Wildfire Alters Spatial Patterns of Available Soil Nitrogen and Understory Environments in a Valley Boreal Larch Forest




Abstract
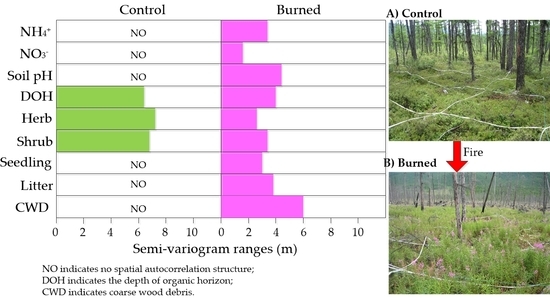
:
1. Introduction
2. Materials and Methods
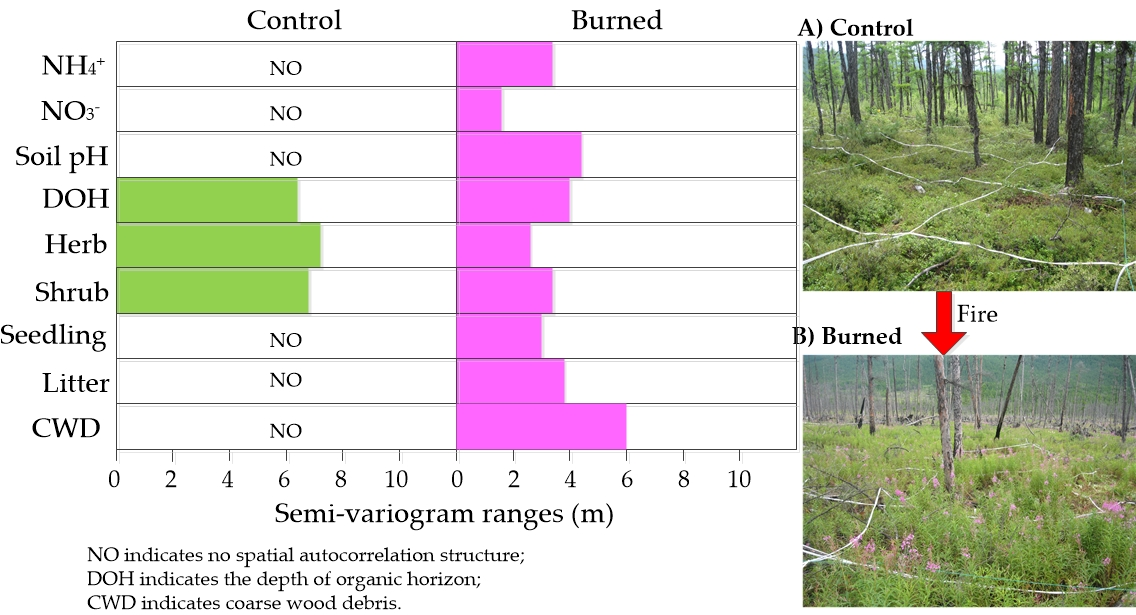
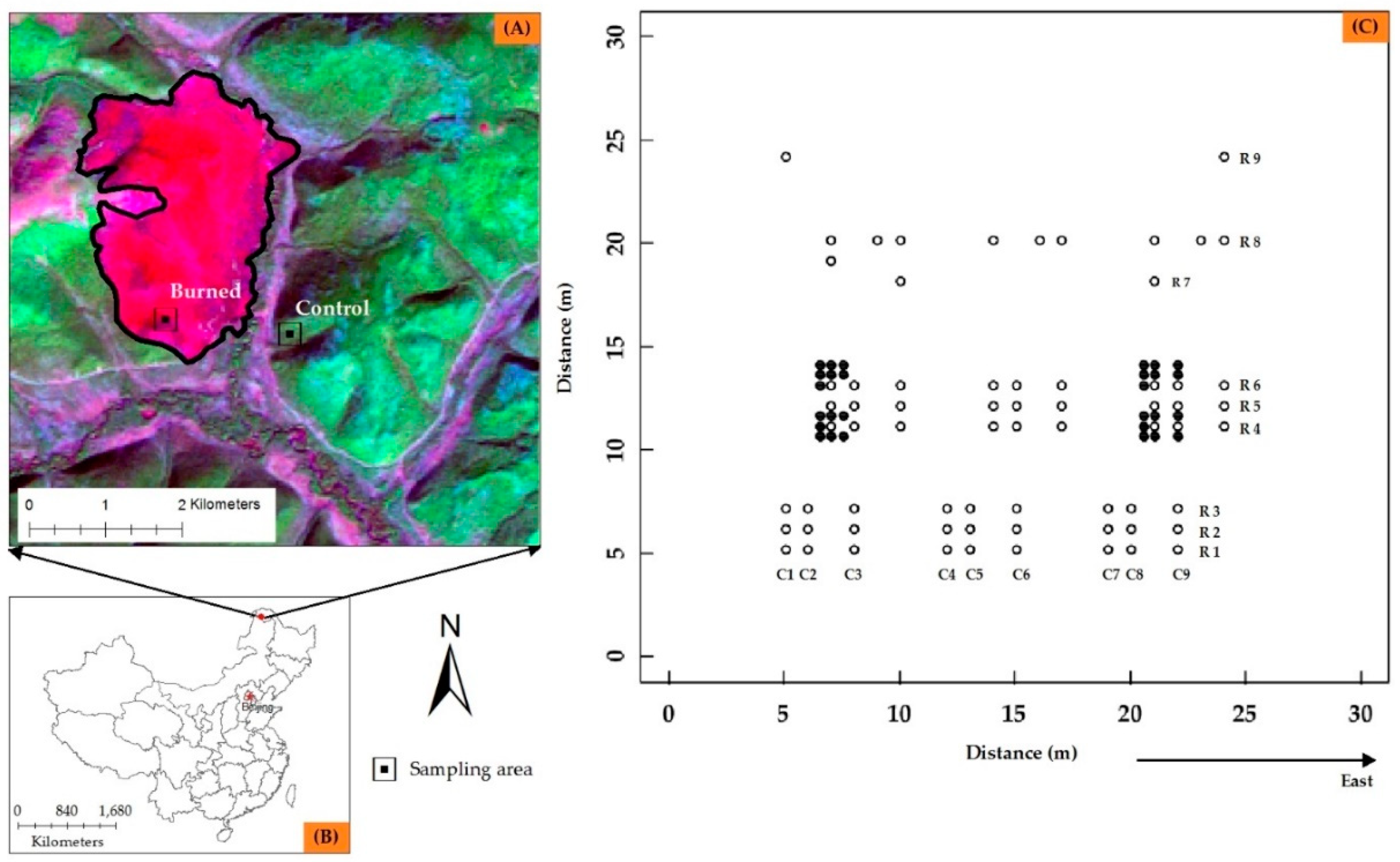
2.1. Study Area
2.2. Field Sampling and Laboratory
2.3. Statistical Analysis
3. Results
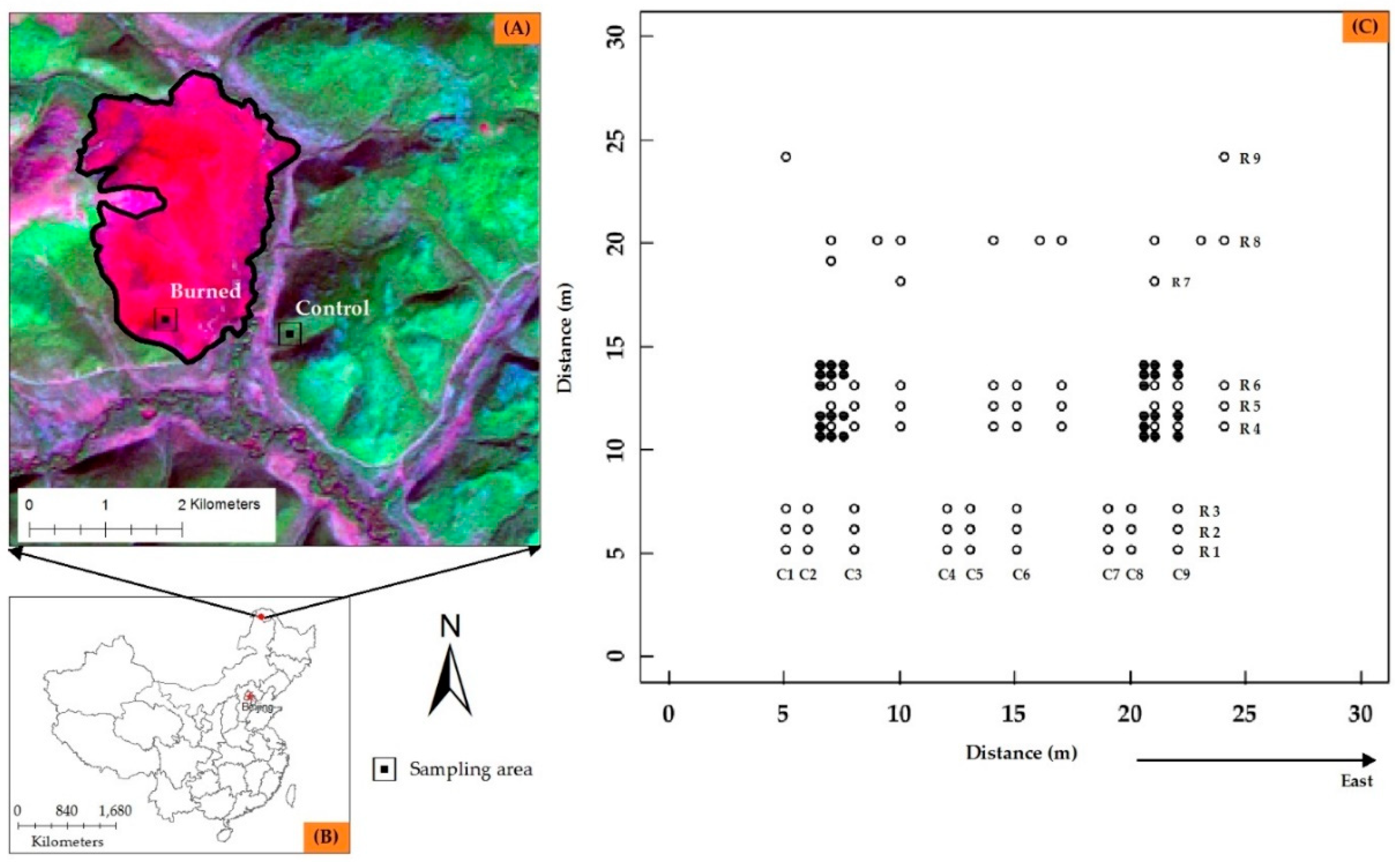
3.1. Within-Plot Variability of Soil N Availability and Site Characteristics
3.2. Spatial Structure Andheterogeneity of within-Plot Variability
3.3. Relationships between Soil N Availability and Local Site Characteristics
4. Discussion
4.1. Within-Plot Variability of Soil N and Site Characteristics
4.2. Changes in Spatial Heterogeneity in Soil N and Site Characteristics
4.3. Relationship between Soil N and Site Characteristics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 3-Year-Post-Fire | |
|---|---|---|
| Forest Floor | ||
| Organic horizon depth (cm) | 12–55 | 12–40 |
| Litter load (kg m−2) | 3.3 × 104–5.4 × 104 | - |
| CWD load (m3 ha−2) | 5.92–43.53 | - |
| Understory vegetation | ||
| Herbaceous height (cm) | 10–40 | 5–100 |
| Shrub height (cm) | 3–30 | |
| Rhododendron dauricum L. | 50–130 | - |
| Vaccinium uliginosum L. | 23–35 | 2–6 |
| Ledum palustre L. | 9–40 | 2–8 |
| Vacciniumvitis-idaea L. | 5–10 | 2–5 |
| Pinuspumila | 60–150 | - |
| Larch trees | ||
| Height (m) | 15–30 | - |
| DBH (cm) | 20–28 | - |
References
- Cohn, J.S.; Di Stefano, J.; Christie, F.; Cheers, G.; York, A. How do heterogeneity in vegetation types and post-fire age-classes contribute to plant diversity at the landscape scale? For. Ecol. Manag. 2015, 346, 22–30. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Hutson, M.A.; Evans, F.C.; Tiedje, J.M. Spatial variability in a successional plant community: Patterns of nitrogen availability. Ecology 1988, 69, 1517–1524. [Google Scholar] [CrossRef]
- Gallardo, A.; Paramá, R.; Covelo, F. Differences between soil ammonium and nitrate spatial pattern in six plant communities. Simulated effect on plant populations. Plant Soil 2006, 279, 333–346. [Google Scholar] [CrossRef]
- Hutchings, M.J.; John, E.A.; Wijesinghea, D.K. Toward understanding the consequences of soil heterogeneity for plant populations and communities. Ecology 2003, 84, 2322–2334. [Google Scholar] [CrossRef]
- De Long, J.R.; Dorrepaal, E.; Kardol, P.; Nilsson, M.-C.; Teuber, L.M.; Wardle, D.A. Understory plant functional groups and litter species identity are stronger drivers of litter decomposition than warming along a boreal forest post-fire successional gradient. Soil Biol. Biochem. 2016, 98, 159–170. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Smithwick, E.A.H.; Tinker, D.B.; Zhu, J. Variation in aboveground cover influences soil nitrogen availability at fine spatial scales following severe fire in subalpine conifer forests. Ecosystems 2011, 14, 1081–1095. [Google Scholar] [CrossRef]
- Saar, S.; Semchenko, M.; Barel, J.M.; de Deyn, G.B. Spatial heterogeneity in root litter and soil legacies differentially affect legume root traits. Plant Soil 2018, 428, 253–264. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Wallenstein, M.D.; Quero, J.L.; Ochoa, V.; Gozalo, B.; Garcia-Gomez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef]
- Ford, S.; Kleinman, J.; Hart, J. Spatial patterns of canopy disturbance, structure, and species composition in a multi-cohort hardwood stand. Forests 2017, 8, 93. [Google Scholar] [CrossRef]
- Lavoie, M.; Mack, M.C. Spatial heterogeneity of understory vegetation and soil in an Alaskan upland boreal forest fire chronosequence. Biogeochemistry 2012, 107, 227–239. [Google Scholar] [CrossRef]
- Fang, L.; Yang, J.; White, M.; Liu, Z. Predicting potential fire severity using vegetation, topography and surface moisture availability in a eurasian boreal forest landscape. Forests 2018, 9, 130. [Google Scholar] [CrossRef]
- Kasischke, E.S.; Turetsky, M.R.; Ottmar, R.D.; French, N.H.F.; Hoy, E.E.; Kane, E.S. Evaluation of the composite burn index for assessing fire severity in Alaskan black spruce forests. Int. J. Wildland Fire 2008, 17, 515–526. [Google Scholar] [CrossRef]
- Flannigan, M.; Stocks, B.; Turetsky, M.; Wotton, M. Impacts of climate change on fire activity and fire management in the circumboreal forest. Glob. Chang. Biol. 2009, 15, 549–560. [Google Scholar] [CrossRef]
- Tokuchi, N.; Hirobe, M.; Kondo, K.; Arai, H.; Hobara, S.; Fukushima, K.; Matsuura, Y. Soil Nitrogen Dynamics in Larch Ecosystem. In Permafrost Ecosystems: Siberian Larch Forests; Springer: New York, NY, USA, 2010; pp. 229–244. [Google Scholar]
- DeLuca, T.; Sala, A. Frequent fire alters nitrogen transformations in ponderosa pine stands of the inland northwest. Ecology 2006, 87, 2511–2522. [Google Scholar] [CrossRef]
- Harden, J.W.; Mack, M.; Veldhuis, H.; Gower, S.T. Fire dynamics and implications for nitrogen cycling in boreal forests. J. Geophys. Res. 2002, 108, 8223–8230. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Neary, D.G.; Ryan, K.C.; de Bano, L.F. Wildland Fire in Ecosystems: Effects of Fire on Soil and Water; Gen. Tech. Rep. RMRS-GTR-42-vol.4; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2005; p. 250. [Google Scholar]
- Smithwick, E.A.H.; Turner, M.G.; Mack, M.C.; Chapin, F.S. Postfire soil N cycling in northern conifer forests affected by severe, stand-replacing wildfires. Ecosystems 2005, 8, 163–181. [Google Scholar] [CrossRef]
- Kong, J.-J.; Yang, J.; Chu, H.; Xiang, X. Effects of wildfire and topography on soil nitrogen availability in a boreal larch forest of northeastern China. Int. J. Wildland Fire 2015, 24, 433–442. [Google Scholar] [CrossRef]
- Wan, S.; Hui, D.; Luo, Y. Fire effects on nitrogen pools and dynamics in terrestrial ecosystems: A meta-analysis. Ecol. Appl. 2001, 11, 1349–1365. [Google Scholar] [CrossRef]
- Adkins, J.; Sanderman, J.; Miesel, J. Soil carbon pools and fluxes vary across a burn severity gradient three years after wildfire in Sierra Nevada mixed-conifer forest. Geoderma 2019, 333, 10–22. [Google Scholar] [CrossRef]
- Koyama, A.; Kavanagh, K.; Stephan, K. Wildfire effects on soil gross nitrogen transformation rates in coniferous forests of Central Idaho, USA. Ecosystems 2010, 13, 1112–1126. [Google Scholar] [CrossRef]
- Rodríguez, A.; Durán, J.; Fernández-Palacios, J.M.; Gallardo, A. Wildfire changes the spatial pattern of soil nutrient availability in Pinus canariensis forests. Ann. For. Sci. 2009, 66, 210. [Google Scholar] [CrossRef]
- Das Gupta, S.; MacKenzie, M.D. Recovery from fire affects spatial variability of nutrient availability in boreal aspen ecosystems. bioRxiv 2018. [Google Scholar] [CrossRef]
- Osawa, A.; Zyryanova, O.A.; Matsuura, Y.; Kajimoto, T.; Wein, R.W. Permafrost Ecosystems: Siberian Larch Forests; Springer: New York, NY, USA, 2010. [Google Scholar]
- Jackson, R.B.; Caldwell, M.M. Geostatistical patterns of soil heterogeneity around individual perennial plants. J. Ecol. 1993, 81, 683–692. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Raikes, J.A.; Hartley, A.E.; Cross, A.F. On the spatial pattern of soil nutrients in desert ecosystems. Ecology 1996, 77, 364–374. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Mack, M.C.; Turner, M.G.; Chapin, F.S.; Zhu, J.; Balser, T.C. Spatial heterogeneity and soil nitrogen dynamics in a burned black spruce forest stand: Distinct controls at different scales. Biogeochemistry 2005, 76, 517–537. [Google Scholar] [CrossRef]
- Boby, L.A.; Schuur, E.A.G.; Mack, M.C.; Verbyla, D.; Johnstone, J.F. Quantifying fire severity, carbon, and nitrogen emissions in Alaska’s boreal forest. Ecol. Appl. 2010, 20, 1633–1647. [Google Scholar] [CrossRef]
- Alexander, H.D.; Natali, S.M.; Loranty, M.M.; Ludwig, S.M.; Spektor, V.V.; Davydov, S.; Zimov, N.; Trujillo, I.; Mack, M.C. Impacts of increased soil burn severity on larch forest regeneration on permafrost soils of far northeastern Siberia. For. Ecol. Manag. 2018, 417, 144–153. [Google Scholar] [CrossRef]
- Cai, W.; Yang, J.; Liu, Z.; Hu, Y.; Weisberg, P.J. Post-fire tree recruitment of a boreal larch forest in Northeast China. For. Ecol. Manag. 2013, 307, 20–29. [Google Scholar] [CrossRef]
- Brown, C.D.; Liu, J.; Yan, G.; Johnstone, J.F. Disentangling legacy effects from environmental filters of postfire assembly of boreal tree assemblages. Ecology 2015, 96, 3023–3032. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.P.; Hoffman, C.; Battaglia, M.; Mell, W. Spatially explicit measurements of forest structure and fire behavior following restoration treatments in dry forests. For. Ecol. Manag. 2017, 386, 1–12. [Google Scholar] [CrossRef]
- Hirobe, M.; Tokuchi, N.; Wachrinrat, C.; Takeda, H. Fire history influences on the spatial heterogeneity of soil nitrogen transformations in three adjacent stands in a dry tropical forest in Thailand. Plant Soil 2003, 249, 309–318. [Google Scholar] [CrossRef]
- Liu, B.; Yang, J.; Johnstone, J.F. Understory vascular plant community assembly in relation to time-since-fire and environmental variables in a Chinese boreal forest. J. Mt. Sci. 2017, 14, 1317–1328. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, J. Quantifying ecological drivers of ecosystem productivity of the early-successional borealLarix gmeliniiforest. Ecosphere 2014, 5, 1–16. [Google Scholar] [CrossRef]
- Xu, H.C. Forests in Daxing’anling Mountains China; Science Press: Beijing, China, 1998. [Google Scholar]
- Gong, Z.T. Chinese Soil Taxonomy; Science Press: Beijing, China, 2003. [Google Scholar]
- Chang, Y.; He, H.S.; Hu, Y.; Bu, R.; Li, X. Historic and current fire regimes in the Great Xing’an Mountains, northeastern China: Implications for long-term forest management. For. Ecol. Manag. 2008, 254, 445–453. [Google Scholar] [CrossRef]
- Hu, T.; Zhou, G. Drivers of lightning- and human-caused fire regimes in the Great Xing’an Mountains. For. Ecol. Manag. 2014, 329, 49–58. [Google Scholar] [CrossRef]
- Liu, Z.H.; Yang, J.; Chang, Y.; Weisberg, P.J.; He, H.S. Spatial patterns and drivers of fire occurrence and its future trend under climate change in a boreal forest of Northeast China. Glob. Chang. Biol. 2012, 18, 2041–2056. [Google Scholar] [CrossRef]
- Clayton, M.K.; Hudelson, B.D. Confidence intervals for autocorrelations based on cyclic samples. J. Am. Stat. Assoc. 1995, 90, 753–757. [Google Scholar] [CrossRef]
- Fraterrigo, J.M.; Rusak, J.A. Disturbance-driven changes in the variability of ecological patterns and processes. Ecol. Lett. 2008, 11, 756–770. [Google Scholar] [CrossRef]
- Available online: https://en.wikipedia.org/wiki/Coefficient_of_variation (accessed on 25 January 2019).
- Cressie, N.A.C. Statistics for Spatial Data, Revised Edition; Wiley: New York, NY, USA, 1993. [Google Scholar]
- Legendre, P.; Fortin, M.J. Spatial pattern and ecological analysis. Vegetation 1989, 80, 107–138. [Google Scholar] [CrossRef]
- Turner, M.G.; Smithwick, E.A.; Metzger, K.L.; Tinker, D.B.; Romme, W.H. Inorganic nitrogen availability after severe stand-replacing fire in the Greater Yellowstone ecosystem. Proc. Natl. Acad. Sci. USA 2007, 104, 4782–4789. [Google Scholar] [CrossRef] [PubMed]
- Ste-Marie, C.; Paré, D. Soil, pH and N availability effects on net nitrification in the forest floors of a range of boreal forest stands. Soil Biol. Biochem. 1999, 31, 1579–1589. [Google Scholar] [CrossRef]
- Kong, J.J.; Yang, J.; Cai, W. Topography controls post-fire changes in soil properties in a Chinese boreal forest. Sci. Total Environ. 2019, 651, 2662–2670. [Google Scholar] [CrossRef]
- Hautala, H.; Tolvanen, A.; Nuortila, C. Regeneration strategies of dominant boreal forest dwarf shrubs in response to selective removal of understorey layers. J. Veg. Sci. 2001, 12, 503–510. [Google Scholar] [CrossRef]
- Baret, M.; Pepin, S.; Ward, C.; Pothier, D. Long-term changes in belowground and aboveground resource allocation of boreal forest stands. For. Ecol. Manag. 2015, 350, 62–69. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Roberts, M.R. Response of the herbaceous layer to natural disturbance in North American forests. Can. J. Bot. 2004, 82, 1273–1283. [Google Scholar] [CrossRef]
- Kong, J.J.; Yang, J.; Bai, E. Long-term effects of wildfire on available soil nutrient composition and stoichiometry in a Chinese boreal forest. Sci. Total Environ. 2018, 642, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Simard, D.; Fyles, J.; Paré, D.; Nguyen, T. Impacts of clearcut harvesting and wildfire on soil nutrient status in the Quebec boreal forest. Can. J. Soil Sci. 2001, 81, 229–237. [Google Scholar] [CrossRef]
- DeLuca, T.H.; MacKenzie, M.D.; Gundale, M.J.; Holben, W.E. Wildfire-produced charcoal directly influences nitrogen cycling in ponderosa pine forests. Soil Sci. Soc. Am. J. 2006, 70, 448–453. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Hollingsworth, T.N.; Chapin, F.S. A Key for Predicting Postfire Succession Trajectories in Black Spruce Stands of Interior Alaska; Pacific Northwest Research Station PNW GTR-767; US Department of Agriculture, Forest Service: Portland, OR, USA, 2008.
- DeLuca, T.H.; Aplet, G.H. Charcoal and carbon storage in forest soils of the Rocky Mountain West. Front. Ecol. Environ. 2008, 6, 18–24. [Google Scholar] [CrossRef]
- García-Palacios, P.; Maestre, F.T.; Bardgett, R.D.; de Kroon, H. Plant responses to soil heterogeneity and global environmental change. J. Ecol. 2012, 100, 1303–1314. [Google Scholar] [CrossRef]
- Guo, D.; Mou, P.; Jones, R.H.; Mitchell, R.J. Spatio-temporal patterns of soil available nutrients following experimental disturbance in a pine forest. Oecologia 2004, 138, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Stark, J.M.; Hart, S.C. High rates of nitrification and nitrate turnover in undisturbed coniferous forests. Nature 1997, 385, 61–64. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Turner, M.G.; Metzger, K.L.; Balser, T.C. Variation in mineralization and microbial communities with stand age in lodgepole pine (Pinus contorta) forests, Yellowstone National Park (USA). Soil Biol. Biochem. 2005, 37, 1546–1559. [Google Scholar] [CrossRef]
- Romme, W.H.; Tinker, D.B.; Stakes, G.K.; Turner, M.G. Does inorganic nitrogen limit plant growth 3–5 years after fire in a Wyoming, USA, lodgepole pine forest? For. Ecol. Manag. 2009, 257, 829–835. [Google Scholar] [CrossRef]
- Tsuyuzaki, S.; Narita, K.; Sawada, Y.; Kushida, K. The establishment patterns of tree seedlings are determined immediately after wildfire in a black spruce (Picea mariana) forest. Plant Ecol. 2014, 215, 327–337. [Google Scholar] [CrossRef]

| Treatment | Mean | Median | Min | Max | CV (%) | |
|---|---|---|---|---|---|---|
| Soil N variables (μg N 10 cm−2) | ||||||
| NO3− | Control | 2.4 b | 1.8 | 0.0 | 18.5 | 122 |
| Burned | 17.7 a | 9.4 | 0.0 | 71.5 | 114 | |
| NH4+ | Control | 5.4 a | 4.4 | 2.3 | 13.9 | 45 |
| Burned | 4.6 b | 4.2 | 2.5 | 10.6 | 33 | |
| Soil resource variables | ||||||
| pH | Control | 4.7 b | 4.7 | 4.2 | 5.1 | 4 |
| Burned | 5.1 a | 5.1 | 4.0 | 6.1 | 7 | |
| DOH (cm) | Control | 30.5 a | 28.0 | 12.0 | 55.0 | 33 |
| Burned | 26.0 b | 27.0 | 12.0 | 40.0 | 30 | |
| Aboveground cover variables (%) | ||||||
| Herb | Control | 7.2 b | 2.8 | 0.0 | 40.0 | 132 |
| Burned | 50.6 a | 50.0 | 10.0 | 98.0 | 46 | |
| Shrub | Control | 60.8 a | 55.0 | 2.0 | 100.0 | 43 |
| Burned | 3.5 b | 0.0 | 0.0 | 48.0 | 224 | |
| RTS | Control | 0.5 a | 0.0 | 0.0 | 25.0 | 598 |
| Burned | 0.3 a | 0.0 | 0.0 | 5.0 | 248 | |
| Moss | Control | 83.4 a | 100.0 | 0.0 | 100.0 | 43 |
| Burned | 77.9 a | 100.0 | 0.0 | 100.0 | 40 | |
| Lichen | Control | 21.4 b | 6.5 | 0.0 | 100.0 | 145 |
| Burned | 40.0 a | 30.0 | 0.0 | 100.0 | 90 | |
| Litter | Control | 9.6 b | 0.0 | 0.0 | 80.0 | 203 |
| Burned | 17.7 a | 5.0 | 0.0 | 95.0 | 130 | |
| CWD | Control | 1.5 b | 0.0 | 0.0 | 30. | 359 |
| Burned | 12.9 a | 0.0 | 0.0 | 100.0 | 170 | |
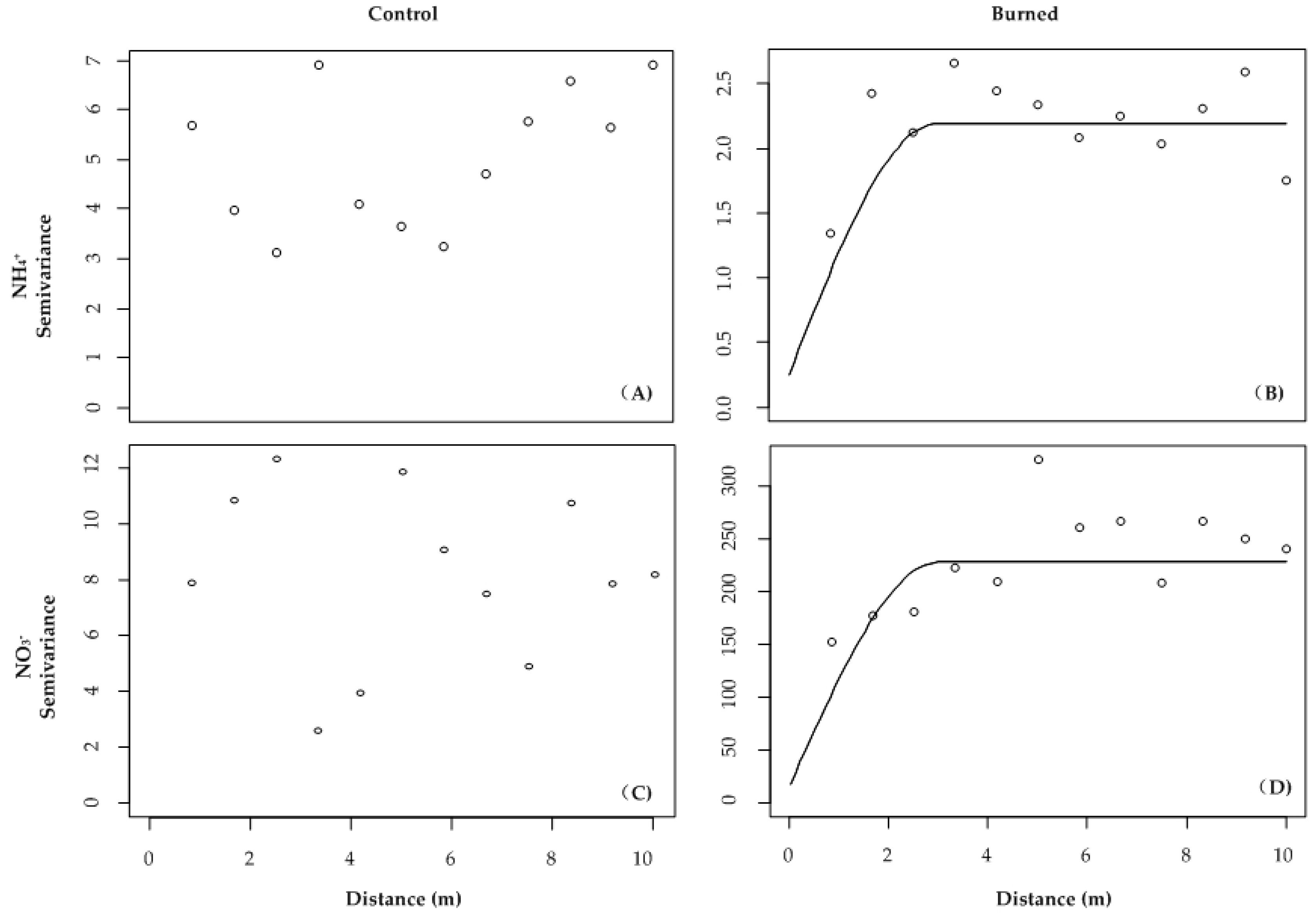
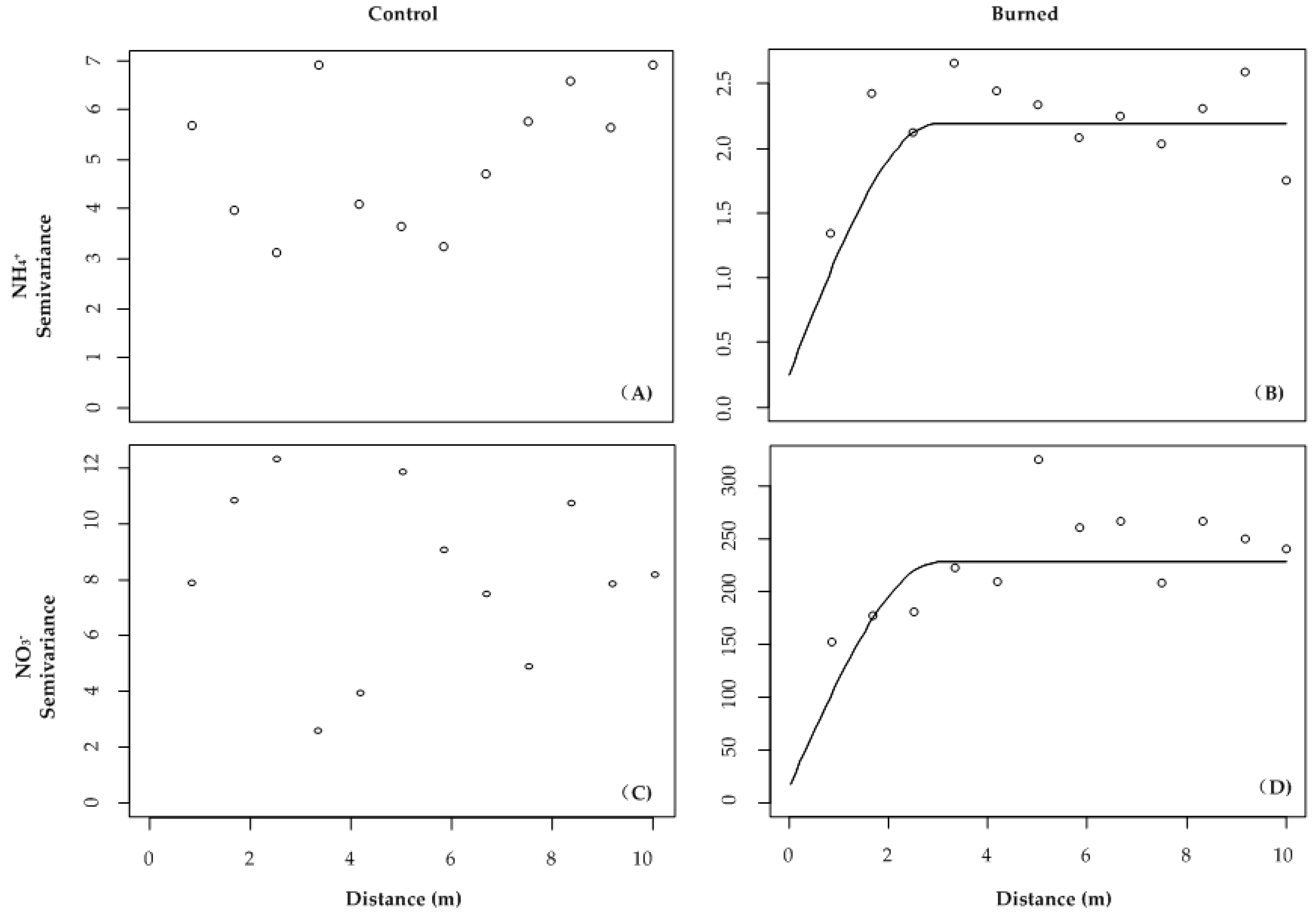
| Treatment | Range (m) | Sill | Nugget | C/(C + C0) (%) | Model | |
|---|---|---|---|---|---|---|
| Soil N variables (μg N 10 cm−2) | ||||||
| NH4+ | Control | - | - | - | - | Nugget |
| Burned | 3.5 | 2.08 | 0.27 | 90 | Spherical | |
| NO3− | Control | - | - | - | - | Nugget |
| Burned | 1.6 | 245 | 0 | 100 | Exponential | |
| Soil environmental variables | ||||||
| pH | Control | - | - | - | - | Nugget |
| Burned | 4.3 | 0.11 | 0.01 | 91 | Spherical | |
| DOH (cm) | Control | 6.5 | 0.06 | 0.03 | 50 | Spherical |
| Burned | 4.0 | 53.3 | 9.5 | 82 | Spherical | |
| Aboveground cover variables (%) | ||||||
| Herb | Control | 7.3 | 67.8 | 6.4 | 90 | Spherical |
| Burned | 2.7 | 0.21 | 0.07 | 67 | Spherical | |
| Shrub | Control | 6.9 | 86.3 | 15.7 | 82 | Exponential |
| Burned | 3.5 | 52.1 | 8.4 | 84 | Spherical | |
| RTS | Control | - | - | - | - | Nugget |
| Burned | 3.0 | 0.64 | 0.05 | 92 | Spherical | |
| Nugget | ||||||
| Litter | Control | - | - | - | - | Nugget |
| Burned | 3.8 | 514 | 38 | 93 | Spherical | |
| CWD | Control | - | - | - | - | Nugget |
| Burned | 6.0 | 28.8 | 3.44 | 89 | Exponential | |
| Local Site | NO3− | NH4+ | ||
|---|---|---|---|---|
| Characteristics | Control | Burned | Control | Burned |
| Soil resource variables | ||||
| DOH (cm) | 0.03 | 0.21 (0.002) | −0.21 (0.01) | −0.21 (0.002) |
| pH | 0.00 | 0.18 (0.01) | 0.06 | −0.14 (0.053) |
| Aboveground cover variables (%) | ||||
| RTS | 0.02 | −0.13 | −0.05 | 0.20 (0.004) |
| Herb | −0.11 | 0.12 | 0.07 | −0.16 (0.02) |
| Shrub | 0.02 | −0.10 | 0.03 | 0.04 |
| Litter | 0.00 | −0.04 | −0.20 (0.02) | 0.08 |
| CWD | −0.03 | 0.01 | 0.01 | 0.06 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, J.; Yang, J.; Liu, B.; Qi, L. Wildfire Alters Spatial Patterns of Available Soil Nitrogen and Understory Environments in a Valley Boreal Larch Forest. Forests 2019, 10, 95. https://doi.org/10.3390/f10020095
Kong J, Yang J, Liu B, Qi L. Wildfire Alters Spatial Patterns of Available Soil Nitrogen and Understory Environments in a Valley Boreal Larch Forest. Forests. 2019; 10(2):95. https://doi.org/10.3390/f10020095
Chicago/Turabian StyleKong, Jianjian, Jian Yang, Bo Liu, and Lin Qi. 2019. "Wildfire Alters Spatial Patterns of Available Soil Nitrogen and Understory Environments in a Valley Boreal Larch Forest" Forests 10, no. 2: 95. https://doi.org/10.3390/f10020095
APA StyleKong, J., Yang, J., Liu, B., & Qi, L. (2019). Wildfire Alters Spatial Patterns of Available Soil Nitrogen and Understory Environments in a Valley Boreal Larch Forest. Forests, 10(2), 95. https://doi.org/10.3390/f10020095