1. Introduction
Many processes lead to periodic pulses of mortality in forest stands and landscapes which can result in changes of forest structure and function that are evident for decades to centuries [
1]. These disturbance effects are dependent on the spatial scale examined, as well as the spatial heterogeneity of the disturbance: the death of a few trees can lead to significant change at the scale of 100 m
2, but may be viewed as part of “normal” mortality at the scale of more than 1 ha. Different disturbances may lead to similar cumulative mortality over broad regions, but those with diffuse spatial patterns will have less-apparent broad-scale impact on structure and function than those that are concentrated. The realization that disturbance expresses itself differently depending on hierarchical level has meant that perspectives on disturbance have evolved from the simple idealized case in which these disturbances completely “reset” the system to one in which disturbances create heterogeneous mosaics of structural and functional change [
2].
The coastal forests of Oregon provide an example of the heterogeneous, hierarchical mosaic of disturbance. In this region, fire and wind are the primary processes causing major episodes of mortality that lead to long-term structural and functional change. In some ways, fire and wind disturbance are opposites in terms of temporal and spatial patterns. Fires are infrequent to rare in this region given the wet climate, with an approximate return interval of 230 years [
3], but when climatic and weather conditions are favorable, fires do occur, and they are often severe because of the high accumulation of fuels [
4]. In contrast, winds of sufficient velocity to fell trees occur most every year in the region, while severe wind events with hurricane-level winds in excess of 119 km per hour (74 miles per hour), occur on average once every 25 years for any given location [
5]. Fire disturbance can be quite extensive, but even historically large, severe fires such as the Tillamook and Nestucca burns modified only a small fraction (≈6%) of the Oregon Coast Range landscape [
6,
7]. In comparison, high wind events, such as the 1962 Columbus Day Storm, can impact areas larger than the Oregon coastal region [
8,
9], even though patches of high mortality are often localized [
10]. Thus, despite influencing over 20 times more area than the 1933 Tillamook burn, the 1962 Columbus Day Storm killed a similar volume of timber: 11 billion board feet versus 10 billion board feet, respectively [
6,
7,
8].
The broad-scale spatial and temporal patterns created by these two types of disturbances in coastal Oregon have been examined by Harcombe et al. [
11], as has the importance of wind-related mortality on forest growth [
12,
13]. However, there has not been a detailed examination of how repeated wind disturbance has influenced these forests at a fine scale over long time periods. Indeed, as noted by Mitchell [
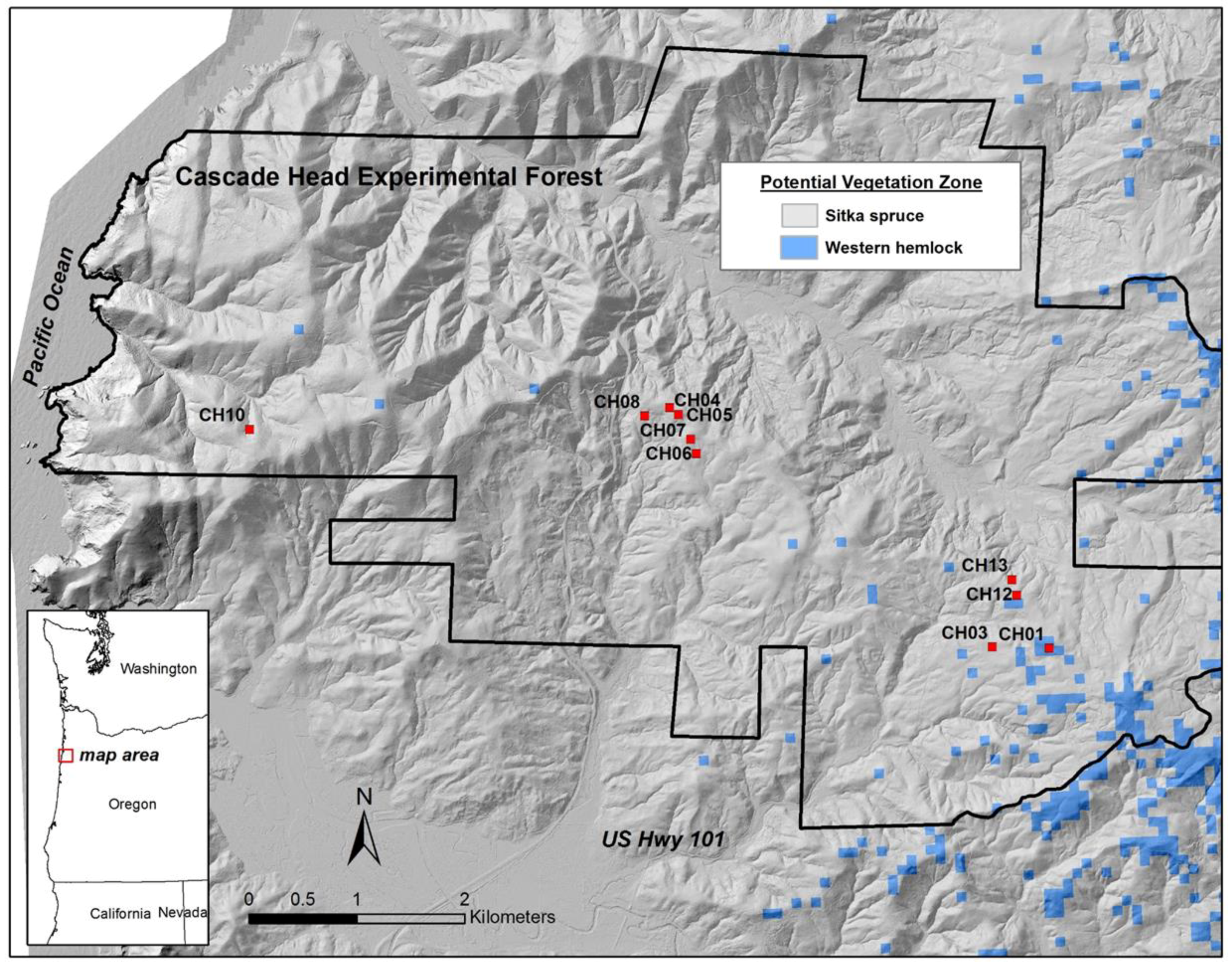
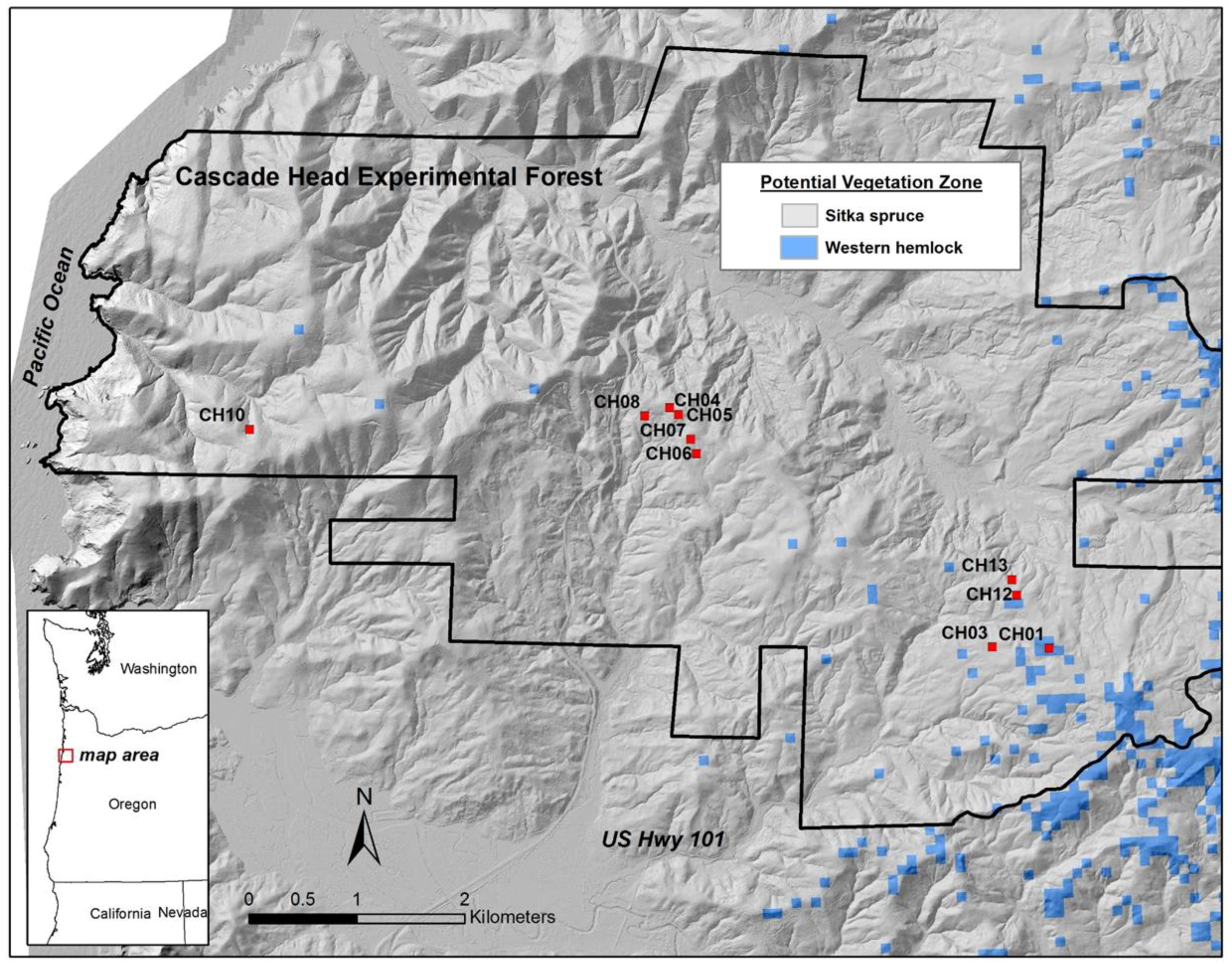
14], “the long-term and cumulative impacts of recurrent windthrow are often over looked by scientists and managers”. We therefore conducted a descriptive study using a set of ten, 0.4-ha plots established in 1935 and measured periodically since then to address the following questions regarding wind at the scale of the individual plot (0.4 ha) and all plots (4.0 ha):
Is there a correspondence between major windstorms, defined as those with hurricane-strength winds, and the occurrence of major tree mortality periods?
What is the magnitude of major tree mortality periods and at what magnitudes are there net losses in tree density and biomass?
What are the specific mechanisms of overall (e.g., suppression) and wind-related mortality (e.g., breakage, uprooting, and crushing), and how many trees involved in the latter are associated with “immediate” versus delayed mortality?
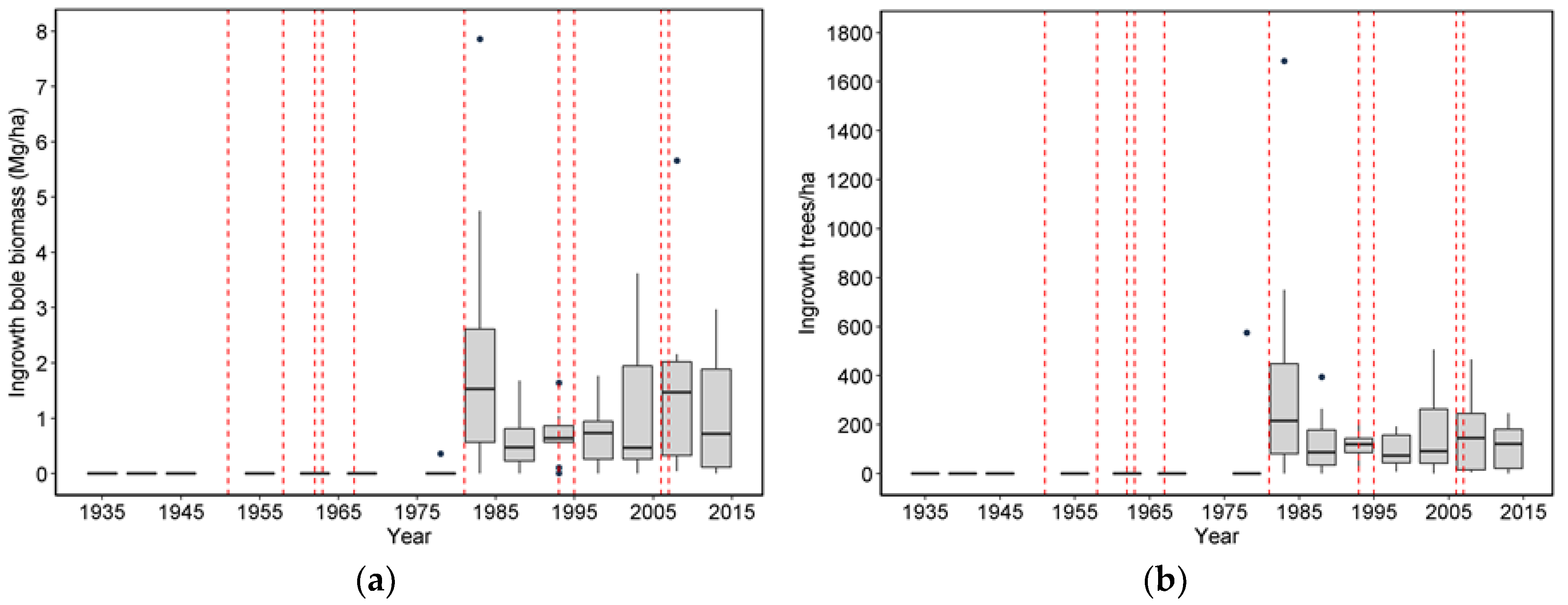
To what degree are pulses of tree ingrowth related to major tree mortality periods, and what is the time lag in tree ingrowth after a major tree mortality period?
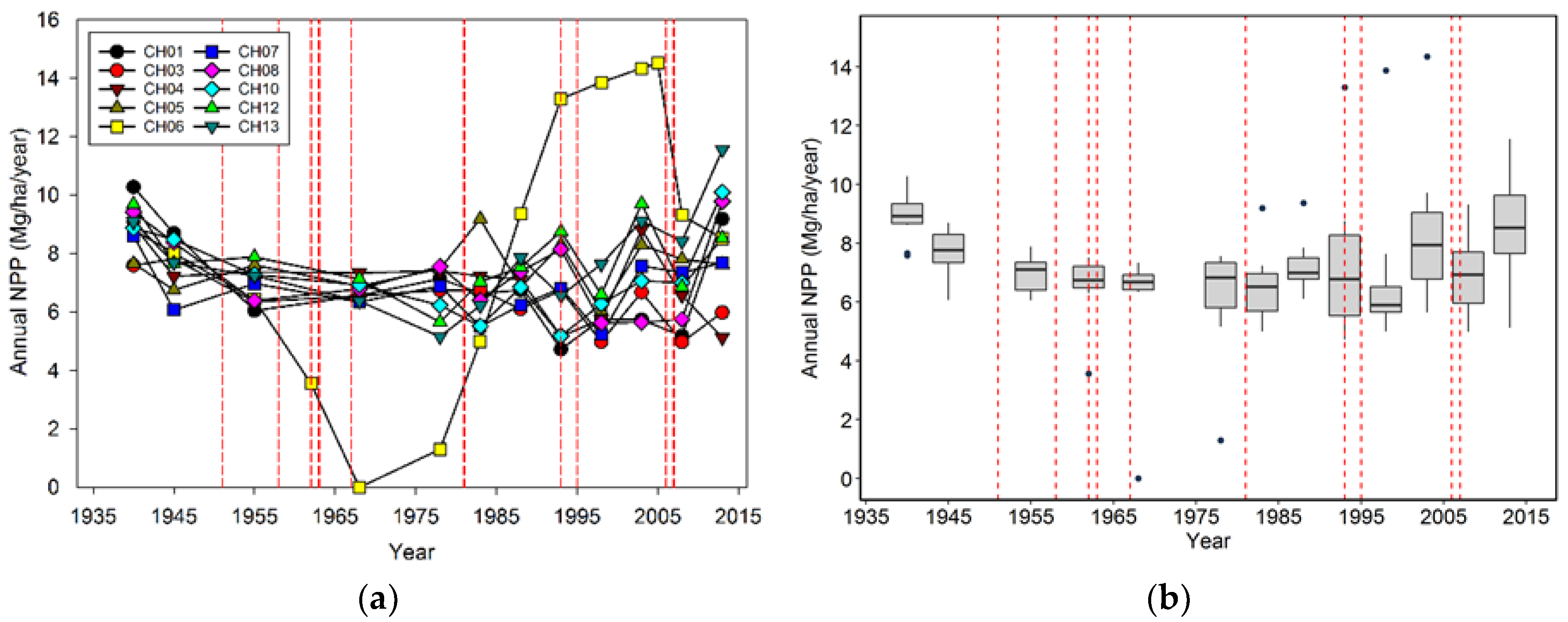
How does net primary productivity (NPP) respond to the magnitude of mortality?
4. Discussion
Wind is an important disturbance agent in many forested regions of the globe, though its effects on forest structure vary with the frequency, intensity and duration of each event, as well as the interaction with topography and antecedent conditions such as soil moisture [
14,
20]. The majority of research on wind-related effects on forests has focused on the period immediately after an event, characterizing damage and mortality and how these change the species composition and structure of stands over the short-term (<10 years). Few studies have examined subsequent changes over the long term and fewer yet have examined the effects of multiple wind disturbance events [
14]. Our study is one of the few that has examined multiple wind disturbance events over multiple decades (i.e., 78 years) in the same plots. Inference from our findings to the whole study area is limited, because the plots were not located randomly or systematically across the study area; however, they do provide some sense of the degree to which the effects of wind disturbance are synchronous versus asynchronous over a broad area. Thus, our results potentially provide new insights into the impact of wind disturbance on forests over time and at multiple spatial scales.
Although studies of wind disturbance generally focus on hurricanes in the eastern portion of North America, wind is also important in the Pacific Northwest, particularly along the Pacific coast and in the Coast Range where storms rival hurricanes in intensity and impact [
21]. In some Pacific Northwest forests, wind is the dominant disturbance because fires are extremely rare [
22]. The Cascades Mountains, the crest of which parallels the coast roughly 200 km inland from the coast, restrict the eastern extent of these storms. For example, little damage was observed from the 1962 Columbus Day Storm east of the Cascades [
8].
Wind disturbance has been characterized as spatially heterogeneous, creating patches of severely impacted forest that are mostly less than 2 ha in extent [
20,
23] but dispersed over a wide area. This is consistent with our observations, in which major wind events influenced plots somewhere in the study area, but rarely across all plots at once. Harcombe et al. [
11] examined a nearby forest landscape dominated by Sitka Spruce and western hemlock using aerial photography. They found that wind could create openings multiple ha in size, although even these were comprised of mixtures of high- and low-severity patches. They also found that openings from one event were often spatially adjacent to those created by previous or subsequent events. If these previous and subsequent events are in close temporal proximity, a larger, effectively homogeneous patch might be created. On the other hand, if wind disturbance events are separated by decades, landscape-level heterogeneity may be increased.
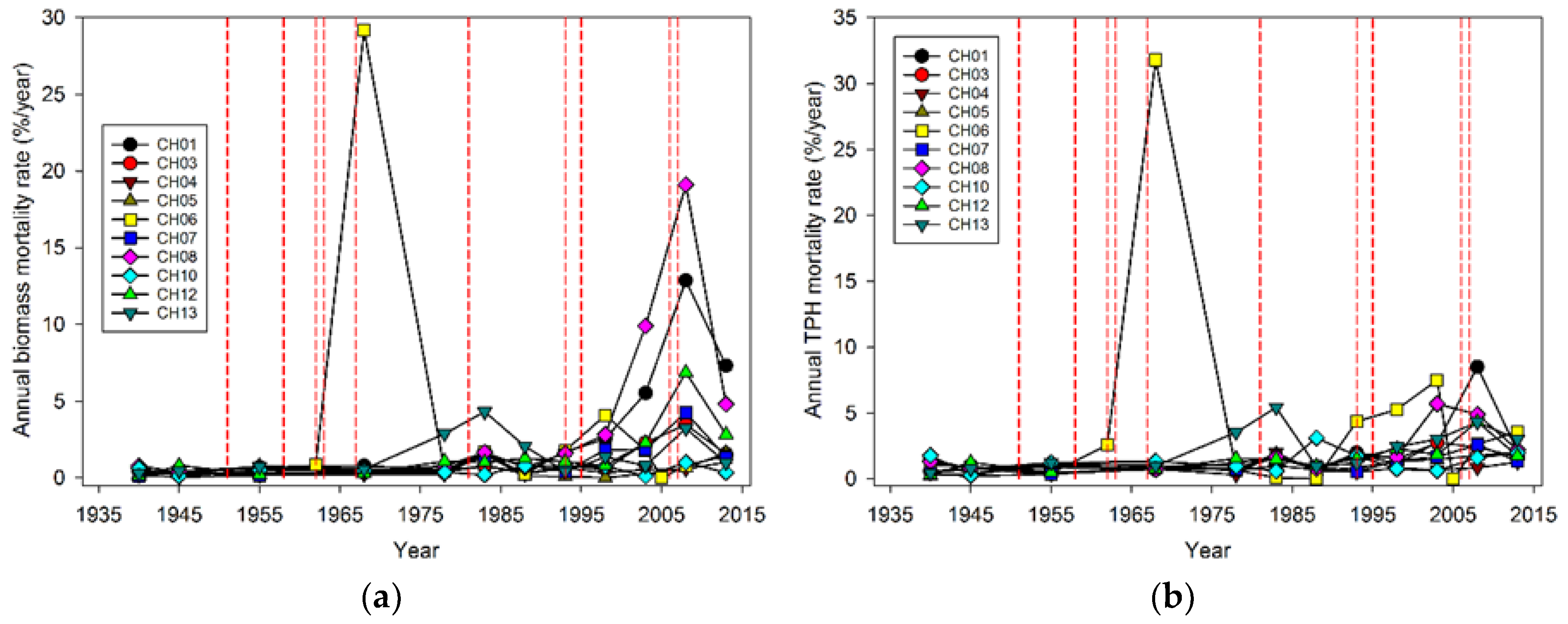
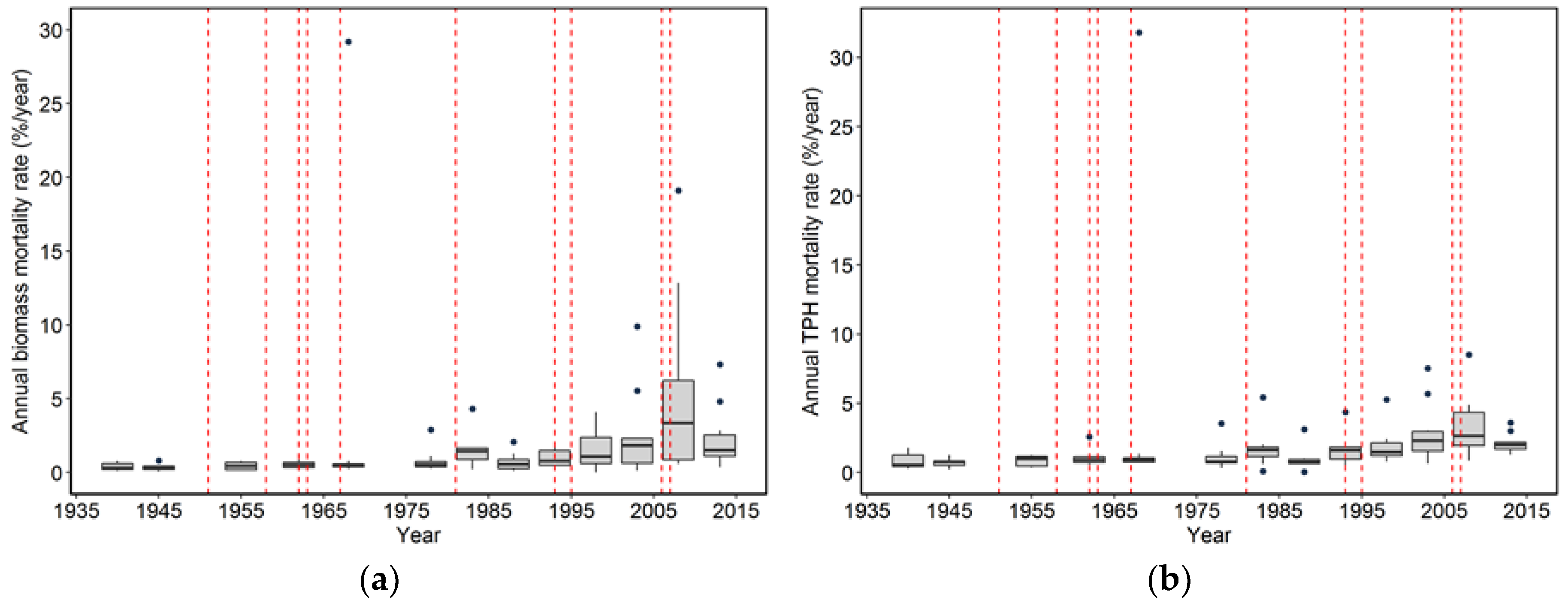
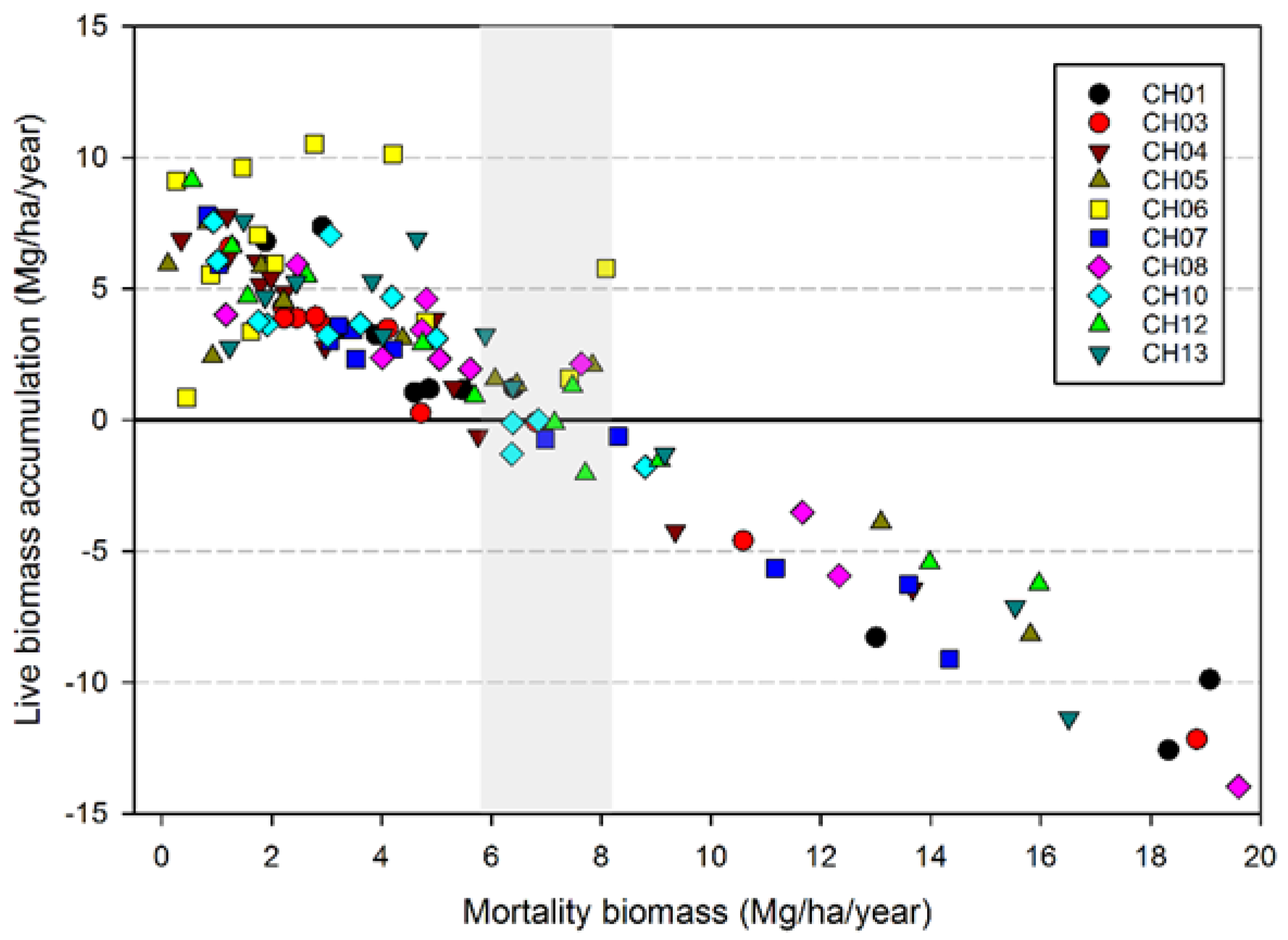
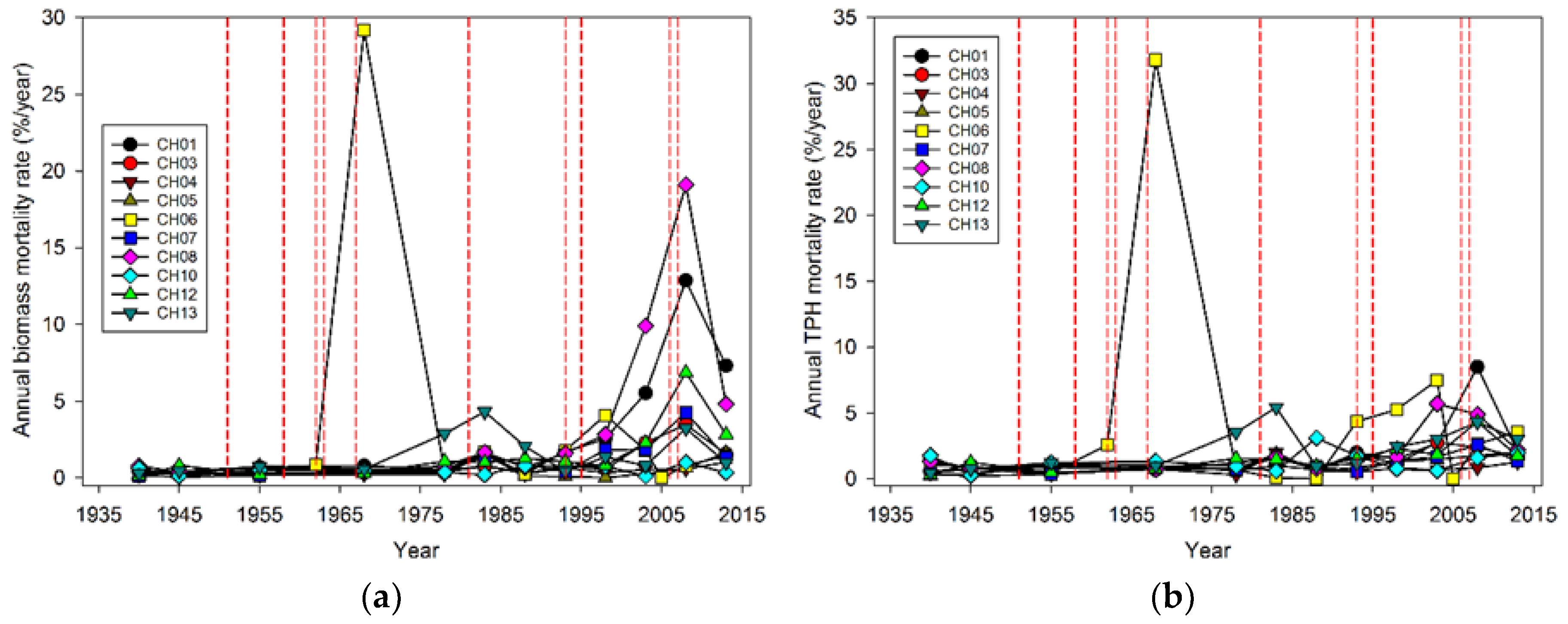
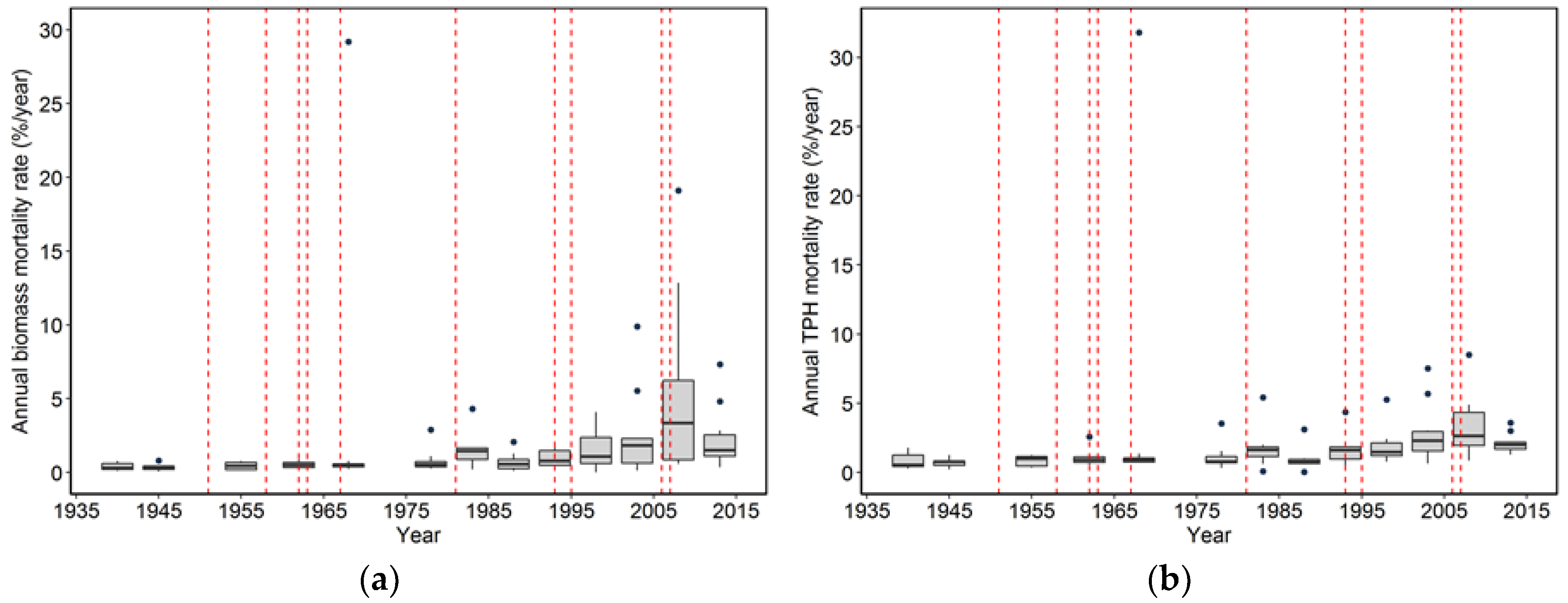
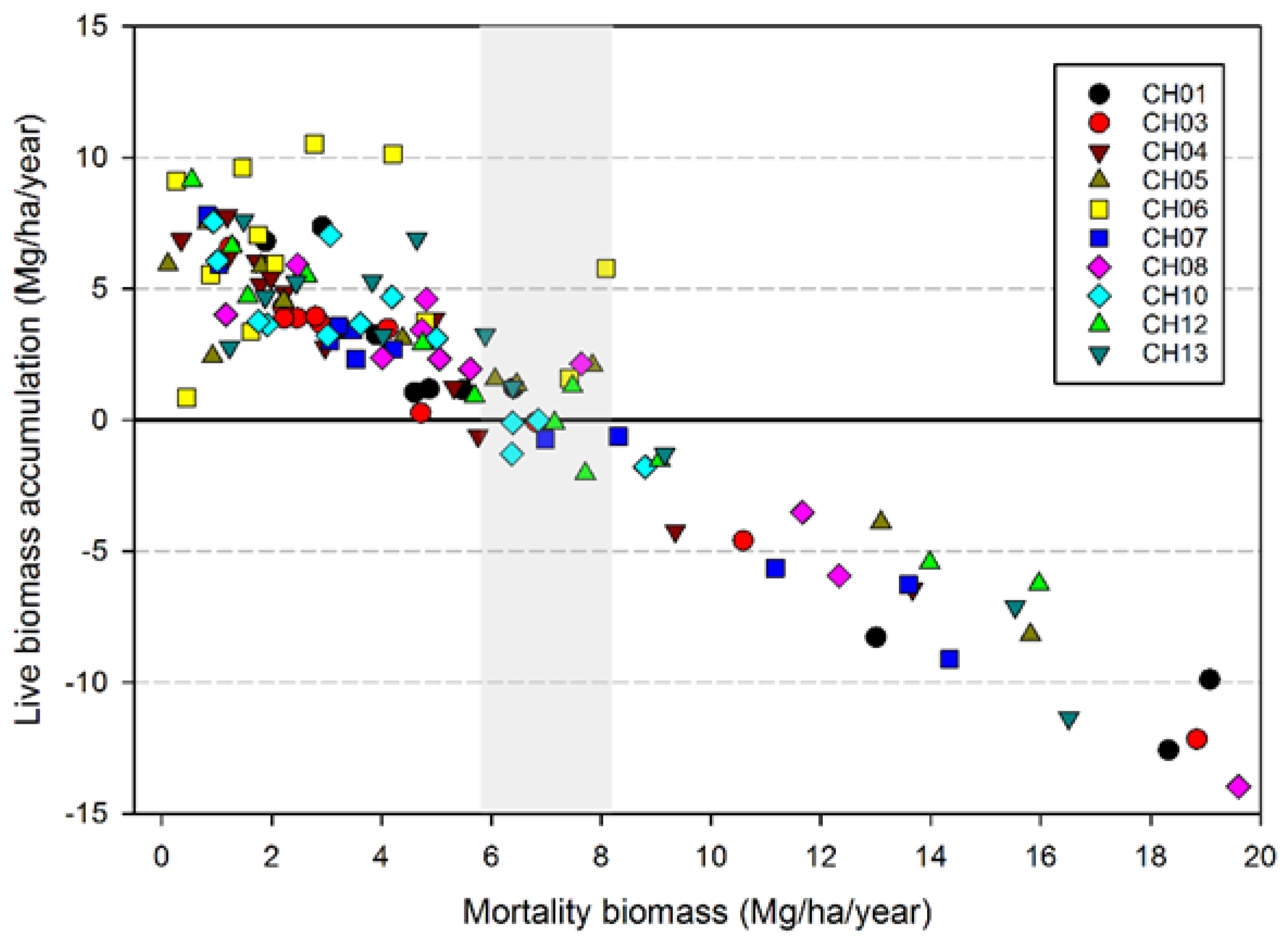
While the severity of wind disturbance depends on the scale being examined, Foster and Boose [
23] observed a maximum mortality of 75% for areas ranging in size from 0.15 to 0.6 ha. Our maximum estimate is 88% for an area of 0.4 ha (plot CH06) in the 1962 Columbus Day Storm, but most of the events impacting our plots caused far lower losses in terms of magnitude. Our estimate of the maximum magnitude of wind-related mortality may be too high given that salvage logging followed the wind disturbance in CH06, compounding losses in trees and biomass [
24]. However, it is likely that the majority of trees were sufficiently damaged in CH06 to have eventually died from wind-related effects on tree vigor such as snapped tops and crushing. The observation that wind-related mortality events rarely kill the majority of stems is consistent with the notion that wind disturbance along the Pacific coast is a very “inefficient” disturbance, meaning that with the exception of very small spatial extents, it does not cause anything approaching complete tree mortality [
20].
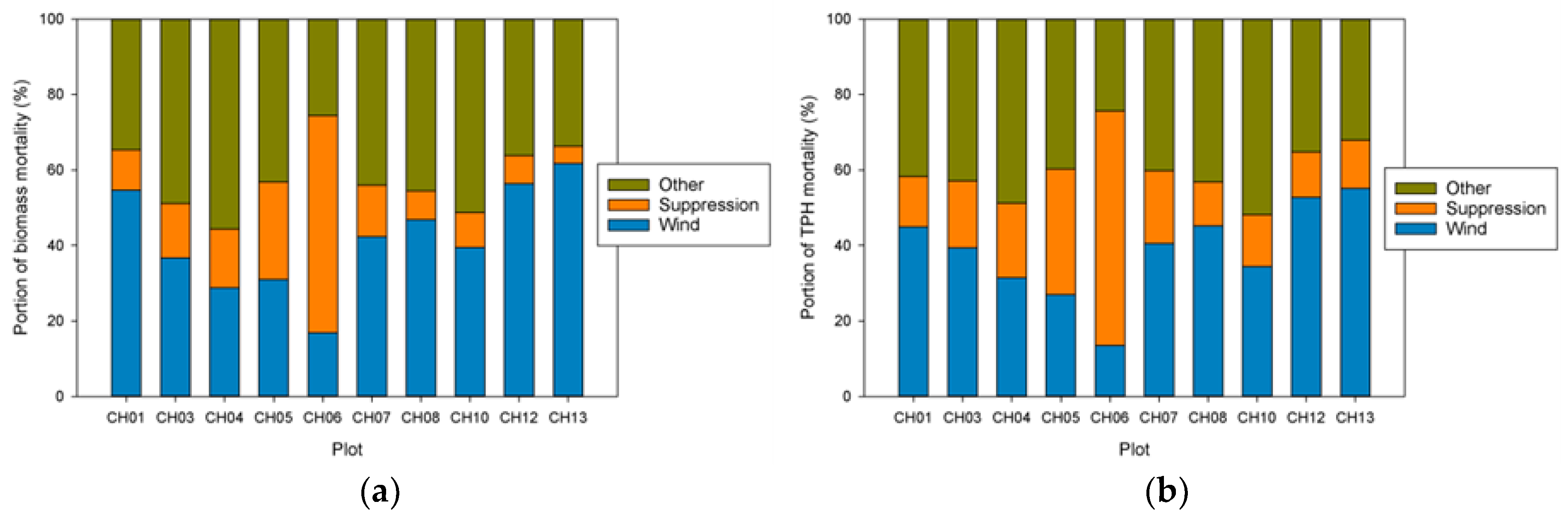
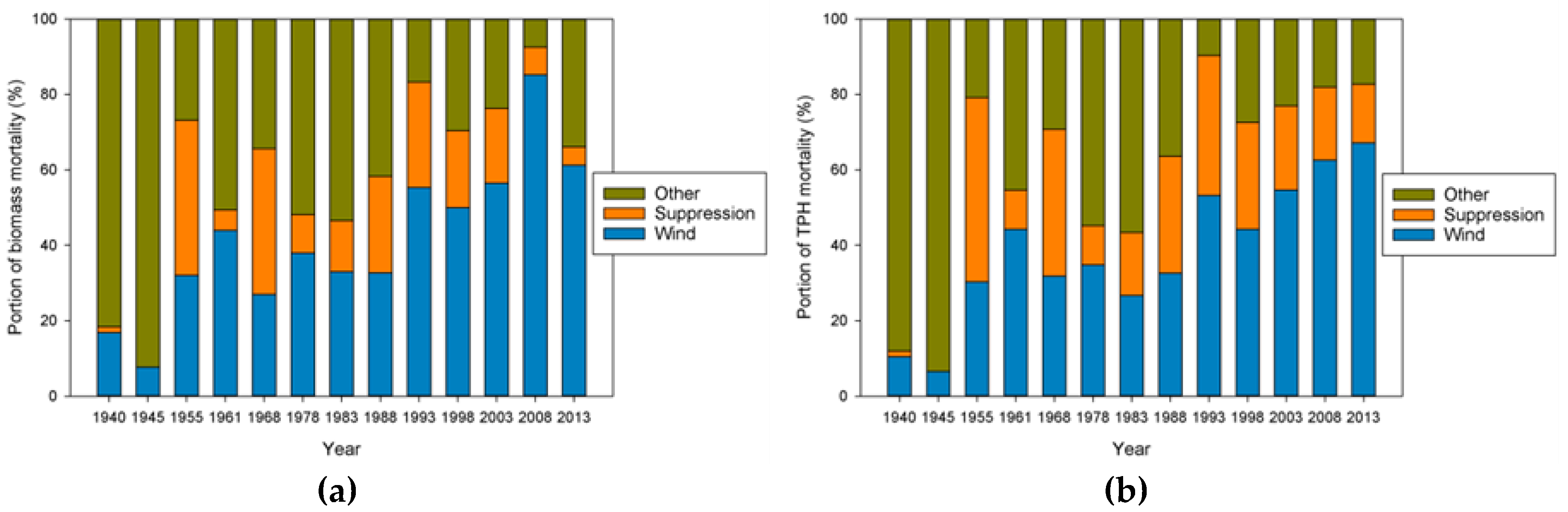
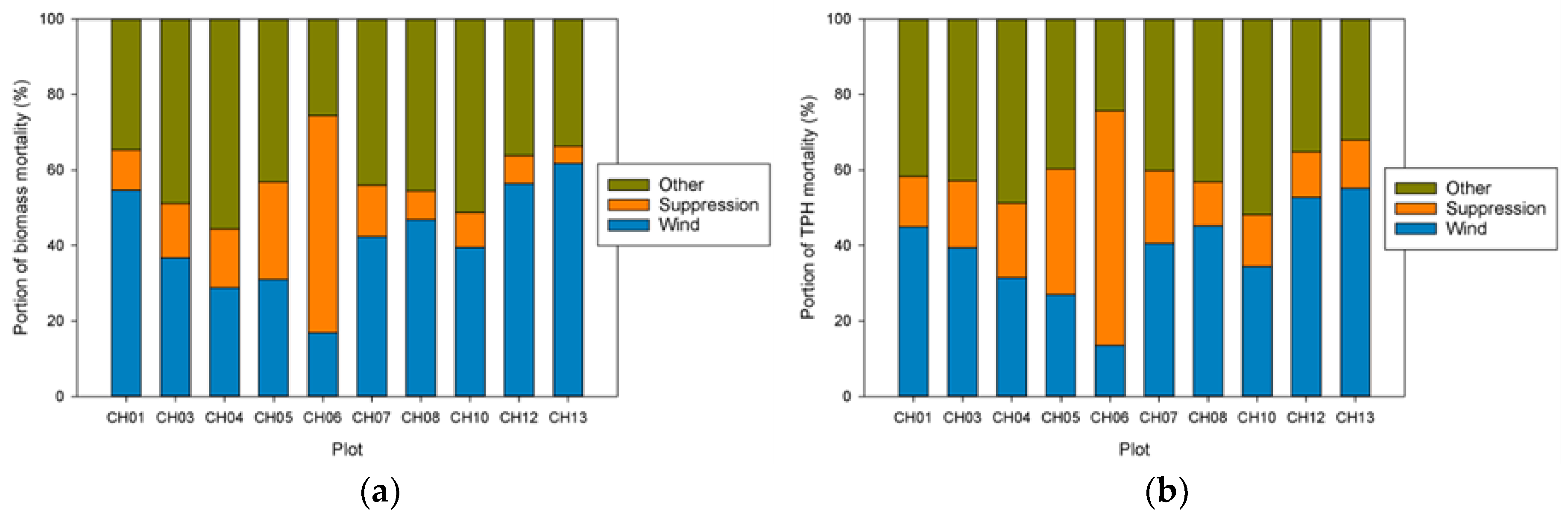
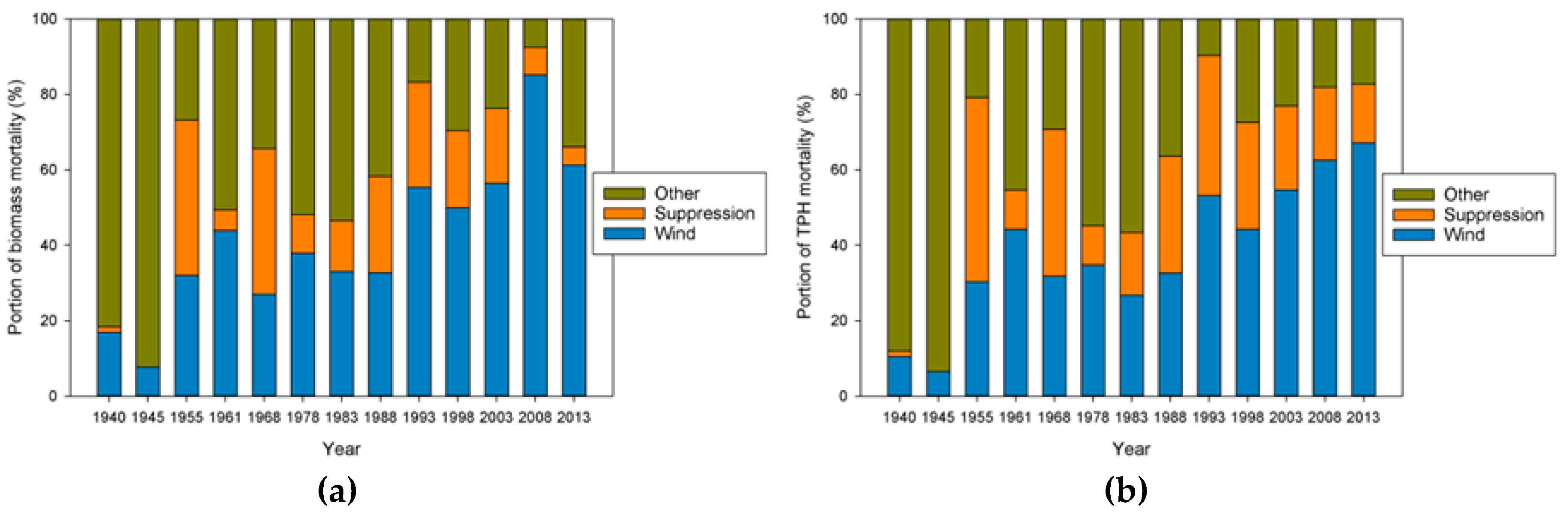
Our analysis focused on mortality losses of trees and biomass caused by wind. Nonetheless, there were other changes brought about by wind disturbance in these plots such as defoliation, branch losses, and damage to stems. We did not quantify defoliation or branch loss, and although both forms of damage occurred, we believe these were relatively small compared to what was observed in regions with broad-leaved species. This may be due to the fact that needle-shaped leaves are less resistant to the wind than broad ones and that the branches of excurrent tree forms (typical of gymnosperms) may be less exposed than those of decurrent forms (typical of angiosperms). It is also likely that the majority of branch loss in the coastal Northwest is associated either with stems snapping or wind-fallen stems impacting other stems. Our records indicate that 319 trees were damaged by wind between 1978 and 2013, as opposed to 1,233 trees that died directly from wind-related mortality. The majority of wind-related effects (80%) therefore resulted in “immediate” mortality. Of those trees that were damaged, 144 (about 45%) died within 35 years, which means that in these coastal Northwest forests only a small share (144 of 1552, or 9%) of the trees dying from wind are associated with indirect causes.
Ruth and Yoder [
25] observed that Sitka spruce and Douglas-fir were more wind-firm than western hemlock. We also observed species differences, with western hemlock having at least an order of magnitude higher mortality rate than Sitka spruce, depending on the time interval examined. We found that the effects of size, as indicated by DBH, were species dependent, even though the smallest DBH size class for both species had low wind-related mortality rates. Trees of small size are likely protected from the full force of the wind either by surrounding larger trees or boundary layer effects [
14,
20]. For Sitka spruce, the largest-diameter trees appeared to be more wind-firm than the intermediate-sized ones. For western hemlock, the wind-related mortality rate appeared to increase with DBH, although it is possible the few largest western hemlocks were more wind-firm than intermediate-sized ones. These findings on size-class responses to wind mortality (i.e., intermediate highest versus largest highest) have been observed in other studies [
20]. Moreover, although we reported the average effects by DBH size class in the results, the ranking of mortality rates by DBH class was not the same over time, suggesting that the effect of DBH on wind-related mortality varies over time (
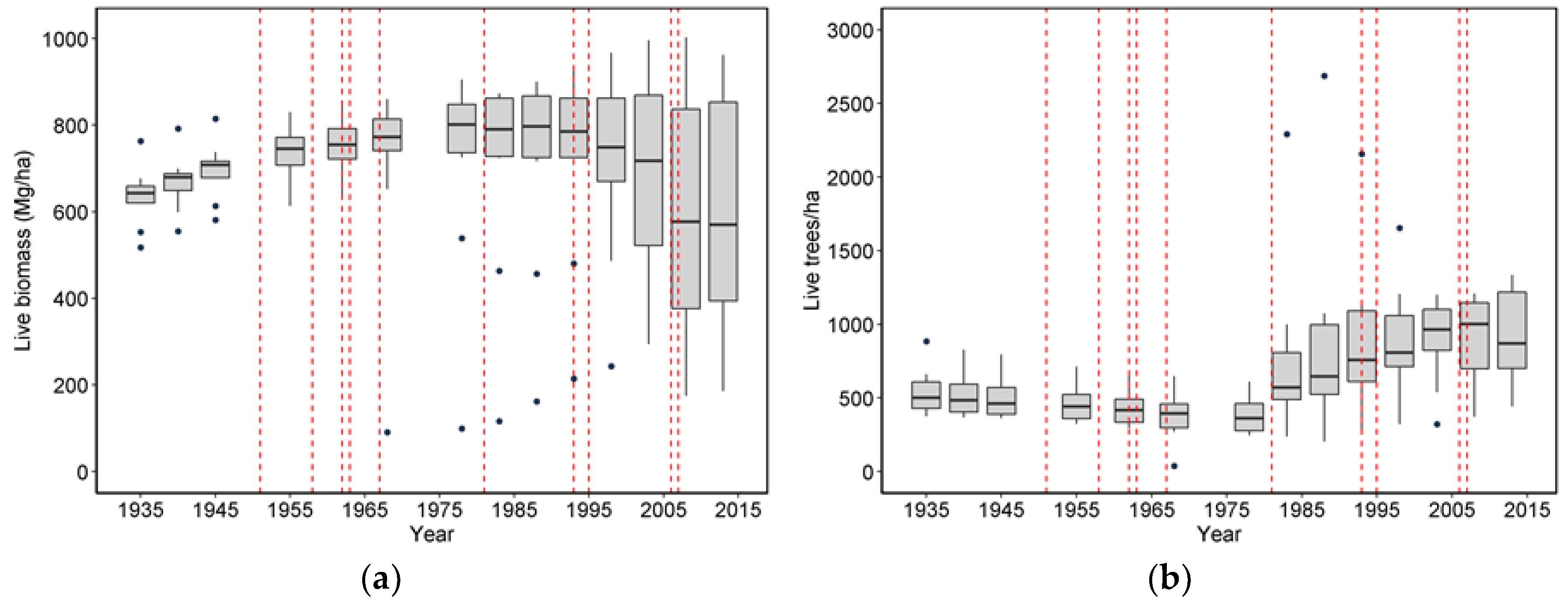
Figure S3). This means that while the smallest DBH class is likely to have the lowest wind-related mortality rate, any of the larger DBH size classes can have the highest rate depending on the features of a particular storm. Specifically, a low-intensity storm might preferentially kill the large trees with stems or roots compromised by decay, whereas the most intense storms might kill the majority of all large diameter stems regardless of condition.
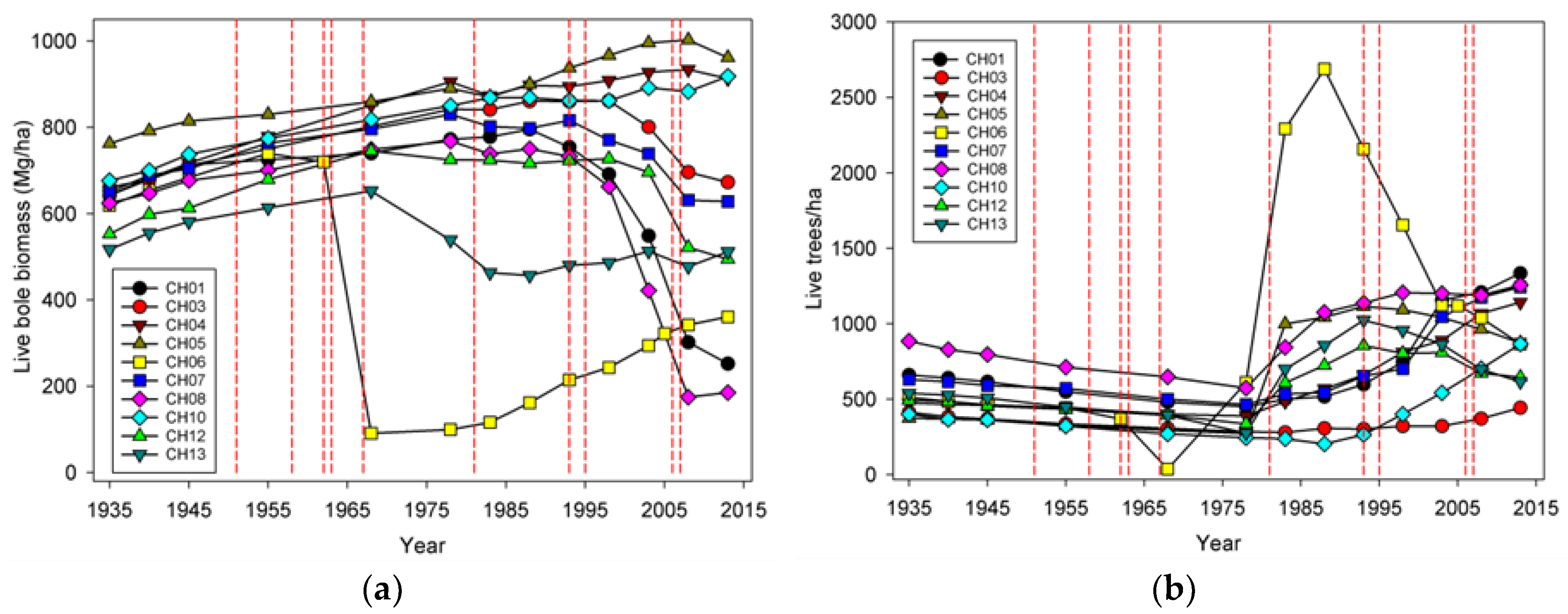
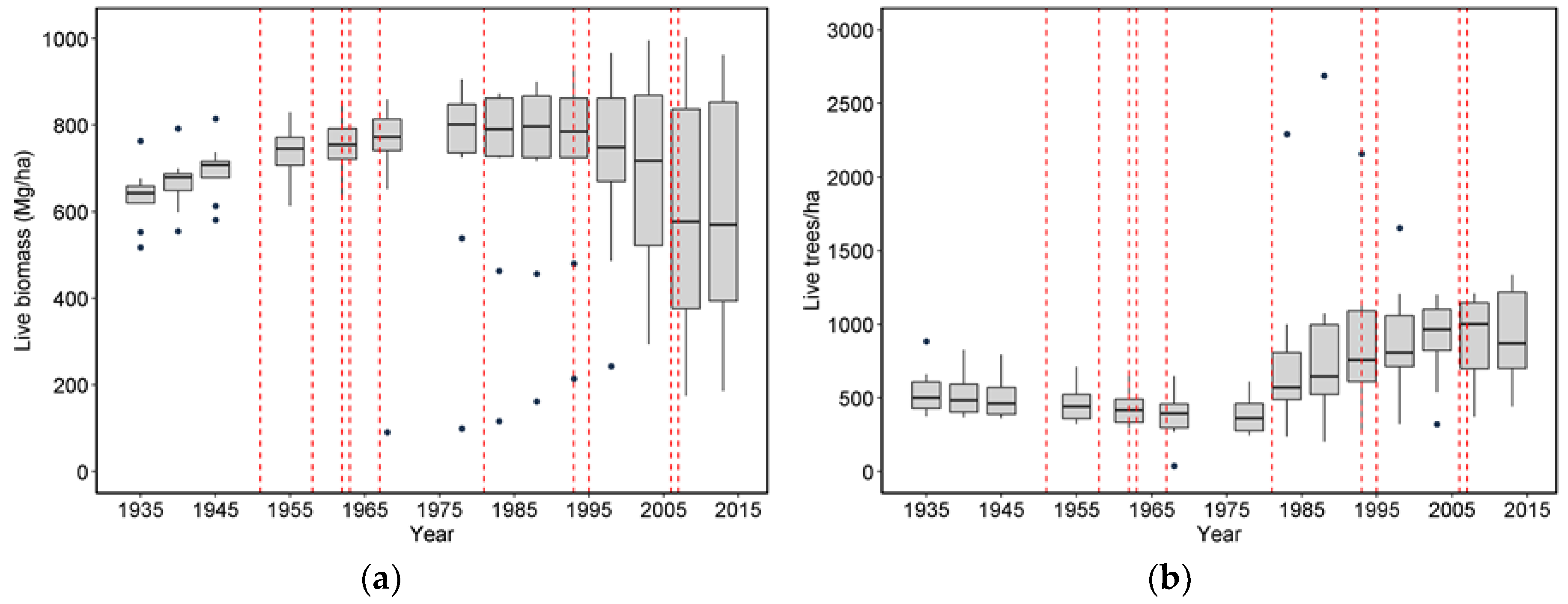
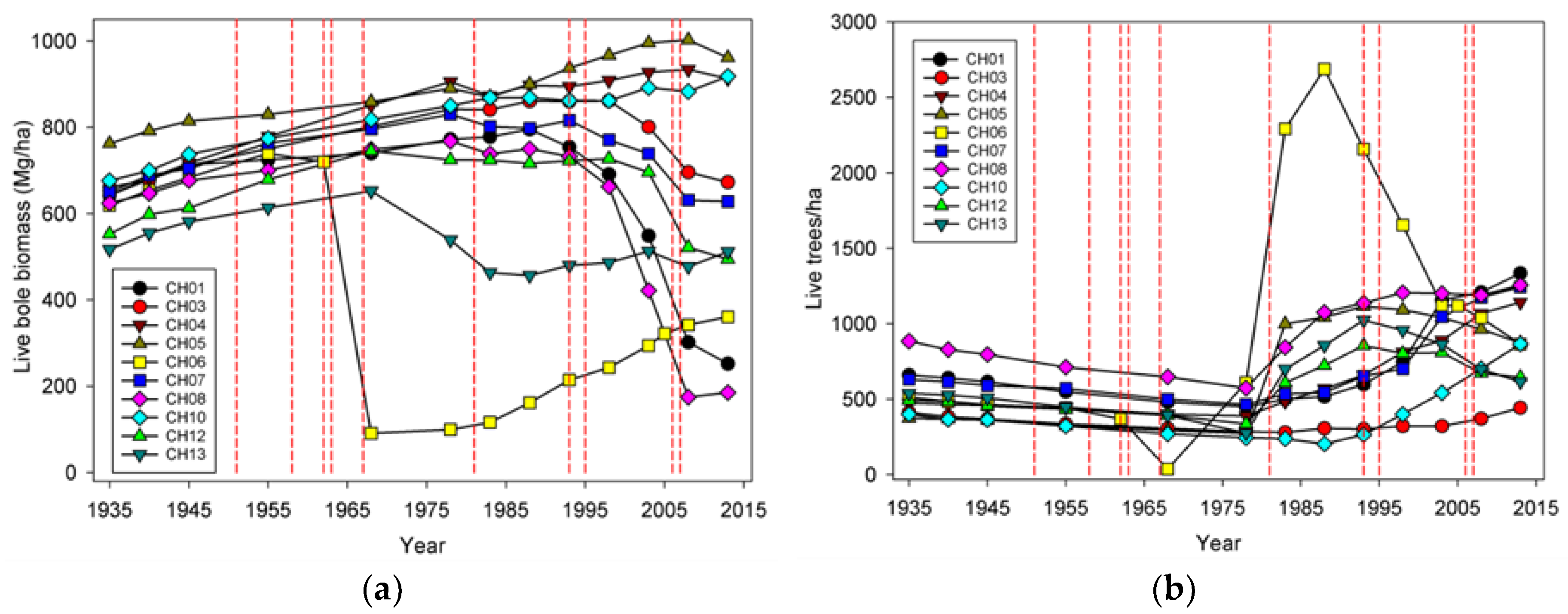
Wind-related mortality has become increasingly important in these plots as the dominant trees have grown and aged, from 85 years at the time of plot establishment (1935) to 163 years of age at last measurement (2013). Although none of the plots was severely impacted by wind until the forest reached an age of 112 years, it is likely that areas within this landscape were impacted at a younger age. The windstorm in 1951 did not seem to impact the plots we examined; however, there were stands in the Cascade Head Experimental Forest that were impacted by this storm, particularly along the boundaries of recent harvest areas [
25]. Moreover, CH06, which had a mortality event in 1962 that was severe enough to cause a new, even-aged cohort of trees to establish, also had a major wind-related mortality loss in 2007 effectively at an age of 45 years. While age is often correlated with the extent of wind damage and mortality, other structural features of the local topography and forest stand are likely more important [
20,
26]. In the case of CH06, the area impacted in 2007 was relatively open, suggesting wind could penetrate this part of the plot. Most of the other plots were relatively intact and evenly stocked by trees for many decades after plot establishment, and thus may have been less susceptible to wind disturbance until gaps began to develop at an older age. In the case of CH08 and CH13, which were impacted by multiple storm events, an earlier event created openings that likely enhanced the impact of subsequent events. This is similar, albeit at a smaller spatial scale, to the spreading of wind-related openings observed by Harcombe et al. [
11] and along cutting boundaries by Ruth and Yoder [
25].
Everham and Brokaw [
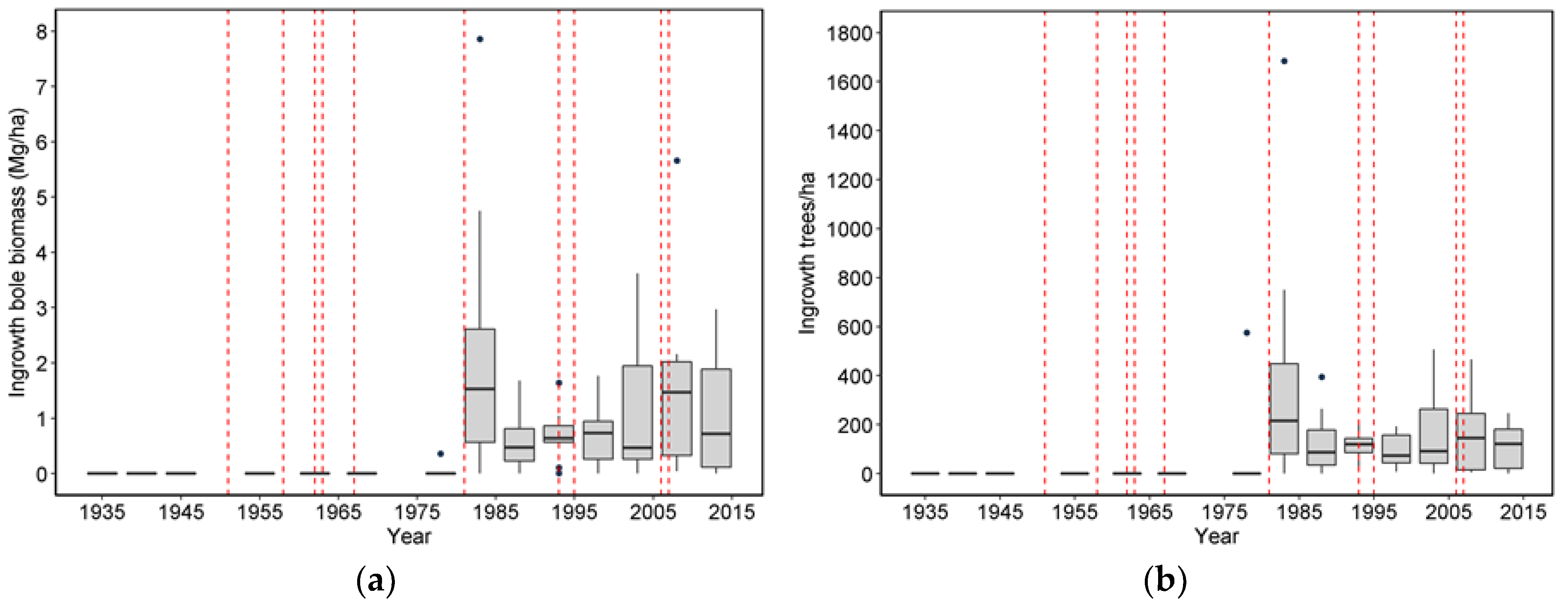
20] list four pathways or mechanisms for recovery following wind disturbance: regrowth, recruitment, release, and repression. Although some foliage and branches were lost in wind disturbance and these likely regrew, the primary process of regrowth in the forests we studied is likely related to sprouting of branches along snapped stems (Sitka spruce can produce epicormics branches). However, a relatively small number of trees suffered broken stems (125), and about half of these died. In comparison, approximately 5000 trees were tagged as ingrowth over the course of the study, indicating that recruitment and release are much more important in these forests. It should be noted that while ingrowth was certainly influenced by wind-related mortality, it also occurred, sometimes prolifically, in plots without major wind-related mortality. Since we did not track seedlings in these plots it was difficult to distinguish recovery via new recruitment versus recovery via release of existing (pre-disturbance) tree seedlings. To some extent, these two mechanisms could be distinguished by the lag between wind disturbance and the ingrowth pulse; in the case of CH06, the lag of 10 years indicated a substantial contribution to recovery by recruitment. However, in most of the other plots a lag was not readily apparent, and a pulse of ingrowth started before wind disturbance impacted most of the plots. This suggests that for most plots, the predominant recovery mechanism was via the release of existing small trees. For the most part, we did not observe the process of repression being important despite a fairly dense understory of herbs and shrubs that developed in openings created by wind.
As noted by Foster and Boose [
23], the majority of studies examining succession following wind disturbance consider either small gap-sized areas or large areas of catastrophic disturbance, but largely ignores situations between these extremes which happen to be quite common. In the case of gap-sized disturbances, regrowth and expansion of surviving trees could lead to little change in density, regeneration, composition, NPP, or biomass at the stand level. With larger gaps, these variables may change over time; if the disturbance repeats in other locations in the stand, wind disturbance may maintain the forest live biomass in quasi-equilibrium [
19,
27,
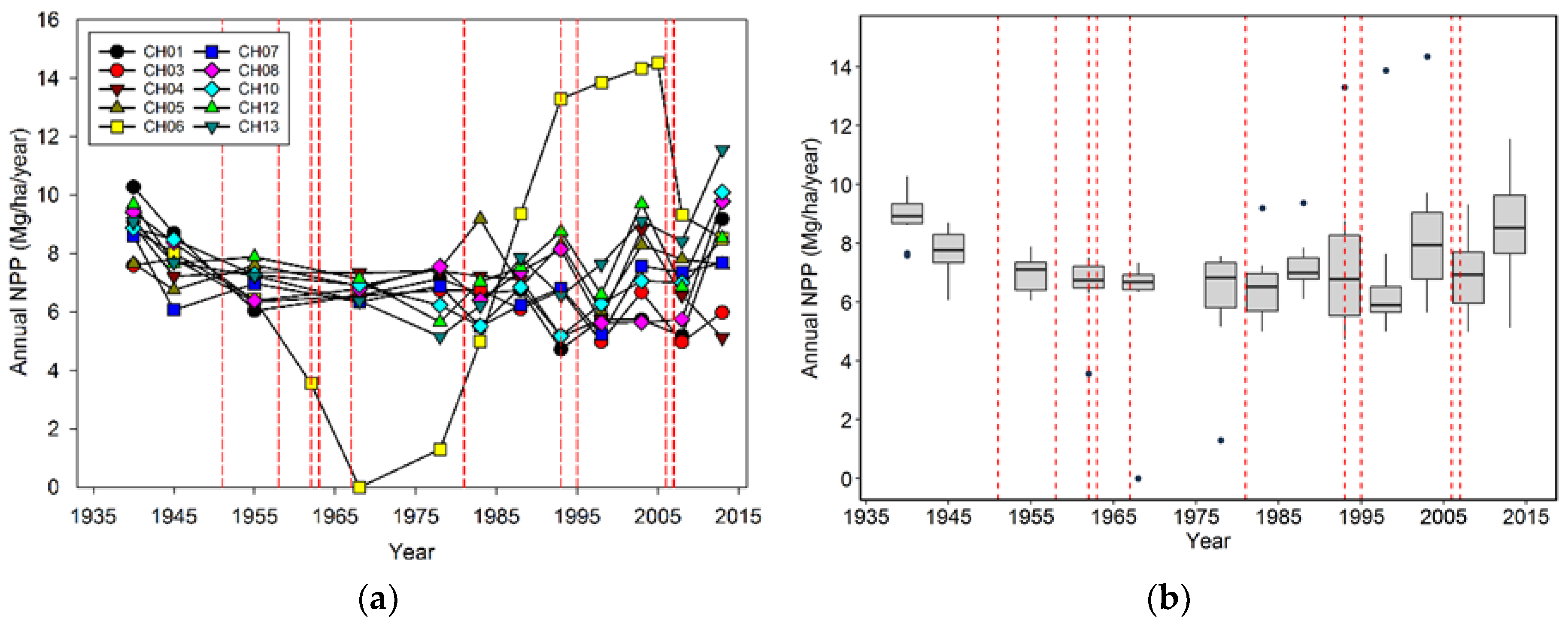
28]. In cases where severe wind mortality impacts an entire stand, a “typical” successional scenario might be a major pulse of mortality followed by pulse of ingrowth followed by a second period of increased mortality due to competition/suppression. Also, if the disturbance is severe enough, then there could be a decrease in woody-related NPP, followed by an increase, and quite possibly a decrease as the new stand develops [
29]. While we observed this temporal pattern of NPP in CH06, that was the exception. The changes in the plots we observed strongly suggest that the majority of stands do not undergo solely either gap or catastrophic wind-related mortality; instead there are more subtle changes in ingrowth, mortality, and NPP that are hard to distinguish from stands not undergoing wind disturbance. Moreover, at the landscape level, even individual catastrophic events do not necessarily have an impact on overall species composition, tree density, biomass, ingrowth, mortality, or NPP. Rather it is the accumulation of multiple events in previously and newly disturbed stands that have the largest impact on these variables.
Of all the variables we examined in this forest, NPP seems the least responsive to wind-related disturbance. In part this result may have been due to our use of a 5–10-year measurement interval to determine NPPb; had we determined this variable annually, then we might have observed a greater and more short-lived reduction after wind disturbance. Therefore, one could make the case at the plot scale that there is a dip in NPP after major wind mortality and then a rebound that overshoots the long-term average. However, this only seems to have occurred when mortality was very high (CH06), while for most of the plots, temporal changes in NPP were muted. Moreover, at the multi-plot scale, NPP was even more stable than that observed at the plot level. Since we did not include the NPP of trees smaller than 5.0 cm dbh, non-woody parts (i.e., foliage) nor understory vegetation, NPP is probably less influenced by wind-related disturbance than our analysis indicates. In this particular environment, water, light, and nutrient resources are relatively abundant, thus surviving or establishing trees and understory plants will use them, keeping NPP relatively high at both the plot and multi-plot scale.
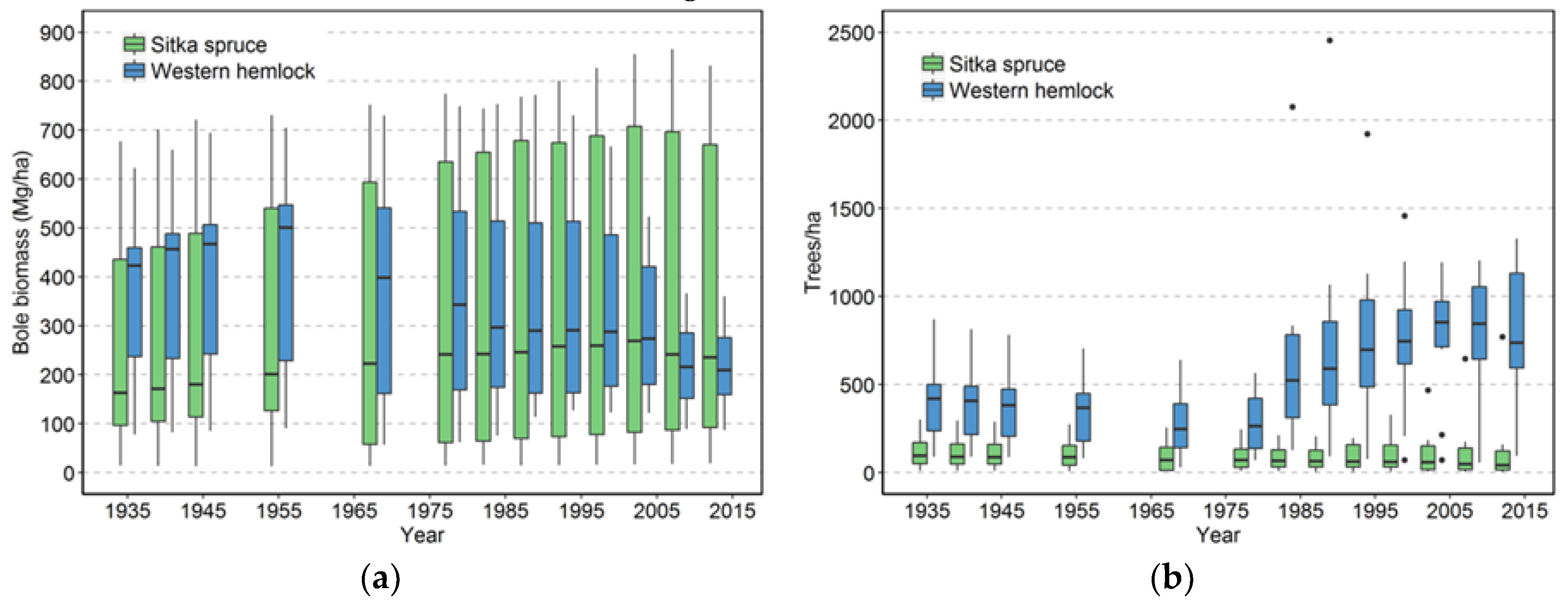
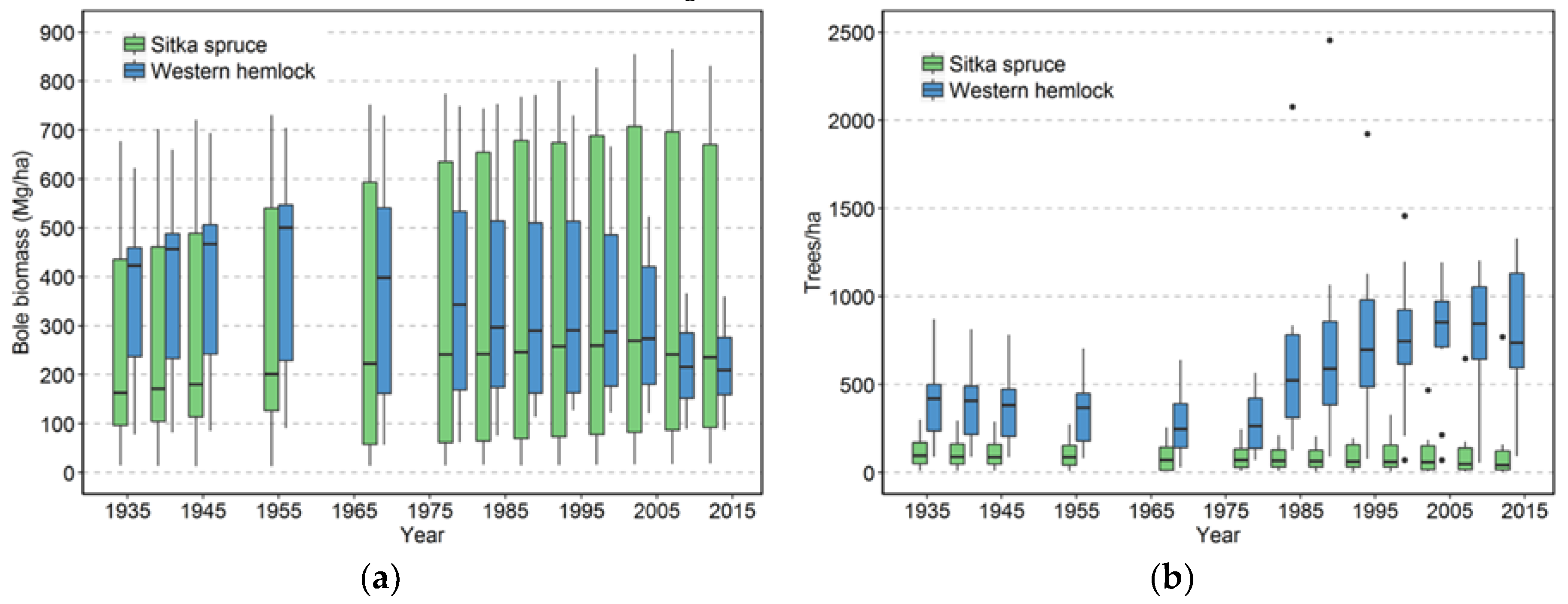
The effects on species composition also seem to be minimal and counterintuitive. The limited amount of change is probably a function of the limited number of species present in this system. The only truly early successional tree species, red alder, is not particularly abundant in these plots, although stands dominated by alder do occur nearby [
30]. The counterintuitive result is that the climax species, western hemlock, is increasing its numbers more than Sitka spruce, a species thought to require large gaps for regeneration [
19]. In this moist, resource-rich environment, western hemlock is not restricted to the later stages of succession and can successfully regenerate in open settings as well as in shade. Moreover, western hemlock can maintain itself in the understory without substantial growth for many decades. Both attributes give this species an advantage over Sitka spruce in terms of numbers. Sitka spruce’s advantage appears to be in its longevity and possibly its wind firmness relative to western hemlock [
25]. This likely explains why the proportion of biomass is increasing in Sitka spruce, but the proportion of TPH is increasing in western hemlock. This pattern may be similar to that observed in the Rocky Mountains where Engelmann spruce (
Picea engelmannii Parry ex. Engelm.) was able to maintain itself in the presence of subalpine fir (
Abies lasiocarpus (Hook.) Nutt.), not by abundantly reproducing after disturbances, but by having a lower mortality rate than subalpine fir [
28].
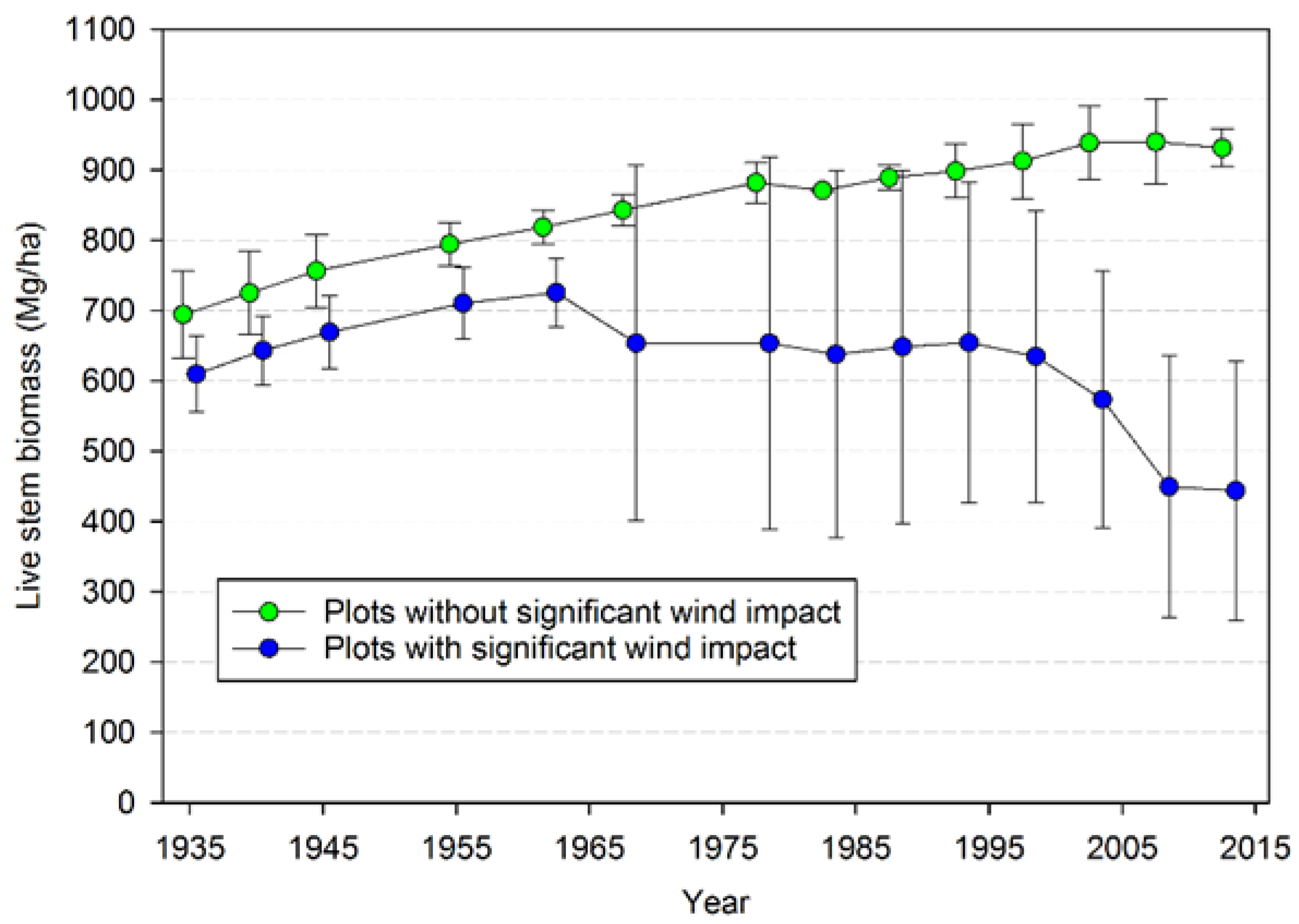
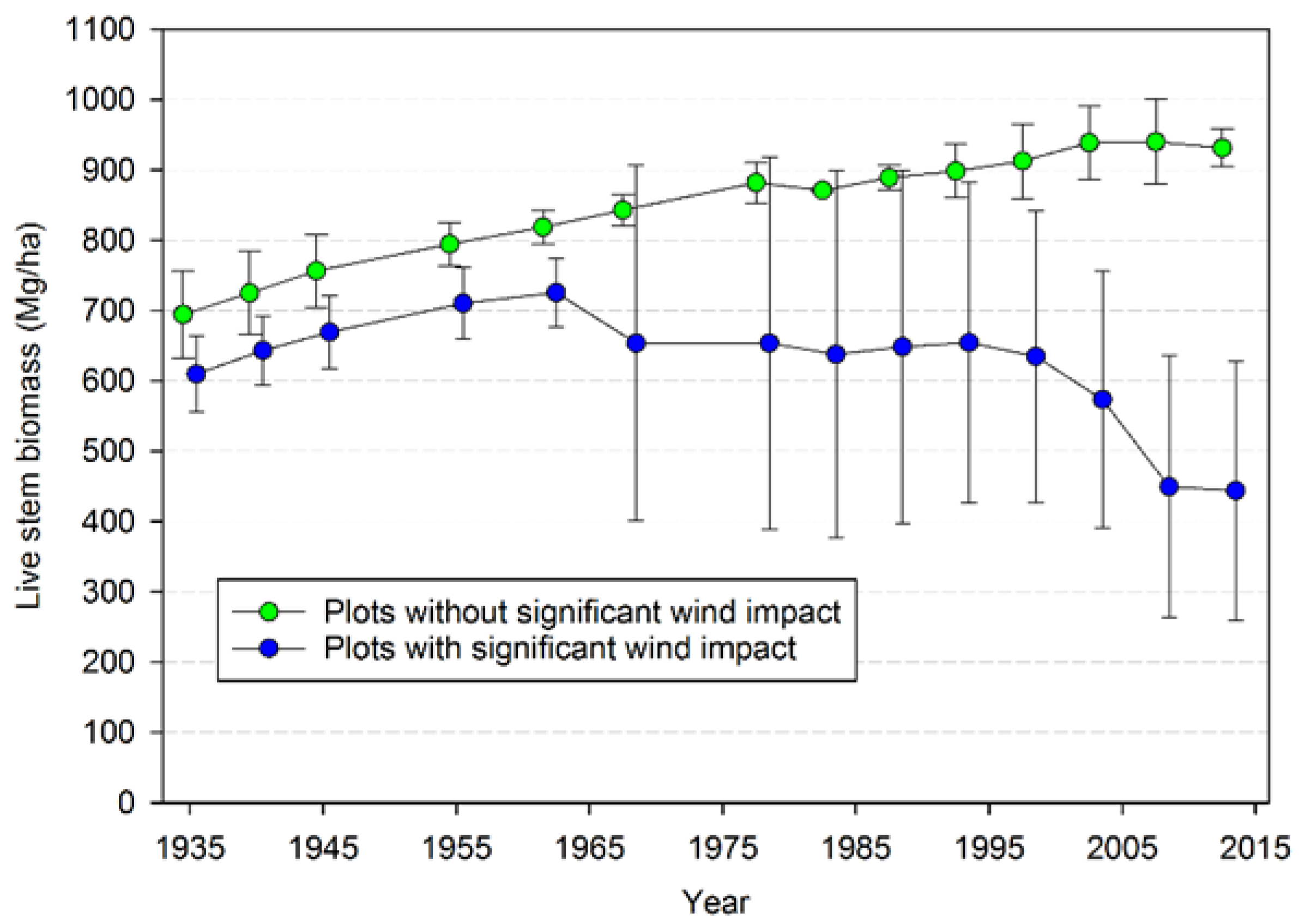
Input-output balances are important in understanding how both TPH and biomass have evolved in this landscape. When NPP, biomass mortality, and net change in biomass are considered for all the plots, biomass mortality has generally exceeded NPP
b since 1968, leading to an overall loss of biomass since that time (
Figure S5). The opposite is true for TPH, with ingrowth generally exceeding stem mortality since 1978, even in plots not impacted significantly by wind disturbance (
Figure S6). For the three plots that were not impacted by major wind disturbance there is some evidence that the rate of biomass accumulation has gradually slowed over time and that mortality became approximately equal to NPP
b (
Figure S7). In contrast, mortality generally exceeded NPP
b since 1968 in the seven plots that have experienced major wind-related losses (
Figure S7). Examination of the possible factors responsible for the divergence of plots with and without major wind disturbance suggests that initial species composition was more important than soil depth or aspect. Soil depth in all the plots exceeded 1 m and deeper than the root depth of these species, thus shallow soils did not seem to be an explanation. Two of the plots not strongly influenced by wind (CH04 and CH05) had SW aspects, the predominant direction of strong winds. The remaining plot with little wind disturbance (CH10) had a NW aspect and may have been protected, but one of the plots strongly influenced by wind CH13) had a N aspect suggesting that northerly aspects can be subject to wind disturbance in this landscape. In fact, all aspects except for E (which were not sampled) appeared to have plots with major wind impacts. Those plots without strong wind disturbance effects all had at least 65% of their initial biomass comprised of Sitka spruce. In contrast, only one of the plots with major wind disturbance (CH03) had the majority of initial biomass comprised of Sitka spruce (62%); the remaining plots had 74%–97% of their initial biomass comprised of western hemlock. Thus, at least for this landscape, an initial predominance of more wind-susceptible species (i.e., western hemlock) may eventually lead to greater changes in biomass than if wind-firm species (Sitka spruce) predominate. It is not clear if there will be additional decreases of biomass in these plots, or whether biomass will gradually increase again as released trees and newly regenerated ones gain biomass. The critical factor is likely the near-term reoccurrence of major impacts from wind or other disturbance: if, on one hand, additional major losses recur in the near future, then biomass may reach a new, lower steady-state as hypothesized by Acker et al. [
13]. On the other hand, if future losses are low because these plots are now more wind resistant (which we view as unlikely) or major windstorms do not occur for many years (also unlikely), then biomass would likely begin to significantly increase.
Wind and fire are significant disturbances in the Pacific Northwest region, but leave contrasting imprints on the forests because of differences in their temporal and spatial patterns, and also because of their different biological legacies. As noted by Harcombe et al. [
11], in the coastal landscape fire creates a long-term stage upon which wind disturbances play out, gradually imposing a new pattern as wind effects accumulate. Because wind disturbance leaves a significant legacy of either surviving large trees or advanced regeneration, forest response following wind disturbance events can be more rapid compared to fire. Therefore, one might think of wind as a slow disturbance (particularly when events occur in series) coupled to a rapid response. Fire might be thought of in the opposite manner as a fast disturbance coupled to a slow recovery process given that it can remove seed sources and advanced regeneration. This suggests that to fully understand forest structure and function one needs to examine how multiple disturbances interact over broad areas and long durations.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}