Potential Distribution Shifts of Plant Species under Climate Change in Changbai Mountains, China

Abstract
1. Introduction
2. Materials and Methods
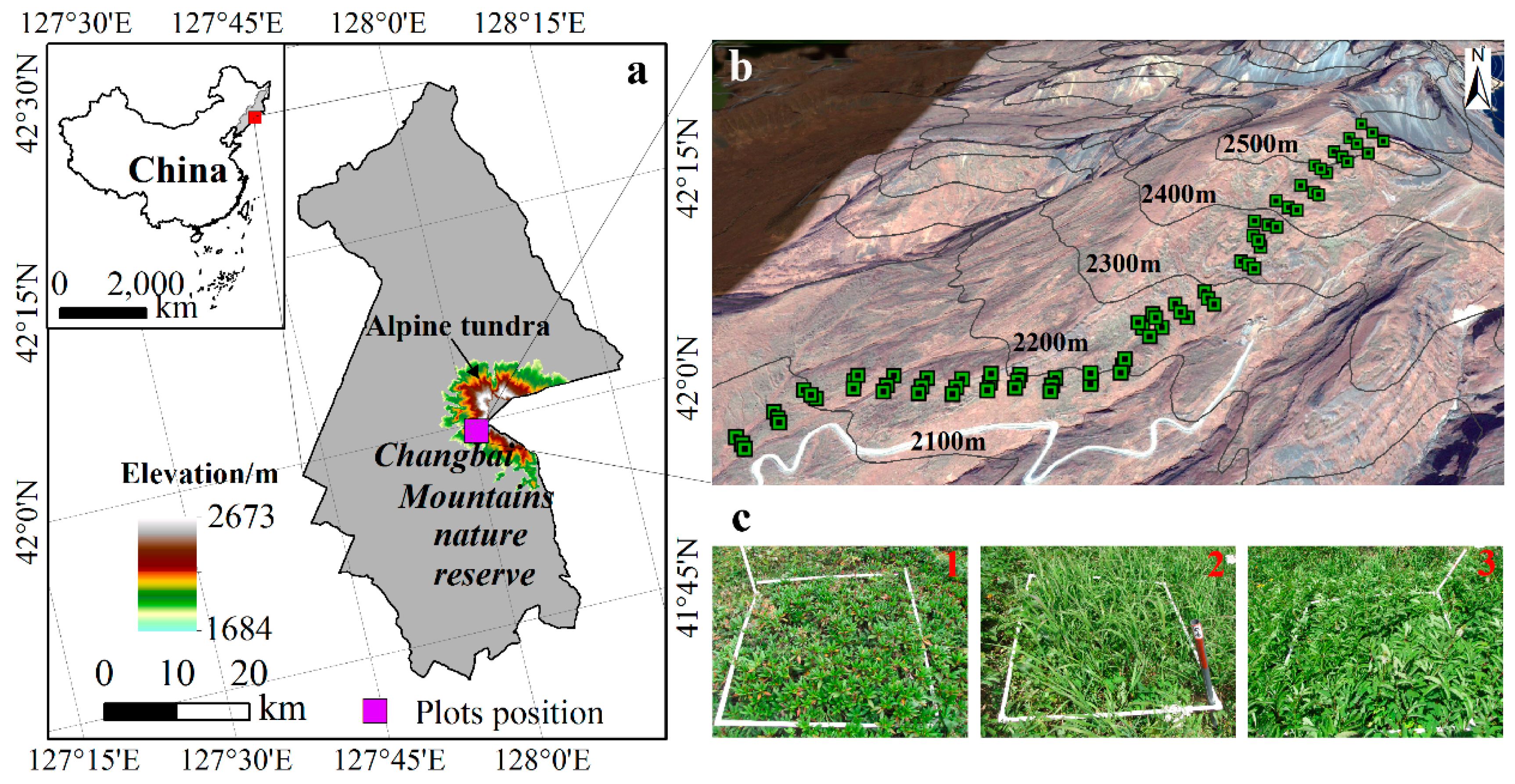
2.1. Study Area
2.2. Historical Vegetation Survey
2.3. Contemporary Vegetation Survey
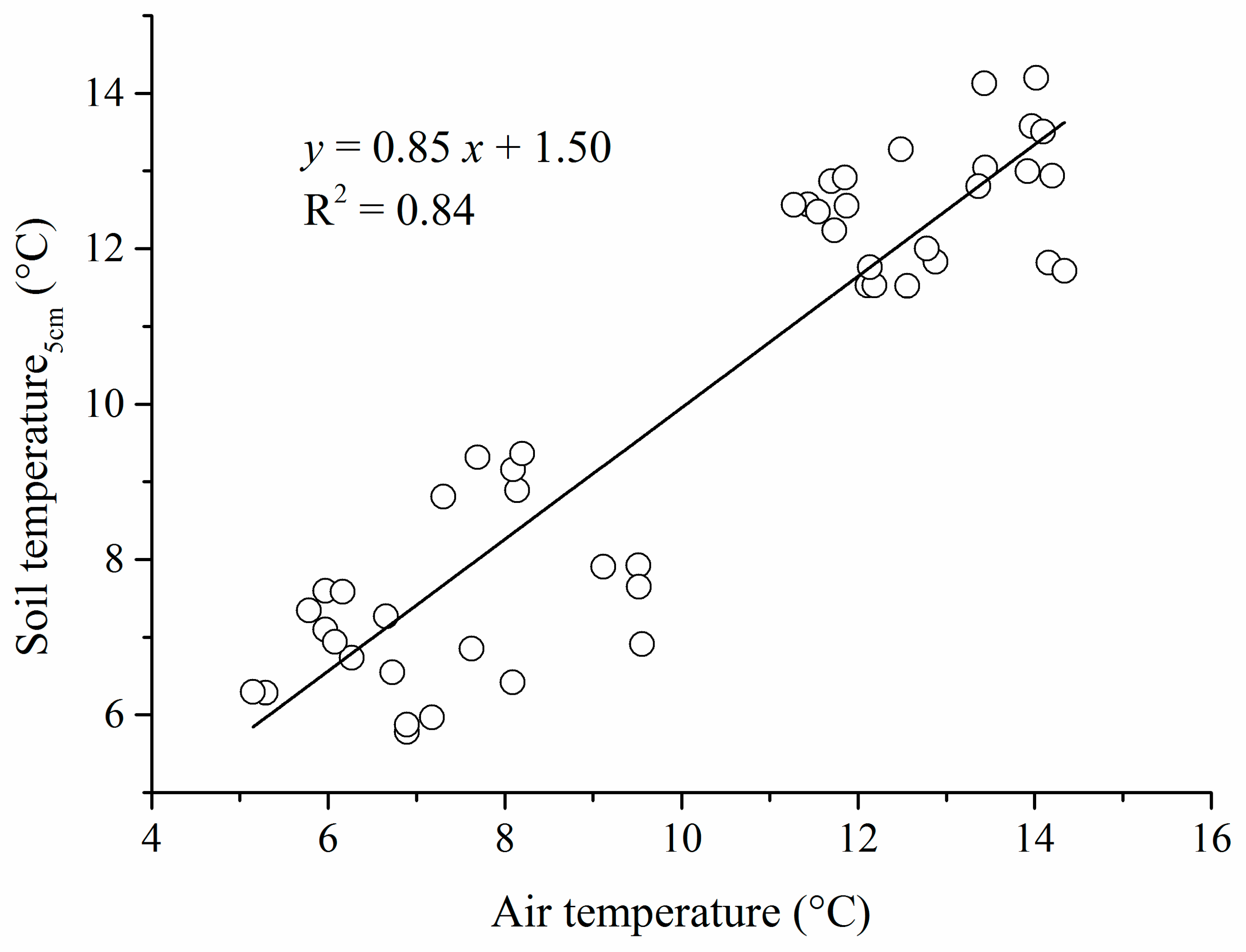
2.4. Historical and Future Climate Data
2.5. Climate Ecological Niche Modeling
3. Results
3.1. Historical and Future Changes in Climate
3.2. Changes in Alpine Tundra Vegetation over the Last 30 Years
3.3. Potential Shifts in Tundra Plant Species under Different Emission Scenarios
4. Discussion
4.1. Historical Plant Species Distribution Shifts within Alpine Tundra
4.2. Possible Factors Controlling Plant Species Shifts
4.3. Potential Plant Species Distribution Shifts within Alpine Tundra
4.4. Uncertainties in Potential Distribution Shifts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Korner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: New York, NY, USA, 2001. [Google Scholar]
- Theurillat, J.P.; Guisan, A. Potential Impact of Climate Change on Vegetation in the European Alps: A Review. Clim. Chang. 2001, 50, 77–109. [Google Scholar] [CrossRef]
- Diaz, H.F.; Eischeid, J.K. Disappearing “alpine tundra” Koppen climatic type in the western United States. Geophys. Res. Lett. 2007, 34, L18707. [Google Scholar] [CrossRef]
- Farrer, E.C.; Ashton, I.W.; Spasojevic, M.J.; Fu, S.; Gonzalez, D.J.X.; Suding, K.N. Indirect effects of global change accumulate to alter plant diversity but not ecosystem function in alpine tundra. J. Ecol. 2015, 103, 351–360. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range dynamics of mountain plants decrease with elevation. Proc. Natl. Acad. Sci. USA 2018, 115, 1848. [Google Scholar] [CrossRef] [PubMed]
- Formica, A.; Farrer, E.C.; Ashton, I.W.; Suding, K.N. Shrub Expansion Over the Past 62 Years in Rocky Mountain Alpine Tundra: Possible Causes and Consequences. Arct. Antarct. Alp. Res. 2002, 46, 616–631. [Google Scholar] [CrossRef]
- Dumais, C.; Ropars, P.; Denis, M.P.; Dufourtremblay, G.; Boudreau, S. Are low altitude alpine tundra ecosystems under threat? A case study from the Parc National de la Gaspésie, Québec. Environ. Res. Lett. 2014, 9, 094001. [Google Scholar] [CrossRef]
- Myerssmith, I.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Maciasfauria, M.; Sassklaassen, U.; Lévesque, E. Shrub expansion in tundra ecosystems: dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef]
- Ken, T.; Matthew, S.; Charles, R. The evidence for shrub expansion in Northern Alaska and the Pan-Arctic. Glob. Chang. Biol. 2006, 12, 686–702. [Google Scholar]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Forbes, B.C.; Macias Fauria, M.; Zetterberg, P. Russian Arctic warming and ‘greening’ are closely tracked by tundra shrub willows. Glob. Chang. Biol. 2010, 16, 1542–1554. [Google Scholar] [CrossRef]
- Cannone, N.; Sgorbati, S.; Guglielmin, M. Unexpected Impacts of Climate Change on Alpine Vegetation. Front. Ecol. Environ. 2007, 5, 360–364. [Google Scholar] [CrossRef]
- Czortek, P.; Eycott, A.E.; Grytnes, J.A.; Delimat, A.; Kapfer, J.; Jaroszewicz, B. Effects of grazing abandonment and climate change on mountain summits flora: A case study in the Tatra Mts. Plant Ecol. 2018, 219, 1–16. [Google Scholar] [CrossRef]
- Kapfer, J.; Hédl, R.; Jurasinski, G.; Kopecký, M.; Schei, F.H.; Grytnes, J.A. Resurveying historical vegetation data-opportunities and challenges. Appl. Veg. Sci. 2016, 20, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.G.; Mokany, K.; Guisan, A. Biodiversity Models: What If Unsaturation Is the Rule? Trends Ecol. Evol. 2017, 32, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W. Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Edwards, T.C.; Graham, C.H.; Pearman, P.B.; Svenning, J.C. New trends in species distribution modelling. Ecography 2011, 33, 985–989. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Chang. Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Taylor, S.; Kumar, L.; Reid, N.; Kriticos, D.J. Climate change and the potential distribution of an invasive shrub, Lantana camara L. PLoS ONE 2012, 7, e35565. [Google Scholar] [CrossRef]
- Walker, G.A.; Robertson, M.P.; Gaertner, M.; Gallien, L.; Richardson, D.M. The potential range of Ailanthus altissima (tree of heaven) in South Africa: the roles of climate, land use and disturbance. Biol. Invasions 2017, 19, 3675–3690. [Google Scholar] [CrossRef]
- Welk, E.; Schubert, K.; Hoffmann, M.H. Present and potential distribution of invasive garlic mustard (Alliaria petiolata) in North America. Divers. Distrib. 2002, 8, 219–233. [Google Scholar] [CrossRef]
- Kem, V.; Oksanen, L.; Oksanen, T.; Pyykönen, A.; Olofsson, J.; Virtanen, R. Open tundra persist, but arctic features decline-Vegetation changes in the warming Fennoscandian tundra. Glob. Chang. Biol. 2017, 23, 3794–3807. [Google Scholar]
- Zong, S.; Xu, J.; Wu, Z. Investigation and Mechanism Analysis on the Invasion of Deyeuxia angustifolia to Tundra Zone in Western Slope of Changbai Mountain. J. MT. Sci.-Engl. 2013, 31, 448–455. [Google Scholar]
- Zong, S.; Wu, Z.; Du, H. Study on Climate Change in Alpine Tundra of the Changbai Mountain in Growing Season in Recent 52 Years. Arid Zone Res. 2013, 30, 41–49. [Google Scholar]
- Hao, Z.; Dai, L.; He, H.S. Potential response of major tree species to climate warming in Changbai Mountain, Northeast China. Chin. J. Appl. Ecol. 2001, 12, 653–658. [Google Scholar]
- Du, H.; Liu, J.; Li, M.H.; Büntgen, U.; Yang, Y.; Wang, L.; Wu, Z.; He, H.S. Warming-induced upward migration of the alpine treeline in the Changbai Mountains, northeast China. Glob. Chang. Biol. 2018, 24, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xing, M.; Lv, J.; Wang, X.; Chen, X. Response of rhizosphere soil microbial to Deyeuxia angustifolia encroaching in two different vegetation communities in alpine tundra. Sci. Rep.-UK 2017, 7, 43150. [Google Scholar] [CrossRef]
- Huang, X.; Li, C. An analysis on the ecology of alpine tundra landscape of Changbai Mountains. Acta Geogr. Sin. 1984, 39, 285–297. [Google Scholar]
- Qian, H. Numerical classification and ordination of plant communities in the alpine tundra of Mt. Changbai. Chin. J. Appl. Ecol. 1990, 1, 254–263. [Google Scholar]
- Samaritani, E.; Mitchell, E.A.D.; Rich, J.; Shrestha, J.; Fournier, B.; Frey, B. Soil bacterial communities and ecosystem functioning change more strongly with season than habitat in a restored floodplain. Appl. Soil Ecol. 2017, 112, 71–78. [Google Scholar] [CrossRef]
- Wang, L.; Wu, Z.; He, H.; Wang, F.; Du, H.; Zong, S. Changes in start, end, and length of frost-free season across Northeast China. Int. J. Climatol. 2017, 37, 271–283. [Google Scholar] [CrossRef]
- Marquardt, D.W. An algorithm for least-squares estimation of nonlinear parameters. J. Soc. Ind. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank correlation methods. Br. J. Psychol. 1990, 25, 86–91. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052. [Google Scholar] [CrossRef] [PubMed]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: New York, NY, USA, 2014. [Google Scholar]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927. [Google Scholar] [CrossRef]
- Fang, J.; Lechowicz, M. Climatic limits for the present distribution of beech (Fagus L.) species in the world. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Kira, T. Forest ecosystems of east and southeast Asia in a global perspective. Ecol. Res. 1991, 6, 185–200. [Google Scholar] [CrossRef]
- Xu, W. The relation between the zonal distribution of types of vegetation and the climate in northeast China. Chin. J. Plant Ecol. 1986, 10, 16–25. [Google Scholar]
- Yim, Y.J. Distribution of forest vegetation and climate in the Korean Peninsula: I. distribution of some indices of thermal climate. Jpn. J. Ecol. 1977, 25, 77–88. [Google Scholar]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Elith, J. Quantitative Methods for Modeling Species Habitat: Comparative Performance and an Application to Australian Plants; Springer: New York, NY, USA, 2000. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Czortek, P.; Delimat, A.; Dyderski, M.K.; Zięba, A.; Jagodziński, A.M.; Jaroszewicz, B. Climate change, tourism and historical grazing influence the distribution of Carex lachenalii Schkuhr—A rare arctic-alpine species in the Tatra Mts. Sci. Total Environ. 2018, 618, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Paroloa, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; De, R.P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, J.; Dvorsky, M.; Kopecky, M.; Liancourt, P.; Hiiesalu, I.; Macek, M.; Altman, J.; Chlumska, Z.; Rehakova, K.; Capkova, K. Vegetation dynamics at the upper elevational limit of vascular plants in Himalaya. Sci. Rep.-UK 2016, 6, 24881. [Google Scholar] [CrossRef] [PubMed]
- Grytnes, J.A.; Kapfer, J.; Jurasinski, G.; Birks, H.H.; Henriksen, H.; Klanderud, K.; Odland, A.; Ohlson, M.; Wipf, S.; Birks, H.J.B. Identifying the driving factors behind observed elevational range shifts on European mountains. Glob. Ecol. Biogeogr. 2014, 23, 876–884. [Google Scholar] [CrossRef]
- Scherrer, D.; Körner, C. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Harvey, J.W.; Bencala, K.E. The effect of streambed topography on surface-subsurface water exchange in mountain catchments. Water Resour. Res. 1993, 29, 89–98. [Google Scholar] [CrossRef]
- Britton, A.J.; Beale, C.M.; Towers, W.; Hewison, R.L. Biodiversity gains and losses: Evidence for homogenisation of Scottish alpine vegetation. Biol. Conserv. 2009, 142, 1728–1739. [Google Scholar] [CrossRef]
- Jin, Y.; Xu, J.; Zong, S.; Wang, P. Experimental study on the effects of nitrogen deposition on the tundra vegetation of the Changbai Mountains. Sci. Geogr. Sin. 2014, 34, 1526–1532. [Google Scholar]
- Jin, S.; Yang, X.; Ma, J. The influence of rainfall on tourist flow in Changbai Mountains from June to October. Jilin Qixiang 2015, 22, 9–13. [Google Scholar]
- Zhu, H.; Ge, Z.; Liu, G.; Chang, S.; Wu, J.; Shi, X.; Xu, J.; Mao, Z. Population Status of Wild Boar (Sus scrofa ussuricus) in the Changbai Mountians, Jilin Province, China. Amino Acid. Biotic Resour. 2011, 33, 13–16. [Google Scholar]
- Czortek, P.; Kapfer, J.; Delimat, A.; Eycott, A.E.; Grytnes, J.A.; Orczewska, A.; Ratyńska, H.; Zięba, A.; Jaroszewicz, B. Plant species composition shifts in the Tatra Mts as a response to environmental change: A resurvey study after 90 years. Folia Geobot. 2018, 1–16. [Google Scholar] [CrossRef]
- Felde, V.A.; Kapfer, J.; Grytnes, J.A. Upward shift in elevational plant species ranges in Sikkilsdalen, central Norway. Ecography 2012, 35, 922–932. [Google Scholar] [CrossRef]
- Koide, D.; Yoshida, K.; Daehler, C.C.; Mueller-Dombois, D. An upward elevational shift of native and non-native vascular plants over 40 years on the island of Hawai’i. J. Veg. Sci. 2017, 28, 939–950. [Google Scholar] [CrossRef]
- Scherrer, D.; Schmid, S.; Körner, C. Elevational species shifts in a warmer climate are overestimated when based on weather station data. Int. J. Biometeorol. 2011, 55, 645–654. [Google Scholar] [CrossRef]
- Ganguly, A.R.; Steinhaeuser, K.; Erickson, D.J.; Branstetter, M.; Parish, E.S.; Singh, N.; Drake, J.B.; Buja, L.; Schneider, S.H. Higher Trends but Larger Uncertainty and Geographic Variability in 21st Century Temperature and Heat Waves. Proc. Natl. Acad. Sci. USA 2009, 106, 15555–15559. [Google Scholar] [CrossRef]
- Dubuis, A.; Giovanettina, S.; Pellissier, L.; Pottier, J.; Vittoz, P.; Guisan, A. Improving the prediction of plant species distribution and community composition by adding edaphic to topo-climatic variables. J. Veg. Sci. 2013, 24, 593–606. [Google Scholar] [CrossRef]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Historic Vegetation Survey (1984) | Current Vegetation Survey (2015) | ||||
|---|---|---|---|---|---|
| Elevation (m) | Vegetation Type | Dominant Plant Species | Elevation (m) | Vegetation Type | Dominant Plant Species |
| 2000–2200 | Dwarf shrub-lichen | I II III | 2000–2100 | Herb-dwarf shrub | VII VIII I |
| 2200–2400 | Dwarf shrub-moss | III IV I | 2100–2300 | Dwarf shrub-herb | I VII VIII IX |
| >2400 | Sparse shrub | V III VI | 2300–2400 | Dwarf shrub-lichen | I II III |
| >2400 | Sparse shrub | V III VI | |||
| Plant Species | Frequency (%) | Important Value |
|---|---|---|
| R. chrysanthum | 65.3 | 21.57 |
| S. sitchensis | 41.3 | 7.61 |
| Sanguisorba teriuifolia Var. | 52.0 | 6.23 |
| D. angustifolia | 29.3 | 4.86 |
| S. tomentosa | 52.0 | 4.79 |
| Ligularia jamesii (Hemsl) Kom. | 32.0 | 4.24 |
| V. uliginosum | 26.7 | 4.10 |
| Polygonum viviparum L. | 46.7 | 3.82 |
| D. octopetala | 22.7 | 3.29 |
| Name | R. chrysanthum | S. sitchensis | D. angustifolia | |
|---|---|---|---|---|
| RCP4.5 | Percentage of plots occupied (%) | 2 ± 12 ↓ | 7 ± 17 ↑ | 2 ± 12 ↑ |
| Lower limit of elevation (m) | 0 | -- | -- | |
| Upper limit of elevation (m) | 50 ± 37 ↑ | 25 ± 37 ↑ | 125 ± 40 ↑ | |
| RCP8.5 | Percentage of plots occupied (%) | 27 ± 16 ↓ | 52 ± 17 ↑ | 38 ± 10 ↑ |
| Lower limit of elevation (m) | 75 ± 43 ↑ | -- | -- | |
| Upper limit of elevation (m) | 50 ± 0 ↑ | 125 ± 52 ↑ | 225 ± 35 ↑ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, W.J.; Wu, Z.; Du, H.; Zong, S.; Ma, S. Potential Distribution Shifts of Plant Species under Climate Change in Changbai Mountains, China. Forests 2019, 10, 498. https://doi.org/10.3390/f10060498
Wang L, Wang WJ, Wu Z, Du H, Zong S, Ma S. Potential Distribution Shifts of Plant Species under Climate Change in Changbai Mountains, China. Forests. 2019; 10(6):498. https://doi.org/10.3390/f10060498
Chicago/Turabian StyleWang, Lei, Wen J. Wang, Zhengfang Wu, Haibo Du, Shengwei Zong, and Shuang Ma. 2019. "Potential Distribution Shifts of Plant Species under Climate Change in Changbai Mountains, China" Forests 10, no. 6: 498. https://doi.org/10.3390/f10060498
APA StyleWang, L., Wang, W. J., Wu, Z., Du, H., Zong, S., & Ma, S. (2019). Potential Distribution Shifts of Plant Species under Climate Change in Changbai Mountains, China. Forests, 10(6), 498. https://doi.org/10.3390/f10060498