Analysis of Climate Change Impacts on Tree Species of the Eastern US: Results of DISTRIB-II Modeling


Abstract
1. Introduction
2. Materials and Methods
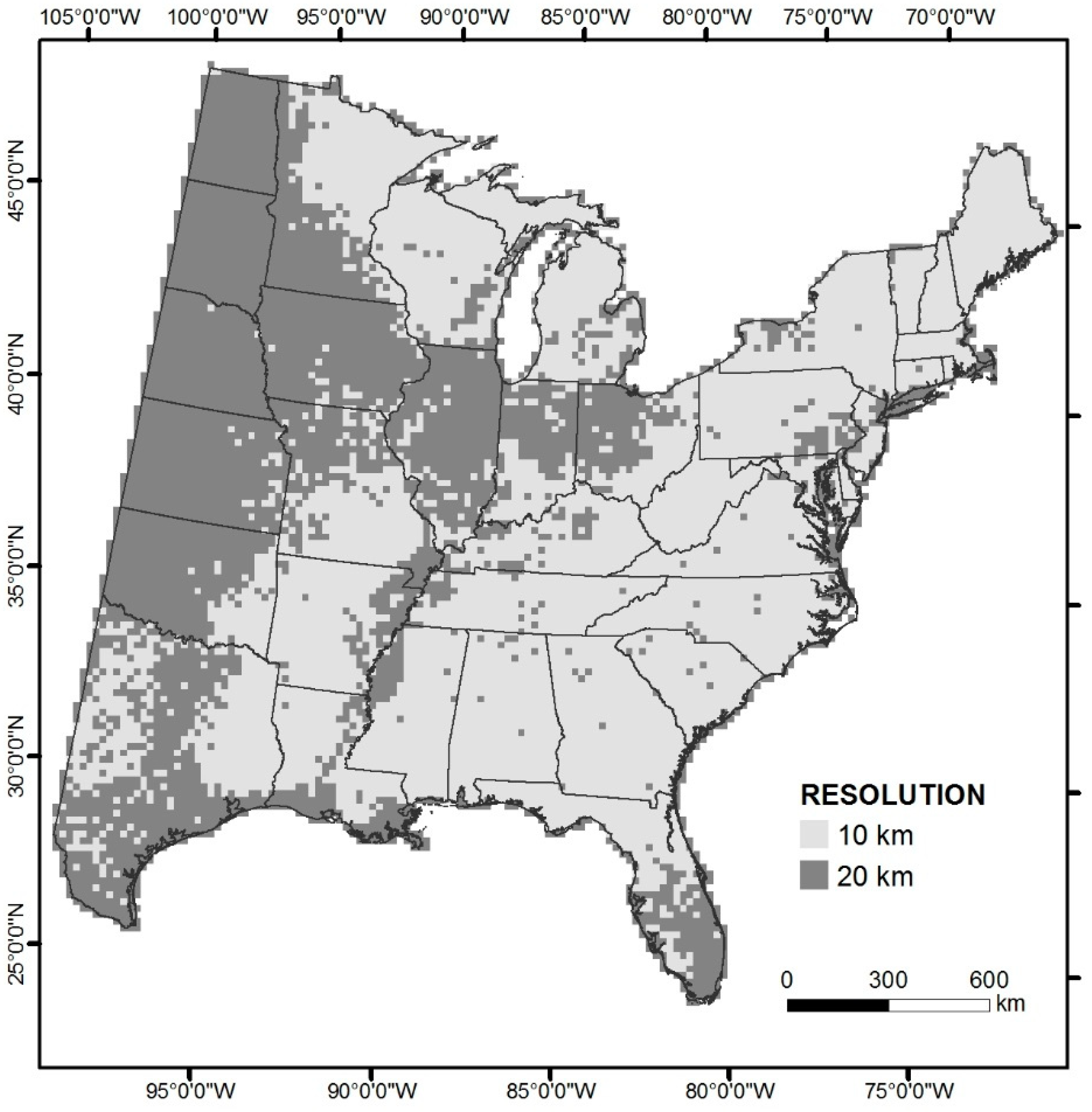
2.1. Data
2.2. Modeling
2.3. Model Reliability
2.4. Variable Importance
2.5. Area-Weighted Importance Values
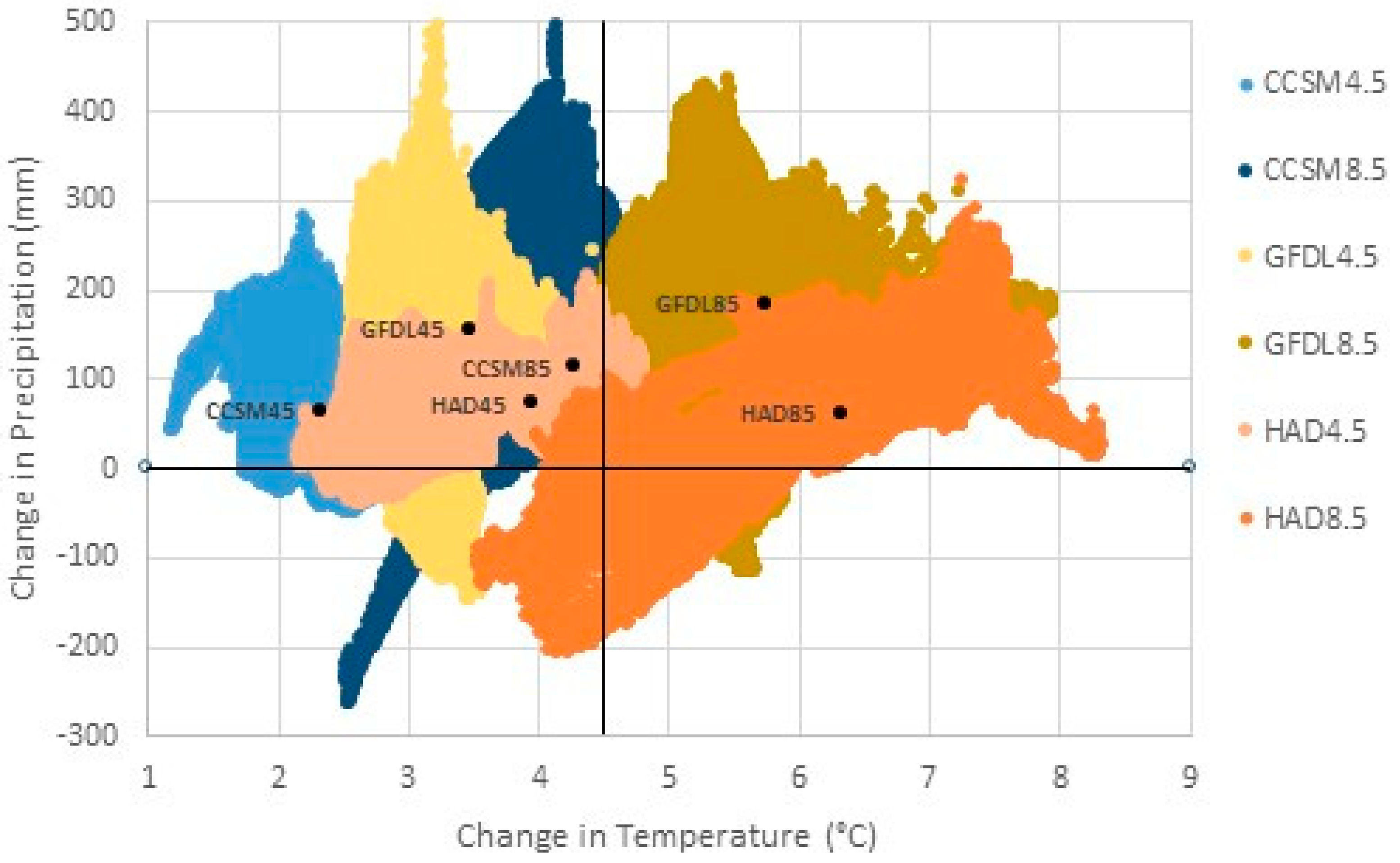
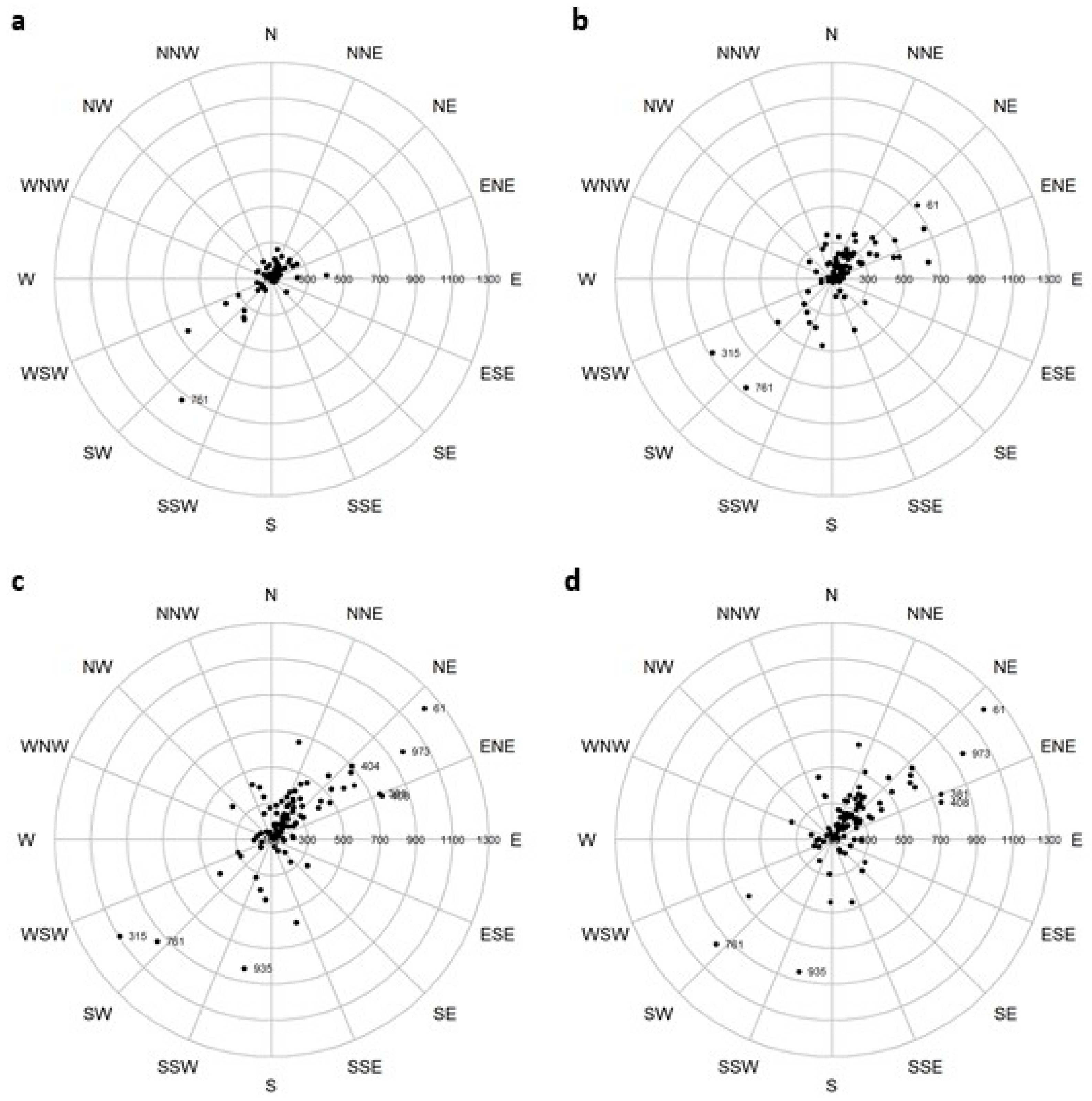
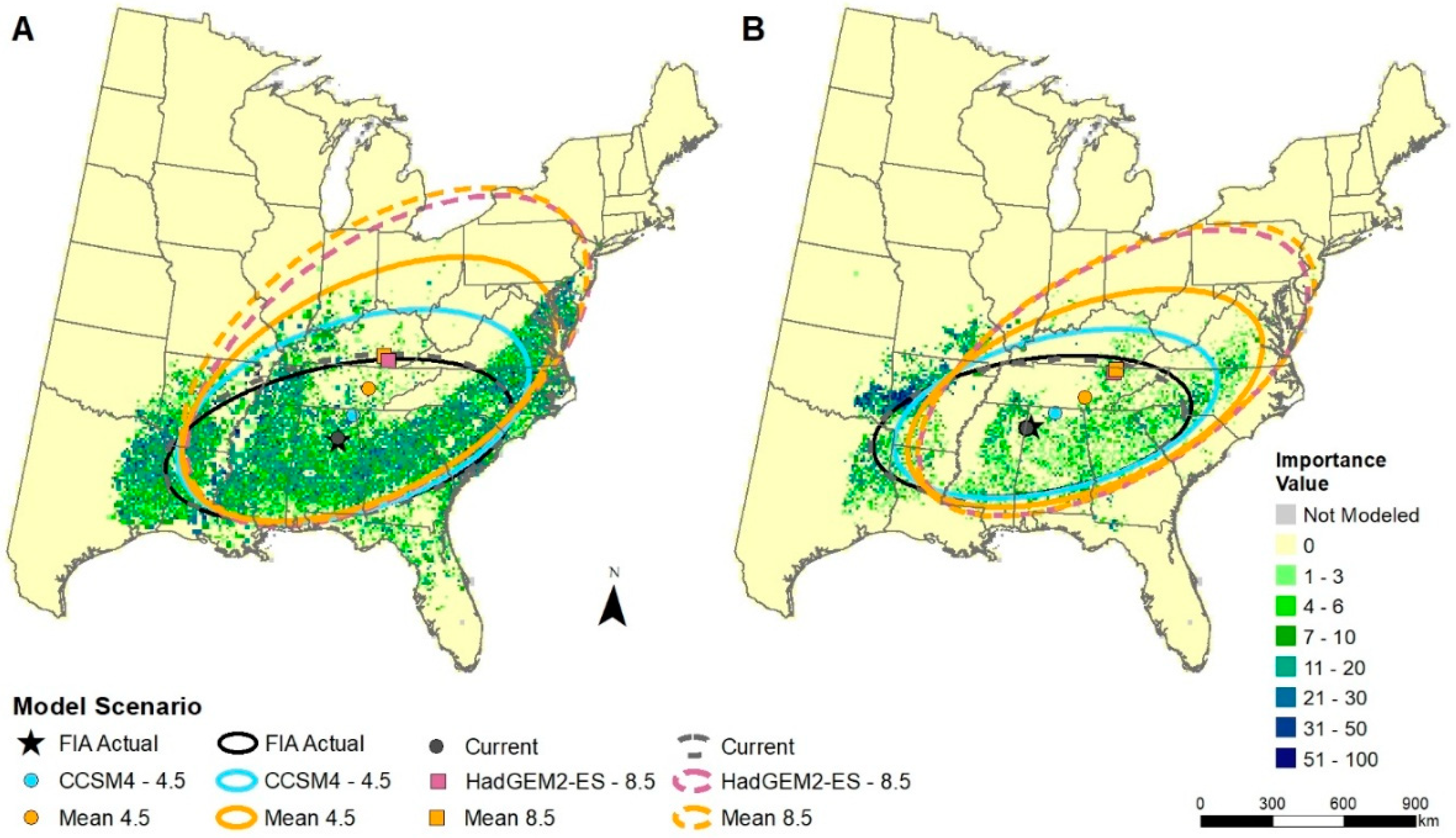
2.6. Changes in Mean Center of Spatial Data
2.7. Analysis of Dominants, Gainers, and Losers
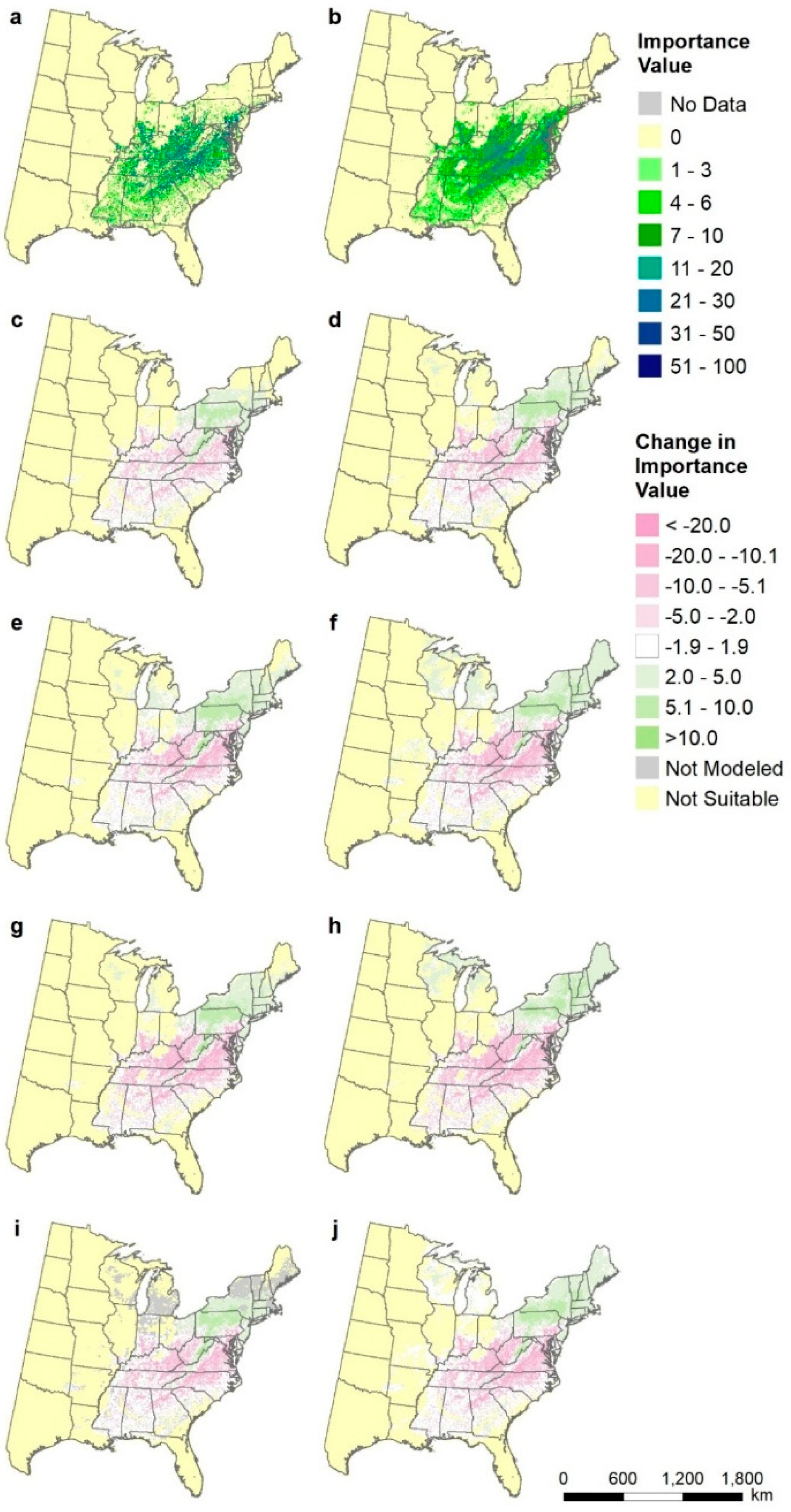
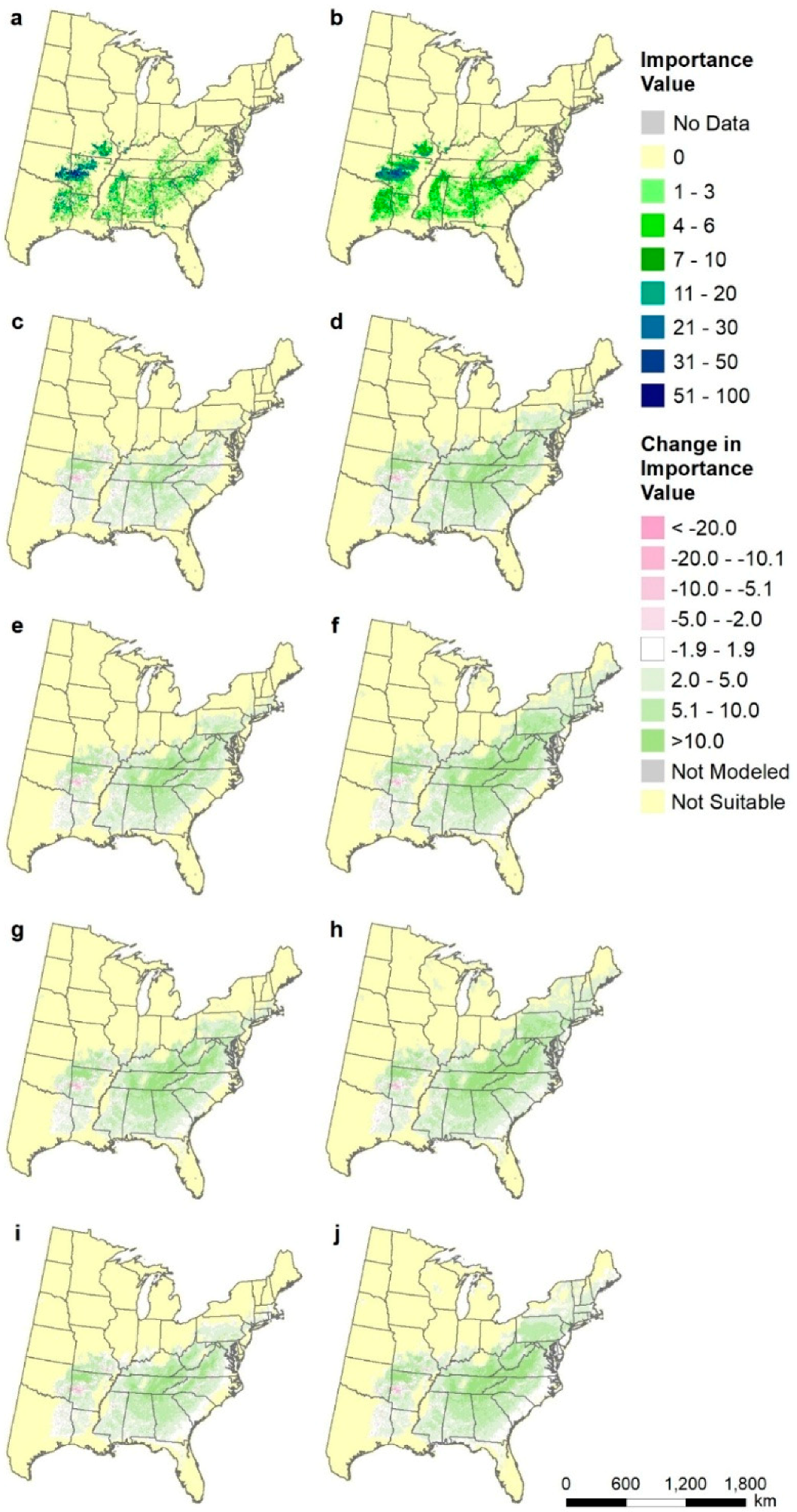
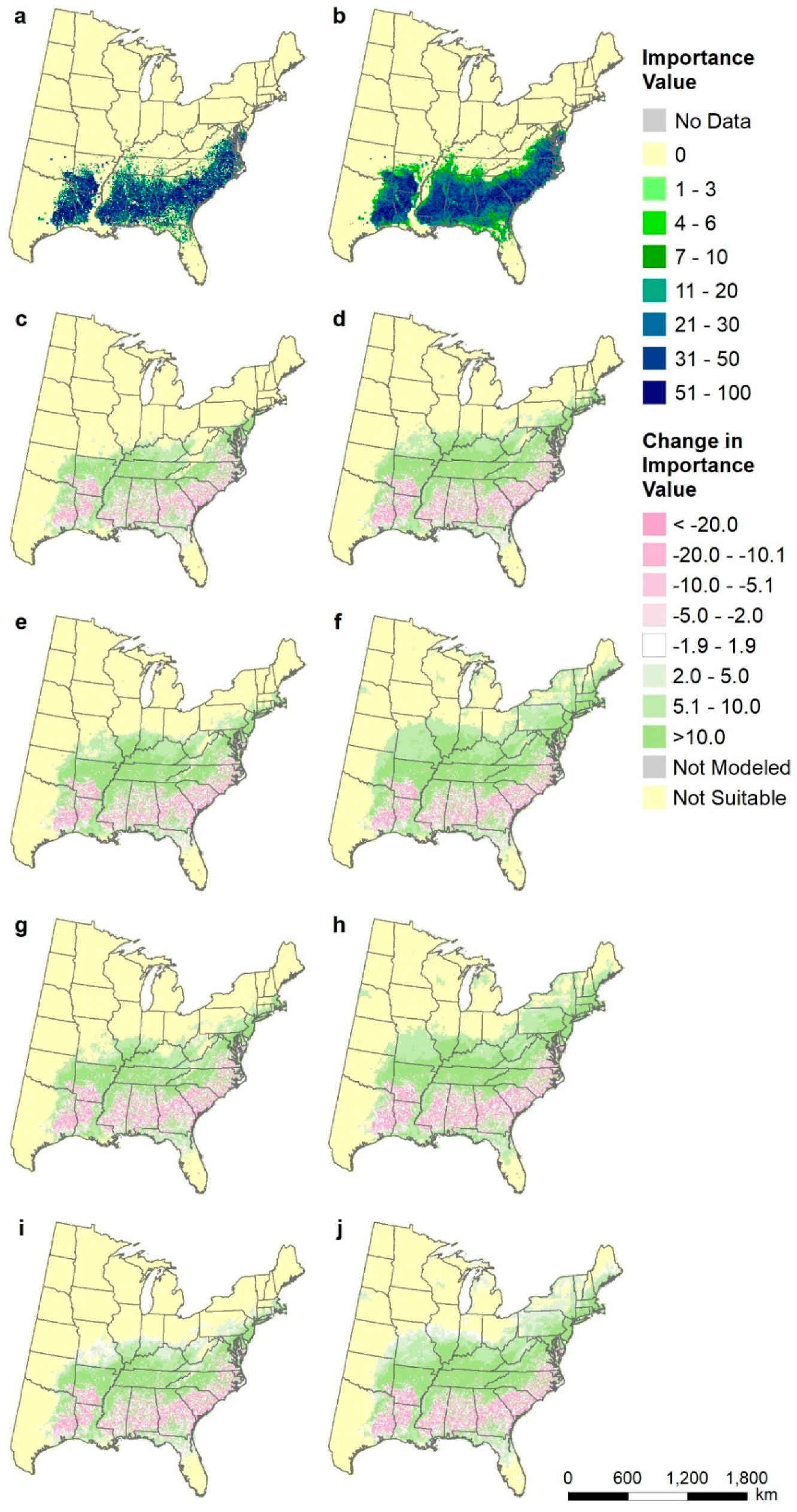
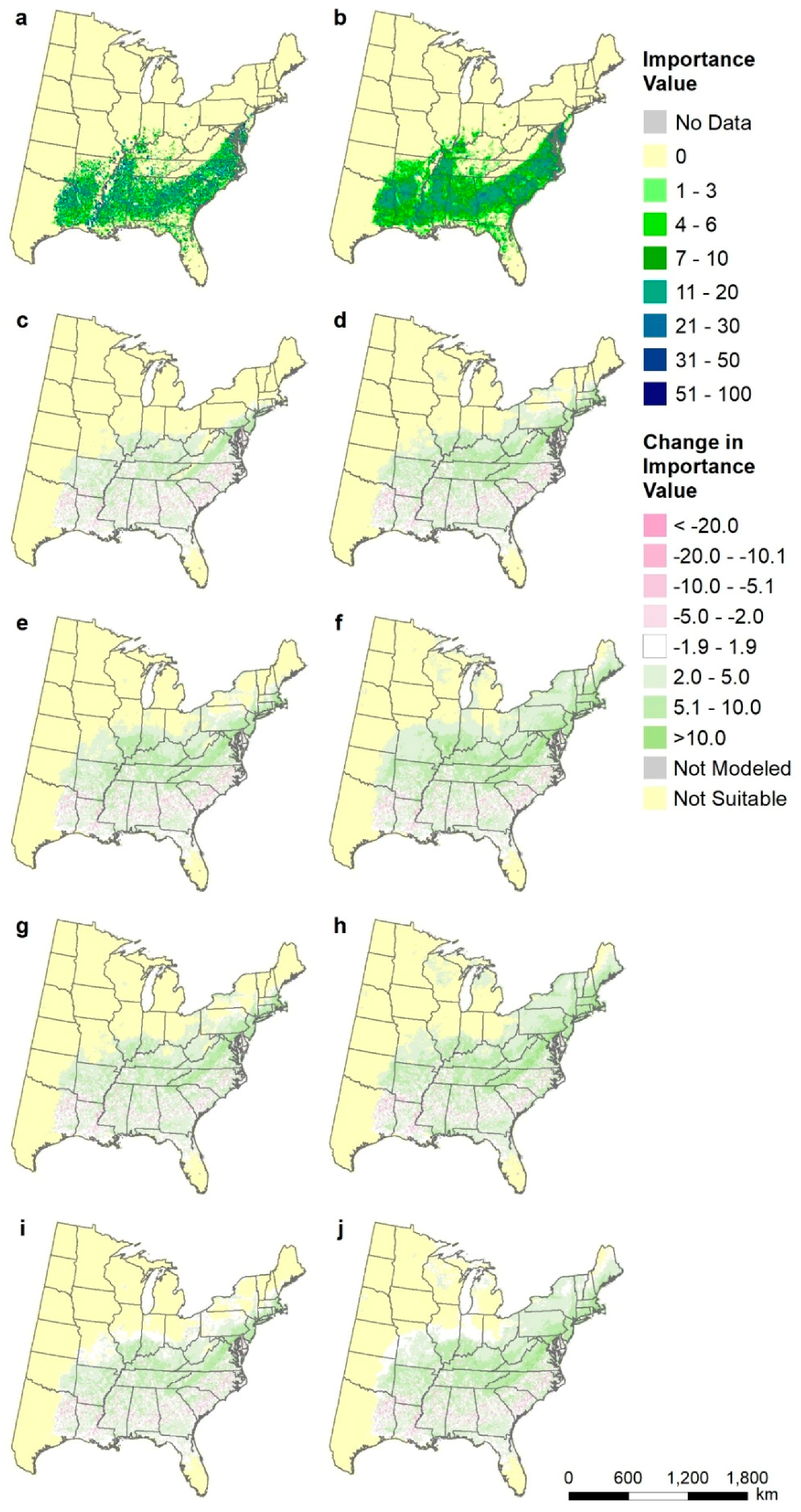
2.8. Species-Level Maps
2.9. Comparison to Earlier DISTRIB Models
2.10. Scope and Limitations
3. Results and Discussion
3.1. Model Reliability and Variable Importance
3.2. Potential Changes in Species Area-weighted Importance Values
3.3. Changes in Mean Center of Spatial Data
3.4. Analysis of Dominants, Gainers, and Losers by State and Region
3.5. Species-Level Maps
3.6. Comparison to Earlier DISTRIB Models
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Model Reliability | FIA sumIV | CCSM45 | GFDL85 | HAD85 | GCM45 | GCM85 | GCMave |
|---|---|---|---|---|---|---|---|---|
| Abies balsamea (L.) Mill. | High | 35,606 | 0.496 | 0.504 | 0.492 | 0.495 | 0.506 | 0.500 |
| Acer barbatus Michx. | Low | 1729 | 1.758 | 5.877 | 5.654 | 3.026 | 5.277 | 3.631 |
| Acer negundo L. | Low | 25,259 | 1.356 | 2.096 | 1.887 | 1.637 | 1.889 | 1.763 |
| Acer nigrum L. | Low | 678 | 0.409 | 0.292 | 0.282 | 0.364 | 0.299 | 0.340 |
| Acer pensylvanicum L. | Medium | 2549 | 0.752 | 0.565 | 0.547 | 0.672 | 0.580 | 0.653 |
| Acer rubrum L. | High | 165,591 | 1.030 | 0.915 | 0.871 | 0.993 | 0.920 | 0.956 |
| Acer saccharinum L. | Low | 14,872 | 1.804 | 3.044 | 2.445 | 2.309 | 2.630 | 2.203 |
| Acer saccharum Marshall | High | 88,010 | 0.945 | 0.906 | 0.819 | 0.932 | 0.880 | 0.906 |
| Acer spicatum Lam. | Low | 472 | 0.214 | 0.177 | 0.171 | 0.177 | 0.164 | 0.220 |
| Aesculus flava Sol. | Low | 1682 | 0.895 | 0.666 | 0.564 | 0.775 | 0.682 | 0.705 |
| Aesculus glabra Willd. | Low | 999 | 0.720 | 0.588 | 0.572 | 0.640 | 0.584 | 0.603 |
| Amelanchier spp. | Low | 2558 | 0.956 | 1.278 | 1.250 | 1.024 | 1.181 | 1.015 |
| Asimina triloba (L.) Dunal | Low | 738 | 0.488 | 0.519 | 0.518 | 0.495 | 0.497 | 0.481 |
| Betula alleghaniensis Britt. | High | 17,123 | 0.828 | 0.635 | 0.634 | 0.753 | 0.654 | 0.703 |
| Betula lenta L. | High | 13,368 | 1.029 | 0.806 | 0.802 | 0.966 | 0.844 | 0.909 |
| Betula nigra L. | Low | 3988 | 1.632 | 6.922 | 8.275 | 4.053 | 6.875 | 4.749 |
| Betula papyrifera Marshall | High | 21,096 | 0.940 | 0.833 | 0.779 | 0.918 | 0.834 | 0.876 |
| Betula populifolia Marsh. | Low | 1622 | 1.045 | 1.293 | 1.157 | 1.154 | 1.191 | 1.095 |
| Carpinus caroliniana Walter | Low | 9337 | 2.417 | 4.278 | 3.472 | 2.913 | 3.756 | 2.954 |
| Carya alba Sarg. | Medium | 22,233 | 1.831 | 2.998 | 3.185 | 2.267 | 2.920 | 2.593 |
| Carya aquatica (F.Michx.) Nutt. | Medium | 2624 | 1.278 | 1.619 | 1.699 | 1.380 | 1.641 | 1.393 |
| Carya cordiformis (Wangenh.) K.Koch | Low | 12,853 | 1.468 | 2.757 | 2.284 | 1.859 | 2.315 | 1.882 |
| Carya glabra Miller | Medium | 22,303 | 1.240 | 1.494 | 1.318 | 1.280 | 1.401 | 1.341 |
| Carya illinoinensis (Wangenh.) K.Koch | Low | 6698 | 2.613 | 13.336 | 15.386 | 5.479 | 12.017 | 7.580 |
| Carya laciniosa (Mill.) K.Koch | Low | 1009 | 1.051 | 1.767 | 1.080 | 1.241 | 1.340 | 1.177 |
| Carya ovate (Mill.) K.Koch | Medium | 19,547 | 1.149 | 1.356 | 1.170 | 1.281 | 1.262 | 1.271 |
| Carya texana Buckley (1861) | High | 9390 | 2.126 | 6.676 | 7.545 | 3.686 | 6.198 | 4.358 |
| Celtis laevigata Willdenow | Medium | 16,556 | 2.469 | 6.786 | 7.940 | 3.893 | 6.580 | 5.236 |
| Celtis occidentalis L. | Medium | 21,798 | 1.516 | 2.420 | 1.974 | 1.861 | 2.076 | 1.968 |
| Cercis Canadensis L. | Low | 4198 | 1.419 | 3.579 | 3.607 | 2.118 | 3.189 | 2.353 |
| Chamaecyparis thyoides (L.) B.S.P. | Low | 821 | 1.464 | 1.927 | 0.697 | 1.704 | 1.542 | 1.475 |
| Cornus florida L. | Medium | 10,589 | 1.545 | 1.868 | 1.631 | 1.607 | 1.765 | 1.554 |
| Diospyros virginiana L. | Low | 7074 | 1.981 | 8.115 | 8.345 | 4.039 | 7.027 | 4.814 |
| Fagus grandifolia Ehrh. | High | 35,486 | 1.118 | 1.291 | 0.996 | 1.163 | 1.193 | 1.178 |
| Fraxinus Americana L. | Medium | 42,548 | 1.314 | 1.633 | 1.547 | 1.455 | 1.559 | 1.507 |
| Fraxinus nigra Marshall | Medium | 13,276 | 1.114 | 0.934 | 0.890 | 1.080 | 0.955 | 0.997 |
| Fraxinus pennsylvanica Marshall | Low | 47,622 | 1.872 | 2.747 | 2.738 | 2.200 | 2.627 | 2.413 |
| Fraxinus quadrangulata Michx. | Low | 631 | 0.870 | 1.045 | 0.709 | 0.888 | 0.889 | 0.827 |
| Gleditsia triacanthos L. | Low | 11,912 | 1.317 | 3.096 | 3.083 | 1.907 | 2.739 | 2.078 |
| Gordonia lasianthus (L.) Ellis | Medium | 2418 | 1.666 | 1.812 | 1.567 | 1.651 | 1.712 | 1.549 |
| Halesia spp. | Low | 150 | 1.354 | 1.627 | 1.314 | 1.486 | 1.518 | 1.391 |
| Ilex opaca Aiton | Medium | 5391 | 1.829 | 2.456 | 2.165 | 2.018 | 2.304 | 1.956 |
| Juglans nigra L. | Low | 24,037 | 1.282 | 2.367 | 2.081 | 1.678 | 2.069 | 1.873 |
| Juniperus ashei J. Buchholz | High | 21,113 | 1.334 | 3.632 | 7.938 | 1.863 | 4.673 | 3.268 |
| Juniperus virginiana L. | Medium | 49,834 | 1.794 | 3.717 | 3.864 | 2.524 | 3.508 | 3.016 |
| Larix laricina (Du Roi) K. Koch | High | 12,797 | 1.198 | 1.170 | 1.161 | 1.238 | 1.215 | 1.176 |
| Liquidambar styraciflua L. | High | 91,344 | 1.342 | 1.816 | 1.802 | 1.479 | 1.730 | 1.604 |
| Liriodendron tulipifera L. | High | 63,276 | 0.895 | 0.821 | 0.645 | 0.807 | 0.766 | 0.786 |
| Maclura pomifera (Raf.) Schneid. | Medium | 11,988 | 1.167 | 2.920 | 2.610 | 1.755 | 2.408 | 1.883 |
| Magnolia acuminata L. | Low | 1734 | 1.058 | 1.106 | 1.066 | 1.039 | 1.086 | 1.008 |
| Magnolia fraseri Walter | Low | 651 | 1.542 | 1.667 | 1.610 | 1.540 | 1.653 | 1.465 |
| Magnolia grandiflora L. | Low | 1407 | 5.634 | 11.214 | 3.819 | 6.490 | 8.227 | 6.387 |
| Magnolia macrophylla Michx. | Low | 292 | 0.891 | 0.979 | 0.678 | 0.879 | 0.889 | 0.823 |
| Magnolia virginiana L. | Medium | 7875 | 2.622 | 3.793 | 1.945 | 2.798 | 3.129 | 2.658 |
| Morus rubra L. | Low | 8905 | 1.432 | 3.160 | 1.991 | 1.955 | 2.345 | 1.904 |
| Nyssa aquatic L. | Medium | 5014 | 1.284 | 1.856 | 1.215 | 1.377 | 1.587 | 1.360 |
| Nyssa biflora Walter | Medium | 14,558 | 1.716 | 2.140 | 1.813 | 1.816 | 1.991 | 1.760 |
| Nyssa sylvatica Marshall | Medium | 25,962 | 1.772 | 2.576 | 2.449 | 2.037 | 2.450 | 2.243 |
| Ostrya virginiana (Mill.) K.Koch | Low | 12,225 | 2.107 | 3.507 | 2.670 | 2.599 | 3.017 | 2.500 |
| Oxydendrum arboretum (L.) DC. | High | 11,224 | 1.020 | 0.911 | 0.701 | 0.885 | 0.860 | 0.877 |
| Persea borbonia (L.) Spreng. | Low | 2757 | 1.729 | 2.491 | 1.563 | 1.839 | 2.078 | 1.776 |
| Picea glauca (Moench) Voss | Medium | 7764 | 0.758 | 0.844 | 0.863 | 0.778 | 0.842 | 0.807 |
| Picea mariana (Mill.) Britton, Sterns & Poggenburg | High | 14,500 | 0.497 | 0.468 | 0.453 | 0.474 | 0.460 | 0.534 |
| Picea rubens Sarg. | High | 13,049 | 0.689 | 0.556 | 0.554 | 0.612 | 0.555 | 0.637 |
| Pinus banksiana Lamb. | Medium | 9280 | 0.719 | 0.635 | 0.642 | 0.689 | 0.661 | 0.702 |
| Pinus clausa (Chapm. ex Engelm.) Sarg. | Medium | 3740 | 1.223 | 1.573 | 0.850 | 1.413 | 1.275 | 1.263 |
| Pinus echinata Mill. | High | 32,601 | 2.147 | 3.848 | 3.985 | 2.796 | 3.665 | 3.231 |
| Pinus elliottii Engelm. | High | 57,498 | 1.450 | 2.093 | 1.622 | 1.577 | 1.858 | 1.718 |
| Pinus glabra Walter | Low | 1039 | 0.507 | 0.608 | 0.297 | 0.551 | 0.487 | 0.533 |
| Pinus palustris Mill. | Medium | 19,920 | 2.677 | 4.428 | 2.167 | 2.933 | 3.507 | 3.220 |
| Pinus pungens Lamb. | Low | 527 | 0.798 | 1.297 | 1.541 | 1.061 | 1.246 | 1.061 |
| Pinus resinosa Sol. ex Aiton | Medium | 17,992 | 0.945 | 0.967 | 0.937 | 0.998 | 0.973 | 0.985 |
| Pinus rigida Mill. | High | 5653 | 0.754 | 0.948 | 0.982 | 0.818 | 0.928 | 0.854 |
| Pinus serotina Michx. | Medium | 4019 | 1.471 | 1.569 | 1.203 | 1.335 | 1.448 | 1.298 |
| Pinus strobus L. | High | 42,529 | 1.137 | 0.882 | 0.827 | 1.090 | 0.921 | 1.006 |
| Pinus taeda L. | High | 271,571 | 1.178 | 1.404 | 1.285 | 1.212 | 1.324 | 1.268 |
| Pinus virginiana Mill. | High | 21,514 | 0.915 | 0.933 | 0.888 | 0.843 | 0.913 | 0.878 |
| Planera aquatic J.F.Gmel. | Low | 932 | 1.940 | 3.579 | 4.961 | 2.526 | 3.987 | 2.854 |
| Platanus occidentalis L. | Low | 11,992 | 1.963 | 4.698 | 4.200 | 2.803 | 4.053 | 3.030 |
| Populus balsamifera L. | Medium | 5854 | 0.431 | 0.314 | 0.423 | 0.380 | 0.385 | 0.445 |
| Populus deltoids W.Bartram ex Marshall | Low | 11,742 | 1.935 | 4.570 | 3.983 | 2.881 | 4.034 | 3.034 |
| Populus grandidentata Michaux | Medium | 12,814 | 1.218 | 1.034 | 0.974 | 1.235 | 1.074 | 1.094 |
| Populus tremuloides Michx. | High | 54,642 | 0.814 | 0.671 | 0.636 | 0.775 | 0.691 | 0.733 |
| Prunus pensylvanica L.f. | Low | 1734 | 0.354 | 0.132 | 0.151 | 0.284 | 0.164 | 0.259 |
| Prunus serotina Ehr. | Medium | 60,388 | 1.380 | 1.529 | 1.404 | 1.477 | 1.466 | 1.472 |
| Quercus alba L. | Medium | 87,470 | 1.225 | 1.364 | 1.300 | 1.317 | 1.330 | 1.324 |
| Quercus bicolor Willd. | Low | 2188 | 1.833 | 3.550 | 2.420 | 2.713 | 3.077 | 2.549 |
| Quercus coccinea Muenchh. | Medium | 17,739 | 1.128 | 1.221 | 1.137 | 1.168 | 1.208 | 1.188 |
| Quercus ellipsoidalis E.J.Hill | Medium | 6120 | 1.679 | 1.915 | 1.454 | 1.909 | 1.733 | 1.677 |
| Quercus falcate Michx. | Medium | 24,747 | 2.091 | 3.581 | 4.291 | 2.697 | 3.659 | 3.178 |
| Quercus imbricaria Michx. | Medium | 4356 | 0.680 | 0.802 | 0.516 | 0.772 | 0.635 | 0.704 |
| Quercus incana Bartram | Low | 624 | 1.050 | 3.781 | 9.120 | 2.678 | 5.097 | 3.404 |
| Quercus laevis Walter | Medium | 2731 | 1.205 | 1.590 | 1.009 | 1.258 | 1.327 | 1.211 |
| Quercus laurifolia Michx. | Medium | 15,945 | 1.947 | 2.739 | 1.708 | 2.046 | 2.263 | 1.974 |
| Quercus lyrata Walter | Medium | 4159 | 1.589 | 3.426 | 3.826 | 2.166 | 3.289 | 2.435 |
| Quercus macrocarpa Michx. | Medium | 19,711 | 1.705 | 1.941 | 1.592 | 1.859 | 1.816 | 1.837 |
| Quercus marilandica Muenchh. | Medium | 10,061 | 2.750 | 10.400 | 11.679 | 5.241 | 9.588 | 6.459 |
| Quercus michauxii Nutt. | Low | 2156 | 2.111 | 3.834 | 2.445 | 2.643 | 3.098 | 2.531 |
| Quercus muehlenbergii Engelm. | Medium | 6459 | 1.230 | 2.054 | 1.388 | 1.484 | 1.613 | 1.433 |
| Quercus nigra L. | High | 46,637 | 2.089 | 3.282 | 3.325 | 2.568 | 3.195 | 2.882 |
| Quercus pagoda Raf. | Medium | 7681 | 2.027 | 3.388 | 3.255 | 2.470 | 3.199 | 2.539 |
| Quercus palustris Münchh. | Low | 4434 | 0.974 | 2.846 | 1.892 | 1.347 | 1.970 | 1.510 |
| Quercus phellos L. | Low | 10,282 | 2.116 | 3.299 | 3.727 | 2.625 | 3.336 | 2.653 |
| Quercus prinus Willd. | High | 34,675 | 1.186 | 1.270 | 1.177 | 1.191 | 1.233 | 1.212 |
| Quercus rubra L. | Medium | 55,330 | 1.410 | 1.460 | 1.337 | 1.478 | 1.416 | 1.447 |
| Quercus shumardii Buckland | Low | 2523 | 1.127 | 5.424 | 4.020 | 2.050 | 3.989 | 2.664 |
| Quercus stellate Wangenh. | High | 58,812 | 2.169 | 4.671 | 5.252 | 3.077 | 4.504 | 3.791 |
| Quercus texana Buckley | Low | 2363 | 1.282 | 1.681 | 1.597 | 1.361 | 1.589 | 1.353 |
| Quercus velutina Lam. | High | 44,550 | 1.598 | 2.469 | 2.301 | 1.962 | 2.273 | 2.118 |
| Quercus virginiana Mill. | High | 25,609 | 2.450 | 7.093 | 10.064 | 3.789 | 7.552 | 5.670 |
| Robinia pseudoacacia L. | Low | 18,414 | 1.380 | 3.449 | 3.471 | 1.922 | 3.030 | 2.476 |
| Sabal palmetto (Walt.) Lodd. | Medium | 4949 | 2.046 | 3.867 | 2.557 | 2.337 | 3.123 | 2.473 |
| Salix nigra Marshall | Low | 13,959 | 1.445 | 4.936 | 5.616 | 2.458 | 4.558 | 3.075 |
| Sassafras albidum (Nutt.) Nees | Low | 14,728 | 1.477 | 2.377 | 2.631 | 1.871 | 2.346 | 1.908 |
| Sideroxylon lanuginosum ssp. lanuginosum | Low | 2109 | 7.622 | 49.696 | 89.291 | 23.994 | 56.912 | 34.731 |
| Sorbus Americana Marshall | Low | 147 | 0.397 | 0.265 | 0.292 | 0.334 | 0.282 | 0.330 |
| Taxodium ascendens Brongn. | Medium | 8177 | 2.305 | 4.612 | 2.979 | 2.467 | 3.457 | 2.646 |
| Taxodium distichum (L.) Rich. | Medium | 8683 | 2.324 | 3.622 | 2.692 | 2.599 | 3.131 | 2.558 |
| Thuja occidentalis L. | High | 20,487 | 0.808 | 0.854 | 0.975 | 0.818 | 0.926 | 0.872 |
| Tilia americana L. | Medium | 20,151 | 1.432 | 1.385 | 1.196 | 1.496 | 1.336 | 1.416 |
| Tsuga Canadensis (L.) Carrière | High | 27,300 | 1.035 | 0.724 | 0.707 | 0.926 | 0.765 | 0.846 |
| Ulmus alata Michx. | Medium | 21,303 | 2.273 | 4.353 | 5.729 | 3.137 | 4.597 | 3.867 |
| Ulmus americana L. | Medium | 55,590 | 1.475 | 2.316 | 2.401 | 1.799 | 2.218 | 2.008 |
| Ulmus crassifolia Nutt. | Medium | 8062 | 3.431 | 14.046 | 23.482 | 7.557 | 15.895 | 10.167 |
| Ulmus rubra Muhl. | Low | 13,179 | 1.359 | 2.543 | 2.828 | 1.774 | 2.453 | 1.907 |



References
- Mora, C.; Spirandelli, D.; Franklin, E.C.; Lynham, J.; Kantar, M.B.; Miles, W.; Smith, C.Z.; Freel, K.; Moy, J.; Louis, L.V.; et al. Broad threat to humanity from cumulative climate hazards intensified by greenhouse gas emissions. Nat. Clim. Chang. 2018. [Google Scholar] [CrossRef]
- Hsiang, S.; Kopp, R.; Jina, A.; Rising, J.; Delgado, M.; Mohan, S.; Rasmussen, D.J.; Muir-Wood, R.; Wilson, P.; Oppenheimer, M.; et al. Estimating economic damage from climate change in the United States. Science 2017, 356, 1362–1369. [Google Scholar] [CrossRef] [PubMed]
- IPCC Summary for Policymakers. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; World Meteorological Organization: Geneva, Switzerland, 2018; p. 32.
- Rogelj, J.; den Elzen, M.; Höhne, N.; Fransen, T.; Fekete, H.; Winkler, H.; Schaeffer, R.; Sha, F.; Riahi, K.; Meinshausen, M. Paris Agreement climate proposals need a boost to keep warming well below 2 °C. Nature 2016, 534, 631. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.; Price, J.; Graham, E.; Forstenhaeusler, N.; VanDerWal, J. The projected effect on insects, vertebrates, and plants of limiting global warming to 1.5 °C rather than 2 °C. Science 2018, 360, 791–795. [Google Scholar] [CrossRef]
- Swanston, C.W.; Janowiak, M.; Brandt, L.; Butler, P.; Handler, S.D.; Shannon, P.D.; Lewis, A.D.; Hall, K.R.; Fahey, R.T.; Scott, L.; et al. Forest Adaptation Resources: Climate Change Tools and Approaches for Land Managers, 2nd ed.; Gen. Tech. Rep. NRS-GTR-87-2; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2016; p. 161.
- Millar, C.; Stephenson, N.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Nagel, L.M.; Palik, B.J.; Battaglia, M.A.; D’Amato, A.W.; Guldin, J.M.; Swanston, C.W.; Janowiak, M.K.; Powers, M.P.; Joyce, L.A.; Millar, C.I.; et al. Adaptive Silviculture for Climate Change: A National Experiment in Manager-Scientist Partnerships to Apply an Adaptation Framework. J. For. 2017, 115, 167–178. [Google Scholar] [CrossRef]
- Halofsky, J.E.; Andrews-Key, S.A.; Edwards, J.E.; Johnston, M.H.; Nelson, H.W.; Peterson, D.L.; Schmitt, K.M.; Swanston, C.W.; Williamson, T.B. Adapting forest management to climate change: The state of science and applications in Canada and the United States. For. Ecol. Manag. 2018, 421, 84–97. [Google Scholar] [CrossRef]
- Iverson, L.; McKenzie, D. Tree-species range shifts in a changing climate—Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.C. Climate-related range shifts—A global multidimensional synthesis and new research directions. Ecography 2014, 38, 15–28. [Google Scholar] [CrossRef]
- Vanderwel, M.C.; Rozendaal, D.M.A.; Evans, M.E.K. Predicting the abundance of forest types across the eastern U.S. through inverse modelling of tree demography. Ecol. Appl. 2017, 27, 2128–2141. [Google Scholar] [CrossRef]
- Clark, J.S.; Nemergut, D.; Seyednasrollah, B.; Turner, P.J.; Zhang, S. Generalized joint attribute modeling for biodiversity analysis: Median-zero, multivariate, multifarious data. Ecol. Monogr. 2016, 87, 34–56. [Google Scholar] [CrossRef]
- DeHayes, D.H.; Jacobson, G.L.; Schaberg, P.G.; Bongarten, B.; Iverson, L.R.; Dieffenbacker-Krall, A. Forest responses to changing climate: Lessons from the past and uncertainty for the future. In Responses of Northern Forests to Environmental Change; Mickler, R.A., Birdsey, R.A., Hom, J.L., Eds.; Springer, Ecological Studies Series: New York, NY, USA, 2000; pp. 495–540. [Google Scholar]
- Delcourt, P.A.; Delcourt, H.R. Late-Quaternary dynamics of temperate forests—Applications of paleoecology to issues of global environmental-change. Quat. Sci. Rev. 1987, 6, 129–146. [Google Scholar] [CrossRef]
- Koo, K.A.; Madden, M.; Patten, B.C. Projection of red spruce (Picea rubens Sargent) habitat suitability and distribution in the Southern Appalachian Mountains, USA. Ecol. Model. 2014, 293, 91–101. [Google Scholar] [CrossRef]
- Prasad, A.M.; Iverson, L.R.; Matthews, S.N.; Peters, M.P. A multistage decision support framework to guide tree species management under climate change via habitat suitability and colonization models, and a knowledge-based scoring system. Landsc. Ecol. 2016, 31, 2187–2204. [Google Scholar] [CrossRef]
- Iverson, L.; Prasad, A.M.; Matthews, S.; Peters, M. Lessons learned while integrating habitat, dispersal, disturbance, and life-history traits into species habitat models under climate change. Ecosystems 2011, 14, 1005–1020. [Google Scholar] [CrossRef]
- Morin, X.; Thuiller, W. Comparing niche- and process-based models to reduce prediction uncertainty in species range shifts under climate change. Ecology 2009, 90, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Iverson, L.R.; Thompson, F.R.; Matthews, S.; Peters, M.; Prasad, A.; Dijak, W.D.; Fraser, J.; Wang, W.J.; Hanberry, B.; He, H.; et al. Multi-model comparison on the effects of climate change on tree species in the eastern U.S.: Results from an enhanced niche model and process-based ecosystem and landscape models. Landsc. Ecol. 2017, 32, 1327–1346. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Kullman, L. Rapid recent range-margin rise of tree and shrub species in the Swedish Scandes. J. Ecol. 2002, 90, 68–77. [Google Scholar] [CrossRef]
- Gamache, I.; Payette, S. Latitudinal response of subarctic tree lines to recent climate change in eastern Canada. J. Biogeogr. 2005, 32, 849–862. [Google Scholar] [CrossRef]
- Holzinger, B.; Hulber, K.; Camenisch, M.; Grabherr, G. Changes in plant species richness over the last century in the eastern Swiss Alps: Elevational gradient, bedrock effects and migration rates. Plant Ecol. 2008, 195, 179–196. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Hillyer, R.; Silman, M.R. Changes in species interactions across a 2.5 km elevation gradient: Effects on plant migration in response to climate change. Glob. Chang. Biol. 2010, 16, 3205–3214. [Google Scholar] [CrossRef]
- Felde, V.A.; Kapfer, J.; Grytnes, J.-A. Upward shift in elevational plant species ranges in Sikkilsdalen, central Norway. Ecography 2012, 35, 922–932. [Google Scholar] [CrossRef]
- Freeman, B.G.; Lee-Yaw, J.A.; Sunday, J.M.; Hargreaves, A.L. Expanding, shifting and shrinking: The impact of global warming on species’ elevational distributions. Glob. Ecol. Biogeogr. 2018, 27, 1268–1276. [Google Scholar] [CrossRef]
- Foster, J.R.; D’Amato, A.W. Montane forest ecotones moved downslope in northeastern US in spite of warming between 1984 and 2011. Glob. Chang. Biol. 2015, 21, 4497–4507. [Google Scholar] [CrossRef] [PubMed]
- Beckage, B.; Osborne, B.; Gavin, D.G.; Pucko, C.; Siccama, T.; Perkins, T. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc. Natl. Acad. Sci. USA 2008, 105, 4197–4202. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M. Invasive forest defoliator contributes to the impending downward trend of oak dominance in eastern North America. For. Int. J. For. Res. 2015, 89, 284–289. [Google Scholar] [CrossRef]
- Dolan, B.K.J. Forest regeneration following emerald ash borer (Agrilus planipennis Fairemaire) enhances mesophication in eastern hardwood forests. Forests 2018, 9, 353. [Google Scholar] [CrossRef]
- Wang, W.; He, H.; Thompson, F., III; Fraser, J.; Hanberry, B.; Dijak, W. The importance of succession, harvest, and climate change in determining future forest composition changes in U.S. Central Hardwood Forests. Ecosphere 2015, 6, art277. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Hansen, M.H. Latitudinal range shifts of tree species in the United States across multi-decadal time scales. Basic Appl. Ecol. 2015, 16, 231–238. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Périé, C.; de Blois, S. Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 2014, 5, 1–33. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Périé, C.; de Blois, S. Divergent responses to climate change and disturbance drive recruitment patterns underlying latitudinal shifts of tree species. J. Ecol. 2019. [Google Scholar] [CrossRef]
- Sittaro, F.; Paquette, A.; Messier, C.; Nock Charles, A. Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Glob. Chang. Biol. 2017, 23, 3292–3301. [Google Scholar] [CrossRef]
- Woodall, C.W.; Westfall, J.A.; D’Amato, A.W.; Foster, J.R.; Walters, B.F. Decadal changes in tree range stability across forests of the eastern U.S. For. Ecol. Manag. 2018, 429, 503–510. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M. Predicting abundance of 80 tree species following climate change in the eastern United States. Ecol. Monogr. 1998, 68, 465–485. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Hale, B.J.; Sutherland, E.K. Atlas of Current and Potential Future Distributions of Common Trees of the Eastern United States; General Technical Report NE-265; Northeastern Research Station, USDA Forest Service: Radnor, PA, USA, 1999; p. 245.
- Prasad, A.M.; Iverson, L.R.; Liaw, A. Newer classification and regression tree techniques: Bagging and random forests for ecological prediction. Ecosystems 2006, 9, 181–199. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For. Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N. Modeling potential climate change impacts on the trees of the northeastern United States. Mitig. Adapt. Strateg. Glob. Chang. 2008, 13, 487–516. [Google Scholar] [CrossRef]
- Butler-Leopold, P.R.; Iverson, L.R.; Thompson, F.R., III; Brandt, L.A.; Handler, S.D.; Janowiak, M.K.; Shannon, P.D.; Swanston, C.W.; Bearer, S.; Bryan, A.M.; et al. Mid-Atlantic Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Mid-Atlantic Climate Change Response Framework Project; Gen. Tech. Rep. NRS-181; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2018; p. 365.
- Butler, P.R.; Iverson, L.; Thompson, F.R.; Brandt, L.; Handler, S.; Janowiak, M.; Shannon, P.D.; Swanston, C.; Karriker, K.; Bartig, J.; et al. Central Appalachians Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Central Appalachians Climate Change Response Framework Project; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2015; p. 310.
- Brandt, L.; He, H.; Iverson, L.; Thompson, F.; Butler, P.; Handler, S.; Janowiak, M.; Swanston, C.; Albrecht, M.; Blume-Weaver, R.; et al. Central Hardwoods Ecosystem Vulnerability Assessment and Synthesis: A Report from the Central Hardwoods Climate Change Response Framework Project; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 254.
- Handler, S.; Duveneck, M.; Iverson, L.; Peters, E.; Scheller, R.; Wythers, K.; Brandt, L.; Butler, P.; Janowiak, M.; Swanston, C.; et al. Minnesota Forest Ecosystem Vulnerability Assessment and Synthesis: A report from the Northwoods Climate Change Response Framework; Gen. Tech. Rep. NRS-133; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014.
- Handler, S.; Duveneck, M.J.; Iverson, L.; Peters, E.; Scheller, R.; Wythers, K.; Brandt, L.; Butler, P.; Janowiak, M.; Swanston, C.; et al. Michigan Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Northwoods Climate Change Response Framework; Gen. Tech. Rep. NRS-129; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014.
- Janowiak, M.K.; Iverson, L.R.; Mladenoff, D.J.; Peters, E.; Wythers, K.R.; Xi, W.; Brandt, L.A.; Butler, P.R.; Handler, S.D.; Shannon, P.D.; et al. Forest Ecosystem Vulnerability Assessment and Synthesis for Northern Wisconsin and Western Upper Michigan: A Report from the Northwoods Climate Change Response Framework Project; Gen. Tech. Rep. NRS-136; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 247.
- Janowiak, M.K.; D’Amato, A.W.; Swanston, C.W.; Iverson, L.; Thompson, F., III; Dijak, W.; Matthews, S.; Peters, M.; Prasad, A.; Fraser, J.; et al. New England and Northern New York Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the New England Climate Change Response Framework Project; Gen. Tech. Rep. NRS-173; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2018; p. 234.
- Brandt, L.A.; Derby Lewis, A.; Scott, L.; Darling, L.; Fahey, R.T.; Iverson, L.; Nowak, D.J.; Bodine, A.R.; Bell, A.; Still, S.; et al. Chicago Wilderness Region Urban Forest Vulnerability Assessment and Synthesis: A Report from the Urban Forestry Climate Change Response Framework Chicago Wilderness Pilot Project; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2017; p. 142.
- Soil_Survey_Staff. Gridded Soil Survey Geographic (gSSURGO) Database for Ohio; United States Department of Agriculture, Natural Resources Conservation Service. Available online: https://gdg.sc.egov.usda.gov/ (accessed on 24 May 2016).
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int. J. Climatol. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- Thrasher, B. Downscaled Climate Projections Suitable for Resource Management. Eos 2013, 94, 321–323. [Google Scholar] [CrossRef]
- Maurer, E.P.; Hidalgo, H.G. Utility of daily vs. monthly large-scale climate data: An intercomparison of two statistical downscaling methods. Hydrol. Earth Syst. Sci. 2008, 12, 551–563. [Google Scholar] [CrossRef]
- Monahan, W.B.; Cook, T.; Melton, F.; Connor, J.; Bobowski, B. Forecasting Distributional Responses of Limber Pine to Climate Change at Management-Relevant Scales in Rocky Mountain National Park. PLoS ONE 2013, 8, e83163. [Google Scholar] [CrossRef]
- Moss, R.; Babiker, W.; Brinkman, S.; Calvo, E.; Carter, T.; Edmonds, J.; Elgizouli, I.; Emori, S.; Erda, L.; Hibbard, K. Towards New Scenarios for the Analysis of Emissions, Climate Change, Impacts, and Response Strategies; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2008; p. 25. ISBN 978-92-9169-124-1. [Google Scholar]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Donner, L.J.; Wyman, B.L.; Hemler, R.S.; Horowitz, L.W.; Ming, Y.; Zhao, M.; Golaz, J.-C.; Ginoux, P.; Lin, S.J.; Schwarzkopf, M.D.; et al. The Dynamical Core, Physical Parameterizations, and Basic Simulation Characteristics of the Atmospheric Component AM3 of the GFDL Global Coupled Model CM3. J. Clim. 2011, 24, 3484–3519. [Google Scholar] [CrossRef]
- Collins, W.J.; Bellouin, N.; Doutriaux-Boucher, M.; Gedney, N.; Halloran, P.; Hinton, T.; Hughes, J.; Jones, C.D.; Joshi, M.; Liddicoat, S.; et al. Development and evaluation of an Earth-System model—HadGEM2. Geosci. Model Dev. 2011, 4, 1051–1075. [Google Scholar] [CrossRef]
- Jones, C.D.; Hughes, J.K.; Bellouin, N.; Hardiman, S.C.; Jones, G.S.; Knight, J.; Liddicoat, S.; O’Connor, F.M.; Andres, R.J.; Bell, C.; et al. The HadGEM2-ES implementation of CMIP5 centennial simulations. Geosci. Model Dev. 2011, 4, 543–570. [Google Scholar] [CrossRef]
- Peters, M.P.; Iverson, L. Projected Drought for the Conterminous United States in the 21st Century. In Drought Impacts on U.S. Forests and Rangelands: Translating Science into Management Responses; Vose, J.M., Peterson, D.L., Luce, C.H., Patel-Weynand, T., Eds.; Gen. Tech. Rep. WO-GTR-xx; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, in press.
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45. [Google Scholar] [CrossRef]
- Forsythe, W.C.; Rykiel, E.J.; Stahl, R.S.; Wu, H.-I.; Schoolfield, R.M. A model comparison for daylength as a function of latitude and day of year. Ecol. Model. 1995, 80, 87–95. [Google Scholar] [CrossRef]
- Schaetzl, R.J.; Krist, F.J.J.; Miller, B.A. A taxonomically based ordinal estimate of soil productivity for landscape-scale analyses. Soil Sci. Soc. Am. J. 2012, 75, 1–8. [Google Scholar] [CrossRef]
- Koch, F.H.; Smith, W.D.; Coulston, J.W. An Improved Method for Standardized Mapping of Drought Conditions. In Forest Health Monitoring: National Status, Trends, and Analysis 2010; Gen. Tech. Rep. SRS-GTR-176; Potter, K.M., Conkling, B.L., Eds.; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2013; p. 164. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- R_Development_Core_Team. R: A Language and Environment for Statistical Computing. Version 3.1.1; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Fry, J.A.; Xian, G.; Jin, S.; Dewitz, J.A.; Homer, C.G.; LIMIN, Y.; Barnes, C.A.; Herold, N.D.; Wickham, J.D. Completion of the 2006 national land cover database for the conterminous United States. Photogramm. Eng. Remote Sens. 2011, 77, 858–864. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Roy, M.-H.; Larocque, D. Robustness of random forests for regression. J. Nonparametric Stat. 2012, 24, 993–1006. [Google Scholar] [CrossRef]
- Hagen-Zanker, A. Comparing Continuous Valued Raster Data: A Cross Disciplinary Literature Scan; Research Institute for Knowledge Systems: Bilthoven, The Netherlands, 2006. [Google Scholar]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A. How fast and far might tree species migrate under climate change in the eastern United States? Glob. Ecol. Biogeogr. 2004, 13, 209–219. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A.M. Potential colonization of new available tree species habitat under climate change: An analysis for five eastern US species. Landsc. Ecol. 2004, 19, 787–799. [Google Scholar] [CrossRef]
- Prasad, A.M.; Gardiner, J.; Iverson, L.; Matthews, S.; Peters, M. Exploring tree species colonization potentials using a spatially explicit simulation model: Implications for four oaks under climate change. Glob. Chang. Biol. 2013, 19, 2196–2208. [Google Scholar] [CrossRef]
- Clark, J.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. Impacts of increasing drought on forest dynamics, structure, diversity, and management. In Effects of Drought on Forests and Rangelands in the United States: A Comprehensive Science Synthesis; Vose, J., Clark, J., Luce, C., Patel-Weynand, T., Eds.; Gen. Tech. Rep. WO-93b; U.S. Department of Agriculture, Forest Service, Washington Office: Washington, DC, USA, 2016; pp. 59–96. [Google Scholar]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017, 3, e1603055. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.N.; Iverson, L.; Peters, M.; Prasad, A.M. Assessing Potential Climate Change Pressures Throughout This Century across the Conterminous United States: Mapping Plant Hardiness Zones, Heat Zones, Growing Degree Days, and Cumulative Drought Severity Throughout this Century Research Map NRS-9; U.S. Department of Agriculture, Forest Service. Northern Research Station: Newtown Square, PA, USA, 2018; p. 31.
- Iverson, L.; Matthews, S.; Prasad, A.; Peters, M.; Yohe, G. Development of risk matrices for evaluating climatic change responses of forested habitats. Clim. Chang. 2012, 114, 231–243. [Google Scholar] [CrossRef]
- Matthews, S.N.; Iverson, L.R.; Prasad, A.M.; Peters, M.P.; Rodewald, P.G. Modifying climate change habitat models using tree species-specific assessments of model uncertainty and life history-factors. For. Ecol. Manag. 2011, 262, 1460–1472. [Google Scholar] [CrossRef]
- Box, G.; Draper, N.R. Empirical Model-Building and Response Surfaces; Wiley: New York, NY, USA, 1987. [Google Scholar]





| Variable | CCSM4 | GFDL CM3 | HadGEM2-ES | ||||
|---|---|---|---|---|---|---|---|
| Current | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | |
| PANN (mm) | 1048 | 1113 | 1162 | 1203 | 1233 | 1121 | 1110 |
| Pgrow (mm) | 509 | 527 | 519 | 573 | 585 | 504 | 457 |
| TANN (°C) | 12.7 | 15.2 | 17.1 | 16.3 | 18.6 | 16.8 | 19.2 |
| Tgrow (°C) | 21.6 | 24.0 | 26.3 | 26.0 | 28.4 | 25.7 | 28.4 |
| TSUMavg (°C) | 24.8 | 26.6 | 28.1 | 28.1 | 29.7 | 28.0 | 29.9 |
| TWINavg (°C) | −0.9 | 1.2 | 2.1 | 1.6 | 2.0 | 1.3 | 3.3 |
| TMin (°C) | −11.3 | −8.6 | −6.5 | −7.2 | −5.7 | −7.4 | −4.0 |
| TMax (°C) | 33.8 | 35.9 | 39.3 | 39.1 | 42.5 | 38.5 | 41.9 |
| Category | Variable | Description | Native Resolution |
|---|---|---|---|
| Climate 1 | Annual precipitation | Mean 30-year (1981–2010) monthly precipitation (mm). | 800 m |
| May–Sept. precipitation | Mean 30-year (1981–2010) monthly precipitation for May–September (mm). | ||
| Annual mean temperature | Mean 30-year (1981–2010) monthly temperature (°C). | ||
| May–Sept. mean temperature | Mean 30-year (1981–2010) monthly temperature for May–September (°C). | ||
| Mean temperature of coldest month | Mean 30-year (1981–2010) monthly temperature of coldest month (°C). | ||
| Mean temperature of warmest month | Mean 30-year (1981–2010) monthly temperature of warmest month (°C). | ||
| Aridity Index | A conditional ratio of precipitation and Thornthwaite potential evapotranspiration (see [66]) | 10 and 20 km | |
| Elevation [63] | Minimum | Minimum value | 90 m |
| Mean | Mean value | ||
| Maximum | Maximum value | ||
| Median | Median value | ||
| Range | Range between minimum and maximum values | ||
| Standard deviation | Amount of deviance among elevation | ||
| Coefficient of variation | The CV of elevation | ||
| Solar [64] | Day length coefficient of variation | The CV of 12 monthly day lengths derived from the latitude of grid cells. | 10 and 20 km |
| Soil [52] | Available water capacity (cm) | The quantity of water that the soil is capable of storing for use by plants | 30 m |
| Available water supply (cm) | The total volume of water that should be available to plants when the soil, inclusive of rock fragments, is at field capacity | ||
| Bulk density (g/cm3) | The ovendry weight of the soil material <2 mm in size per unit volume of soil at water tension of 1/3 bar | ||
| Calcium carbonate | The percent of carbonates, by weight, in the fraction of the soil <2 mm in size | ||
| Cation-exchange capacity | The total amount of extractable cations that can be held by the soil, expressed in terms of milliequivalents per 100 grams of soil at neutrality (pH 7.0) or at some other stated pH | ||
| Depth to water table (cm) | Depth to a saturated zone in the soil | ||
| Permeability (cm/h) | Saturated hydraulic conductivity or the ease with which pores in a saturated soil transmit water | ||
| Erosion K factor | The susceptibility of a soil to sheet and rill erosion by water estimated by the percentage of silt, sand, and organic matter and on soil structure and saturated hydraulic conductivity | ||
| Erosion T factor (tons/acre/year) | An estimate of the maximum average annual rate of soil erosion by wind and/or water that can occur without affecting crop productivity over a sustained period | ||
| Percent clay | Mineral soil particles that are <0.002 mm in diameter | ||
| Percent sand | Mineral soil particles that are 0.05 mm to 2 mm in diameter | ||
| Percent silt | Mineral soil particles that are 0.002 to 0.05 mm in diameter | ||
| Organic matter content (% by weight) | Plant and animal residue in soil material <2 mm in diameter at various stages of decomposition | ||
| pH | A measure of acidity or alkalinity | ||
| Percent passing sieve No. 10 | Soil fraction passing a number 10 sieve (2.00 mm square opening) | ||
| Percent passing sieve No. 200 | Soil fraction passing a number 200 sieve (0.074 mm square opening) | ||
| Soil productivity [65] | Productivity Index derived from family-level Soil Taxonomy information | ||
| Soil taxonomic order | The percentage of taxonomic order | ||
| Soil texture | The percentage of clayey, loamy, sandy, or other texture class defined by USDA standard terms |
| Future: Current Ratios of Importance Values | ||||||
|---|---|---|---|---|---|---|
| Scenario | <0.5 | 0.5–0.9 | 0.9–1.1 | 1.1–2 | >2 | Total |
| CCSM45 | 6 | 18 | 14 | 64 | 23 | 125 |
| CCSM85 | 7 | 16 | 13 | 40 | 49 | 125 |
| GFDL45 | 8 | 15 | 13 | 39 | 50 | 125 |
| GFDL85 | 6 | 18 | 10 | 31 | 60 | 125 |
| HAD45 | 7 | 20 | 12 | 38 | 48 | 125 |
| HAD85 | 8 | 24 | 8 | 35 | 50 | 125 |
| GCM45 | 8 | 17 | 11 | 47 | 42 | 125 |
| GCM85 | 7 | 19 | 9 | 35 | 55 | 125 |
| Scenario | Kilometers | |||||||
|---|---|---|---|---|---|---|---|---|
| <10 | 10–50 | 51–100 | 101–200 | 201–300 | 301–400 | >400 | Total | |
| Northward | ||||||||
| CCSM45 | 1 | 16 | 22 | 43 | 10 | 0 | 1 | 93 |
| GCM45 | 1 | 12 | 19 | 33 | 19 | 7 | 8 | 99 |
| GCM85 | 0 | 9 | 11 | 25 | 23 | 16 | 17 | 101 |
| Had85 | 0 | 8 | 17 | 22 | 19 | 18 | 16 | 100 |
| Southward | ||||||||
| CCSM45 | 0 | 7 | 7 | 10 | 1 | 5 | 2 | 32 |
| GCM45 | 0 | 0 | 5 | 9 | 2 | 5 | 5 | 26 |
| GCM85 | 0 | 3 | 2 | 6 | 4 | 3 | 6 | 24 |
| Had85 | 0 | 2 | 4 | 5 | 7 | 2 | 5 | 25 |
| State/Region | First Species | Low | High | Second Species | Low | High | Third Species | Low | High |
|---|---|---|---|---|---|---|---|---|---|
| Eastern U.S. | Pinus taeda | 1.21 | 1.32 | Acer rubrum | 0.99 | 0.92 | Liquidambar styraciflua | 1.48 | 1.73 |
| Northeast | Acer rubrum | 0.91 | 0.76 | Acer saccharum | 0.93 | 0.88 | Pinus strobus | 1.04 | 0.87 |
| North Central | Populus tremuloides | 0.71 | 0.61 | Acer rubrum | 1 | 0.93 | Acer saccharum | 1.03 | 0.98 |
| Great Plains | Ulmus americana | 1.44 | 1.74 | Celtis occidentalis | 1.53 | 1.58 | Juniperus virginiana | 2.64 | 3.07 |
| Southeast | Pinus taeda | 1.07 | 1.09 | Pinus elliottii | 1.31 | 1.42 | Liquidambar styraciflua | 1.34 | 1.43 |
| South Central | Pinus taeda | 1.2 | 1.2 | Liquidambar styraciflua | 1.35 | 1.42 | Quercus stellata | 2.15 | 2.44 |
| Alabama | Pinus taeda | 0.98 | 0.95 | Liquidambar styraciflua | 1.17 | 1.21 | Quercus nigra | 2.04 | 2.29 |
| Arkansas | Pinus taeda | 1.35 | 1.36 | Pinus echinata | 1.55 | 1.56 | Quercus alba | 0.84 | 0.82 |
| Connecticut | Acer rubrum | 0.7 | 0.54 | Betula lenta | 0.65 | 0.54 | Quercus rubra | 0.89 | 0.77 |
| District of Columbia | Liriodendron tulipifera | 0.34 | 0.29 | Fagus grandifolia | 0.44 | 0.44 | Quercus prinus | 0.5 | 0.49 |
| Delaware | Acer rubrum | 0.77 | 0.69 | Pinus taeda | 2.38 | 2.4 | Liquidambar styraciflua | 1 | 1.07 |
| Florida | Pinus elliottii | 0.93 | 0.89 | Pinus taeda | 1.31 | 1.46 | Quercus laurifolia | 1.17 | 1.15 |
| Georgia | Pinus taeda | 0.97 | 0.95 | Pinus elliottii | 1.19 | 1.27 | Liquidambar styraciflua | 1.23 | 1.29 |
| Iowa | Ulmus americana | 1.23 | 1.27 | Juglans nigra | 1.39 | 1.33 | Celtis occidentalis | 2.24 | 2.02 |
| Illinois | Ulmus americana | 1.29 | 1.47 | Quercus alba | 0.92 | 0.76 | Prunus serotina | 0.77 | 0.66 |
| Indiana | Acer saccharum | 0.73 | 0.64 | Prunus serotina | 0.74 | 0.6 | Fraxinus americana | 0.99 | 0.94 |
| Kansas | Ulmus americana | 1.15 | 1.23 | Celtis occidentalis | 1.07 | 1.06 | Maclura pomifera | 1.21 | 1.22 |
| Kentucky | Liriodendron tulipifera | 0.44 | 0.35 | Acer saccharum | 0.66 | 0.6 | Quercus alba | 1.01 | 0.95 |
| Louisiana | Pinus taeda | 0.97 | 0.92 | Liquidambar styraciflua | 1.13 | 1.14 | Quercus nigra | 2.2 | 2.32 |
| Massachusetts | Acer rubrum | 0.81 | 0.66 | Pinus strobus | 0.46 | 0.38 | Quercus rubra | 1.02 | 0.92 |
| Maryland | Acer rubrum | 0.8 | 0.72 | Pinus taeda | 2.32 | 2.67 | Liriodendron tulipifera | 0.65 | 0.47 |
| Maine | Abies balsamea | 0.53 | 0.5 | Acer rubrum | 1.35 | 1.24 | Picea rubens | 0.64 | 0.57 |
| Michigan | Acer rubrum | 0.98 | 0.87 | Acer saccharum | 0.82 | 0.74 | Populus tremuloides | 0.89 | 0.8 |
| Minnesota | Populus tremuloides | 0.66 | 0.56 | Picea mariana | 0.4 | 0.4 | Larix laricina | 1.21 | 1.17 |
| Missouri | Quercus alba | 0.57 | 0.52 | Quercus velutina | 0.95 | 0.86 | Quercus stellata | 1.74 | 1.94 |
| Mississippi | Pinus taeda | 1.09 | 1.01 | Liquidambar styraciflua | 1.19 | 1.2 | Quercus nigra | 2.05 | 2.27 |
| North Carolina | Pinus taeda | 1.21 | 1.24 | Acer rubrum | 0.79 | 0.77 | Liquidambar styraciflua | 1.28 | 1.39 |
| North Dakota | Quercus macrocarpa | 1.75 | 2.58 | Populus tremuloides | 0.42 | 0.33 | Acer negundo | 1.92 | 1.63 |
| Nebraska | Juniperus virginiana | 1.92 | 2.33 | Quercus macrocarpa | 1.59 | 1.57 | Ulmus americana | 2.1 | 2.51 |
| New Hampshire | Acer rubrum | 1.06 | 0.91 | Pinus strobus | 0.79 | 0.65 | Tsuga canadensis | 0.77 | 0.66 |
| New Jersey | Pinus rigida | 0.39 | 0.39 | Acer rubrum | 1.05 | 0.92 | Quercus alba | 1 | 0.83 |
| New York | Acer rubrum | 0.98 | 0.77 | Acer saccharum | 0.78 | 0.71 | Fagus grandifolia | 0.73 | 0.67 |
| Ohio | Acer rubrum | 0.47 | 0.4 | Acer saccharum | 0.89 | 0.79 | Prunus serotina | 0.67 | 0.55 |
| Oklahoma | Quercus stellata | 1.09 | 1.08 | Juniperus virginiana | 1.24 | 1.34 | Pinus echinata | 1.36 | 1.4 |
| Pennsylvania | Acer rubrum | 0.64 | 0.47 | Prunus serotina | 0.77 | 0.63 | Acer saccharum | 0.91 | 0.86 |
| Rhode Island | Acer rubrum | 0.62 | 0.52 | Pinus strobus | 0.55 | 0.42 | Quercus rubra | 0.72 | 0.62 |
| South Carolina | Pinus taeda | 0.86 | 0.81 | Liquidambar styraciflua | 1.08 | 1.09 | Quercus nigra | 1.8 | 2.03 |
| South Dakota | Quercus macrocarpa | 5.57 | 5.65 | Juniperus virginiana | 5.87 | 6.93 | Fraxinus pennsylvanica | 3.75 | 4.15 |
| Tennessee | Liriodendron tulipifera | 0.51 | 0.42 | Quercus alba | 1.13 | 1.08 | Acer rubrum | 0.81 | 0.83 |
| Texas | Pinus taeda | 0.97 | 0.9 | Juniperus ashei | 1.11 | 1.1 | Quercus virginiana | 1.49 | 1.55 |
| Virginia | Pinus taeda | 1.45 | 1.66 | Liriodendron tulipifera | 0.5 | 0.33 | Acer rubrum | 0.82 | 0.76 |
| Vermont | Acer saccharum | 0.61 | 0.57 | Acer rubrum | 1.39 | 1.25 | Fagus grandifolia | 0.76 | 0.72 |
| Wisconsin | Populus tremuloides | 0.7 | 0.62 | Acer rubrum | 0.97 | 0.8 | Acer saccharum | 0.94 | 0.92 |
| West Virginia | Acer rubrum | 0.71 | 0.61 | Liriodendron tulipifera | 0.84 | 0.56 | Acer saccharum | 0.84 | 0.73 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iverson, L.R.; Peters, M.P.; Prasad, A.M.; Matthews, S.N. Analysis of Climate Change Impacts on Tree Species of the Eastern US: Results of DISTRIB-II Modeling. Forests 2019, 10, 302. https://doi.org/10.3390/f10040302
Iverson LR, Peters MP, Prasad AM, Matthews SN. Analysis of Climate Change Impacts on Tree Species of the Eastern US: Results of DISTRIB-II Modeling. Forests. 2019; 10(4):302. https://doi.org/10.3390/f10040302
Chicago/Turabian StyleIverson, Louis R., Matthew P. Peters, Anantha M. Prasad, and Stephen N. Matthews. 2019. "Analysis of Climate Change Impacts on Tree Species of the Eastern US: Results of DISTRIB-II Modeling" Forests 10, no. 4: 302. https://doi.org/10.3390/f10040302
APA StyleIverson, L. R., Peters, M. P., Prasad, A. M., & Matthews, S. N. (2019). Analysis of Climate Change Impacts on Tree Species of the Eastern US: Results of DISTRIB-II Modeling. Forests, 10(4), 302. https://doi.org/10.3390/f10040302