Do Terrestrial Salamanders Indicate Ecosystem Changes in New England Forests?


Abstract
1. Introduction
2. Materials and Methods
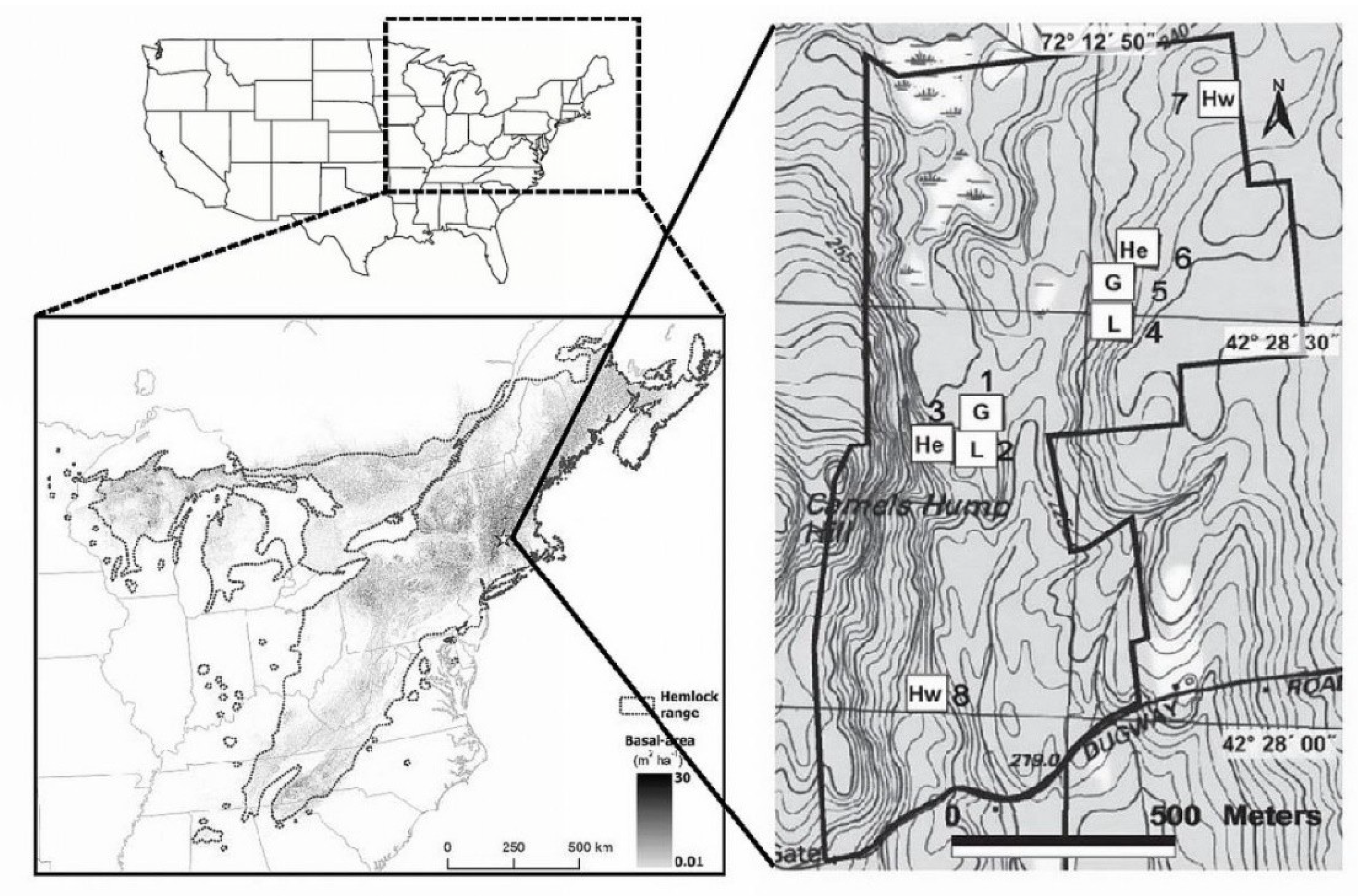
2.1. Study Sites
2.2. Region-Wide Hemlock Decline and the Harvard Forest Hemlock Removal Experiment (HF-HeRE)
2.3. Sampling Plethodontid Salamanders within HF-HeRE
2.4. Data Analysis and Data Availability
3. Results
3.1. Salamanders Are Responsive to Ecological Changes
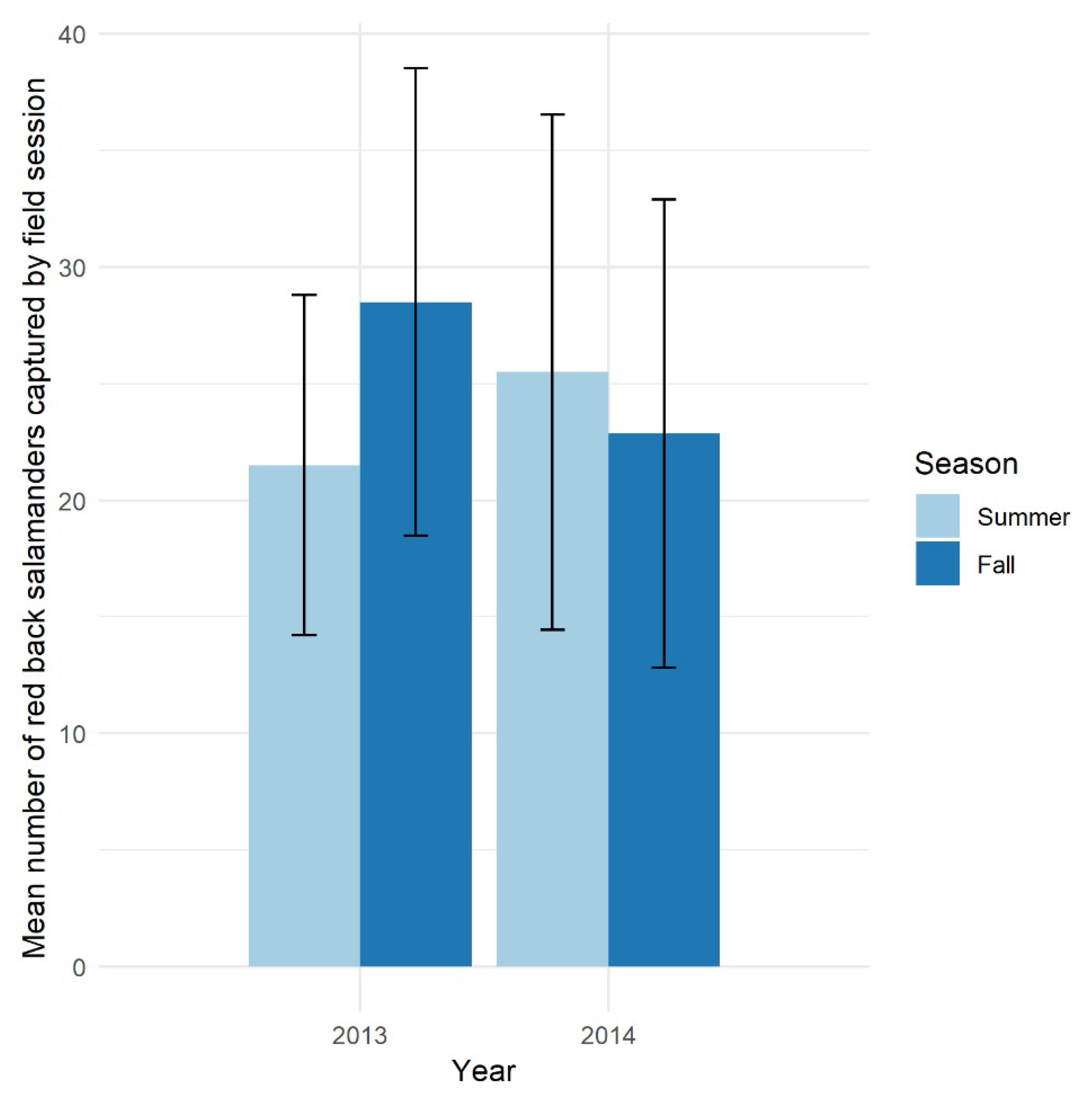
3.2. Abundant with Seasonal and Annual Stability
3.3. Ease of Sampling and Potential for Cost-Effective Monitoring
3.4. Limited Mobility and Very Local Activity
3.5. Indicator Values of Red Backs and Red Efts within HF-HeRE
4. Discussion
4.1. The Utility of Salamanders as Indicator Species
4.2. Examples of Salamanders as Potential Indicators of Ecosystem Change at Other Forested US LTER Sites
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Likens, G.E. Long-Term Studies in Ecology: Approaches and Alternatives; Springer: New York, NY, USA, 1989; ISBN 9781461573586. [Google Scholar]
- Morrison, M.L. Restoring Wildlife: Ecological Concepts and Practical Applications; Island Press: Washington, DC, USA, 2009. [Google Scholar]
- Gitzen, R.A.; Millspaugh, J.J.; Cooper, A.B.; Licht, D.S. Design and Analysis of Long-Term Ecological Monitoring Studies; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Peters, D.P.C.; Laney, C.M.; Lugo, A.E.; Collins, S.L.; Driscoll, C.T.; Groffman, P.M.; Grove, J.M.; Knapp, A.K.; Kratz, T.K.; Ohman, M.D.; et al. Long-Term Trends in Ecological Systems: A Basis for Understanding Responses to Global Change; Technical Bulletin 1931; U.S. Department of Agriculture: Washington, DC, USA, 2013.
- Yoccoz, N.G. Ecological Monitoring. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2012. [Google Scholar]
- Cairns, J., Jr.; Pratt, J.R. A history of biological monitoring using benthic macroinvertebrates. In Freshwater Biomonitoring and Benthic Macroinvertebrates; Rosenberg, D.M., Resh, V.H., Eds.; Chapman & Hall: London, UK, 1993; pp. 10–27. [Google Scholar]
- Siddig, A.A.H.; Ellison, A.M.; Ochs, A.; Villar-Leeman, C.; Lau, M.K. How do ecologists select and use indicator species to monitor ecological change? Insights from 14 years of publication in Ecological Indicators. Ecol. Indic. 2016, 60, 223–230. [Google Scholar] [CrossRef]
- Bartell, S.M. Biomarkers, Bioindicators, and Ecological Risk Assessment—A Brief Review and Evaluation. Environ. Bioindic. 2006, 1, 60–73. [Google Scholar] [CrossRef]
- Burger, J. Bioindicators: Types, Development, and Use in Ecological Assessment and Research. Environ. Bioindic. 2006, 1, 22–39. [Google Scholar] [CrossRef]
- Klinka, K. Indicator Plants of Coastal British Columbia; University of British Columbia Press: Vancouver, BC, Canada, 1989; ISBN 0774803215. [Google Scholar]
- Welsh, H.H.; Droege, S. A case for using plethodontid salamanders for monitoring biodiversity and ecosystem integrity of North American forests. Conserv. Biol. 2001, 15, 558–569. [Google Scholar] [CrossRef]
- Price, S.J.; Howe, R.W.; Hanowski, J.M.; Regal, R.R.; Niemi, G.J.; Smith, C.R. Are Anurans of Great Lakes Coastal Wetlands Reliable Indicators of Ecological Condition? J. Great Lakes Res. 2007, 33, 211–223. [Google Scholar] [CrossRef]
- Welsh, H.H.; Hodgson, G.R. Woodland salamanders as metrics of forest ecosystem recovery: A case study from California’s redwoods. Ecosphere 2013, 4, art59. [Google Scholar] [CrossRef]
- Furness, R.W.; Greenwood, J.J.D. Birds as Monitors of Environmental Change; Springer: Dordrecht, The Netherlands, 1993; ISBN 9789401513227. [Google Scholar]
- Greenwood, J. Birds as biomonitors: Principles and practice. Bird Census News 2004, 13, 1–10. [Google Scholar]
- Jenkins, J.A. Fish Bioindicators of Ecosystem Condition at the Calcasieu Estuary, Louisiana; USGS Open-File Report 2004–1323; Louisiana National Wetlands Research Center: Lafayette, LA, USA, 2004.
- Pearce, J.; Venier, L. Small mammals as bioindicators of sustainable boreal forest management. For. Ecol. Manag. 2005, 208, 153–175. [Google Scholar] [CrossRef]
- McGEOGH, M.A. The selection, testing and application of terrestrial insects as bioindicators. Biol. Rev. 2007, 73, 181–201. [Google Scholar] [CrossRef]
- Heino, J. Taxonomic surrogacy, numerical resolution and responses of stream macroinvertebrate communities to ecological gradients: Are the inferences transferable among regions? Ecol. Indic. 2014, 36, 186–194. [Google Scholar] [CrossRef]
- Chen, K.; Hughes, R.M.; Xu, S.; Zhang, J.; Cai, D.; Wang, B. Evaluating performance of macroinvertebrate-based adjusted and unadjusted multi-metric indices (MMI) using multi-season and multi-year samples. Ecol. Indic. 2014, 36, 142–151. [Google Scholar] [CrossRef]
- Logar, R.M.; Vodovnik, M. The Applications of Microbes in Environmental Monitoring. In Communicating Current Research Educational Topics Trends in Applied Microbiology; Mèndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2007. [Google Scholar]
- Caruso, G. Microbes and their use as Indicators of Pollution. J. Pollut. Eff. Control 2013, 1, 1–3. [Google Scholar] [CrossRef]
- Carignan, V.; Villard, M.-A. Selecting Indicator Species to Monitor Ecological Integrity: A Review. Environ. Monit. Assess. 2002, 78, 45–61. [Google Scholar] [CrossRef]
- NOSS, R.F. Indicators for Monitoring Biodiversity: A Hierarchical Approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Dale, V.H.; Beyeler, S.C. Challenges in the development and use of ecological indicators. Ecol. Indic. 2001, 1, 3–10. [Google Scholar] [CrossRef]
- U.S. EPA. Biological Assessments and Criteria: Crucial Components of Water Quality Programs; United States Environmental Protection Agency, Office of Water: Washington, DC, USA, 2004.
- Trexler, J.C.; Gottlieb, A.D.; Harwell, M.C. Ecological indicators for system-wide assessment of the greater everglades ecosystem restoration program. Ecol. Indic. 2009, 9, S2–S16. [Google Scholar]
- Author, R.; Welsh, H.H.; Ollivier, L.M.; And, J.R. Stream Amphibians as Indicators of Ecosystem Stress: A Case Study from California’s Redwoods. Ecol. Appl. 1998, 8, 1118–1132. [Google Scholar]
- Mathewson, B. Salamanders in a changing environment on hemlock hill. Arnoldia 2007, 65, 19–25. [Google Scholar]
- Mathewson, B. The Relative Abundance of Eastern Red-Backed Salamanders in Eastern Hemlock-dominated and Mixed Deciduous Forests at Harvard Forest. Northeast. Nat. 2009, 16, 1–12. [Google Scholar] [CrossRef]
- Wade, A.S.I.; Barov, B.; Burfield, I.J.; Gregory, R.D.; Norris, K.; Vorisek, P.; Wu, T.; Butler, S.J. A Niche-Based Framework to Assess Current Monitoring of European Forest Birds and Guide Indicator Species’ Selection. PLoS ONE 2014, 9, e97217. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Likens, G.E. Direct Measurement Versus Surrogate Indicator Species for Evaluating Environmental Change and Biodiversity Loss. Ecosystems 2011, 14, 47–59. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of Biodiversity for Ecologically Sustainable Forest Management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Quinn, J.E.; Brandle, J.R.; Johnson, R.J.; Tyre, A.J. Application of detectability in the use of indicator species: A case study with birds. Ecol. Indic. 2011, 11, 1413–1418. [Google Scholar] [CrossRef]
- Urban, N.A.; Swihart, R.K.; Malloy, M.C.; Dunning, J.B. Improving selection of indicator species when detection is imperfect. Ecol. Indic. 2012, 15, 188–197. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species:the need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- U.S. EPA. Climate Change Effects on Stream and River Biological Indicators: A Preliminary Analysis (Final Report); EPA/600/R-07/085F; U.S. Environmental Protection Agency: Washington, DC, USA, 2008.
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Orwig, D.A.; Barker Plotkin, A.A.; Davidson, E.A.; Lux, H.; Savage, K.E.; Ellison, A.M. Foundation species loss affects vegetation structure more than ecosystem function in a northeastern USA forest. PeerJ 2013, 1, e41. [Google Scholar] [CrossRef]
- Burton, T.M.; Likens, G.E. Salamander Populations and Biomass in the Hubbard Brook Experimental Forest, New Hampshire. Copeia 1975, 1975, 541. [Google Scholar] [CrossRef]
- Siddig, A.A.H.; Ellison, A.M.; Mathewson, B.G. Assessing the impacts of the decline of Tsuga canadensis stands on two amphibian species in a New England forest. Ecosphere 2016, 7, e01574. [Google Scholar] [CrossRef]
- Foster, D.R.; Aber, J.D. Forests in Time: The Environmental Consequences of 1,000 Years of Change in New England; Yale University Press: London, UK, 2004; ISBN 0300115377. [Google Scholar]
- Likens, G.E.; Bormann, F.H.; Johnson, N.M.; Pierce, R.S. The Calcium, Magnesium, Potassium, and Sodium Budgets for a Small Forested Ecosystem. Ecology 1967, 48, 772–785. [Google Scholar] [CrossRef]
- Elliott, K.J.; Miniat, C.F. Herbaceous-layer diversity and tree seedling recruitment are enhanced following Rhododendron maximum shrub removal. For. Ecol. Manag. 2018, 430, 403–412. [Google Scholar] [CrossRef]
- Knoepp, J.D.; See, C.R.; Vose, J.M.; Miniat, C.F.; Clark, J.S. Total C and N Pools and Fluxes Vary with Time, Soil Temperature, and Moisture Along an Elevation, Precipitation, and Vegetation Gradient in Southern Appalachian Forests. Ecosystems 2018, 21, 1623–1638. [Google Scholar] [CrossRef]
- Smith, W.B.; Miles, P.D.; Perry, C.H.; Pugh, S.A. Forest Resources of the United States, 2007; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2017; Volume 78.
- Yamasaki, M.; DeGraaf, W.B.; Lanier, J. Wildlife habitat associations in Eastern Hemlock—Birds, smaller mammals, and forest carnivores. In United States Department of Agriculture General Technical Report 267, Proceedings of the Symposium on Sustainable Management of Hemlock Ecosystems in Eastern North America, Newton Square, PA, USA; McManus, K.A., Shields, K.S., Souto, D.R., Eds.; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Newton Square, PA, USA, 2000; pp. 135–143. [Google Scholar]
- Sackett, T.E.; Record, S.; Bewick, S.; Baiser, B.; Sanders, N.J.; Ellison, A.M. Response of macroarthropod assemblages to the loss of hemlock (Tsuga canadensis), a foundation species. Ecosphere 2011, 2, art74. [Google Scholar] [CrossRef]
- Snyder, C.D.; Young, J.A.; Lemarié, D.P.; Smith, D.R. Influence of eastern hemlock (Tsuga canadensis) forests on aquatic invertebrate assemblages in headwater streams. Can. J. Fish. Aquat. Sci. 2002, 59, 262–275. [Google Scholar] [CrossRef]
- Tingley, M.W.; Orwig, D.A.; Field, R.; Motzkin, G. Avian response to removal of a forest dominant: Consequences of hemlock woolly adelgid infestations. J. Biogeogr. 2002, 29, 1505–1516. [Google Scholar] [CrossRef]
- Ellison, A.M.; Chen, J.; Díaz, D.; Kammerer-Burnham, C.; Lau, M.C.F. Changes in Ant Community Structure and Composition Associated with Hemlock Decline in New England. In Proceedings of the 3rd Symposium on Hemlock Woolly Adelgid in the Eastern United States, Asheville, NC, USA, 1–3 February 2005; Onken, B., Reardon, R., Eds.; U.S. Department of Agriculture, U.S. Forest Service Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2005; pp. 280–289. [Google Scholar]
- Dilling, C.; Lambdin, P.; Grant, J.; Buck, L. Insect guild structure associated with eastern hemlock in the southern Appalachians. Environ. Entomol. 2007, 36, 1408–1414. [Google Scholar] [CrossRef]
- Mathewson, B.G. The Relative Abundance of the Juvenile Phase of the Eastern Red-Spotted Newt at Harvard Forest Prior to the Arrival of the Hemlock Woolly Adelgid. Southeast. Nat. 2014. [Google Scholar] [CrossRef]
- Rohr, J.R.; Mahan, C.G.; Kim, K.C. Response of arthropod biodiversity to foundation species declines: The case of the eastern hemlock. For. Ecol. Manag. 2009, 258, 1503–1510. [Google Scholar] [CrossRef]
- Siderhurst, L.A.; Griscom, H.P.; Hudy, M.; Bortolot, Z.J. Changes in light levels and stream temperatures with loss of eastern hemlock (Tsuga canadensis) at a southern Appalachian stream: Implications for brook trout. For. Ecol. Manag. 2010, 260, 1677–1688. [Google Scholar] [CrossRef]
- Mallis, R.E.; Rieske, L.K. Arboreal Spiders in Eastern Hemlock. Environ. Entomol. 2011, 40, 1378–1387. [Google Scholar] [CrossRef]
- Havill, N.P.; Montgomery, M.E.; Yu, G.; Shiyake, S.; Caccone, A.; Caccone, A. Mitochondrial DNA from Hemlock Woolly Adelgid (Hemiptera: Adelgidae) Suggests Cryptic Speciation and Pinpoints the Source of the Introduction to Eastern North America. Ann. Entomol. Soc. Am. 2006, 99, 195–203. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R.; Mausel, D.L. Landscape patterns of hemlock decline in New England due to the introduced hemlock woolly adelgid. J. Biogeogr. 2002, 29, 1475–1487. [Google Scholar] [CrossRef]
- Eschtruth, A.K.; Cleavitt, N.L.; Battles, J.J.; Evans, R.A.; Fahey, T.J. Vegetation dynamics in declining eastern hemlock stands: 9 years of forest response to hemlock woolly adelgid infestation. Can. J. For. Res. 2006, 36, 1435–1450. [Google Scholar] [CrossRef]
- Knoepp, J.D.; Vose, J.M.; Clinton, B.D.; Hunter, M.D. Hemlock Infestation and Mortality: Impacts on Nutrient Pools and Cycling in Appalachian Forests. Soil Sci. Soc. Am. J. 2011, 75, 1935–1945. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R. Forest Response to the Introduced Hemlock Woolly Adelgid in Southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Kendrick, J.A.; Ribbons, R.R.; Classen, A.T.; Ellison, A.M. Changes in canopy structure and ant assemblages affect soil ecosystem variables as a foundation species declines. Ecosphere 2015, 6, art77. [Google Scholar] [CrossRef]
- Foster, D.R.; Orwig, D.A. Preemptive and salvage harvesting of New England forests: When doing nothing is a viable alternative. Conserv. Biol. 2006, 20, 959–970. [Google Scholar] [CrossRef]
- Ellison, A.M.; Barker-Plotkin, A.A.; Foster, D.R.; Orwig, D.A. Experimentally testing the role of foundation species in forests: The Harvard Forest Hemlock Removal Experiment. Methods Ecol. Evolut. 2010, 1, 168–179. [Google Scholar] [CrossRef]
- Orwig, D.A.; Cobb, R.C.; D’amato, A.W.; Kizlinski, M.L.; Foster, D.R. Multi-year ecosystem response to hemlock woolly adelgid infestation in southern New England forests. Can. J. For. Res. 2008, 38, 834–843. [Google Scholar] [CrossRef]
- Orwig, D.A.; Thompson, J.R.; Povak, N.A.; Manner, M.; Niebyl, D.; Foster, D.R. A foundation tree at the precipice: Tsuga canadensis health after the arrival of Adelges tsugae in central New England. Ecosphere 2012, 3, art10. [Google Scholar] [CrossRef]
- Ford, C.R.; Elliott, K.J.; Clinton, B.D.; Kloeppel, B.D.; Vose, J.M. Forest dynamics following eastern hemlock mortality in the southern Appalachians. Oikos 2012, 121, 523–536. [Google Scholar] [CrossRef]
- Siddig, A.A.; Ellison, A.M.; Jackson, S. Calibrating abundance indices with population size estimators of red back salamanders (Plethodon cinereus) in a New England forest. PeerJ 2015, 3, e952. [Google Scholar] [CrossRef] [PubMed]
- Royle, J.A. N-Mixture Models for Estimating Population Size from Spatially Replicated Counts. Biometrics 2004, 60, 108–115. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 1 October 2018).
- Hairston, N.G. Community Ecology and Salamander Guilds/Monograph; Cambridge University Press: Cambridge, UK, 2009; ISBN 9780521310819. [Google Scholar]
- Hesed, K.M. Uncovering Salamander Ecology: A Review of Coverboard Design. J. Herpetol. 2012, 46, 442–450. [Google Scholar] [CrossRef]
- Welsh, H.H.; Pope, K.L.; Wheeler, C.A. Using multiple metrics to assess the effects of forest succession on population status: A comparative study of two terrestrial salamanders in the US Pacific Northwest. Biol. Conserv. 2008, 141, 1149–1160. [Google Scholar] [CrossRef]
- Wyman, R.L. Experimental assessment of salamanders as predators of detrital food webs: Effects on invertebrates, decomposition and the carbon cycle. Biodivers. Conserv. 1998, 7, 641–650. [Google Scholar] [CrossRef]
- Best, M.L.; Welsh, H.H., Jr. The trophic role of a forest salamander: Impacts on invertebrates, leaf litter retention, and the humification process. Ecosphere 2014, 5, art16. [Google Scholar] [CrossRef]
- Hocking, D.J.; Babbitt, K.J. Effects of Red-Backed Salamanders on Ecosystem Functions. PLoS ONE 2014, 9, e86854. [Google Scholar] [CrossRef] [PubMed]
- Eaton, S.W. Wild Turkey (Meleagris gallopavo). In The Birds of North America; Poole, A., Stettenheim, P., Gill, F., Eds.; American Ornithologists’ Union: Chicago, IL, USA, 1992; pp. 1–27. [Google Scholar]
- Arnold, S.J. A Quantitative Approach to Antipredator Performance: Salamander Defense against Snake Attack. Copeia 1982, 1982, 247. [Google Scholar] [CrossRef]
- Lowe, W.H.; Bolger, D.T. Local and Landscape-Scale Predictors of Salamander Abundance in New Hampshire Headwater Streams. Conserv. Biol. 2002, 16, 183–193. [Google Scholar] [CrossRef]
- Grant, E.H.C.; Green, L.E.; Lowe, W.H. Salamander occupancy in headwater stream networks. Freshw. Biol. 2009, 54, 1370–1378. [Google Scholar] [CrossRef]
- Lowe, W.H. Climate change is linked to long-term decline in a stream salamander. Biol. Conserv. 2012, 145, 48–53. [Google Scholar] [CrossRef]
- Stoddard, M.A.; Hayes, J.P.; Stoddard, A. The Influence of Forest Management on Headwater Stream Amphibians At Multiple Spatial Scales. America 2012, 15, 811–823. [Google Scholar]
- Cecala, K.K.; Lowe, W.H.; Maerz, J.C. Riparian disturbance restricts in-stream movement of salamanders. Freshw. Biol. 2014, 59, 2354–2364. [Google Scholar] [CrossRef]
- Vernick, S. Night of the spotted salamanders. Highlights 2012 2012, 4, 10–11. [Google Scholar]
- Price, S.J.; Dorcas, M.E. The Carolina Herp Atlas: An Online, Citizen-Science Approach to Document Amphibian and Reptile Occurrences. Herpetol. Conserv. Biol. 2011, 6, 287–296. [Google Scholar]



{kind=link}
{kind=link}
| Red Backs/m2 | Red Efts/m2 | |||||
|---|---|---|---|---|---|---|
| Experimental Treatment | 2004 | 2005 | 2013 | 2014 | 2004 | 2014 |
| Hemlock control | 1.64 (0.13) | 0.2 (0.03) | 1.28 (0.01) | 1.68 (0.006) | 0.05 (0.004) | 0.04 (0.001) |
| Girdling | 2.32 (0.19) | 0.6 (0.07) | 0.28 (0.006) | 0.32 (0.003) | 0.06 (0.005) | 0.02 (0.0007) |
| Logging | 2 (0.16) | 0.12 (0.03) | 0.32 (0.007) | 0.08 (0.003) | 0.08 (0.004) | 0.02 (0.002) |
| Hardwood control | 1.16 (0.007) | 0.2 (0.03) | 0.92 (0.01) | 0.8 (0.005) | 0.04 (0.002) | 0.03 (0.003) |
| Forest Type | Species | Estimated Population Size (Individuals Per m−2) | Specificity | Occupancy | Naïve IndVal | Robust IndVal |
|---|---|---|---|---|---|---|
| Hemlock | Red back | 13.8 | 0.99 | 1.00 | 0.68 | 0.99 |
| Red eft | 7.4 | 0.77 | 1.00 | 0.47 | 0.77 | |
| Hardwood | Red back | 0.2 | 0.01 | 0.63 | 0.14 | 0.01 |
| Red eft | 2.2 | 0.32 | 1.00 | 0.47 | 0.23 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddig, A.A.H.; Ochs, A.; Ellison, A.M. Do Terrestrial Salamanders Indicate Ecosystem Changes in New England Forests? Forests 2019, 10, 154. https://doi.org/10.3390/f10020154
Siddig AAH, Ochs A, Ellison AM. Do Terrestrial Salamanders Indicate Ecosystem Changes in New England Forests? Forests. 2019; 10(2):154. https://doi.org/10.3390/f10020154
Chicago/Turabian StyleSiddig, Ahmed A. H., Alison Ochs, and Aaron M. Ellison. 2019. "Do Terrestrial Salamanders Indicate Ecosystem Changes in New England Forests?" Forests 10, no. 2: 154. https://doi.org/10.3390/f10020154
APA StyleSiddig, A. A. H., Ochs, A., & Ellison, A. M. (2019). Do Terrestrial Salamanders Indicate Ecosystem Changes in New England Forests? Forests, 10(2), 154. https://doi.org/10.3390/f10020154