Pretreatment with High-Dose Gamma Irradiation on Seeds Enhances the Tolerance of Sweet Osmanthus Seedlings to Salinity Stress

Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Collection, Gamma Irradiation and Storage
2.2. Salt Stress and Tolerance Assays
2.3. Sampling for Physiological Index
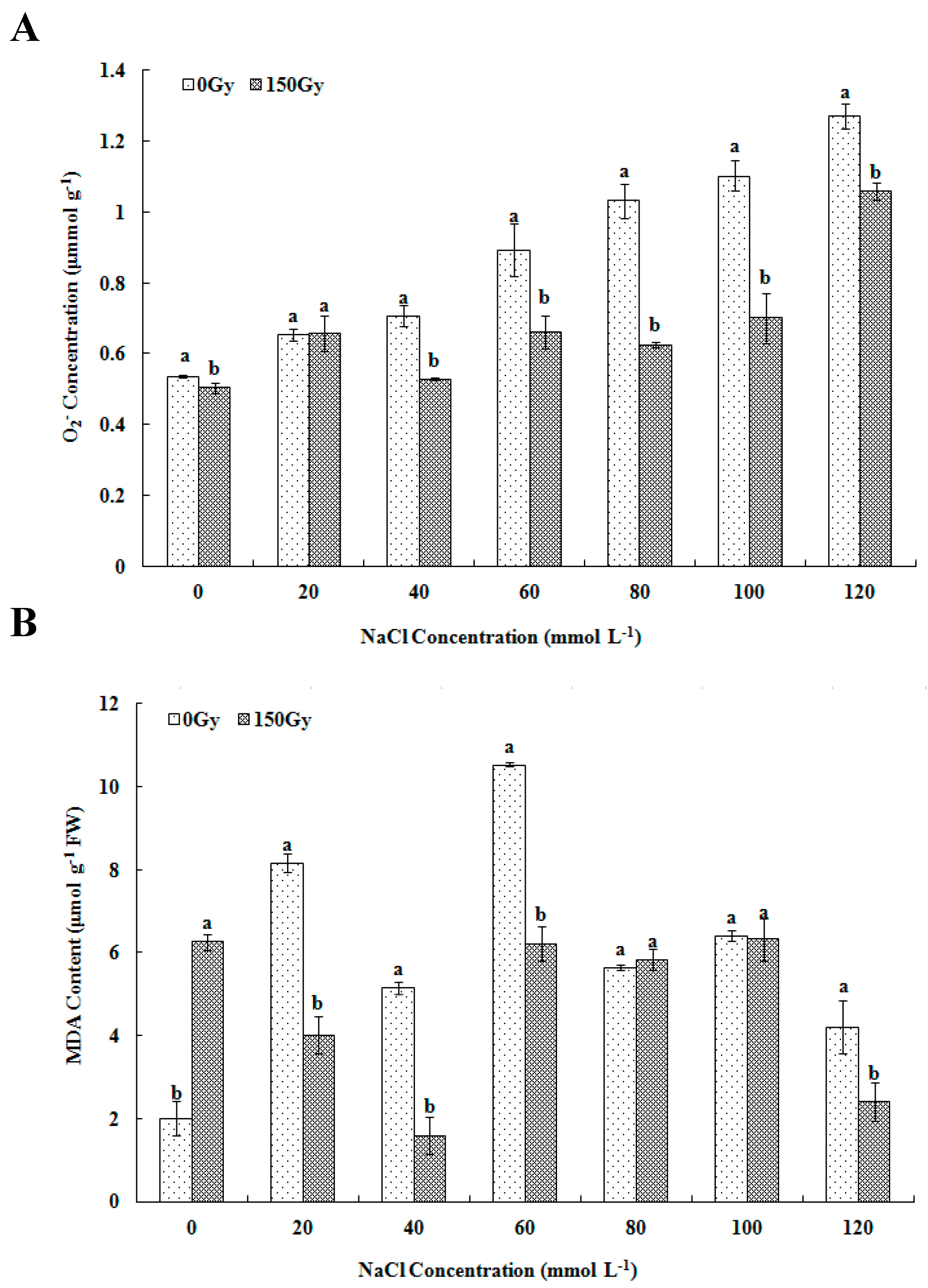
2.3.1. Measurement of O2− Content and MDA Level
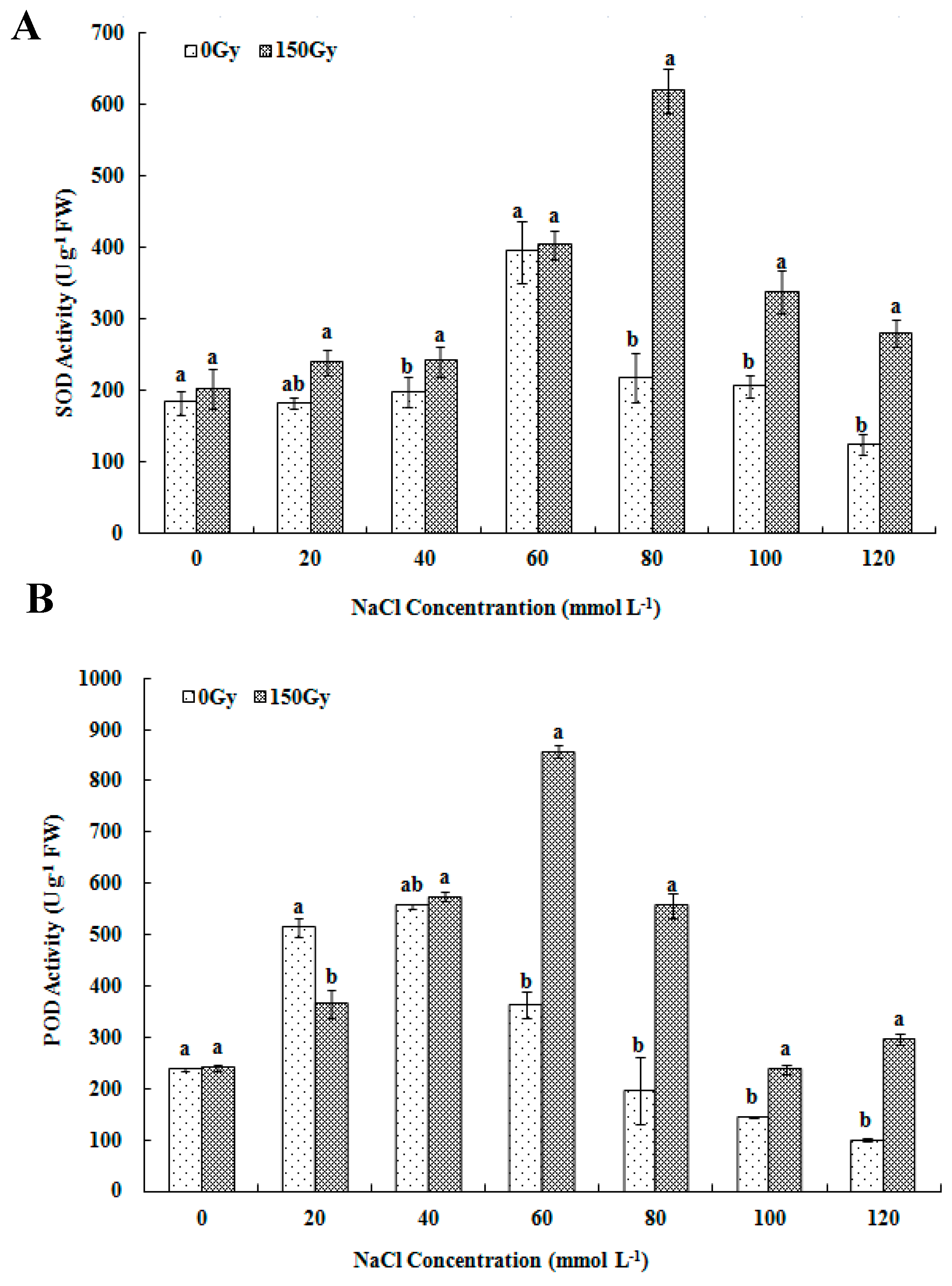
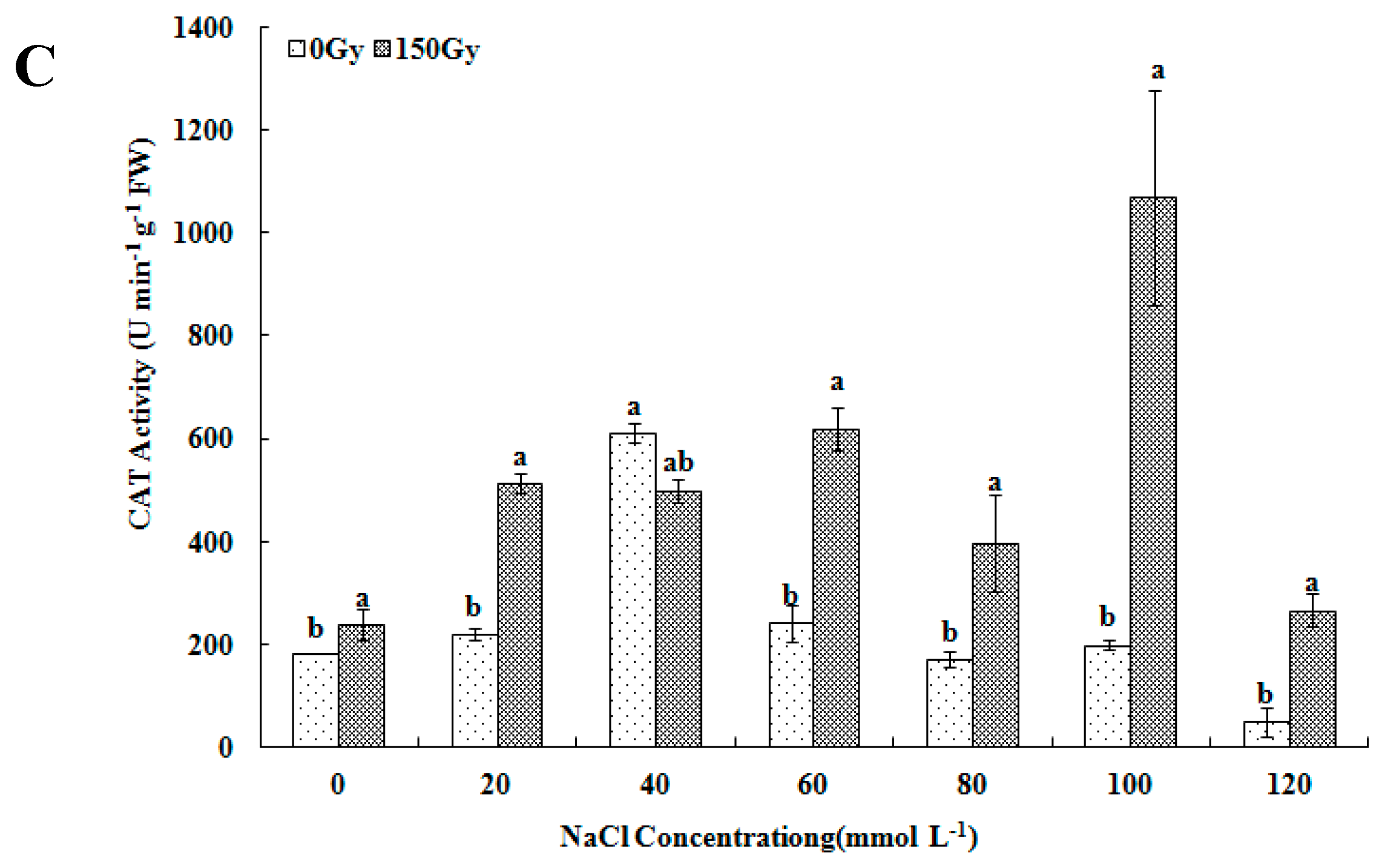
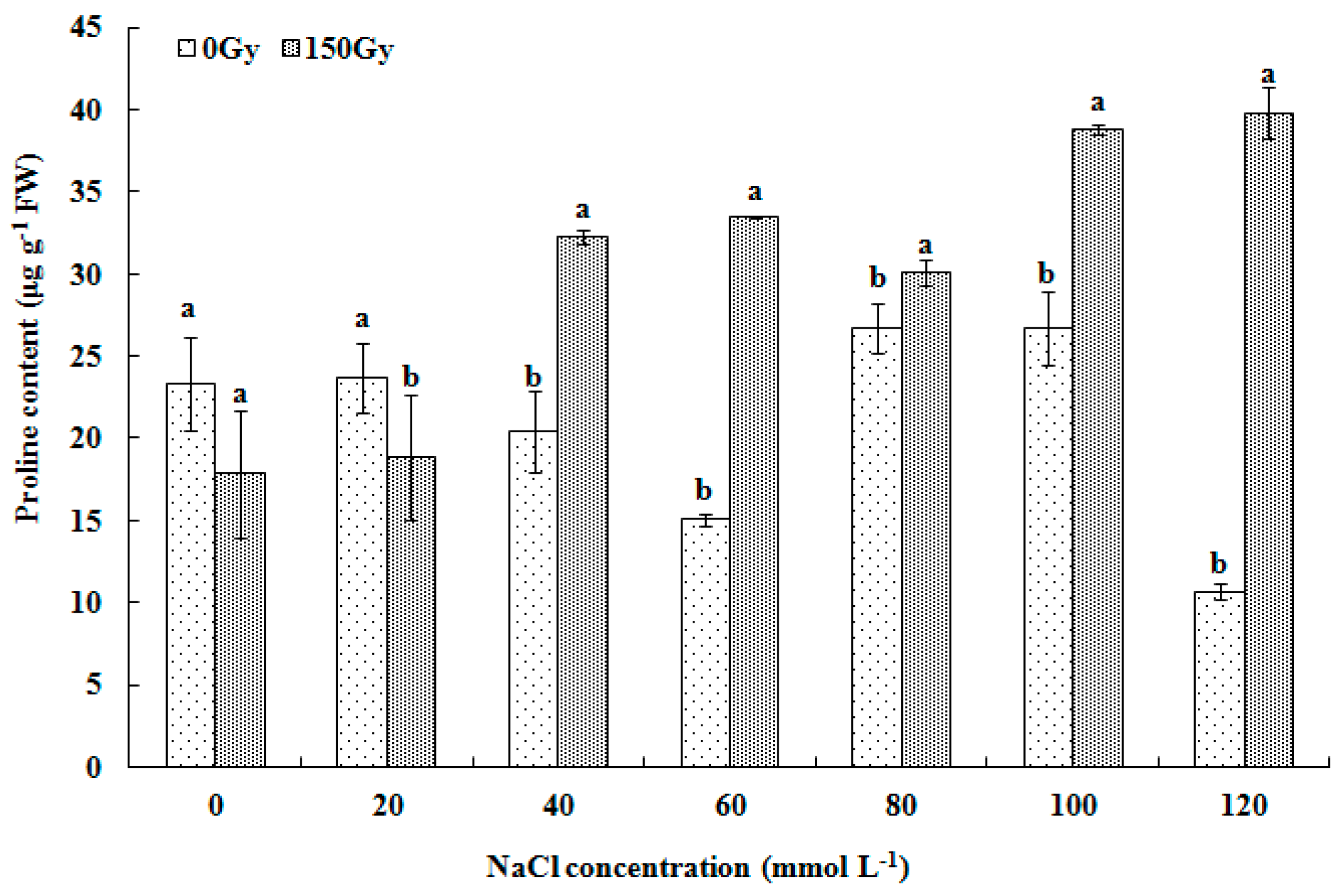
2.3.2. Determination of Antioxidant Enzyme Activity and Proline Level
2.4. Data Analyses
3. Results
3.1. Tolerance in Irradiated Seedlings to Salinity Stress
3.2. Effects of High-Doses Gamma Irradiation on ROS Level and Lipid Peroxidation in Response to Salt Stress
3.3. Effects of High-Doses Gamma Irradiation on ROS Scavenging Systems in Response to Salt Stress
3.4. Effects of High-Doses Gamma Irradiation on Proline Level in Response to Salt Stress
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murtaza, G.; Ghafoor, A.; Owens, G.; Qadir, M.; Kahlon, U.Z. Environmental and economic benefits of saline-sodic soil reclamation using low-quality water and soil amendments in conjunction with a rice-wheat cropping system. J. Agron. Crop Sci. 2009, 195, 124–136. [Google Scholar] [CrossRef]
- Rozema, J.; Flowers, T. Ecology crops for a salinized world. Science 2008, 322, 1478–1480. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good! Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.P.J.C.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Carvalho, M.D. Drought stress and reactive oxygen species, production, scavenging and signaling. Plant Signal Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species, metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Kai, H.; Hirashima, K.; Matsuda, O.; Ikegami, H.; Winkelmann, T.; Nakahara, T.; Iba, K. Thermotolerant cyclamen with reduced acrolein and methyl vinyl ketone. J. Exp. Bot. 2012, 63, 4143–4150. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef]
- Wi, S.G.; Chung, B.Y.; Kim, J.S.; Kim, J.H.; Baek, M.H.; Lee, J.W.; Kim, Y.S. Ultrastructural changes of cell organelles in Arabidopsis stem after gamma irradiation. J. Plant Biol. 2005, 48, 195–200. [Google Scholar] [CrossRef]
- Wi, S.G.; Chung, B.Y.; Kim, J.S.; Kim, J.H.; Baek, M.H.; Lee, J.W.; Kim, Y.S. Effects of gamma irradiation on morphological changes and biological responses in plants. Micron 2007, 38, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, R.; Shu, C.; Zhang, X.C. Screening for cold-resistant tomato under radiation mutagenesis and observation of the submicroscopic structure. Acta Physiol. Plant 2016, 38, 1–12. [Google Scholar] [CrossRef]
- Cho, H.S.; Lee, H.S.; Pai, H.S. Expression patterns of diverse genes in response to gamma irradiation in Nicotiana tabacum. J. Plant Biol. 2000, 43, 82–87. [Google Scholar] [CrossRef]
- Moussa, H.R. Gamma irradiation effects on antioxidant enzymes and G6PDH activities in Vicia Faba plants. J. New Seeds 2008, 9, 89–99. [Google Scholar] [CrossRef]
- Zaka, R.; Vandecasteele, C.M.; Misset, M.T. Effects of low chronic doses of ionizing radiation on antioxidant enzymes and G6PDH activities in Stipa capillata (Poaceae). J. Exp. Bot. 2002, 53, 1979–1987. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Chung, B.Y.; Kim, J.S.; Wi, S.G. Effects of in planta gamma-irradiation on growth, photosynthesis, and antioxidative capacity of red pepper (Capsicum annuum L.). J. Plant Biol. 2005, 48, 47–56. [Google Scholar] [CrossRef]
- Chen, S.; Chai, M.L.; Jia, Y.F.; Gao, Z.S.; Zhang, L.; Gu, M.X. In Vitro Selection of salt tolerant variants following 60Co gamma irradiation of long-term callus cultures of zoysia matrella [L.] merr. Plant Cell Tiss. Organ Cult. 2011, 107, 493–500. [Google Scholar] [CrossRef]
- Qi, W.C.; Zhang, L.; Xu, H.B.; Wang, L.; Jiao, Z. Physiological and molecular characterization of the enhanced salt tolerance induced by low-dose gamma irradiation in Arabidopsis seedlings. Biochem. Biophys. Res. Commun. 2014, 450, 1010–1015. [Google Scholar] [CrossRef]
- Xiang, Q.B.; Liu, Y.L. An Illustrated Monograph of the Sweet Osmanthus Cultivars in China; Zhejiang Science and Technology Press: Hangzhou, China, 2008. [Google Scholar]
- Ke, D.S.; Wang, A.G.; Sun, G.C.; Dong, L.F. The effect of active oxygen on the activity of ACC synthase induced by exogenous IAA. Acta Bot. Sin. 2002, 44, 551–556. [Google Scholar]
- Geng, X.M.; Liu, J.; Lu, J.G.; Hu, F.R.; Okubo, H. Effects of cold storage and different pulsing treatments on postharvest quality of cut OT Lily ‘Mantissa’ flowers. J. Facul. Agric. Kyushu Univ. 2009, 54, 41–45. [Google Scholar]
- Dhindsa, R.A.; Plumb, D.P.; Thorpe, T.A. Leaf senescence, correlated with increased permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. Exp. Bot. 1981, 126, 93–101. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination offree proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Geng, X.M.; Liu, X.; Ji, M.; Hoffmann, W.A.; Grunden, A.; Xiang, Q.Y.J. Enhancing heat tolerance of the little dogwood cornus canadensis L. f. with introduction of a superoxide reductase gene from the hyperthermophilic archaeon pyrococcus furiosus. Front. Plant Sci. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Marcu, D.; Cristea, V.; Daraban, L. Dose-dependent effects of gamma radiation on lettuce (Lactuca sativa var. capitata) seedlings. Int. J. Radiat. Biol. 2013, 89, 219–223. [Google Scholar] [CrossRef]
- Geng, X.M.; Wang, L.G.; Li, N.; Yang, X.L. Study on the seed germination and seedling growth of Osmanthus fragrans under 6°Co-γ irradiation. J. Nuclear Agric. Sci. 2016, 30, 0216–0223, (In Chinese with English abstract). [Google Scholar]
- Shereen, A.; Ansari, R.; Mumtaz, S.; Bughio, H.R.; Mujtaba, S.M.; Shirazi, M.U.; Khan, M.A. Impact of gamma irradiation induced changes on growth and physiological responses of rice under saline conditions. Pakist. J. Bot. 2009, 41, 2487–2495. [Google Scholar]
- He, S.Z.; Han, Y.F.; Wang, Y.P.; Zhai, H.; Liu, Q.C. In vitro selection and identification of sweet potato (Ipomoea batatas (L.) Lam.) plants tolerant to NaCl. Plant Cell Tiss. Organ Cult. 2009, 96, 69–74. [Google Scholar] [CrossRef]
- Haruhiko, W.; Koshiba, T.; Matsui, T.; Satô, M. Involvement of peroxidase in differential sensitivity to γ-radiation in seedlings of two Nicotiana species. Plant Sci. 1998, 132, 109–119. [Google Scholar]
- Zaka, R.; Chenal, C.; Misset, M.T. Effects of low doses of short-term gamma irradiation on growth and development through two generations of Pisum sativum. Sci. Total Environ. 2004, 320, 121–129. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef]
- Helaly, M.N.M.; El-Hosieny, A.M.R. Effectiveness of gamma irradiated protoplasts on improving salt tolerance of lemon (Citrus limon L. Burm. f.). Am. J. Plant Physiol. 2011, 6, 190–208. [Google Scholar] [CrossRef]
- Qi, W.C.; Zhang, L.; Feng, W.S.; Xu, H.B.; Wang, L.; Jiao, Z. ROS and ABA Signaling Are Involved in the Growth Stimulation Induced by Low-Dose Gamma Irradiation in Arabidopsis Seedling. Appl. Biochem. Biotechnol. 2015, 175, 1490–1506. [Google Scholar] [CrossRef]
- Goh, E.J.; Kim, J.B.; Kim, W.J.; Ha, B.K.; Kim, S.H.; Kang, S.Y.; Seo, Y.W.; Kim, D.S. Physiological changes and anti-oxidative responses of Arabidopsis plants after acute and chronic γ-irradiation. Radiat. Environ. Biophys. 2014, 53, 677–693. [Google Scholar] [CrossRef]
- Aghaei, K.; Ehsanpour, A.A.; Komatsu, S. Potato responds to salt stress by increased activity of antioxidant enzymes. J. Integr. Plant Biol. 2009, 51, 1095–1103. [Google Scholar] [CrossRef]
- Nikam, A.A.; Devarumath, R.M.; Ahuja, A.; Babu, H.; Shitole, M.G.; Suprasanna, P. Radiation-induced in vitro mutagenesis system for salt tolerance and other agronomic characters in sugarcane (Saccharum officinarum L.). Crop J. 2015, 3, 46–56. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Protection of plasma membrane of onion epidermal cells by glycinebetaine and proline against NaCl stress. Plant Physiol. Biochem. 1998, 36, 767–772. [Google Scholar] [CrossRef]
- Rejeb, K.B.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Hossain, M.A.; Fujita, M. Evidence for a role of exogenous glycinebetaine and proline inantioxidant defense and methylglyoxal detoxification systems in mung bean seedlings under salt stress. Physiol. Mol. Biol. Plants 2010, 16, 19–29. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Salinity Damage Level | Description |
|---|---|---|
| 0 | No damage | No morphological damage symptoms of the whole plant |
| 1 | Mild | Less than 20% of the leaves have scorched margin and dehydration symptoms |
| 2 | Moderate | Nearly 50% of the leaves scorch, yellow with rust or wither |
| 3 | Severe | More than 50% of the leaves scorch, yellow with rust or wither |
| 4 | Very severe | More than 90% of the leaves scorch and wither or even the whole plant dies |
| NaCl Concentration (mmol/L) | Salt Injury Index | Salt Injury Rate (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 Gy | 50 Gy | 100 Gy | 150 Gy | 0 Gy | 50 Gy | 100 Gy | 150 Gy | |
| 0 | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a | 0.00 a |
| 20 | 0.08 ± 0.01 a | 0.07 ± 0.01 a | 0.07 ± 0.01 a | 0.00 ± 0.00 b | 63.33 a | 30.00 b | 26.67 c | 0.00 d |
| 40 | 0.55 ± 0.05 a | 0.37 ± 0.03 b | 0.36 ± 0.01 b | 0.28 ± 0.04 c | 100.0 a | 90.00 b | 70.00 c | 33.33 d |
| 60 | 0.70 ± 0.05 a | 0.56 ± 0.06 b | 0.52 ± 0.06 b | 0.40 ± 0.04 c | 100.0 a | 100.0 a | 96.67 b | 80.00 c |
| 80 | 0.90 ± 0.02 a | 0.74 ± 0.05 b | 0.67 ± 0.05 c | 0.54 ± 0.04 c | 100.0 a | 100.0 a | 100.0 a | 100.0 a |
| 100 | 0.90 ± 0.01 a | 0.89 ± 0.01 a | 0.78 ± 0.06 c | 0.85 ± 0.03 b | 100.0 a | 100.0 a | 100.0 a | 100.0 a |
| 120 | 0.98 ± 0.01 a | 0.98 ± 0.01 a | 0.95 ± 0.01 b | 0.97 ± 0.01 a | 100.0 a | 100.0 a | 100.0 a | 100.0 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, X.; Zhang, Y.; Wang, L.; Yang, X. Pretreatment with High-Dose Gamma Irradiation on Seeds Enhances the Tolerance of Sweet Osmanthus Seedlings to Salinity Stress. Forests 2019, 10, 406. https://doi.org/10.3390/f10050406
Geng X, Zhang Y, Wang L, Yang X. Pretreatment with High-Dose Gamma Irradiation on Seeds Enhances the Tolerance of Sweet Osmanthus Seedlings to Salinity Stress. Forests. 2019; 10(5):406. https://doi.org/10.3390/f10050406
Chicago/Turabian StyleGeng, Xingmin, Yuemiao Zhang, Lianggui Wang, and Xiulian Yang. 2019. "Pretreatment with High-Dose Gamma Irradiation on Seeds Enhances the Tolerance of Sweet Osmanthus Seedlings to Salinity Stress" Forests 10, no. 5: 406. https://doi.org/10.3390/f10050406
APA StyleGeng, X., Zhang, Y., Wang, L., & Yang, X. (2019). Pretreatment with High-Dose Gamma Irradiation on Seeds Enhances the Tolerance of Sweet Osmanthus Seedlings to Salinity Stress. Forests, 10(5), 406. https://doi.org/10.3390/f10050406