans, R. Biodiversity in short-rotation coppice. Renew. Sustain. Energy Rev. 2019, 111, 34–43. [Google Scholar] [CrossRef]
- Ferrarini, A.; Serra, P.; Almagro, M.; Trevisan, M.; Amaducci, S. Multiple ecosystem services provision and biomass logistics management in bioenergy buffers: A state-of-the-art review. Renew. Sustain. Energy Rev. 2017, 73, 277–290. [Google Scholar] [CrossRef]
- Rockwood, D.; Naidu, C.; Carter, D.; Rahmani, M.; Spriggs, T.; Lin, C.; Alker, G.; Isebrands, J.; Segrest, S. Short-rotation woody crops and phytoremediation: Opportunities for agroforestry? In New Vistas in Agroforestry. A Compendium for the 1st World Congress of Agroforestry; Kluwer Academic Publisher: Dordrecht, The Netherlands; Boston, MA, USA; New York, NY, USA, 2004; pp. 51–63. [Google Scholar]
- Dipesh, K.; Will, R.E.; Hennessey, T.C.; Penn, C.J. Evaluating performance of short-rotation woody crops for bioremediation purposes. New For. 2015, 46, 267–281. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.S., Jr.; Orlović, S.; Drekić, M.; Pekeč, S.; Katanić, M.; Poljaković-Pajnik, L. Growth and physiological responses of three poplar clones grown on soils artificially contaminated with heavy metals, diesel fuel, and herbicides. Int. J. Phytoremediat. 2020, 22, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Bañuelos, G.S.; Hallett, R.A.; Hass, A.; Stange, C.M.; Perdue, J.H.; Young, T.M.; Coyle, D.R. Environmental technologies of woody crop production systems. Bioenergy Res. 2016, 9, 492–506. [Google Scholar] [CrossRef]
- Evangelou, M.W.; Conesa, H.M.; Robinson, B.H.; Schulin, R. Biomass production on trace element–contaminated land: A review. Biomass Bioenergy 2012, 29, 823–839. [Google Scholar] [CrossRef]
- IRENA. Global Renewables Outlook: Energy Transformation 2050; International Renewable Energy Agency: Abu Dhabi, UAE, 2020; p. 212. [Google Scholar]
- GBEP. The Global Bioenergy Partnership Sustainability Indicators for Bioenergy; Climate, Energy and Tenure Division, Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2011. [Google Scholar]
- Marron, N. Agronomic and environmental effects of land application of residues in short-rotation tree plantations: A literature review. Biomass Bioenergy 2015, 81, 378–400. [Google Scholar] [CrossRef]
- de Miguel, A.; Meffe, R.; Leal, M.; González-Naranjo, V.; Martínez-Hernández, V.; Lillo, J.; Martín, I.; Salas, J.J.; Bustamante, I.d. Treating municipal wastewater through a vegetation filter with a short-rotation poplar species. Ecol. Eng. 2014, 73, 560–568. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Evett, S.R.; Kandil, N.F.; Soriano, C. Opportunities for woody crop production using treated wastewater in Egypt. I. Afforestation strategies. Int. J. Phytoremediat. 2011, 13, 102–121. [Google Scholar] [CrossRef]
- San Miguel, A.; Montoya, J. Resultados de los primeros 5 años de produccion de talleres de chopo en rotacion corta (2–5 anõs). In Anales del Instituto Nacional de Investigaciones Agrarias. Serie Forestal; Instituto Nacional de Investigaciones Agrarias: Madrid, Spain, 1984; pp. 73–91. [Google Scholar]
- Marcos, F. Cultivos energéticos forestales. In Proceedings of the V Conferencia sobre Planificación, Ahorro y Alternativas Energéticas, Zaragoza, Spain, 1985. [Google Scholar]
- Ciria, M.P.; Mazon, P.; Carrasco, J. Productivity of poplar grown on short rotation in Spain: Influence of the rotation age and plant density. In Proceedings of the 2nd biomass conference of the Americas: Energy, Environment, Agriculture, and Industry, Portland, OR, USA, 21–24 August 1995; pp. 235–243. [Google Scholar]
- Pérez-Cruzado, C.; Sanchez-Ron, D.; Rodríguez-Soalleiro, R.; Hernández, M.; Sánchez-Martín, M.; Cañellas, I.; Sixto, H. Biomass production assessment from Populus spp. short-rotation irrigated crops in Spain. Glob. Chage Biol. Bioenergy 2013, 6, 312–326. [Google Scholar] [CrossRef]
- Huber, J.A.; May, K.; Siegl, T.; Schmid, H.; Gerl, G.; Hülsbergen, K.J. Yield potential of tree species in organic and conventional short-rotation agroforestry systems in Southern Germany. Bioenergy Res. 2016, 9, 955–968. [Google Scholar] [CrossRef]
- Havlíčková, K.; Weger, J.; Zánová, I. Short rotation coppice for energy purposes—Economy conditions and landscape functions in the Czech Republic. In Proceedings of ISES World Congress 2007; Springer: Berlin/Heidelberg, Germany, 2008; pp. 2482–2487. [Google Scholar]
- Stankova, T.; Gyuleva, V.; Tsvetkov, I.; Popov, E.; Velinova, K.; Velizarova, E.; Dimitrov, D.N.; Hristova, H.; Kalmukov, K.; Dimitrova, P. Aboveground dendromass allometry of hybrid black poplars for energy crops. Ann. For. Res. 2016, 59, 61–74. [Google Scholar] [CrossRef]
- Pilipović, A.; Orlović, S.; Kovačević, B.; Galović, V.; Stojnić, S. Selection and Breeding of Fast Growing Trees for Multiple Purposes in Serbia. In Forests of Southeast Europe Under a Changing Climate; Springer: Berlin/Heidelberg, Germany, 2019; pp. 239–249. [Google Scholar]
- Niemczyk, M.; Kaliszewski, A.; Jewiarz, M.; Wróbel, M.; Mudryk, K. Productivity and biomass characteristics of selected poplar (Populus spp.) cultivars under the climatic conditions of northern Poland. Biomass Bioenergy 2018, 111, 46–51. [Google Scholar] [CrossRef]
- Benbrahim, M.; Gavaland, A.; Gauvin, J. Growth and yield of mixed polyclonal stands of Populus in short-rotation coppice. Scand. J. For. Res. 2000, 15, 605–610. [Google Scholar] [CrossRef]
- Nardin, F.; Alasia, F. Use of selected fast growth poplar trees for a woody biomass production die along Po valley. In Proceedings of the 2nd World Conference on Biomass for Energy, Industry and Climate Protection, Rome, Italy, 10–14 May 2004; pp. 247–249. [Google Scholar]
- Sixto, H.; Gil, P.; Ciria, P.; Camps, F.; Sánchez, M.; Cañellas, I.; Voltas, J. Performance of hybrid poplar clones in short rotation coppice in Mediterranean environments: Analysis of genotypic stability. Bioenegy 2013, 6, 661–671. [Google Scholar] [CrossRef]
- Nordborg, M.; Berndes, G.; Dimitriou, I.; Henriksson, A.; Mola-Yudego, B.; Rosenqvist, H. Energy analysis of poplar production for bioenergy in Sweden. Biomass Bioenergy 2018, 112, 110–120. [Google Scholar] [CrossRef]
- Tullus, A.; Rosenvald, K.; Lutter, R.; Kaasik, A.; Kupper, P.; Sellin, A. Coppicing improves the growth response of short-rotation hybrid aspen to elevated atmospheric humidity. Forest Ecol. Manag. 2020, 459, 117825. [Google Scholar] [CrossRef]
- Clifton-Brown, J.; Harfouche, A.; Casler, M.D.; Dylan Jones, H.; Macalpine, W.J.; Murphy-Bokern, D.; Smart, L.B.; Adler, A.; Ashman, C.; Awty-Carroll, D. Breeding progress and preparedness for mass-scale deployment of perennial lignocellulosic biomass crops switchgrass, miscanthus, willow and poplar. Glob. Chang. Biol. Bioenergy 2019, 11, 118–151. [Google Scholar] [CrossRef]
- Scholz, V.; Ellerbrock, R. The growth productivity, and environmental impact of the cultivation of energy crops on sandy soil in Germany. Biomass Bioenergy 2002, 23, 81–92. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Yu, Q.; Chauret, G.; Koubaa, A. Selection for both growth and wood properties in hybrid poplar clones. For. Sci. 2003, 49, 901–908. [Google Scholar] [CrossRef]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- FAO. Working Party on Sustainable Livelihoods, Land-Use, Products and Bioenergy. Available online: http://www.fao.org/forestry/ipc/69631/en/ (accessed on 15 September 2020).
- Glas, D.; Schimer, R. Prüfung von Pappelsorten aus Anderen EU-Staaten für Kurzumtriebsplantagen (EU-POP); Bundesministerium für Ernährung und Landwirtschaft: Teisendorf, Germany, 2018. [Google Scholar]
- MAPA. Estadísticas Comisión Nacional del Chopo. Available online: https://www.mapa.gob.es/es/desarrollo-rural/temas/politica-forestal/comision-nacional-del-chopo/estadisticas.aspx (accessed on 20 September 2020).
- ProPopulus. La Escasez de Chopo Amenaza a la Industria Española de Contrachapado; ProPopulus; Nature, Society, Future; The European Poplar Initiative: Spain, 2019; Available online: http://propopulus.eu/es/la-escasez-de-chopo-amenaza-a-la-industria-espanola-de-contrachapado/ (accessed on 10 December 2020).
- Rueda, J.; García Caballero, J.L.; Cuevas, Y.; García-Jiménez, C.; Villar, C. Cultivo de Chopos en Castilla y León; Consejería de Fomento y Medio Ambiente, Junta de Castilla y León: Valladolid, Spain, 2019. [Google Scholar]
- ESYRCE. Encuesta Sobre Superficies y Rendimientos de Cultivos. Informe Sobre Regadíos en España; Subsecretaría de Agricultura, Pesca y Alimentación y Subdirección General de Análisis, Coordinación y Estadística: Madrid, Spain, 2019. [Google Scholar]
- Sixto, H.; Hernández, M.; Barrio, M.; Carrasco, J.; Cañellas, I. Plantaciones del género Populus para la producción de biomasa con fines energéticos: Revisión. For. Syst. 2007, 16, 277–294. [Google Scholar]
- Corcuera, L.; Maestro, C.; Notivol, E. La ecofisiología como herramienta para la selección de clones más adaptados y productivos en el marco de una selvicultura clonal con chopos. Invest. Agrar. Sist. Recur. For. 2005, 14, 394–407. [Google Scholar] [CrossRef][Green Version]
- Pérez-Cruzado, C.; Merino, A.; Rodríguez-Soalleiro, R. A management tool for estimating bioenergy production and carbon sequestration in Eucalyptus globulus and Eucalyptus nitens grown as short rotation woody crops in north-west Spain. Biomass Bioenerg 2011, 35, 2839–2851. [Google Scholar] [CrossRef]
- Pita, P.; Soria, F.; Canas, I.; Toval, G.; Pardos, J. Carbon isotope discrimination and its relationship to drought resistance under field conditions in genotypes of Eucalyptus globulus Labill. Forest Ecol. Manag. 2001, 141, 211–221. [Google Scholar] [CrossRef]
- Valdés, A.E.; Irar, S.; Majada, J.P.; Rodríguez, A.; Fernández, B.; Pagès, M. Drought tolerance acquisition in Eucalyptus globulus (Labill.): A research on plant morphology, physiology and proteomics. J. Proteom. 2013, 79, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Perez, I.; Perez, J.; Carrasco, J.; Ciria, P. Siberian elm responses to different culture conditions under short rotation forestry in Mediterranean areas. Turk. J. Agric. For. 2014, 38, 652–662. [Google Scholar] [CrossRef]
- Manzone, M.; Bergante, S.; Facciotto, G. Energy and economic sustainability of woodchip production by black locust (Robinia pseudoacacia L.) plantations in Italy. Fuel 2015, 140, 555–560. [Google Scholar] [CrossRef]
- Böhm, C.; Quinkenstein, A.; Freese, D. Yield prediction of young black locust (Robinia pseudoacacia L.) plantations for woody biomass production using allometric relations. Ann. For. Res. 2011, 54, 215–227. [Google Scholar]
- Oliveira, N.; del Río, M.; Forrester, D.I.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Cañellas, I.; Sixto, H. Mixed short rotation plantations of Populus alba and Robinia pseudoacacia for biomass yield. For. Ecol. Manag. 2018, 410, 48–55. [Google Scholar] [CrossRef]
- Sixto, H.; Cañellas, I.; van Arendonk, J.; Ciria, P.; Camps, F.; Sánchez, M.; Sánchez-González, M. Growth potential of different species and genotypes for biomass production in short rotation in Mediterranean environments. For. Ecol. Manag. 2015, 354, 291–299. [Google Scholar] [CrossRef]
- Rosso, L.; Facciotto, G.; Bergante, S.; Vietto, L.; Nervo, G. Selection and testing of Populus alba and Salix spp. as bioenergy feedstock: Preliminary results. Appl. Energy 2013, 102, 87–92. [Google Scholar] [CrossRef]
- Karp, A.; Shield, I. Bioenergy from plants and the sustainable yield challenge. New Phytol. 2008, 179, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, N.; Ciria, P.; Camps, F.; Sánchez-González, M.; Sánchez, M.; Cañellas, I.; Sixto, H. Assessing genotypes in second rotation for lignocellulosic biomass production. In Proceedings of the 23rd European Biomass Conference and Exhibition (EUBCE), Viena, Austria, 1–4 June 2015. [Google Scholar]
- Sixto, H.; Calvo, R.; Sánchez, M.M.; Arrieta, J.A.; Otero, J.M.; Salvia, J.; Cañellas, I. Fine scale site variation correlated to growth in a Salicaceae plantations (Salix and Populus) during the first vegetative period. In Proceedings of the 24th International Poplar Commission. Improving lives with poplars and willows, Dehradun, India, 30 October–2 November 2012. [Google Scholar]
- Zuazo, V.H.D.; Bocanegra, J.A.J.; Torres, F.P.; Pleguezuelo, C.R.R.; Martínez, J.R.F. Biomass yield potential of Paulownia trees in a semi-arid Mediterranean environment (Spain). Int. J. Renew. Energy Res. 2013, 3, 789–793. [Google Scholar]
- García-Morote, F.A.; López-Serrano, F.R.; Martínez-García, E.; Andrés-Abellán, M.; Dadi, T.; Candel, D.; Rubio, E.; Lucas-Borja, M.E. Stem biomass production of Paulownia elongata × P. fortunei under low irrigation in a semi-arid environment. Forests 2014, 5, 2505–2520. [Google Scholar] [CrossRef]
- Hernández Garasa, M.J.; Sixto, H.; Ciria, P.; Carrasco, J.; Cañellas, I. Paulownia plantations for bioenergy in Spain. In Proceedings of the 16th European Biomass confernce and Exhibition, Valencia, Spain, 2–6 May 2008. [Google Scholar]
- Bradshaw, A.D. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 1965, 13, 115–155. [Google Scholar]
- Zalesny, R.S., Jr.; Hall, R.B.; Zalesny, J.A.; McMahon, B.G.; Berguson, W.E.; Stanosz, G.R. Biomass and genotype × environment interactions of Populus energy crops in the Midwestern United States. Bioenergy Res. 2009, 2, 106–122. [Google Scholar] [CrossRef]
- Mohn, C.; Randall, W. Interaction of cottonwood clones with site and planting year. Can. J. For. Res. 1973, 3, 329–332. [Google Scholar] [CrossRef]
- Orlovic, S.; Guzina, V.; Krstic, B.; Merkulov, L. Genetic variability in anatomical, physiological and growth characteristics of hybrid poplar (Populus × euramericana Dode (Guinier)) and eastern cottonwood (Populus deltoides Bartr.) clones. Silvae Genet. 1998, 47, 183–189. [Google Scholar]
- Becker, H.; Leon, J. Stability analysis in plant breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Bergante, S.; Facciotto, G.; Minotta, G. Identification of the main site factors and management intensity affecting the establishment of Short-Rotation-Coppices (SRC) in Northern Italy through stepwise regression analysis. Cent. Eur. J. Biol. 2010, 5, 522–530. [Google Scholar] [CrossRef]
- Li, Y.; Suontama, M.; Burdon, R.D.; Dungey, H.S. Genotype by environment interactions in forest tree breeding: Review of methodology and perspectives on research and application. Tree Genet. Genomes 2017, 13, 60. [Google Scholar] [CrossRef]
- Sabatti, M.; Fabbrini, F.; Harfouche, A.; Beritognolo, I.; Mareschi, L.; Carlini, M.; Paris, P.; Scarascia-Mugnozza, G. Evaluation of biomass production potential and heating value of hybrid poplar genotypes in a short-rotation culture in Italy. Ind. Crop. Prod. 2014, 61, 62–73. [Google Scholar] [CrossRef]
- Sixto, H.; Gil, P.M.; Ciria, P.; Camps, F.; Cañellas, I.; Voltas, J. Interpreting genotype-by-environment interaction for biomass production in hybrid poplars under short-rotation coppice in Mediterranean environments. Glob. Chang. Biol. Bioenergy 2016, 8, 1124–1135. [Google Scholar] [CrossRef][Green Version]
- Hernández, P.M.G.; Voltas, J.; González, M.S.; Martín, M.M.S.; Camps, F.; Ciria, M.P.; Cañellas, I.; Sixto, H. Análisis de estabilidad en genotipos híbridos de “Populus” L. para producción de biomasa. Cuad. Soc. Esp. Cienc. For. 2012, 105–110. [Google Scholar]
- Sixto, H.; Salvia, J.; Barrio, M.; Ciria, M.P.; Canellas, I. Genetic variation and genotype-environment interactions in short rotation Populus plantations in southern Europe. New For. 2011, 42, 163–177. [Google Scholar] [CrossRef]
- Stanton, B. Poplar hybridisation and clonal evaluation: James River, Lower Columbia River Fibber Farm, Westport Research Station. In Proceedings of the International Poplar Symposium, Seattle, WA, USA, 20–25 August 1995; pp. 114–115. [Google Scholar]
- Fladung, M.; Polak, O. Ac/Ds-transposon activation tagging in poplar: A powerful tool for gene discovery. BMC Genom. 2012, 13, 61. [Google Scholar] [CrossRef]
- Marchadier, H.; Sigaud, P. Los álamos en la investigación biotecnológica. Unasylva 2005, 56, 38–39. [Google Scholar]
- Fan, D.; Liu, T.; Li, C.; Jiao, B.; Li, S.; Hou, Y.; Luo, K. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 12217. [Google Scholar] [CrossRef]
- Zhou, X.; Jacobs, T.B.; Xue, L.J.; Harding, S.A.; Tsai, C.J. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate: CoA ligase specificity and redundancy. New Phytol. 2015, 208, 298–301. [Google Scholar] [CrossRef]
- Liu, T.; Fan, D.; Ran, L.; Jiang, Y.; Liu, R.; Luo, K. Highly efficient CRISPR/Cas9-mediated targeted mutagenesis of multiple genes in Populus. Yi Chuan 2015, 37, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Pilate, G.; Allona, I.; Boerjan, W.; Déjardin, A.; Fladung, M.; Gallardo, F.; Häggman, H.; Jansson, S.; Van Acker, R.; Halpin, C. Lessons from 25 years of GM tree field trials in Europe and prospects for the future. In Biosafety of Forest Transgenic Trees; Springer: Berlin/Heidelberg, Germany, 2016; pp. 67–100. [Google Scholar]
- Porth, I.; Klapšte, J.; Skyba, O.; Hannemann, J.; McKown, A.D.; Guy, R.D.; DiFazio, S.P.; Muchero, W.; Ranjan, P.; Tuskan, G.A. Genome-wide association mapping for wood characteristics in Populus identifies an array of candidate single nucleotide polymorphisms. New Phytologist. 2013, 200, 710–726. [Google Scholar] [CrossRef] [PubMed]
- Ludovisi, R.; Tauro, F.; Salvati, R.; Khoury, S.; Mugnozza Scarascia, G.; Harfouche, A. UAV-based thermal imaging for high-throughput field phenotyping of black poplar response to drought. Front. Plant Sci. 2017, 8, 1681. [Google Scholar] [CrossRef] [PubMed]
- Gömöry, D.; Hrivnák, M.; Krajmerová, D.; Longauer, R. Epigenetic memory effects in forest trees: A victory of “Michurinian biology”? Cent. Eur. For. J. 2017, 63, 173–179. [Google Scholar] [CrossRef][Green Version]
- Le Gac, A.-L.; Lafon-Placette, C.; Chauveau, D.; Segura, V.; Delaunay, A.; Fichot, R.; Marron, N.; Le Jan, I.; Berthelot, A.; Bodineau, G. Winter-dormant shoot apical meristem in poplar trees shows environmental epigenetic memory. J. Exp. Bot. 2018, 69, 4821–4837. [Google Scholar] [CrossRef]
- Jing, Z.P.; Gallardo, F.; Pascual, M.B.; Sampalo, R.; Romero, J.; De Navarra, A.T.; Cánovas, F.M. Improved growth in a field trial of transgenic hybrid poplar overexpressing glutamine synthetase. New Phytol. 2004, 164, 137–145. [Google Scholar] [CrossRef]
- Conde, D.; Moreno-Cortés, A.; Dervinis, C.; Ramos-Sánchez, J.M.; Kirst, M.; Perales, M.; González-Melendi, P.; Allona, I. Overexpression of DEMETER, a DNA demethylase, promotes early apical bud maturation in poplar. Plant Cell Environ. 2017, 40, 2806–2819. [Google Scholar] [CrossRef]
- Moreno-Cortés, A.; Hernández-Verdeja, T.; Sánchez-Jiménez, P.; González-Melendi, P.; Aragoncillo, C.; Allona, I. CsRAV1 induces sylleptic branching in hybrid poplar. New. Phytol. 2012, 194, 83–90. [Google Scholar] [CrossRef]
- Moreno-Cortés, A.; Ramos-Sánchez, J.M.; Hernández-Verdeja, T.; González-Melendi, P.; Alves, A.; Simões, R.; Rodrigues, J.C.; Guijarro, M.; Canellas, I.; Sixto, H. Impact of RAV1-engineering on poplar biomass production: A short-rotation coppice field trial. Biotechnol. Biofuels 2017, 10, 110. [Google Scholar] [CrossRef]
- Sixto, H. Populus alba clones and their hybrids growing under water deficit conditions for biomass production in short rotation forestry. In Proceedings of the VI International Poplar Symposium (IUFRO), Vancouver, BC, Canada, 20–28 July 2014. [Google Scholar]
- Marcos, F.; Izquierdo, I.; Gracia, R.; Godino, M.; Ruiz, J.; Vilegas, S. Estudio de plantaciones energéticas de chopo (Populus × euramericana cv. I-214) a turnos muy cortos; Documento interno del Departamento de Ingeniería Forestal ETSI de Montes: Madrid, Spain, 2002. [Google Scholar]
- San Miguel, A.; San Miguel, J.; Yagüe, S. Tallares de chopo a turno corto. In Proceedings of the 19ª Sesión de la Comisión Internacional del Álamo, Zaragoza, Spain, 22–25 September 1992; pp. 143–156. [Google Scholar]
- Sixto, H.; Grau, J.M.; García-Baudín, J.M. Assessment of the effect of broad-spectrum pre-emergence herbicides in poplar nurseries. Crop Prot. 2001, 20, 121–126. [Google Scholar] [CrossRef]
- Sixto, H.; Barrio, M.; Aranda, I. Evaluación de criterios para la selección de clones de chopo como productores de biomasa. In Proceedings of the XVII Reunión de la Sociedad Española de Fisiología Vegetal y X Congreso Hispano-Luso de Fisiología Vegetal Alcalá de Henares, Madrid, Spain, 18–21 September 2007. [Google Scholar]
- Sixto, H.; Grau, J.; Alba, N.; Alia, R. Response to sodium chloride in different species and clones of genus Populus L. Forestry 2005, 78, 93–104. [Google Scholar] [CrossRef]
- Álvarez-Álvarez, P.; Pizarro, C.; Barrio-Anta, M.; Cámara-Obregón, A.; Bueno, J.L.M.; Álvarez, A.; Gutiérrez, I.; Burslem, D.F. Evaluation of tree species for biomass energy production in Northwest Spain. Forests 2018, 9, 160. [Google Scholar] [CrossRef]
- Castaño Díaz, M. Producción de Biomasa Procedente de Cultivos Energéticos Leñosos en Terrenos Ociosos de Minería en el PRINCIPADO de Asturias. Ph.D. Thesis, Biología de Organismos y Sistemas, Departamento de, University of Oviedo, Oviedo, Spain, 2018. [Google Scholar]
- Fiala, M.; Bacenetti, J. Economic, energetic and environmental impact in short rotation coppice harvesting operations. Biomass Bioenergy 2012, 42, 107–113. [Google Scholar] [CrossRef]
- Cañellas, I.; Huelin, P.; Hernández, M.; Ciria, P.; Calvo, R.; Gea-Izquierdo, G.; Sixto, H. The effect of density on Short Rotation Populus spp. plantations in the Mediterranean area. Biomass Bioenergy 2012, 46, 645–652. [Google Scholar] [CrossRef]
- Hernández Garasa, M.; Montes Pita, F. Evolución del índice de área foliar en plantaciones de chopo con fines energéticos. Comparación entre métodos semidirectos e indirectos de estimación. In Proceedings of the 5° Congreso Forestal Español, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Sixto, H.; Rueda, J. Evaluación de genotipos para la producción de biomasa con fines energéticos en la Comunidad de Castilla y León. In Proceedings of the 5° Congresos Forestales, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Klasnja, B.; Orlovic, S.; Drekic, M.; Markovic, M. Energy production from short rotation poplar plantations. In Proceedings of the 7th International Symposium on Interdisciplinary Regional Research, Hungary, Serbia and Montenegro, Hunedoara, Romania, 25–25 September 2003; pp. 161–166. [Google Scholar]
- Fiala, M.; Bacenetti, J.; Scaravonati, A.; Bergonzi, A. Short rotation coppice in northern Italy: Comprenhensive Sustainability. In Proceedings of the 18th European Biomass Conference and Exhibition, Lyon, France, 3–7 May 2010; pp. 342–348. [Google Scholar]
- DeBell, D.S.; Clendenen, G.W.; Zasadat, J.C. Growing Populus biomass: Comparison of woodgrass versus wider-spaced short-rotation systems. Biomass Bioenergy 1993, 4, 305–313. [Google Scholar] [CrossRef]
- Ciria, M.; González, E.; Carrasco, J. The effect of fertilization and planting density on biomass productivity of poplar harvested after three-years rotation. In Proceedings of the 12th European Conference and Technology Exhibition on Biomass for Energy, Industry and Climate Protection, Amsterdam, The Netherlands, 17–21 June 2002; pp. 28–286. [Google Scholar]
- Ciria, P. Efecto del Turno de Corta y de la Densidad de Plantación Sobre la Productividad de Diversos Clones de Chopo en Condiciones de Corta Rotación. Ph.D. Thesis, Universidad Politécnica de Madrid, Madrid, Spain, 1999. [Google Scholar]
- Sevigne, E.; Gasol, C.M.; Brun, F.; Rovira, L.; Pagés, J.M.; Camps, F.; Rieradevall, J.; Gabarrell, X. Water and energy consumption of Populus spp. bioenergy systems: A case study in Southern Europe. Renew. Sustain. Energy Rev. 2011, 15, 1133–1140. [Google Scholar] [CrossRef]
- Dillen, S.Y.; Djomo, S.N.; Al Afas, N.; Vanbeveren, S.; Ceulemans, R. Biomass yield and energy balance of a short-rotation poplar coppice with multiple clones on degraded land during 16 years. Biomass Bioenergy 2013, 56, 157–165. [Google Scholar] [CrossRef]
- Friedrich, E. Anbautechnische Untersuchungen in Forstlichen Schnellwuchsplantagen und Demonstration des Leistungsvermögens Schnellwachsender Baumarten; Landwirtschaftsverlag Münster: Leipzig, Germany, 1999. [Google Scholar]
- Menéndez, J.; Loredo Pérez, J.L. Biomass production in surface mines: Renewable energy source for power plants. WSEAS Trans. Environ. Dev. 2018, 14, 205–211. [Google Scholar]
- Martín-García, J.; Merino, A.; Diez, J.J. Relating visual crown conditions to nutritional status and site quality in monoclonal poplar plantations (Populus × euramericana). Eur. J. For. Res. 2012, 131, 1185–1198. [Google Scholar] [CrossRef]
- Bungart, R.; Hüttl, R.F. Growth dynamics and biomass accumulation of 8-year-old hybrid poplar clones in a short-rotation plantation on a clayey-sandy mining substrate with respect to plant nutrition and water budget. Eur. J. For. Res. 2004, 123, 105–115. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Planting density and site effects on stem dimensions, stand productivity, biomass partitioning, carbon stocks and soil nutrient supply in hybrid poplar plantations. Forests 2018, 9, 293. [Google Scholar] [CrossRef]
- O Di Nasso, N.N.; Guidi, W.; Ragaglini, G.; Tozzini, C.; Bonari, E. Biomass production and energy balance of a 12 year old short rotation coppice poplar stand under different cutting cycles. Bioenergy 2010, 2, 89–97. [Google Scholar] [CrossRef]
- Vanbeveren, S.P.P.; Ceulemans, R. Genotypic differences in biomass production during three rotations of short-rotation coppice. Biomass Bioenergy 2018, 119, 198–205. [Google Scholar] [CrossRef]
- Gómez-Martín, J.M.; Castaño-Díaz, M.; Cámara-Obregón, A.; Álvarez-Álvarez, P.; Folgueras-Díaz, M.B.; Diez, M.A. On the chemical composition and pyrolytic behavior of hybrid poplar energy crops from northern Spain. Energy Rep. 2020, 6, 764–769. [Google Scholar] [CrossRef]
- Garasa, M.J.H.; Cañellas, I.; Viscasillas, E.; García, A.; Carrasco, J.E.; Sixto, H. Biomass yield in a short rotation clonal poplar trial. In Proceedings of the 18th European Biomass Conference and Exhibitions, Lyon, France, 3–7 May 2010. [Google Scholar]
- Oliveira, N.; De la Iglesia, J.P.; Viscasillas, E.; Bachiller, A.; Parras, A.; González, I.; Grau, J.M.; Otero, J.M.; Cañellas, I.; Sixto, H. Adecuación de genotipos para la producción de biomasa en la meseta septentrional. In Proceedings of the II Simposio del Chopo, Valladolid, Spain, 17–19 October 2018. [Google Scholar]
- Oliveira, N. Evaluación de la productividad clonal en biomasa, en un cultivo energético experimental de chopo instalado por INIA-CIFOR en Almazán (Soria). Master’s Thesis, University of Santiago de Compostela, Lugo, Spain, 2012. [Google Scholar]
- Willebrand, E.; Verwijst, T. Population dynamics of willow coppice systems and their implications for management of short-rotation forests. Forest Chron. 1993, 69, 699–704. [Google Scholar] [CrossRef][Green Version]
- Kopp, R.; Abrahamson, L.; White, E.; Volk, T.; Nowak, C.; Fillhart, R. Willow biomass production during ten successive annual harvests. Biomass Bioenergy 2001, 20, 1–7. [Google Scholar] [CrossRef]
- Tubby, I.; Armstrong, A. Establishment and Management of Short Rotation Coppice. 2002. Available online: https://www.cabdirect.org/cabdirect/abstract/20023172097 (accessed on 15 November 2020).
- Sixto, H.; Hernández, M.J.; de Miguel, J.; Cañellas, I. Red de Parcelas de Cultivos Leñosos en Alta Densidad y Turno Corto; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, Ministerio de Economía y Competitividad: Madrid, Spain, 2013; p. 188. [Google Scholar]
- Fernández, M.J.; Barro, R.; Pérez, J.; Losada, J.; Ciria, P. Influence of the agricultural management practices on the yield and quality of poplar biomass (a 9-year study). Biomass Bioenergy 2016, 93, 87–96. [Google Scholar] [CrossRef]
- Sixto, H.; Ciria, P.; Rueda, J.; Perez, J.; García Caballero, J.; Mazón, P.; Montoto, J.L.; Cañellas, I. Comparison of two cutting scenarios and clone response in the first rotation period of a poplar crop for biomass energy production. In Proceedings of the 17th European Biomass Conference and Exhibition, Hamburg, Germany, 28 June–3 July 2009. [Google Scholar]
- Štochlová, P.; Novotná, K.; Costa, M.; Rodrigues, A. Biomass production of poplar short rotation coppice over five and six rotations and its aptitude as a fuel. Biomass Bioenergy 2019, 122, 183–192. [Google Scholar] [CrossRef]
- Fernández, M.J.; Barro, R.; Pérez, J.; Ciria, P. Production and composition of biomass from short rotation coppice in marginal land: A 9-year study. Biomass Bioenergy 2020, 134, 105478. [Google Scholar] [CrossRef]
- Oliveira, N.; Otero, J.M.; Pasalodos, M.; Cañellas, I.; Pereira-Espinel, J.; Rodríguez-Soalleiro, R.; Sixto, H. Analisis de los costes de cultivos forestales de turno corto de chopo en España para produccion de bioenergia. In Proceedings of the XII Congreso de Economía Agraria, Lugo, Spain, 4–6 September 2019. [Google Scholar]
- Sixto, H.; Grau, J.M.; Eugenio, M.E.; Ibarra, D.; Hernandez, J.J.; Monedero, E.; Oliveira, N.; Otero, J.M.; Cañellas, I. Evolution of biomass production with poplar hybrids in southern Europe. In Proceedings of the 7th International Poplar Symposium. New Bioeconomies: Exploring the role of Salicaceae, Buenos Aires, Argentina, 28 October–4 November 2018. [Google Scholar]
- Dawson, J.O.; Hansen, E.A. Effect of Alnus glutinosa on hybrid populus growth and soil nitrogen concentration in a mixed plantation. In Intensive Plantation Culture: 12 Years Research; Hansen, E.A., Ed.; Gen. Tech. Rep. NC-91; U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1983; pp. 29–34. [Google Scholar]
- Radwan, M.; DeBell, D. Nutrient relations in coppiced black cottonwood and red alder. Plant Soil 1988, 106, 171–177. [Google Scholar] [CrossRef]
- Pelleri, F.; Plutino, M.; Manetti, M.C.; Sansone, D.; Bergante, S.; Castro, G.; Moya, J.F.; Chiarabaglio, P.M.; Urban, M.I. Plantaciones policíclicas mixtas: Nogales, chopos y biomasa de rotación corta. In Proceedings of the II Simposio del Chopo, Valladolid, Spain, 17–19 October 2018. [Google Scholar]
- Gana, C.; Epron, D.; Gérant, D.; Maillard, P.; Plain, C.; Priault, P.; Marron, N. How does poplar / black locust mixture influence growth and functioning of each species in a short rotation plantation? In Proceedings of the VI International Poplar Symposium, Vancouver, BC, Canada, 20–23 July 2014. [Google Scholar]
- Marron, N.; Priault, P.; Gana, C.; Gérant, D.; Epron, D. Prevalence of interspecific competition in a mixed poplar/black locust plantation under adverse climate conditions. Ann. For. Sci. 2018, 75, 23. [Google Scholar] [CrossRef]
- Oviedo, P.A.; Gutiérrez, J.A.; Martín, R.T.; Martínez, M.F. Cultivo mixto de especies forestales de turno corto (Robinia pseudoacacia y Populus × euroamericana clon AF2) con fines energéticos. Cuad. Soc. Esp. Cienc. For. 2016, 17–30. [Google Scholar] [CrossRef]
- Oliveira, N.; Del Río, M.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Cañellas, I.; Sixto, H. Mixed short rotation plantation of Populus alba and Robinia pseudoacacia for biomass yield over the course of two rotations. In Proceedings of the 7th International Poplar Symposium, Buenos Aires, Argentina, 28 October–4 November 2018. [Google Scholar]
- González, I.; Sixto, H.; Rodríguez-Soalleiro, R.; Oliveira, N. Nutrient Contribution of Litterfall in a Short Rotation Plantation of Pure or Mixed Plots of Populus alba L. and Robinia pseudoacacia L. Forests 2020, 11, 1133. [Google Scholar] [CrossRef]
- Grünewald, H.; Böhm, C.; Quinkenstein, A.; Grundmann, P.; Eberts, J.; von Wühlisch, G. Robinia pseudoacacia L.: A lesser known tree species for biomass production. Bioenergy Res. 2009, 2, 123–133. [Google Scholar] [CrossRef]
- DeBell, D.S.; Harrington, C.A. Deploying genotypes in short-rotation plantations: Mixtures and pure cultures of clones and species. For. Chron. 1993, 69, 705–713. [Google Scholar] [CrossRef]
- DeBell, D.S.; Harrington, C.A. Productivity of Populus in monoclonal and polyclonal blocks at three spacings. Can. J. For. Res. 1997, 27, 978–985. [Google Scholar] [CrossRef]
- Miot, S.; Frey, P.; Pinon, J. Varietal mixture of poplar clones: Effects on infection by Melampsora larici-populina and on plant growth. Eur. J. For. Pathol. 1999, 29, 411–423. [Google Scholar] [CrossRef]
- Elferjani, R.; DesRochers, A.; Tremblay, F. Effects of mixing clones on hybrid poplar productivity, photosynthesis and root development in northeastern Canadian plantations. For. Ecol. Manag. 2014, 327, 157–166. [Google Scholar] [CrossRef]
- Dillen, M.; Vanhellemont, M.; Verdonckt, P.; Maes, W.H.; Steppe, K.; Verheyen, K. Productivity, stand dynamics and the selection effect in a mixed willow clone short rotation coppice plantation. Biomass Bioenergy 2016, 87, 46–54. [Google Scholar] [CrossRef]
- Schweier, J.; Arranz, C.; Nock, C.A.; Jaeger, D.; Scherer-Lorenzen, M. Impact of increased genotype or species diversity in Short Rotation Coppice on biomass production and wood characteristics. Bioenergy Res. 2019, 12, 497–508. [Google Scholar] [CrossRef]
- Broeckx, L.S.; Verlinden, M.S.; Ceulemans, R. Establishment and two-year growth of a bio-energy plantation with fast-growing Populus trees in Flanders (Belgium): Effects of genotype and former land use. Biomass Bioenergy 2012, 42, 151–163. [Google Scholar] [CrossRef]
- Morhart, C.; Sheppard, J.; Seidl, F.; Spiecker, H. Influence of different tillage systems and weed treatments in the establishment year on the final biomass production of short rotation coppice poplar. Forests 2013, 4, 849. [Google Scholar] [CrossRef]
- Albertsson, J.; Hansson, D.; Bertholdsson, N.O.; Åhman, I. Site-related set-back by weeds on the establishment of 12 biomass willow clones. Weed Res. 2014, 54, 398–407. [Google Scholar] [CrossRef]
- Heilman, P.E.; Xie, F.-G. Effects of nitrogen fertilization on leaf area, light interception, and productivity of short-rotation Populus trichocarpa × Populus deltoides hybrids. Can. J. For. Res. 1994, 24, 166–173. [Google Scholar] [CrossRef]
- Ceulemans, R.; Deraedt, W. Production physiology and growth potential of poplars under short-rotation forestry culture. For. Ecol. Manag. 1999, 121, 9–23. [Google Scholar] [CrossRef]
- Liberloo, M.; Calfapietra, C.; Lukac, M.; Godbold, D.; Luo, Z.-b.; Polle, A.; Hoosbeek, M.R.; Kull, O.; Marek, M.; Raines, C. Woody biomass production during the second rotation of a bio-energy Populus plantation increases in a future high CO2 world. Glob. Chang. Biol. 2006, 12, 1094–1106. [Google Scholar] [CrossRef]
- Van Veen, J.; Breteler, H.; Olie, J.; Frissel, M. Nitrogen and energy balance of a short-rotation poplar forest system. Neth. J. Agri. Sci. 1981, 29, 163–172. [Google Scholar] [CrossRef]
- Coleman, M.; Tolsted, D.; Nichols, T.; Johnson, W.D.; Wene, E.G.; Houghtaling, T. Post-establishment fertilization of Minnesota hybrid poplar plantations. Biomass Bioenergy 2006, 30, 740–749. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Van Oosten, C.; Netzer, D.A.; Coleman, M.D.; Portwood, C.J. Ecology and silviculture of poplar plantations. In Poplar Culture in North America; Dickmann, D.I., Isebrands, J.G., Eckenwalder, J.E., Richardson, J., Eds.; NRC Research Press: Ottawa, ON, Canada, 2002; pp. 153–206. [Google Scholar]
- Yan, X.-L.; Dai, T.-F.; Zhao, D.; Jia, L.-M. Combined surface drip irrigation and fertigation significantly increase biomass and carbon storage in a Populus × euramericana cv. Guariento plantation. J. For. Res. 2016, 21, 280–290. [Google Scholar] [CrossRef]
- Euring, D.; Löfke, C.; Teichmann, T.; Polle, A. Nitrogen fertilization has differential effects on N allocation and lignin in two Populus species with contrasting ecology. Trees 2012, 26, 1933–1942. [Google Scholar] [CrossRef]
- Coleman, M.D.; Dickson, R.E.; Isebrands, J. Growth and physiology of aspen supplied with different fertilizer addition rates. Physiol. Plant. 1998, 103, 513–526. [Google Scholar] [CrossRef]
- Paris, P.; Mareschi, L.; Sabatti, M.; Tosi, L.; Scarascia-Mugnozza, G. Nitrogen removal and its determinants in hybrid Populus clones for bioenergy plantations after two biennial rotations in two temperate sites in northern Italy. iForest 2015, 8, 657–665. [Google Scholar] [CrossRef]
- Rennenberg, H.; Wildhagen, H.; Ehlting, B. Nitrogen nutrition of poplar trees. Plant Biol. 2010, 12, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xi, B.; Bloomberg, M.; Moltchanova, E.; Li, G.; Jia, L. Response of diameter growth, biomass allocation and N uptake to N fertigation in a triploid Populus tomentosa plantation in the North China Plain: Ontogenetic shift does not exclude plasticity. Eur. J. Forest Res. 2015, 134, 889–898. [Google Scholar] [CrossRef]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293. [Google Scholar] [CrossRef] [PubMed]
- Erisman, J.W.; van Grinsven, H.; Leip, A.; Mosier, A.; Bleeker, A. Nitrogen and biofuels; an overview of the current state of knowledge. Nut. Cycl. Agroecosyst. 2010, 86, 211–223. [Google Scholar] [CrossRef]
- Heller, M.C.; Keoleian, G.A.; Volk, T.A. Life cycle assessment of a willow bioenergy cropping system. Biomass Bioenergy 2003, 25, 147–165. [Google Scholar] [CrossRef]
- Sugiura, A.; Tyrrel, S.; Seymour, I.; Burgess, P. Water Renew systems: Wastewater polishing using renewable energy crops. Water Sci. Technol. 2008, 57, 1421–1428. [Google Scholar] [CrossRef]
- Werner, A.; McCracken, A. The use of short rotation coppice poplar and willow for the bioremediation of sewage effluent. Asp. Appl. Biol. 2008, 317–324. [Google Scholar]
- Dimitriou, I.; Rosenqvist, H. Sewage sludge and wastewater fertilisation of Short Rotation Coppice (SRC) for increased bioenergy production—Biological and economic potential. Biomass Bioenergy 2011, 35, 835–842. [Google Scholar] [CrossRef]
- Martínez-Hernández, V.; Leal, M.; Meffe, R.; De Miguel, Á.; Alonso-Alonso, C.; De Bustamante, I.; Lillo, J.; Martín, I.; Salas, J. Removal of emerging organic contaminants in a poplar vegetation filter. J. Hazard. Mater. 2018, 342, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Pradana, R.; Sixto, H.; González, B.; Demaria, I.; González, I.; Martínez-Hernández, V.; de Bustamante, I. Poplar Vegetation Filters for the beer industry: Wastewater treatment combined with biomass production. In Proceedings of the XI Congreso Ibérico de Gestión y Planificación del Agua, Madrid, Spain, 3–5 September 2020. [Google Scholar]
- Paniagua, S.; Escudero, L.; Escapa, C.; Coimbra, R.N.; Otero, M.; Calvo, L.F. Effect of waste organic amendments on Populus sp biomass production and thermal characteristics. Renew. Energy 2016, 94, 166–174. [Google Scholar] [CrossRef]
- Sixto, H.; Garasa, M.H.; Ciria, P.C.; Garcia, J.C.; de Vinas, I.C.R. Manual de Cultivo de Populus spp. Para la Producción de Biomasa Con Fines Energéticos. 2010. Available online: https://agris.fao.org/agris-search/search.do?recordID=XF2015034695 (accessed on 15 November 2020).
- Kollert, W. La populicultura en Europa y España. In Proceedings of the II Simposio del chopo, Valladolid, Spain, 17–19 October 2018. [Google Scholar]
- Newcombe, G.; Ostry, M.; Hubbes, M.; Périnet, P.; Mottet, M.-J. Poplar diseases. In Poplar Culture in North America; Dickmann, D.I., Isebrands, J.G., Eckenwalder, J.E., Richardson, J., Eds.; NRC Research Press: Ottawa, ON, Canada, 2001; pp. 249–276. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nebeker, T.E.; Hart, E.R.; Mattson, W.J. Biology and management of insect pests in North American intensively managed hardwood forest systems. Annu. Rev. Entomol. 2005, 50, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Duplessis, S.; Major, I.; Martin, F.; Séguin, A. Poplar and pathogen interactions: Insights from Populus genome-wide analyses of resistance and defense gene families and gene expression profiling. Crit. Rev. Plant Sci. 2009, 28, 309–334. [Google Scholar] [CrossRef]
- Broderick, N.A.; Vasquez, E.; Handelsman, J.; Raffa, K.F. Effect of clonal variation among hybrid poplars on susceptibility of gypsy moth (Lepidoptera: Lymantriidae) to Bacillus thuringiensis subsp. kurstaki. J. Econ. Entomol. 2010, 103, 718–725. [Google Scholar] [CrossRef]
- Steenackers, J.; Steenackers, M.; Steenackers, V.; Stevens, M. Poplar diseases, consequences on growth and wood quality. Biomass Bioenergy 1996, 10, 267–274. [Google Scholar] [CrossRef]
- Soria, S. Patología de las Choperas; Caja Rural del Duero. Colegio de Ingenieros de Montes: Valladolid, Spain, 1992; pp. 61–75. [Google Scholar]
- Nordman, E.E.; Robison, D.J.; Abrahamson, L.P.; Volk, T.A. Relative resistance of willow and poplar biomass production clones across a continuum of herbivorous insect specialization: Univariate and multivariate approaches. For. Ecol. Manag. 2005, 217, 307–318. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Richardson, J. Poplars and Willows: Trees for Society and the Environment; The Food and Agricultural Organization of the United Nations (FAO) and CAB International: Rome, Italy, 2014. [Google Scholar]
- Bloemen, J.; Fichot, R.; Horemans, J.A.; Broeckx, L.S.; Verlinden, M.S.; Zenone, T.; Ceulemans, R. Water use of a multi-genotype poplar short-rotation coppice from tree to stand scale. Glob. Chang. Biol. Bioenegy 2016, 9, 370–384. [Google Scholar] [CrossRef]
- Padró, A.; Orensanz, J. El Chopo y su Cultivo; Ministerio de Agricultura, Pesca y Alimentación (MAPA): Madrid, Spain, 1987. [Google Scholar]
- Sixto, H.; Ruiz, V.; Grau, J.; Montoto, J. Primeros resultados de un ensayo de riego en vivero de planta de chopo. 1. In Proceedings of the 1er Simposio del Chopo, Zaragoza, Spain, 9–11 May 2001; pp. 159–166. [Google Scholar]
- Sixto, H.; Grau, J.; González-Antoñanzas, F. Populicultura: Populus spp. e híbridos. Capítulo IV. In Compendio de Selvicultura Aplicada en España; Serrada, R., Montero, G., Reque, J.A., Eds.; Institulo Nacional de Investigación y Tecnología Agraria y Alimentaria, Ministerio de Educación y Ciencia: Madrid, Spain, 2008. [Google Scholar]
- IPCC. Climate Change 2007: Impacts, adaptation and vulnerability. In Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; p. 976. [Google Scholar]
- González-González, B.D.; Oliveira, N.; González, I.; Cañellas, I.; Sixto, H. Poplar biomass production in short rotation under irrigation: A case study in the Mediterranean. Biomass Bioenergy 2017, 107, 198–206. [Google Scholar] [CrossRef]
- Hennig, A.; Kleinschmit, J.R.; Schoneberg, S.; Löffler, S.; Janßen, A.; Polle, A. Water consumption and biomass production of protoplast fusion lines of poplar hybrids under drought stress. Front. Plant Sci. 2015, 6, 330. [Google Scholar] [CrossRef]
- Liang, Z.-S.; Yang, J.-W.; Shao, H.-B.; Han, R.-L. Investigation on water consumption characteristics and water use efficiency of poplar under soil water deficits on the Loess Plateau. Colloid Surface B 2006, 53, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Viger, M.; Smith, H.; Cohen, D.; Dewoody, J.; Trewin, H.; Steenackers, M.; Bastien, C.; Taylor, G. Adaptive mechanisms and genomic plasticity for drought tolerance identifed in European black poplar (Populus nigra L.). Tree Physiol. 2016, 36, 909–928. [Google Scholar] [CrossRef] [PubMed]
- Niemczyk, M.; Hu, Y.; Thomas, B.R. Selection of Poplar Genotypes for Adapting to Climate Change. Forests 2019, 10, 1041. [Google Scholar] [CrossRef]
- Garasa, M.J.H. Respuesta Anatómico Fisiológicas Frente a Estrés Hídrico en Plantaciones de Especies de Crecimiento Rápido Para la Producción de Biomasa. Ph.D. Thesis, Universidad Politécnica de Madrid, Madrid, Spain, 2015. [Google Scholar]
- Navarro, A.; Facciotto, G.; Campi, P.; Mastrorilli, M. Physiological adaptations of five poplar genotypes grown under SRC in the semi-arid Mediterranean environment. Trees 2014, 28, 983–994. [Google Scholar] [CrossRef]
- Navarro, A.; Portillo-Estrada, M.; Arriga, N.; Vanbeveren, S.P.; Ceulemans, R. Genotypic variation in transpiration of coppiced poplar during the third rotation of a short-rotation bio-energy culture. Glob. Chang. Biol. Bioenergy 2018, 10, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Portillo-Estrada, M.; Ceulemans, R. Identifying the best plant water status indicator for bio-energy poplar genotypes. Glob. Chang. Biol. Bioenergy 2020, 12, 426–444. [Google Scholar] [CrossRef]
- Fuertes, A.; Oliveira, N.; Cañellas, I.; Sixto, H.; Rodríguez-Soalleiro, R. An economic overview of Populus spp. in Short Rotation Coppice systems under Mediterranean conditions: An assessment tool for decision-making. Sustain. Energy Rev. 2020. under review. [Google Scholar]
- de Bustamante, I. Filtros verdes. Un sistema para la depuración y reutilización de aguas residuales. Tecnoambiente 1998, 79, 73–75. [Google Scholar]
- de Miguel, Á.; Martínez-Hernández, V.; Leal, M.; González-Naranjo, V.; de Bustamante, I. Del residuo al recurso: Reutilización de aguas para riego de biocombustibles-From waste to resource: Reuse of water for biofuel irrigation. Water 2012, 79, 68–70. [Google Scholar]
- Makeschin, F. Short rotation forestry in Central and Northern Europe-introduction and conclusions. For. Ecol. Manag. 1999, 121, 1–7. [Google Scholar]
- Ciria, M.; Mazón, M.; Carrasco, J. Poplar productivity evolution on short rotation during three consecutive cycles in extreme continental climate. In Proceedings of the 2nd World Conference on Biomass for Energy, Industry and Climate Protection, Rome, Italy, 10–14 May 2004. [Google Scholar]
- Laureysens, I.; Deraedt, W.; Indeherberge, T.; Ceulemans, R. Population dynamics in a 6-year old coppice culture of poplar. I. Clonal differences in stool mortality, shoot dynamics and shoot diameter distribution in relation to biomass production. Biomass Bioenerg 2003, 24, 81–95. [Google Scholar] [CrossRef]
- Benetka, V.; Novotná, K.; Štochlová, P. Biomass production of Populus nigra L. clones grown in short rotation coppice systems in three different environments over four rotations. iForest 2014, 7, 233–239. [Google Scholar] [CrossRef]
- Trnka, M.; Trnka, M.; Fialová, J.; Koutecky, V.; Fajman, M.; Zalud, Z.; Hejduk, S. Biomass production and survival rates of selected poplar clones grown under a short-rotation system on arable land. Plant Soil Environ. 2008, 54, 78–88. [Google Scholar] [CrossRef]
- Berthelot, A. Mélange de clones en taillis à courtes rotations de peuplier: Influence sur la productivité et l’homogénéité des produits récoltés. Can. J. For. Res. 2001, 31, 1116–1126. [Google Scholar] [CrossRef]
- Hartmann, L.; Lamersdorf, N. Site conditions, initial growth and nutrient and litter cycling of newly installed short rotation coppice and agroforestry systems. In Bioenergy from Dendromass for the Sustainable Development of Rural Areas; Manning, D.B., Bemmann, A., Bredemeier, N., Lamersdorf, N., Ammer, C., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2015. [Google Scholar]
- Kern, J.; Germer, S.; Ammon, C.; Balasus, A.; Bischoff, W.-A.; Schwarz, A.; Forstreuter, M.; Kaupenjohann, M. Environmental effects over the first 2½ rotation periods of a fertilised poplar short rotation coppice. Bioenergy Res. 2018, 11, 152–165. [Google Scholar] [CrossRef]
- Cabrera, A.; Tozzini, C.; Espinoza, S.; Santelices, R.; Bonari, E. Cálculo del balance energético de una plantación de Populus deltoides clon Lux con fines energéticos en un sitio con ambiente mediterráneo. Bosque 2014, 35, 133–139. [Google Scholar] [CrossRef]
- Paris, P.; Mareschi, L.; Sabatti, M.; Pisanelli, A.; Ecosse, A.; Nardin, F.; Scarascia-Mugnozza, G. Comparing hybrid Populus clones for SRF across northern Italy after two biennial rotations: Survival, growth and yield. Biomass Bioenergy 2011, 35, 1524–1532. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Krzyżaniak, M.; Szczukowski, S.; Tworkowski, J.; Bieniek, A. Short rotation woody crops grown on marginal soil for biomass energy. Pol. J. Environ. Stud. 2014, 23, 1727–1739. [Google Scholar]
- Proe, M.F.; Griffiths, J.H.; Craig, J. Effects of spacing, species and coppicing on leaf area, light interception and photosynthesis in short rotation forestry. Biomass Bioenergy 2002, 23, 315–326. [Google Scholar] [CrossRef]
- Baraza, C. Cultivos demostrativos de chopos (Populus spp.) a altas densidades con fines energéticos realizados en Aragón. Primeros datos y resultados. In Proceedings of the 6° Congreso Forestal Español, Vitoria, Spain, 10–14 June 2013. [Google Scholar]
- Ciria, M.; González, E.; Mazon, P.; Carrasco, J. Influence of the rotation age and plant density on the composition and quality of poplar biomass. Biomass for energy and the environment. In Proceedings of the 9th European Bioenergy Conference and 1st European Energy from Biomass Technology Exhibition, Copenhagen, Denmark, 24–27 June 1996; pp. 968–973. [Google Scholar]
- Eimil-Fraga, C.; Proupín-Castiñeiras, X.; Rodríguez-Añón, J.A.; Rodríguez-Soalleiro, R. Effects of Shoot Size and Genotype on Energy Properties of Poplar Biomass in Short Rotation Crops. Energies 2019, 12, 2051. [Google Scholar] [CrossRef]
- Godino, M. Análisis de los Cultivos de Populus × euroamericana, Clon I-214, a Turnos Muy Cortos con Fines Energéticos. Ph.D. Thesis, Universidad Politécnica de Madrid, Madrid, Spain, 2005. [Google Scholar]
- Ibarra, D.; Eugenio, M.E.; Cañellas, I.; Sixto, H.; Martín-Sampedro, R. Potential of different poplar clones for sugar production. Wood Sci. Technol. 2017, 51, 669–684. [Google Scholar] [CrossRef]
- Martín-Sampedro, R.; Eugenio, M.E.; Cañellas, I.; Sixto, H.; Ibarra, D. Characterization of different poplar clones for sugar production. In Proceedings of the 1st International Workshop on Biorefinery of Lignocellulosic Materials, Cordoba, Spain, 9–12 June 2015. [Google Scholar]
- Djomo, S.N.; Ac, A.; Zenone, T.; De Groote, T.; Bergante, S.; Facciotto, G.; Sixto, H.; Ciria Ciria, P.; Weger, J.; Ceulemans, R. Energy performances of intensive and extensive short rotation cropping systems for woody biomass production in the EU. Renew. Sustain. Energy Rev. 2015, 41, 845–854. [Google Scholar] [CrossRef]
- Oliveira, N.; Pérez-Cruzado, C.; Sixto, H.; Cañellas, I.; Rodrígez-Soalleiro, R. Modelización alométrica para la estimación de biomasa en un cultivo energético experimental de chopo. In Proceedings of the II Encuentro Juventud Investigadora. Universidad de Santiago de Compostela, Santiago de Compostela, Spain, 29–31 January 2014. [Google Scholar]
- San Miguel, G.; Corona, B.; Ruiz, D.; Landholm, D.; Laina, R.; Tolosana, E.; Sixto, H.; Cañellas, I. Environmental, energy and economic analysis of a biomass supply chain based on a poplar short rotation coppice in Spain. J. Clean. Prod. 2015, 94, 93–101. [Google Scholar] [CrossRef]
- Valbuena-Castro, J.; Oliveira, N.; Rodríguez-Soalleiro, R.; Sixto, H.; Cañellas, I. Biomass productivity assessment at a clonal forest of Populus spp. at Valtierra (Navarra, Spain). In Proceedings of the Xth Young Researchers Meeting on Conservation and Sustainable Use of Forest Systems, Valsaín, Spain, 25–26 January 2016. [Google Scholar]
- Valbuena-Castro, J.; Oliveira, N.; Rodríguez-Soalleiro, R.; Cañellas, I.; Sixto, H. Assessment of biomass productivity and effect of planting density in a short rotation coppice poplar plantation in the north of Spain. In Proceedings of the 25th Session of the International Poplar Commission, Berlin, Germany, 12–16 September 2016. [Google Scholar]
- Picard, N.; Saint-André, L.; Henry, M. Manual for Building Tree Volume and Biomass Allometric Equations: From Field Measurement to Prediction; Food and Agricultural Organization of the United Nations and Centre de Coopération Internationale en Recherche Agronomique pour le Développement, Montpellier: Rome, Italy, 2012; p. 215. [Google Scholar]
- Peichl, M.; Arain, M.A. Allometry and partitioning of above- and belowground tree biomass in an age-sequence of white pine forests. For. Ecol. Manag. 2007, 253, 68–80. [Google Scholar] [CrossRef]
- Shaiek, O.; Loustau, D.; Trichet, P.; Meredieu, C.; Bachtobji, B.; Garchi, S.; Aouni, M.H.E. Generalized biomass equations for the main aboveground biomass components of maritime pine across contrasting environments. Ann. For. Sci. 2011, 68, 443–452. [Google Scholar] [CrossRef]
- Verwijst, T.; Lundkvist, A.; Edelfeldt, S.; Albertsson, J. Development of sustainable willow short rotation forestry in Northern Europe. In Biomass Now-Sustainable Growth and Use; Matovic, D.M.D., Ed.; BoD–Books on Demand: Croatia, 2013; pp. 479–502. Available online: https://books.google.com.hk/books?hl=zh-CN&lr=&id=WdmgDwAAQBAJ&oi=fnd&pg=PA479&dq=Development+of+sustainable+willow+short+rotation+forestry+in+Northern+Europe&ots=BJwT6IWByQ&sig=TZ6h3-l6e8g3YMKL1hdp77ywZ2w&redir_esc=y&hl=zh-CN&sourceid=cndr#v=onepage&q=Development%20of%20sustainable%20willow%20short%20rotation%20forestry%20in%20Northern%20Europe&f=false (accessed on 15 November 2020). [CrossRef]
- Mosseler, A.; Major, J.; Labrecque, M.; Larocque, G. Allometric relationships in coppice biomass production for two North American willows (Salix spp.) across three different sites. For. Ecol. Manag. 2014, 320, 190–196. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Broeckx, L.S.; Ceulemans, R. First vs. second rotation of a poplar short rotation coppice: Above-ground biomass productivity and shoot dynamics. Biomass Bioenergy 2015, 73, 174–185. [Google Scholar] [CrossRef]
- Cannell, M.; Willett, S. Shoot growth phenology, dry matter distribution and root: Shoot ratios of provenances of Populus trichocarpa, Picea sitchensis and Pinus contorta growing in Scotland. Silvae Genet. 1976, 25, 49–59. [Google Scholar]
- Ruark, G.A.; Martin, G.L.; Bockheim, J.G. Comparison of constant and variable allometric ratios for estimating Populus tremuloides biomass. For. Sci. 1987, 33, 294–300. [Google Scholar] [CrossRef]
- Oliveira, N.; Rodríguez-Soalleiro, R.; Hernández, M.J.; Cañellas, I.; Sixto, H.; Pérez-Cruzado, C. Improving biomass estimation in a Populus short rotation coppice plantation. For. Ecol. Manag. 2017, 391, 194–206. [Google Scholar] [CrossRef]
- Di Matteo, G.; Sperandio, G.; Verani, S. Field performance of poplar for bioenergy in southern Europe after two coppicing rotations: Effects of clone and planting density. iForest 2012, 5, 224–229. [Google Scholar] [CrossRef]
- Rock, J. Suitability of published biomass equations for aspen in Central Europe-Results from a case study. Biomass Bioenergy 2007, 31, 299–307. [Google Scholar] [CrossRef]
- Coyle, D.R.; Coleman, M.D. Forest production responses to irrigation and fertilization are not explained by shifts in allocation. For. Ecol. Manag. 2005, 208, 137–152. [Google Scholar] [CrossRef]
- Oliveira, N.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Cañellas, I.; Sixto, H. On the genetic affinity of individual tree biomass allometry in poplar short rotation coppice. Bioenergy Res. 2017, 10, 525–535. [Google Scholar] [CrossRef]
- Pérez-Cruzado, C.; Fehrmann, L.; Magdon, P.; Cañellas, I.; Sixto, H.; Kleinn, C. On the site-level suitability of biomass models. Environ. Modell. Softw. 2015, 73, 14–26. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and Stem Volume Equations for Tree Species in Europe; Tammer-Paino Oy: Tampere, Finland, 2005. [Google Scholar]
- Dowell, R.C.; Gibbins, D.; Rhoads, J.L.; Pallardy, S.G. Biomass production physiology and soil carbon dynamics in short-rotation-grown Populus deltoides and P. deltoides × P. nigra hybrids. For. Ecol. Manag. 2009, 257, 134–142. [Google Scholar] [CrossRef]
- Johansson, T.; Karacic, A. Increment and biomass in hybrid poplar and some practical implications. Biomass Bioenergy 2011, 35, 1925–1934. [Google Scholar] [CrossRef]
- Zabek, L.M.; Prescott, C.E. Biomass equations and carbon content of aboveground leafless biomass of hybrid poplar in Coastal British Columbia. For. Ecol. Manag. 2006, 223, 291–302. [Google Scholar] [CrossRef]
- Ben Brahim, M.; Gavaland, A.; Cabanettes, A. Generalized allometric regression to estimate biomass of Populus in short-rotation coppice. Scand. J. For. Res. 2000, 15, 171–176. [Google Scholar] [CrossRef]
- Casella, E.; Sinoquet, H. A method for describing the canopy architecture of coppice poplar with allometric relationships. Tree Physiol. 2003, 23, 1153–1170. [Google Scholar] [CrossRef]
- Berhongaray, G.; Janssens, I.A.; King, J.S.; Ceulemans, R. Fine root biomass and turnover of two fast-growing poplar genotypes in a short-rotation coppice culture. Plant Soil 2013, 373, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Berhongaray, G.; Verlinden, M.S.; Broeckx, L.S.; Ceulemans, R. Changes in belowground biomass after coppice in two Populus genotypes. For. Ecol. Manag. 2015, 337, 1–10. [Google Scholar] [CrossRef]
- Oliveira, N.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Cañellas, I.; Sixto, H.; Ceulemans, R. Above- and below-ground carbon accumulation and biomass allocation in poplar short rotation plantations under Mediterranean conditions. For. Ecol. Manag. 2018, 428, 57–65. [Google Scholar] [CrossRef]
- Amichev, B.Y.; Johnston, M.; Van Rees, K.C.J. Hybrid poplar growth in bioenergy production systems: Biomass prediction with a simple process-based model (3PG). Biomass Bioenergy 2010, 34, 687–702. [Google Scholar] [CrossRef]
- Headlee, W.L.; Zalesny, R.S., Jr.; Donner, D.M.; Hall, R.B. Using a process-based model (3-PG) to predict and map hybrid poplar biomass productivity in Minnesota and Wisconsin, USA. BioEnergy Res. 2013, 6, 196–210. [Google Scholar] [CrossRef]
- Hart, Q.J.; Prilepova, O.; Merz, J.R.; Bandaru, V.; Jenkins, B.M. Modeling poplar growth as a short rotation woody crop for biofuels. UC Davis: Institute of Transportation Studies: EScholarship. University of California. 2014. Available online: https://escholarship.org/uc/item/1cc1p27b (accessed on 15 November 2020).
- Hart, Q.J.; Tittmann, P.W.; Bandaru, V.; Jenkins, B.M. Modeling poplar growth as a short rotation woody crop for biofuels in the Pacific Northwest. Biomass Bioenergy 2015, 79, 12–27. [Google Scholar] [CrossRef]
- Beringer, T.; Lucht, W.; Schaphoff, S. Bioenergy production potential of global biomass plantations under environmental and agricultural constraints. Glob. Chang. Biol. Bioenergy 2011, 3, 299–312. [Google Scholar] [CrossRef]
- Rodrigues, A.; Gonçalves, A.B.; Casquilho, M.; Gomes, A.A. A GIS-based evaluation of the potential of woody short rotation coppice (SRC) in Portugal aiming at co-firing and decentralized co-generation. Biomass Bioenergy 2020, 137, 105554. [Google Scholar] [CrossRef]
- Djomo, S.N.; De Groote, T.; Gobin, A.; Ceulemans, R.; Janssens, I.A. Combining a land surface model with life cycle assessment for identifying the optimal management of short rotation coppice in Belgium. Biomass Bioenergy 2019, 121, 78–88. [Google Scholar] [CrossRef]
- Castaño-Díaz, M.; Álvarez-Álvarez, P.; Tobin, B.; Nieuwenhuis, M.; Afif-Khouri, E.; Cámara-Obregón, A. Evaluation of the use of low-density LiDAR data to estimate structural attributes and biomass yield in a short-rotation willow coppice: An example in a field trial. Ann. For. Sci. 2017, 74, 69. [Google Scholar] [CrossRef]
- Sánchez-González, M.; Durbán, M.; Lee, D.-J.; Cañellas, I.; Sixto, H. Smooth additive mixed models for predicting aboveground biomass. J. Agric. Biol. Environ. Stat. 2017, 22, 23–41. [Google Scholar] [CrossRef]
- Barrio-Anta, M.; Sixto-Blanco, H.; Viñas, I.C.-R.D.; Castedo-Dorado, F. Dynamic growth model for I-214 poplar plantations in the northern and central plateaux in Spain. Forest Ecol. Manag. 2008, 255, 1167–1178. [Google Scholar] [CrossRef]
- de Ron, D.S.; Pérez-Cruzado, C.; González-Estevez, V.; Rodríguez-Soalleiro, R.; Cañellas, I.; Sixto, H. Visor cartográfico sobre producción de biomasa de chopo (Populus spp.) con fines energéticos en España. In Proceedings of the 6° Congreso Forestal Español, Vitoria-Gasteiz, Spain, 10–14 June 2013. [Google Scholar]
- Gasol, C.M.; Gabarrell, X.; Rigola, M.; González-García, S.; Rieradevall, J. Environmental assessment:(LCA) and spatial modelling (GIS) of energy crop implementation on local scale. Biomass Bioenergy 2011, 35, 2975–2985. [Google Scholar] [CrossRef]
- Andújar, D.; Fernández-Quintanilla, C.; Dorado, J. Matching the best viewing angle in depth cameras for biomass estimation based on poplar seedling geometry. Sensors 2015, 15, 12999–13011. [Google Scholar] [CrossRef] [PubMed]
- Andújar, D.; Rosell-Polo, J.R.; Sanz, R.; Rueda-Ayala, V.; Fernández-Quintanilla, C.; Ribeiro, A.; Dorado, J. A LiDAR-based system to assess poplar biomass. Gesunde Pflanz. 2016, 68, 155–162. [Google Scholar] [CrossRef]
- Oliveira, N.; Sixto, H.; Cañellas, I.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C. Productivity model and reference diagram for short rotation biomass crops of poplar grown in Mediterranean environments. Biomass Bioenergy 2015, 72, 309–320. [Google Scholar] [CrossRef]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef]
- Sannigrahi, P.; Ragauskas, A.J.; Tuskan, G.A. Poplar as a feedstock for biofuels: A review of compositional characteristics. Biofuel. Bioprod. Biorefin. 2010, 4, 209–226. [Google Scholar] [CrossRef]
- Bioplat. Manual Sobre las Biorrefinerías en España; Bioplat, Suschem y Agencia estatal de investigación del Ministerio de Economía, Industría y Competitividad: Madrid, Spain, 2017. [Google Scholar]
- Tharakan, P.J.; Volk, T.A.; Abrahamson, L.P.; White, E.H. Energy feedstock characteristics of willow and hybrid poplar clones at harvest age. Biomass Bioenerg 2003, 25, 571–580. [Google Scholar] [CrossRef]
- Obernberger, I.; Brunner, T.; Bärnthaler, G. Chemical properties of solid biofuels-significance and impact. Biomass Bioenergy 2006, 30, 973–982. [Google Scholar] [CrossRef]
- Elder, T.; Groom, L.H. Pilot-scale gasification of woody biomass. Biomass Bioenergy 2011, 35, 3522–3528. [Google Scholar] [CrossRef]
- Kloss, S.; Zehetner, F.; Dellantonio, A.; Hamid, R.; Ottner, F.; Liedtke, V.; Schwanninger, M.; Gerzabek, M.H.; Soja, G. Characterization of slow pyrolysis biochars: Effects of feedstocks and pyrolysis temperature on biochar properties. J. Environ. Qual. 2012, 41, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Bridgwater, T. Challenges and opportunities in fast pyrolysis of biomass: Part I. Johnson Matthey Technol. Rev. 2018, 62, 118–130. [Google Scholar] [CrossRef]
- Raud, M.; Kikas, T.; Sippula, O.; Shurpali, N.J. Potentials and challenges in lignocellulosic biofuel production technology. Renew. Sustain. Energy Rev. 2019, 111, 44–56. [Google Scholar] [CrossRef]
- Moreno, A.D.; Alvira, P.; Ibarra, D.; Tomás-Pejó, E. Production of ethanol from lignocellulosic biomass. In Production of Platform Chemicals from Sustainable Resources; Springer: Berlin/Heidelberg, Germany, 2017; pp. 375–410. [Google Scholar]
- González-García, S.; Dias, A.C.; Clermidy, S.; Benoist, A.; Bellon Maurel, V.; Gasol, C.M.; Gabarrell, X.; Arroja, L. Comparative environmental and energy profiles of potential bioenergy production chains in Southern Europe. J. Clean. Prod. 2014, 76, 42–54. [Google Scholar] [CrossRef]
- Martínez, M.; Duret, X.; Minh, D.P.; Nzihou, A.; Lavoie, J. Conversion of lignocellulosic biomass in biobutanol by a NOVEL thermal process. Int. J. Energy Prod. Manag. 2019, 4, 298–310. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, C.; Lu, M.; Tu, M. Effect of overliming and activated carbon detoxification on inhibitors removal and butanol fermentation of poplar prehydrolysates. Biotechnol. Biofuels 2018, 11, 178. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J.; Warmiński, K. Life cycle assessment of poplar production: Environmental impact of different soil enrichment methods. J. Clean. Prod. 2019, 206, 785–796. [Google Scholar] [CrossRef]
- Díaz-Ramírez, M.; Nogués, F.S.; Royo, J.; Rezeau, A. Combustion Behavior of Novel Energy Crops in Domestic Boilers: Poplar and Brassica Experiences. In Alternative Energies; Springer: Berlin/Heidelberg, Germany, 2013; pp. 27–45. [Google Scholar]
- Díaz-Ramírez, M.C. Grate-Fired Energy Crop Conversion: Experiences with Brassica Carinata and Populus sp.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Díaz-Ramírez, M.; Boman, C.; Sebastián, F.; Royo, J.; Xiong, S.; Boström, D. Environmental performance of three novel opportunity biofuels: Poplar, brassica and cassava during fixed bed combustion. In Herbaceous Platns: Cultivation Methods, Grazing and Environmental Impacts; Wallner, F., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2013. [Google Scholar]
- Santos, J.I.; Fillat, Ú.; Martín-Sampedro, R.; Ballesteros, I.; Manzanares, P.; Ballesteros, M.; Eugenio, M.E.; Ibarra, D. Lignin-enriched fermentation residues from bioethanol production of fast-growing poplar and forage sorghum. Bioresources 2015, 10, 5215–5232. [Google Scholar] [CrossRef][Green Version]
- Paniagua, S.; García-Pérez, A.I.; Calvo, L.F. Biofuel consisting of wheat straw–poplar wood blends: Thermogravimetric studies and combustion characteristic indexes estimation. Biomass Convers. Biorefin. 2019, 9, 433–443. [Google Scholar] [CrossRef]
- Serrano, D.; Coronado, J.M.; Melero, J.A. Conversion of cellulose and hemicellulose into platform molecules: Chemical routes. In Biorefinery: From Biomass to Chemicals and Fuels; Aresta, M., Dibenedetto, A., Dumeignil, F., Eds.; The Gruyter: Berlin/Boston, Germany, 2012; pp. 123–140. [Google Scholar]
- Berrueco Martínez, B.; Langa Morales, E.; Maestro Tejada, C.; Mainar Fernández, A.M.; Urieta Navarro, J.S. Potencial de las yemas de chopo como fuente de compuestos bioactivos: Actividad antioxidante de sus extractos supercríticos. In Proceedings of the 5° Congreso Forestal Español, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Martín-Sampedro, R.; Santos, J.I.; Fillat, Ú.; Wicklein, B.; Eugenio, M.E.; Ibarra, D. Characterization of lignins from Populus alba L. generated as by-products in different transformation processes: Kraft pulping, organosolv and acid hydrolysis. Int. J. Biol. Macromol. 2019, 126, 18–29. [Google Scholar] [CrossRef]
- McKendry, P. Energy production from biomass (part 1): Overview of biomass. Bioresour. Technol. 2002, 83, 37–46. [Google Scholar] [CrossRef]
- Kenney, W.A.; Sennerby-Forsse, L.; Layton, P. A review of biomass quality research relevant to the use of poplar and willow for energy conversion. Biomass 1990, 21, 163–188. [Google Scholar] [CrossRef]
- Fernández, M.J.; Ciria, P.; Barro, R.; Losada, J.; Pérez, J.; Sixto, H.; Carrasco, J.E. Quality of the biomass produced in Short Rotation Coppices of poplar, willow, black locust and sycamore in two different spanish locations. Biomass Resour. 2013, 134–139. [Google Scholar] [CrossRef]
- Monedero, E.; Hernández, J.J.; Cañellas, I.; Otero, J.M.; Sixto, H. Thermochemical and physical evaluation of poplar genotypes as short rotation forestry crops for energy use. Energy Convers. Manag. 2016, 129, 131–139. [Google Scholar] [CrossRef]
- Pliura, A.; Zhang, S.; MacKay, J.; Bousquet, J. Genotypic variation in wood density and growth traits of poplar hybrids at four clonal trials. For. Ecol. Manag. 2007, 238, 92–106. [Google Scholar] [CrossRef]
- Marcos, F.; García, R.; García, F.; Godino, M.; Relova, I.; Villegas, S. Caracterización energética de la biomasa de chopo (Populus x euramericana I-214). In Proceedings of the 4° Congreso Forestal Español, Zaragoza, Spain, 26–30 September 2005. [Google Scholar]
- Marcos, F.; Godino, M.; García, F.; Izquierdo, I.; Villegas, S. Tallares de chopo I-214 a turnos muy cortos. In Proceedings of the 4° Congreso Forestal Español, Zaragoza, Spain, 26–30 September 2005. [Google Scholar]
- Marcos, F.; Villegas, S.; García, F.; Godino, M. Caracterización energética de la biomasa de chopo (Populus x euramericana I-214) en turnos muy cortos. Rev. Energ. 2006. Available online: file:///C:/Users/MDPI/AppData/Local/Temp/16541-Texto%20del%20art%C3%ADculo-16533-1-10-20140611-1.pdf (accessed on 15 November 2020).
- Llorente, M.J.F.; Laplaza, J.M.M.; Cuadrado, R.E.; García, J.E.C. Ash behaviour of lignocellulosic biomass in bubbling fluidised bed combustion. Fuel 2006, 85, 1157–1165. [Google Scholar] [CrossRef]
- Bartolomé, C.; Gil, A. Ash deposition and fouling tendency of two energy crops (cynara and poplar) and a forest residue (pine chips) co-fired with coal in a pulverized fuel pilot plant. Energy Fuel 2013, 27, 5878–5889. [Google Scholar] [CrossRef]
- Díaz-Ramírez, M.; Boman, C.; Sebastián, F.; Royo, J.; Xiong, S.; Boström, D. Ash characterization and transformation behavior of the fixed-bed combustion of novel crops: Poplar, brassica, and cassava fuels. Energy Fuel 2012, 26, 3218–3229. [Google Scholar] [CrossRef]
- Díaz-Ramírez, M.; Sebastián, F.; Royo, J.; Rezeau, A. Influencing factors on NOX emission level during grate conversion of three pelletized energy crops. Appl. Energy 2014, 115, 360–373. [Google Scholar] [CrossRef]
- Cai, J.; He, Y.; Yu, X.; Banks, S.W.; Yang, Y.; Zhang, X.; Yu, Y.; Liu, R.; Bridgwater, A.V. Review of physicochemical properties and analytical characterization of lignocellulosic biomass. Renew. Sustain. Energy Rev. 2017, 76, 309–322. [Google Scholar] [CrossRef]
- Vassilev, S.V.; Baxter, D.; Andersen, L.K.; Vassileva, C.G. An overview of the chemical composition of biomass. Fuel 2010, 89, 913–933. [Google Scholar] [CrossRef]
- Bartolomé, C.; Gil, A. Emissions during co-firing of two energy crops in a PF pilot plant: Cynara and poplar. Fuel Process. Technol. 2013, 113, 75–83. [Google Scholar] [CrossRef]
- González-García, S.; Moreira, M.T.; Feijoo, G.; Murphy, R.J. Comparative life cycle assessment of ethanol production from fast-growing wood crops (black locust, eucalyptus and poplar). Biomass Bioenergy 2012, 39, 378–388. [Google Scholar] [CrossRef]
- Dallemand, J.F.; Petersen, J.E.; Karp, A. Short Rotation Forestry, Short Rotation Coppice and Perennial Grasses in the European Union: Agro-Environmental Aspects, Present Use and Perspectives; JRC Scientific and Technical Reports: Luxembourg, 2007; pp. 106–111. [Google Scholar]
- Dimitriou, I.; Rutz, D. Sustainability Criteria and Recommendations for Short Rotation Woody Crops; IEE Project SRCplus: Munich, Germany, 2014. [Google Scholar]
- Zalesny, R.S., Jr.; Donner, D.M.; Coyle, D.R.; Headlee, W.L. An approach for siting poplar energy production systems to increase productivity and associated ecosystem services. For. Ecol. Manag. 2012, 284, 45–58. [Google Scholar] [CrossRef]
- Schweier, J.; Molina-Herrera, S.; Ghirardo, A.; Grote, R.; Díaz-Pinés, E.; Kreuzwieser, J.; Haas, E.; Butterbach-Bahl, K.; Rennenberg, H.; Schnitzler, J.P. Environmental impacts of bioenergy wood production from poplar short-rotation coppice grown at a marginal agricultural site in Germany. Bioenergy 2017, 9, 1207–1221. [Google Scholar] [CrossRef]
- El Kasmioui, O.; Ceulemans, R. Financial analysis of the cultivation of poplar and willow for bioenergy. Biomass Bioenergy 2012, 43, 52–64. [Google Scholar] [CrossRef]
- Manzone, M.; Bergante, S.; Facciotto, G. Energy and economic evaluation of a poplar plantation for woodchips production in Italy. Biomass Bioenergy 2014, 60, 164–170. [Google Scholar] [CrossRef]
- Gasol, C. Environmental and Economic Integrated Assessment of Local Energy Crops Production in Southern Europe. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2009. [Google Scholar]
- González-García, S.; Gasol, C.M.; Gabarrell, X.; Rieradevall, J.; Moreira, M.T.; Feijoo, G. Environmental profile of ethanol from poplar biomass as transport fuel in Southern Europe. Renew. Energy 2010, 35, 1014–1023. [Google Scholar] [CrossRef]
- Blanc, S.; Gasol, C.M.; Martínez-Blanco, J.; Muñoz, P.; Coello, J.; Casals, P.; Mosso, A.; Brun, F. Economic profitability of agroforestry in nitrate vulnerable zones in Catalonia (NE Spain). Span. J. Agric. Res. 2019, 17, 16. [Google Scholar] [CrossRef]
- Butnar, I.; Rodrigo, J.; Gasol, C.M.; Castells, F. Life-cycle assessment of electricity from biomass: Case studies of two biocrops in Spain. Biomass Bioenergy 2010, 34, 1780–1788. [Google Scholar] [CrossRef]
- Gasol, C.M.; Gabarrell, X.; Anton, A.; Rigola, M.; Carrasco, J.; Ciria, P.; Rieradevall, J. LCA of poplar bioenergy system compared with Brassica carinata energy crop and natural gas in regional scenario. Biomass Bioenergy 2009, 33, 119–129. [Google Scholar] [CrossRef]
- Relaño, R.L.; Esteban, E.T.; Rodríguez, S.J.H. Economic and Life Cycle Assessment of integrated wood and chips harvesting from hybrid poplar plantations in the Genil Valley (Spain). Comparison with chips harvesting from Poplar SRCs. In Proceedings of the From Theory to Practice: Challenges for Forest Engineering, Warsaw, Poland, 4–7 September 2016; p. 249. [Google Scholar]
- Gasol, C.M.; Martínez, S.; Rigola, M.; Rieradevall, J.; Anton, A.; Carrasco, J.; Ciria, P.; Gabarrell, X. Feasibility assessment of poplar bioenergy systems in the Southern Europe. Renew. Sustain. Energy Rev. 2009, 13, 801–812. [Google Scholar] [CrossRef]
- Li, W.; Ciais, P.; Makowski, D.; Peng, S. A global yield dataset for major lignocellulosic bioenergy crops based on field measurements. Sci. Data 2018, 5, 180169. [Google Scholar] [CrossRef] [PubMed]
- Don, A.; Osborne, B.; Hastings, A.; Skiba, U.; Carter, M.S.; Drewer, J.; Flessa, H.; Freibauer, A.; Hyvönen, N.; Jones, M.B. Land-use change to bioenergy production in Europe: Implications for the greenhouse gas balance and soil carbon. Glob. Chang. Biol. Bioenergy 2012, 4, 372–391. [Google Scholar] [CrossRef]
- Dimitriou, I.; Rutz, D. Sustainable Short Rotation Coppice: A Handbook; Rutz, D., Ed.; WIP Renewable Energies: Munich, Germany, 2015. [Google Scholar]
- Rowe, R.L.; Street, N.R.; Taylor, G. Identifying potential environmental impacts of large-scale deployment of dedicated bioenergy crops in the UK. Renew. Sustain. Energy Rev. 2009, 13, 271–290. [Google Scholar] [CrossRef]
- Griffiths, N.A.; Rau, B.M.; Vaché, K.B.; Starr, G.; Bitew, M.M.; Aubrey, D.P.; Martin, J.A.; Benton, E.; Jackson, C.R. Environmental effects of short-rotation woody crops for bioenergy: What is and isn’t known. GCB-Bioenergy 2019, 11, 554–572. [Google Scholar] [CrossRef]
- Pilipović, A.; Orlović, S.; Rončević, S.; Nikolić, N.; Župunski, M.; Spasojević, J. Results of selection of poplars and willows for water and sediment phytoremediation. Agric. For. 2015, 61, 205. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Headlee, W.L.; Gopalakrishnan, G.; Bauer, E.O.; Hall, R.B.; Hazel, D.W.; Isebrands, J.G.; Licht, L.A.; Negri, M.C.; Nichols, E.G. Ecosystem services of poplar at long-term phytoremediation sites in the Midwest and Southeast, United States. Willey Interdiscip. Rev. Energy Environ. 2019, 8, e349. [Google Scholar] [CrossRef]
- Pandey, V.C.; Bajpai, O.; Singh, N. Energy crops in sustainable phytoremediation. Renew. Sustain. Energy Rev. 2016, 54, 58–73. [Google Scholar] [CrossRef]
- Menéndez, J.; Loredo, J. Use of Reclaimed Surface Mines for Heat and Power Production. Int. J. Renew. Energy Res. 2018, 3, 2367–9123. [Google Scholar]
- Castaño-Díaz, M.; Barrio-Anta, M.; Afif-Khouri, E.; Cámara-Obregón, A. Willow short rotation coppice trial in a former mining area in Northern Spain: Effects of clone, fertilization and planting density on yield after five years. Forests 2018, 9, 154. [Google Scholar] [CrossRef]
- de Bustamante, I.; Lillo, J.; Hernández, J.; Leal, M.; Meffe, R.; de Santiago, A.; Martínez-Hernández, V. El chopo como materia prima e instrumento medioambiental. In Proceedings of the II Simposio del chopo, Valladolid, Spain, 17–19 October 2018. [Google Scholar]
- Rodríguez-Soalleiro, R.; Eimil-Fraga, C.; Gómez-García, E.; García-Villabrille, J.D.; Rojo-Alboreca, A.; Muñoz, F.; Oliveira, N.; Sixto, H.; Pérez-Cruzado, C. Exploring the factors affecting carbon and nutrient concentrations in tree biomass components in natural forests, forest plantations and short rotation forestry. For. Ecosyst. 2018, 5, 35. [Google Scholar] [CrossRef]
- Oliveira, N.; Fuertes, A.; González, I.; de la Iglesia, J.P.; Cañellas, I.; Rodríguez-Soalleiro, R.; Sixto, H. Cultivo de chopo para la producción de biomasa: Estrategia de mitigación mediante la acumulación de carbono en las diferentes fracciones. In Proceedings of the VIII Remedia Workshop. Economía Circular como Catalizador de la Sostenibilidad Medioambiental del Sector Primario Español, Elche, Spain, 22–23 September 2020. [Google Scholar]
- Rodríguez, F.; Serrano, L.; Aunós, A. El papel del chopo como sumidero de CO2 atmosférico. In Proceedings of the 4° Congreso Forestal Español, Zaragoza, Spain, 26–30 September 2005. [Google Scholar]
- Djomo, S.N.; Kasmioui, O.E.; Ceulemans, R. Energy and greenhouse gas balance of bioenergy production from poplar and willow: A review. Glob. Chang. Biol. Bioenergy 2011, 3, 181–197. [Google Scholar] [CrossRef]
- Pérez-Cruzado, C.; Mohren, G.M.; Merino, A.; Rodríguez-Soalleiro, R. Carbon balance for different management practices for fast growing tree species planted on former pastureland in southern Europe: A case study using the CO2 Fix model. Eur. J. For. Res. 2012, 131, 1695–1716. [Google Scholar] [CrossRef]
- Pérez-Cruzado, C.; Mansilla-Salinero, P.; Rodríguez-Soalleiro, R.; Merino, A. Influence of tree species on carbon sequestration in afforested pastures in a humid temperate region. Plant Soil 2012, 353, 333–353. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Broeckx, L.S.; Zona, D.; Berhongaray, G.; De Groote, T.; Serrano, M.C.; Janssens, I.A.; Ceulemans, R. Net ecosystem production and carbon balance of an SRC poplar plantation during its first rotation. Biomass Bioenergy 2013, 56, 412–422. [Google Scholar] [CrossRef]
- Ceulemans, R.; Horemans, J.A.; Vanbeveren, S.P.P. Short-rotation crops reduce greenhouse gas emissions and contribute to climate change mitigation. In Proceedings of the 7th International Poplar Symposium. New Bioeconomies: Exploring the potential role of Salicaceae, Buenos Aires, Argentina, 28 October–4 November 2018. [Google Scholar]
- Folch, A.; Ferrer, N. The impact of poplar tree plantations for biomass production on the aquifer water budget and base flow in a Mediterranean basin. Sci. Total Environ. 2015, 524, 213–224. [Google Scholar] [CrossRef]
- MINETAD. La Energía en España 2016; Ministerio de Energía, Turismo y Agenda Digital: Madrid, Spain, 2017; p. 342. [Google Scholar]
- Unión por la Biomasa. Balance Socioeconómico de las Biomasas en ESPAÑA 2017–2021; Unión por la Biomasa: Spain, 2018; p. 30. Available online: https://www.interempresas.net/Energia/Articulos/219726-Union-por-Biomasa-presenta-’Balance-socioeconomico-de-biomasas-en-Espana-2017-2021’.html (accessed on 15 November 2020).
- IDAE. Biomasa: Cultivos Energéticos; Instituto para la Diversificación y Ahorro de la Energía: Madrid, Spain, 2007. [Google Scholar]
- Eurostat. Renewable Energy Statistics; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- WBA. Global Bioenergy Statistics; World Bioenergy Association: Stockholm, Sweden, 2019; p. 58. [Google Scholar]
- APPA. Estudio del Impacto Macroeconómico de las ENERGÍAS Renovables en España; Asociación de Empresas de Energías Renovables: Spain, 2018; Available online: https://www.appa.es/wp-content/uploads/2018/10/Estudio_del_impacto_Macroeconomico_de_las_energias_renovables_en_Espa%C3%B1a_2017.pdf (accessed on 15 November 2020).





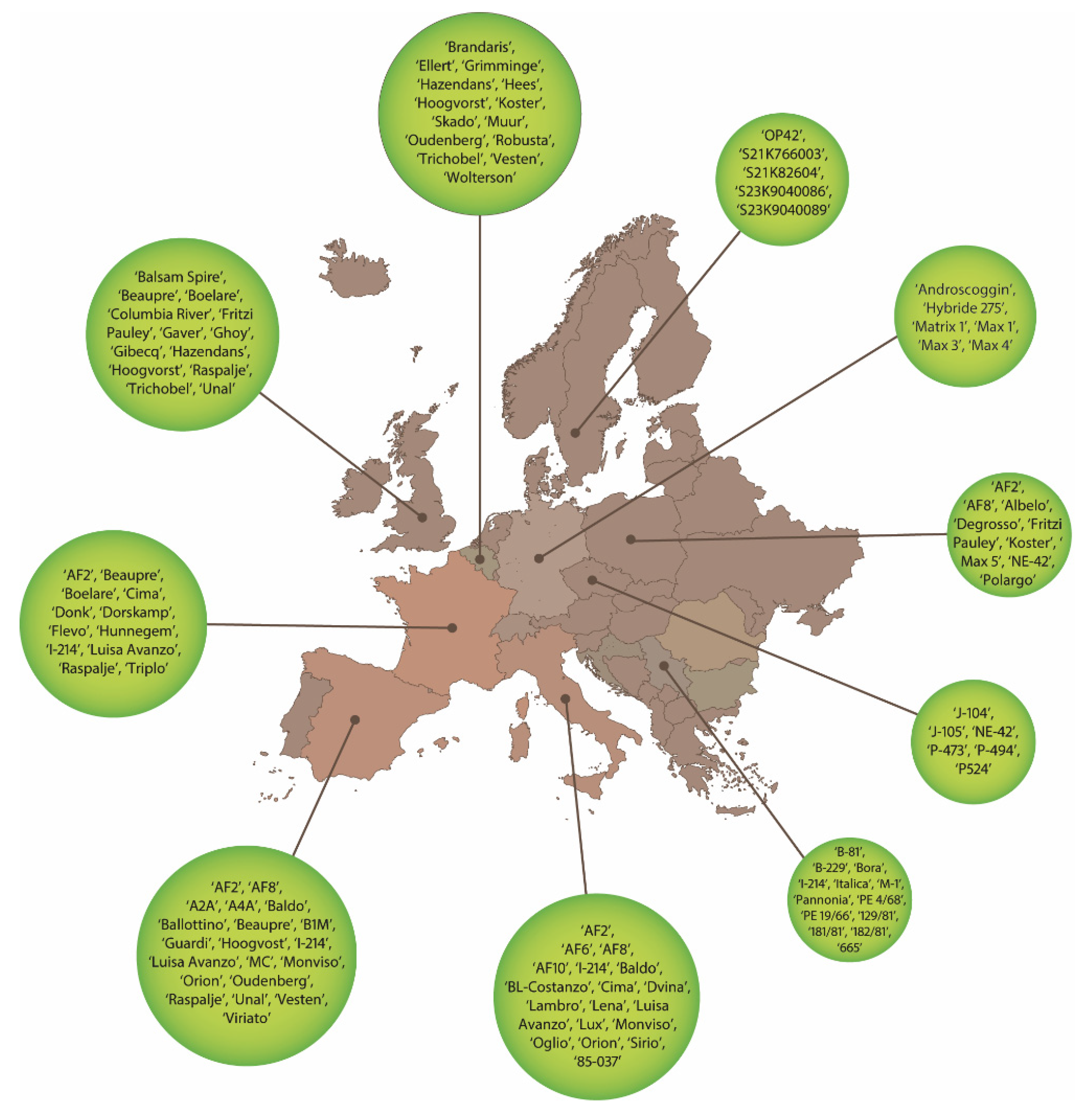{kind=link}
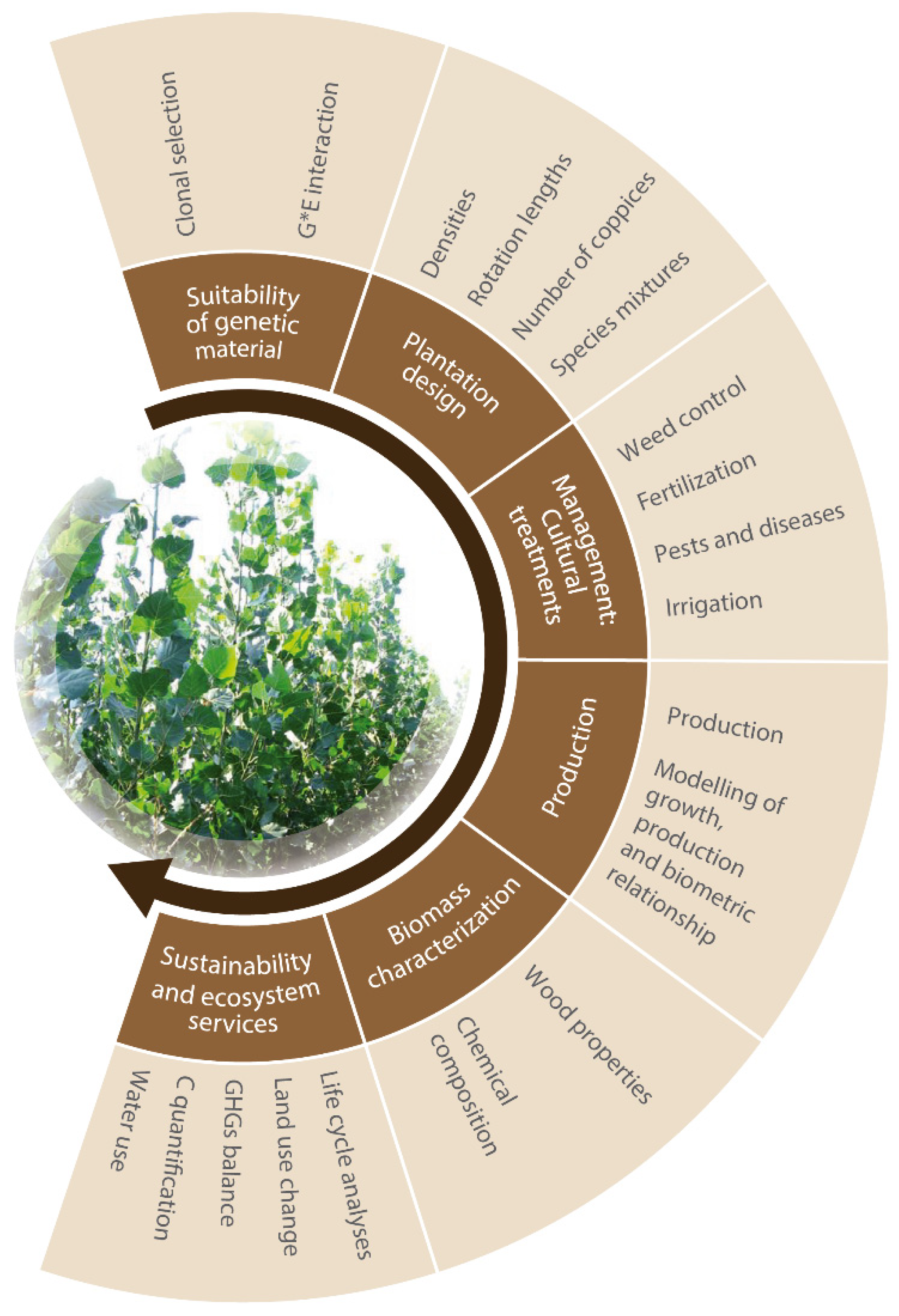{kind=link}
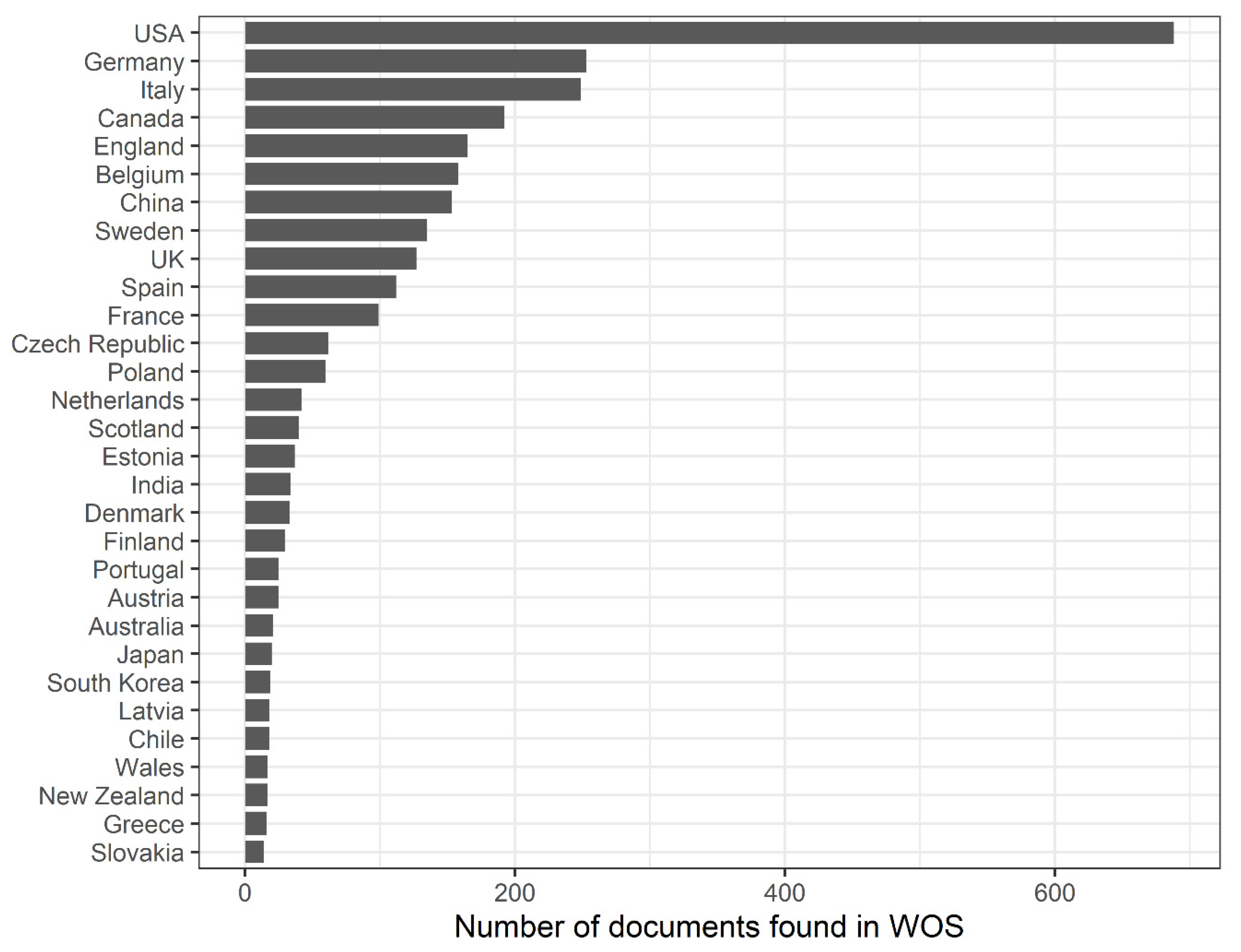{kind=link}
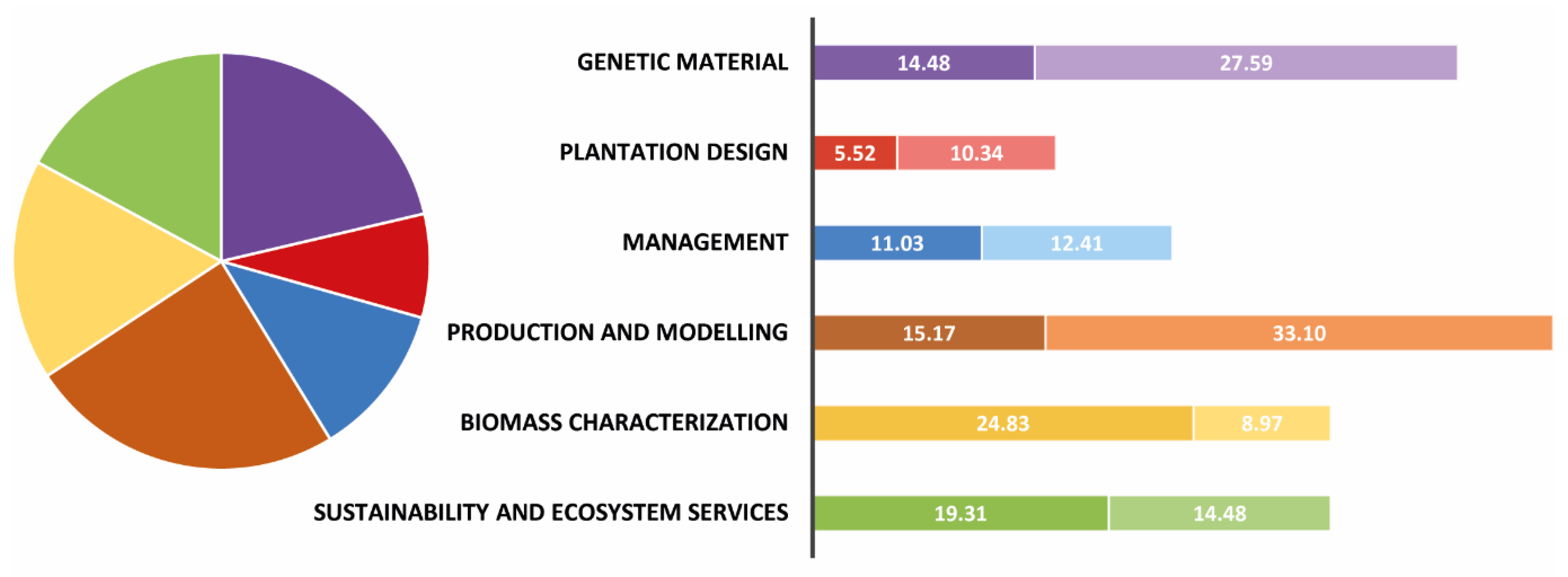{kind=link}
| Parentage | Section | Genotypes | |
|---|---|---|---|
| P. × canadensis Mönch | D × N | Aigeiros | ‘2000 Verde’, ‘Agathe F’ a, ‘E-298′ a, ‘Branagesi’, ‘B-1M’, ‘Canadá Blanco’, ‘Dorskamp’, ‘Flevo’, ‘Guardi’, ‘I-214′ b, ‘Campeador’ b, ‘I-454/40′, ‘Luisa Avanzo’, ‘MC’, ‘Triplo’ |
| P. deltoides W. Bartram ex Marshall | D | Aigeiros | ‘Lux’, ‘Viriato’ |
| P. × generosa Henry | T × D | Tacamahaca × Aigeiros | ‘Beaupre’, ‘Boelare’, ‘Raspalje’, ‘Unal’, ‘USA 49-177’ |
| P. × generosa Henry × P. alba L. | (T × D) × A | (Tacamahaca × Aigeiros) × Populus | ‘I-114/69′ |
| P. nigra L. | N | Aigeiros | ‘Tr 56/75’, ‘Bordils’, ‘Lombardo leones’ |
| Species and Poplar Hybrids | Genotype | Density | Age | Country | Yield (Mg dm ha−1 year−1) by Rotation | Reference | ||
|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | ||||||
| P. deltoides × P. trichocarpa | ‘IBW2’ | 10,000 | 4 | Belgium | 2.5 | 1.6 | [19,219] | |
| P. nigra | ‘Wolterson’ | 8.1 | 9.7 | |||||
| P. trichocarpa | ‘Columbia River’ | 7.8 | ||||||
| ‘Fritzi Pauley’ | 8.1 | 8.2 | ||||||
| ‘Trichobel’ | 8.4 | 8.2 | ||||||
| P. × canadensis | ‘Gibecq’ | 1.6 | 3.5 | |||||
| P. × generosa | ‘Hazendans’ | 10.8 | 3 | |||||
| ‘Hoogvorst’ | 10.1 | 8.2 | ||||||
| P. deltoides × P. × generosa | ‘Grimminge’ | 8000 | 2 (In the 3rd rot the age is 3 year) | Belgium | 4.3 | 11.7 | 8.4 | [136] * |
| P. nigra | ‘Brandaris’ | 1.4 | 7.0 | 8.7 | ||||
| P. nigra × P. maximowiczii | ‘Wolterson’ | 2.7 | 11.7 | 14.6 | ||||
| P. trichocarpa × P. maximowiczii | ‘Bakan’ | 4.9 | 14.3 | 18.1 | ||||
| ‘Skado’ | 5.7 | 16.8 | 20.8 | |||||
| P. × canadensis | ‘Ellert’ | 3.3 | 11.1 | 19.5 | ||||
| ‘Hees’ | 6.5 | 15.5 | 26.0 | |||||
| ‘Koster’ | 2.8 | 9.5 | 14.1 | |||||
| ‘Muur’ | 3.9 | 12.4 | 17.1 | |||||
| ‘Oudenberg’ | 3.7 | 13.4 | 14.0 | |||||
| ‘Robusta’ | 2.5 | 8.1 | 15.6 | |||||
| ‘Vesten | 4.7 | 12.6 | 16.1 | |||||
| P. maximowiczii × P. trichocarpa | ‘NE-42’ | 2222 | 3 (2nd rot: 4 year) | Czech Republic | 1.0–1.4 | 9.4–9.8 | 9.1–11.4 | [220] |
| 3 (2nd rot: 2 year) | 8.3 | 15.4 | 18.9 | |||||
| P. balsamifera × P. tremula | ‘P-524’ | 10,000 | 6 | Czech Republic | 8.1 | [221] | ||
| P. maximowiczii × P. berolinensis | ‘P-494’ | 10.2 | ||||||
| P. nigra | P. nigra | 2.6 | ||||||
| P. nigra × P. maximowiczii | ‘J-104’ | 11.9 | ||||||
| ‘J-105’ | 13.9 | |||||||
| P. × generosa | ‘P-473’ | 9.7 | ||||||
| P. × generosa | ‘Beaupré’ | 3030 | 3 | France | 1.57–11.13 | [222] | ||
| 4 | 2.73–12.7 | |||||||
| ‘Boelare’ | 3 | 1.37–10.53 | ||||||
| 4 | 2.85–11.6 | |||||||
| ‘Hunnegem’ | 3 | 12.1 | ||||||
| 4 | 13.68 | |||||||
| ‘Raspalje’ | 3 | 2.2 | ||||||
| 4 | 3.63 | |||||||
| P. × generosa | ‘Beaupré’ | 8333 | 8 | Germany | 6.1 | [133] * | ||
| ‘Rap’ | 5.8 | |||||||
| P. nigra × P. maximowiczii | ‘Max 1’ | 3.3 | ||||||
| ‘Max 3’ | 3.5 | |||||||
| ‘Max 4’ | 3 | |||||||
| P. maximowiczii × P. trichocarpa | ‘Androscoggin’ | 3.3 | ||||||
| ‘Hybride 275’ | 4.2 | |||||||
| P. trichocarpa | ‘Muhle Larsen’ | 3.7 | ||||||
| P. maximowiczii × P. nigra | ‘Max 1’ | 8890 | 2 | Germany | 1.0–1.59 | [223] | ||
| 9250 | 6.81 | |||||||
| P. maximowiczii × P. nigra | ‘Max 3’ | 17,778 | 4 | Germany | 8.6 | [48] | ||
| P. maximowiczii × P. trichocarpa | ‘Androscoggin’ | 10.5 | ||||||
| P. maximowiczii × P. nigra | ‘Max 4’ | 11,000 | 2 | Germany | 5.4–6.3 | [224] | ||
| P. deltoides | ‘Baldo’ | 5747 | 2 | Italy | 4.75 | [90] | ||
| 10,000 | 17.5 | |||||||
| ‘Dvina’ | 10,000 | 11.8 | ||||||
| ‘Lambro’ | 10,000 | 9.5 | ||||||
| ‘Lena’ | 10,000 | 14.2 | ||||||
| ‘Lux’ | 5714 | 7.05 | ||||||
| 8333 | 4.3 | |||||||
| ‘Oglio’ | 10,000 | 14.1 | ||||||
| P. × canadensis | ‘BL-Costanzo’ | 5714 | 3.25 | |||||
| 7142 | 4.45 | |||||||
| ‘Cima’ | 5714 | 4.55 | ||||||
| 7142 | 5.7 | |||||||
| ‘I-214’ | 5747 | 4.35 | ||||||
| 10,000 | 8.4 | |||||||
| ‘Luisa Avanzo’ | 5714 | 5.55 | ||||||
| 7142 | 6.25 | |||||||
| ‘Orion’ | 5747 | 4.46 | ||||||
| 10,000 | 17.0 | |||||||
| P. deltoides | ‘Lux’ | 10,000 | 2 | Italy | 12.38 | 6.99 | 4.99 | [225] |
| 3 | 7.07 | 14.53 | ||||||
| P. deltoides | ‘Lux’ | 10,000 | 1 | Italy | 16.4 | [135] | ||
| 2 | 22.5 | |||||||
| 3 | 24.3 | |||||||
| P. deltoides | ‘Lux’ | 5900 | 2 | Italy | 3.31–9.33 | 2.17–18.85 | [226] | |
| P. maximowiczii × P. nigra | ‘AF10’ | 14.84 | 20.74 | |||||
| P. × canadensis | ‘AF2’ | 5.66–15.77 | 7.54–17.01 | |||||
| ‘I-214’ | 3.63–11.87 | 4.2–18.44 | ||||||
| ‘Sirio’ | 5.65 | 8.55 | ||||||
| P. × generosa × P. nigra | ‘AF6’ | 5.76–15.04 | 5.79–17.07 | |||||
| ‘Monviso’ | 6.79–17.92 | 9.31–23.55 | ||||||
| P. × generosa × P. trichocarpa | ‘AF8’ | 4.75–14.93 | 5.04–24.05 | |||||
| P. × canadensis | ‘AF2’ | 6061 | 2 | Italy | 16 | 17 | 17 | [92] * |
| ‘I-214’ | 11.5 | 18 | 13.5 | |||||
| P. × generosa × P. nigra | ‘AF6’ | 15 | 17.5 | 15 | ||||
| ‘Monviso’ | 17.5 | 23.5 | 17.5 | |||||
| P. × generosa × P. trichocarpa | ‘AF8’ | 15 | 24 | 19 | ||||
| P. maximowiczii × P. trichocarpa | ‘NE-42’ | 1333 | 7 | Poland | 8 | [52] | ||
| P. trichocarpa × P. trichocarpa | ‘Fritzi Pauley’ | 8.1 | ||||||
| P. × canadensis | ‘AF2’ | 4.1 | ||||||
| ‘Albelo’ | 4 | |||||||
| ‘Degrosso’ | 6.8 | |||||||
| ‘Koster’ | 5.6 | |||||||
| ‘Polargo’ | 4.3 | |||||||
| P. × generosa × P. trichocarpa | ‘AF8’ | 2.5 | ||||||
| P. nigra × P. maximowiczii | ‘Max 5’ | 11,110 | 3 | Poland | 7.8 | [227] | ||
| P. trichocarpa | ‘Columbia River’ | 10,000 | 3 | United Kingdom | 6.71 | 6.62 | [13] | |
| ‘Fritzi Pauley’ | 8.59 | 8.24 | ||||||
| ‘Trichobel’ | 9.08 | 9.59 | ||||||
| P. trichocarpa × P. balsamifera | ‘Balsam Spire’ | 7.24 | 7.03 | |||||
| P. × canadensis | ‘Gaver’ | 6.58 | 5.58 | |||||
| ‘Ghoy’ | 6.45 | 5.77 | ||||||
| ‘Gibecq’ | 5.7 | 4.73 | ||||||
| P. × generosa | ‘Beaupré’ | 7.34 | 4.87 | |||||
| ‘Boelare’ | 6.23 | 4.2 | ||||||
| ‘Hazendans’ | 7.23 | 7.56 | ||||||
| ‘Hoogvorst’ | 8.84 | 8.12 | ||||||
| ‘Raspalje’ | 6.69 | 4.66 | ||||||
| ‘Unal’ | 7.55 | 5.25 | ||||||
| P. balsamifera × P. trichocarpa | ‘Balsam Spire’ | 10,000 | 5 | United Kingdom | 14.42 | [228] | ||
| 4444 | 5 | 11.20 | ||||||
| 10,000 | 4 | 13.98 | ||||||
| Species and Poplar Hybrids | Genotype | Density | Age | Location | Yield (Mg dm ha−1 year−1) by Rotation | Reference | ||
|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | ||||||
| P. × canadensis | ‘AF2’ | 13,333 | 3 | Aragón | 24.72 | [229] * | ||
| ‘Ballottino’ | 25.08 | |||||||
| ‘I-214’ | 28.64 | |||||||
| P. × generosa × P. nigra | ‘Monviso’ | 26.91 | ||||||
| P. × canadensis | ‘Campeador’ | 10,000 | 5 | Soria | 12.10 | [230] | ||
| ‘Dorskamp’ | 6.3 | |||||||
| P. × generosa | ‘Boelare’ | 16.6 | ||||||
| ‘Raspalje’ | 19.8 | |||||||
| P. × canadensis | ‘Campeador’ | 10,000 | 6 | Soria | 9.3 | [126,127] | ||
| ‘I-214’ | 9.9 | |||||||
| P. × generosa | ‘Boelare’ | 12.9 | ||||||
| ‘Raspalje’ | 2500 | 8.0 | ||||||
| 5000 | 9.2 | |||||||
| 10,000 | 6 (2nd–3rd:5year) | 16.5 | 17.2 | 14.8 | ||||
| 10,000 | 3 | 9.8 | ||||||
| 20,000 | 12.7 | |||||||
| 40,000 | 15.6 | |||||||
| P. × canadensis | ‘AF2’ | 8000 | 7 | Lugo | 0.84 | [231] Without irrigation in a transition zone between Mediterranean and Atlantic climate | ||
| ‘I-214’ | 2.96 | |||||||
| P. × generosa | ‘Beaupré’ | 3.07 | ||||||
| ‘Raspalje’ | 3.1 | |||||||
| ‘Unal’ | 1.97 | |||||||
| P. × generosa × P. nigra | ‘AF6’ | 1.14 | ||||||
| ‘Monviso’ | 0.93 | |||||||
| P. trichocarpa | ‘Trichocarpa’ | 3.93 | ||||||
| P. × canadensis | ‘AF2’ and ‘I-214’ | 33,333 | 3 | Soria | 7–12 | 2–9.5 | 1.5–4.5 | [145] * |
| P. × canadensis | ‘I-214’ | 10,000 | 3 | Asturias | 9 | 11 | 6 | [148] * Without irrigation in mining zone |
| P. × canadensis | ‘I-214’ | 37,037 | 2 | Salamanca | 12.41 | 9.99 | 24.83 | [232] |
| P. × canadensis | ‘AF2’ | 13,333 | 3 | Aragón | 15.53 | [233] *, [234] | ||
| ‘I-214’ | 10.3 | |||||||
| ‘Guardi’ | 9.03 | |||||||
| P. deltoides | ‘Viriato’ | 14 | ||||||
| P. × generosa | ‘Unal’ | 9.23 | ||||||
| P. × generosa × P. nigra | ‘Monviso’ | 13.13 | ||||||
| P. × canadensis | ‘I-214’ | 33,333 | 2 | Spain | 15.1 | [112] | ||
| Populus spp. | 4 genotypes | 13,333 | 3 | Granada | 13.7 | [235] | ||
| Populus spp. | 3 genotypes | 19,700 | 4 | Soria | 12.0 | |||
| Populus spp. | 4 genotypes | 13,333 | 3 | Zamora | 7.7 | |||
| Populus spp. | 4 genotypes | 33,333 | 3 | León | 6.9 | |||
| Populus spp. | 4 genotypes | 20,000 | 3 | Navarra | 16 | |||
| P. nigra | ‘Lombardo leones’ | 17,316 | 3 | Soria | 9.1 | 17.1 | [236] | |
| P. × canadensis | ‘A4A’ | 20.0 | 12.2 | |||||
| ‘A2A’ | 11.8 | 15.3 | ||||||
| ‘I-214’ | 22.0 | 30.1 | ||||||
| ‘Luisa Avanzo’ | 17.3 | 27.1 | ||||||
| ‘MC’ | 16.8 | 18.4 | ||||||
| ‘2000 Verde’ | 13.64 | 14.25 | ||||||
| P. deltoindes | ‘Baldo’ | 10,000 | 3 | Gerona, Madrid, Soria | 8.25–19.07 | 8.59–29.94 | [77,80] | |
| P. × canadensis | ‘Ballottino’ | 8.25–15.94 | 11.45–19.20 | |||||
| ‘I-214’ | 5.45–16.16 | 11.09–37.27 | ||||||
| ‘Orion’ | 12.53–22.65 | 12.70–34.56 | ||||||
| ‘Oudenberg’ | 9.43–14.27 | 9.66–19.96 | ||||||
| P. × canadensis | ‘I-214’ | 6666-33,333 | 3 | Barcelona | 4.2 | [47] | ||
| Gerona | 5.7 | |||||||
| Gerona | 11.6 | |||||||
| Granada | 18.3 | |||||||
| León | 4.7 | |||||||
| León | 5.8 | |||||||
| León | 15.2 | |||||||
| Madrid | 13.2 | |||||||
| Navarra | 18.5 | |||||||
| Soria | 8.1 | |||||||
| Soria | 14.2 | |||||||
| Zamora | 8.2 | |||||||
| P. × canadensis | ‘I-214’ | 5000 | 4 | Madrid | 12.4 | [44] | ||
| ‘Campeador’ | 10.0 | |||||||
| ‘Canada Blanco’ | 8.4 | |||||||
| ‘I-214’ | 2 | 10.90 | ||||||
| ‘I-214’ | 3 | 10.10 | ||||||
| ‘I-214’ | 5 | 17.30 | ||||||
| ‘I-214’ | 10,000 | 4 | 16.9 | |||||
| P. × canadensis | ‘AF2’ | 13,333 | 3 | Granada | 17.1 | [237] | ||
| P. × canadensis | ‘AF2’ | 33,333 | 3 | León | 16.75 | [122] | ||
| ‘Guardi’ | 16.14 | |||||||
| ‘I-214’ | 15.14 | |||||||
| ‘MC’ | 15.01 | |||||||
| ‘2000 Verde’ | 10.05 | |||||||
| P. × generosa | ‘Unal’ | 11.38 | ||||||
| ‘USA 49-177’ | 15.92 | |||||||
| P. × generosa × P. nigra | ‘Monviso’ | 15.16 | ||||||
| ‘Pegaso’ | 7.37 | |||||||
| P. × canadensis | ‘AF2’ | 33,333 | 3 | León, Gerona, Madrid, Soria | 16.8 | [55] | ||
| ‘Guardi’ | 12.44 | |||||||
| ‘I-214’ | 12.21 | |||||||
| ‘MC’ | 15.32 | |||||||
| ‘2000 Verde’ | 15.38 | |||||||
| P. × generosa | ‘Unal’ | 11.13 | ||||||
| P. × generosa × P. nigra | ‘Monviso’ | 14.47 | ||||||
| ‘Pegaso’ | 6.65 | |||||||
| P. deltoides | ‘Viriato’ | 13,333 | 3 | Granada | 9.55 | 18.20 | 12.82 | [150] |
| P. × canadensis | ‘AF2’ | 13.47 | 16.66 | 13.99 | ||||
| ‘Ballottino’ | 9.48 | 16.75 | 12.27 | |||||
| ‘I-214’ | 9.11 | 13.44 | 15.81 | |||||
| P. × generosa × P. nigra | ‘Monviso’ | 12.29 | 11.57 | 13.92 | ||||
| P. × canadensis | ‘AF2’ | 20,000 | 3 (2nd rot: 2 year) | Navarra | 15.59 | [238] | ||
| ‘A4A’ | 23.80 | 22.12 | ||||||
| ‘Guardi’ | 15.59 | 6.43 | ||||||
| ‘I-214’ | 23.80 | 18.51 | ||||||
| ‘MC’ | 11.21 | 26.51 | ||||||
| ‘Triplo’ | 18.07 | 32.92 | ||||||
| P. deltoides | ‘Viriato’ | 16.68 | 38.18 | |||||
| P. × generosa | ‘Beaupré’ | 21.29 | 15.29 | |||||
| ‘Unal’ | 25.86 | 12.15 | ||||||
| P. × generosa × P. nigra | ‘Monviso’ | 15.41 | ||||||
| ‘Pegaso’ | 13.99 | |||||||
| P. × canadensis | ‘I-214’ | 15,000 | 3 (2nd rot:2 year) | Navarra | 18.20 | 14.46 | [239] | |
| 20,000 | 18.68 | 18.51 | ||||||
| 25,000 | 23.36 | 14.65 | ||||||
| 33,333 | 18.85 | 17.13 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, N.; Pérez-Cruzado, C.; Cañellas, I.; Rodríguez-Soalleiro, R.; Sixto, H. Poplar Short Rotation Coppice Plantations under Mediterranean Conditions: The Case of Spain. Forests 2020, 11, 1352. https://doi.org/10.3390/f11121352
Oliveira N, Pérez-Cruzado C, Cañellas I, Rodríguez-Soalleiro R, Sixto H. Poplar Short Rotation Coppice Plantations under Mediterranean Conditions: The Case of Spain. Forests. 2020; 11(12):1352. https://doi.org/10.3390/f11121352
Chicago/Turabian StyleOliveira, Nerea, César Pérez-Cruzado, Isabel Cañellas, Roque Rodríguez-Soalleiro, and Hortensia Sixto. 2020. "Poplar Short Rotation Coppice Plantations under Mediterranean Conditions: The Case of Spain" Forests 11, no. 12: 1352. https://doi.org/10.3390/f11121352
APA StyleOliveira, N., Pérez-Cruzado, C., Cañellas, I., Rodríguez-Soalleiro, R., & Sixto, H. (2020). Poplar Short Rotation Coppice Plantations under Mediterranean Conditions: The Case of Spain. Forests, 11(12), 1352. https://doi.org/10.3390/f11121352