Long-term Growth Variation and Selection of Geographical Provenances of Cunninghamialanceolata (Lamb.) Hook

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Test Design
2.3. Data Collection
2.4. Data Analysis
3. Results and Analysis
3.1. Variance Analysis of Growth Traits of Chinese Fir with Different Provenances
3.2. Growth Variation of Chinese Firwith Different Provenances
3.3. Correlation Analysis of Growth Traits and Geographical Factors of Chinese Fir with Different Provenances
3.4. Bioclimatic Analyses of Chinese Fir
3.5. Correlation between Juvenile–Mature Growth Traits and the Early Selection Effect of Chinese Fir
3.6. Cluster Analysis of Chinese Fir with Different Provenances
3.7. Selection Analysis and Risk Estimation of Chinese Fir with Different Provenances in Early and Late Growth Stages
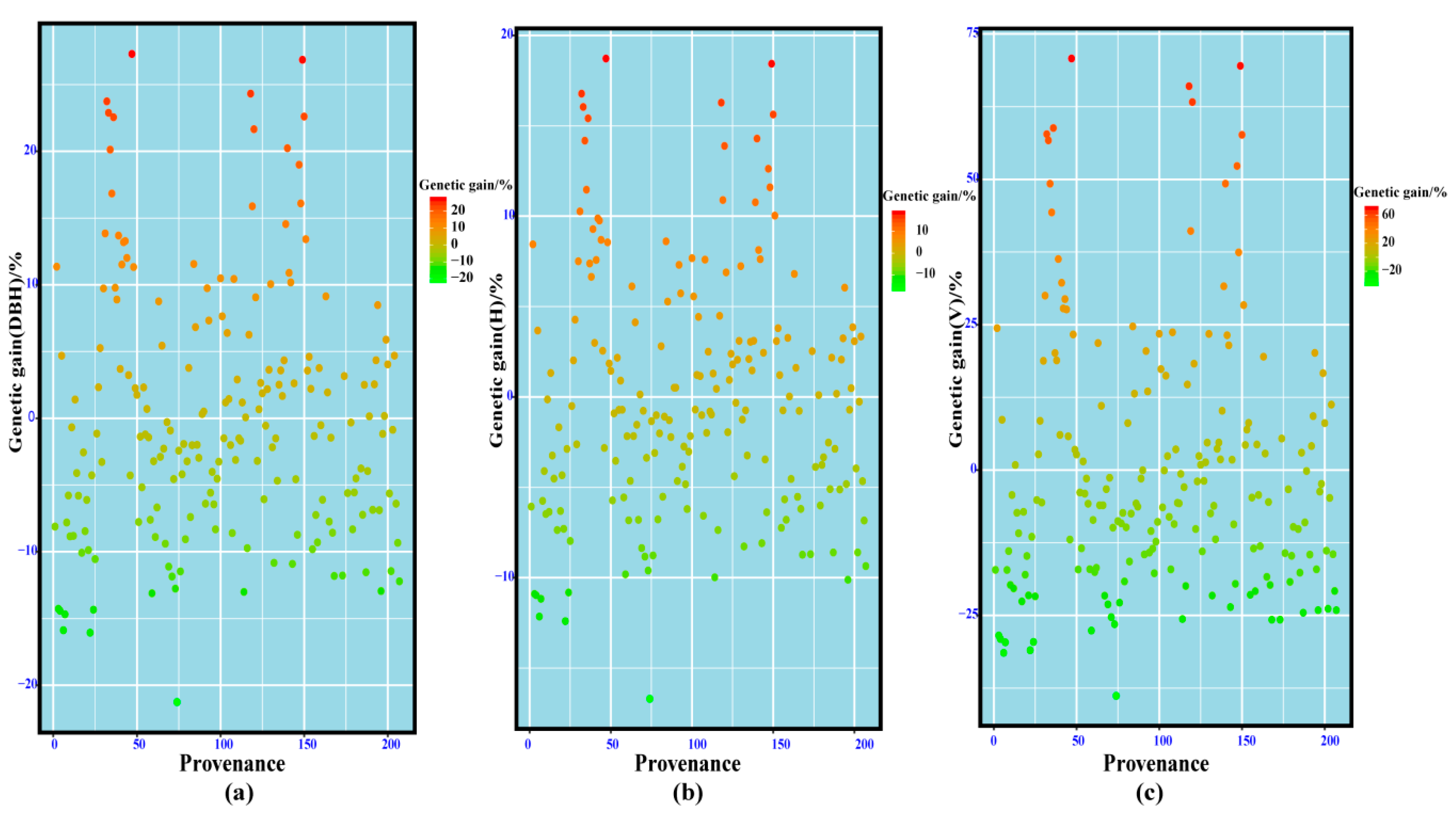
3.8. Genetic Gain Evaluation of Excellent Provenances of Chinese Fir
4. Discussion
4.1. Genetic Variation Analysis of Growth Traits of Chinese Firwith Different Provenances
4.2. Analysis of Geographical Variation Model of Chinese Fir Provenance
4.3. Appropriate Forest Age for the Early Selection of Excellent Provenance Areas of Chinese Fir
4.4. Evaluation of the Effect of Excellent Provenances of Chinese Fir
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zobel, B.; Talbert, J. Applied Forest Tree Improvement; John Wiley & Sons: New York, NY, USA, 1984; p. 496. [Google Scholar]
- Yu, Q.; Pulkkinen, P. Genotype–environment interaction and stability in growth of aspen hybrid clones. For. Ecol. Manag. 2003, 173, 25–35. [Google Scholar] [CrossRef]
- Larsen, C.P.S.; Macdonald, G.M. Relations between tree-ring widths, climate, and annual area burned in the boreal forest of Alberta. Can. J. For. Res. 1995, 25, 1746–1755. [Google Scholar] [CrossRef]
- Wang, Y.L.; Li, Y.; Chen, X.Y. Phenotypic diversity of natural populations in Picea crassifolia in Qilian Mountains. Sci. Silvae Sin. 2008, 44, 70–77. [Google Scholar]
- Nagamitsu, T.; Nagasaka, K.; Yoshimaru, H.; Tsumura, Y. Provenance tests for survival and growth of 50-year-old Japanese larch (Larix kaempferi) trees related to climatic conditions in central Japan. Tree Genet. Genomes 2014, 10, 87–99. [Google Scholar] [CrossRef]
- Libby, W.J. Domestication Strategies for Forest Trees. Can. J. For. Res. 1973, 3, 265–276. [Google Scholar] [CrossRef]
- Gwaze, D.P.; Bridgwater, F.E.; Byram, T.D.; Lowe, W.J. Genetic parameter estimates for growth and wood density in loblolly pine (Pinus taeda L.). Int. J. For. Genet. 2001, 8, 47–55. [Google Scholar]
- Lambeth, C.C.; Dill, L.A. Prediction models for juvenile-mature correlations for loblolly pine growth traits within, between and across sites. For. Genet. 2001, 8, 101–108. [Google Scholar]
- Mckeand, S.E. Optimum Age For Family Selection for Growth in Genetic Tests of Loblolly Pine. For. Sci. 1988, 34, 400–411. [Google Scholar]
- Jansson, G.; Li, B.; Hannrup, B. Time Trends in Genetic Parameters for Height and Optimal Age for Parental Selection in Scots Pine. For. Sci. 2003, 49, 696–705. [Google Scholar]
- Sheng, W.T. Plantation Forest and Their Silviculture Systems in China; China Forestry Press: Beijing, China, 2014; pp. 2–6. [Google Scholar]
- SFA. Reports of Chinese Forestry Resource: The 8th National Forest Resource Inventory; Chinese Forestry Publishing House: Beijing, China, 2013; p. 16. [Google Scholar]
- Yu, X.T.; Chen, C.D.; He, Z.Y.; Ye, Z.C.; Cheng, S.X. Growth and wood properties of Cunninghamia lanceolata provenance forest (23 years). J. Fujian Coll. For. 1984, 4, 1–8. [Google Scholar]
- Ye, Z.H.; Shi, J.S.; Wen, Y.Z.; Li, S.M.; Yu, R.Z.; Chen, R.X. Geographic variation and inheritance, correlation and selection of traits of provenances of Cunninghamia lanceolata. J. Nanjing For. Univ. 1991, 15, 7–10. [Google Scholar]
- Hong, J.S. Excellent provenance selection of Cunninghamia lanceolata afforestation. For. Res. 1994, 7, 1–25. [Google Scholar]
- Ulbrichová, I.; Podrázský, V.; Beran, F.; Zahradník, D.; Fulin, M.; Procházka, J.; Kubeček, J. Picea abies provenance test in the Czech Republic after 36 years—Central European provenances. J. For. Sci. 2015, 61, 465–477. [Google Scholar] [CrossRef]
- Soil Systematic Classification Research Group, Nanjing Institute of Soil Science, Chinese Academy of Sciences, Cooperative Group on Soil Systematic Classification Research in China. Soil Systematic Classification Retrieval of China, 3rd ed.; Press of China University of Science and Technology: Hefei, China, 2001; pp. 111–113. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Guarino, L.; Gruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data(1): DIVA-GIS. Plant Genet. Resour. Newsl. 2001, 127, 15–19. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Klisz, M.; Ukalski, K.; Ukalska, J.; Jastrzębowski, S.; Puchałka, R.; Przybylski, P.; Mionskowski, M.; Matras, J. What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments? Forests 2018, 9, 441. [Google Scholar] [CrossRef]
- Klisz, M.; Ukalska, J.; Koprowski, M.; Tereba, A.; Puchałka, R.; Przybylski, P.; Jastrzębowski, S.; Nabais, C. Effect of provenance and climate on intra-annual density fluctuations of Norway spruce Picea abies (L.) Karst. in Poland. Agric. For. Meteorol. 2019, 269, 145–156. [Google Scholar] [CrossRef]
- Wang, H.F. Introduction to Econometric Geography; Shandong Education Press: Jinan, China, 2001; pp. 90–92. [Google Scholar]
- Tang, Q.Y.; Feng, M.G. DPS Data Processing System; Science Press: Beijing, China, 2007; pp. 661–665. [Google Scholar]
- Xia, G.Q.; Wu, Y.C. Multicharacter selection plus Provenances of Cunninghamia lanceolata. J. Zhejiang For. Sci. Technol. 1999, 19, 1–5. [Google Scholar]
- Zhou, C.; Zeng, Z.G. Comparison research on growth characteristics of Cunninghamia lanceolata provenance’s young stage and maturity stage. Jiangxi For. Sci. Technol. 2006, 3, 10–12. [Google Scholar]
- Fan, L.; Jiang, B.; Shen, A.H.; Hong, C.D.; Xu, J.L.; Wu, Y.C.; Xu, G.C. Multi-traits selection of plus provenances of fast-growing Cunninghamia lanceolata. J. Zhejiang For. Sci. Technol. 2010, 30, 24–32. [Google Scholar]
- Xiang, B.; Li, B.; Isik, F. TimeTrend of Genetic Parameters in Growth Traits of Pinus taeda L. Silvae Genet. 2003, 52, 114–121. [Google Scholar]
- Wu, J.Y.; Cheng, Y.; Chen, J.F.; Xie, H.M.; Liao, D.Z.; Li, H.J.; Wei, Z.H.; Wu, Z. Heritability Analysis of Growth Traits and Selection of Semi-sib Families of Pinus elliottii. Agric. Biotechnol. 2018, 7, 160–162. [Google Scholar]
- Barzdajn, W. Results of a 24-year provenance trial with Scots pine in the Suprasl Forest District. Sylwan 2008, 152, 21–29. [Google Scholar]
- Rzeźnik, Z. Scots pine (Pinus sylvestris L.) of European provenances in the Suprasl forest inspectorate. Rocz. Akad. Rol. W Pozn. Rozpr. Nauk. 1991, 219, 56–67. [Google Scholar]
- Stephan, B.R.; Liesbach, M. Results of the IUFRO 1982 Scots pine (Pinus sylvestris L.) provenance experiment in southwestern Germany. Silvae Genet. 1996, 45, 342–349. [Google Scholar]
- Androsiuk, P.; Kaczmarek, Z.; Urbaniak, L. The morphological traits of needles as markers of geographical differentiation in European Pinus sylvestris populations. Dendrobiology 2011, 65, 3–16. [Google Scholar]
- Amateis, R.L.; Prisley, S.P.; Burkhart, H.E.; Liu, J.P. The effect of physiographic region and geographic locale on predicting the dominant height and basal area of loblolly pine plantations. South. J. Appl. For. 2006, 30, 147–153. [Google Scholar] [CrossRef]
- Shen, X.H. Selection and Breeding of Pinus Tabulaeformis and Larix Principis-Rupprechtii: Practice and Theory; Science Press: Beijing, China, 2015. [Google Scholar]
- Fang, S.; Xu, X.; Lu, S.; Tang, L. Growth dynamics and biomass production in short-rotation poplar plantations: 6-year results for three clones at four spacings. Biomass Bioenerg. 1999, 17, 415–425. [Google Scholar] [CrossRef]
- Caliński, T.; Czajka, S.; Kaczmarek, Z.; Krajewski, P.; Pilarczyk, W. A mixed model analysis of variance for multi-environment variety trials. Stat. Pap. 2009, 50, 735–759. [Google Scholar] [CrossRef]
- Sheng, W.T.; Hui, G.Y.; Zhang, S.G. Optimum Cultivation Pattern of Chinese Fir Plantation; China Science and Technology Publishing House: Beijing, China, 2004; pp. 93–98. [Google Scholar]
- Ma, C.G. Research status of accelerated tree breeding of the world. World For. Res. 1996, 9, 15–23. [Google Scholar]
- Yang, X.Y.; Ji, K.S. Early selection in forest tree improvement. World For. Res. 2004, 17, 6–8. [Google Scholar]
- He, H.; Dai, H. Research on theories and methods of early selection of forest tree. Rural Econ. Sci. Technol. 2012, 23, 33–36. [Google Scholar]
- Wang, Z.R.; Chen, T.H.; Zhou, Z.C.; Zheng, J.J.; Wang, Y.L. Early-late correlation and early selection of the growth traits of Pinus massoniana Lamb. in Huaan county, Fujian province. J. Nanjing For. Univ. 1987, 11, 41–47. [Google Scholar]
- Jin, G.Q.; Qin, G.F.; Liu, W.H.; Chu, D.Y.; Feng, Z.P.; Zhou, Z.C. Provenance selection effect at different stand age of Pinus massoniana. Sci. Silvae Sin. 2011, 47, 39–45. [Google Scholar]
- Zhou, H.; Fries, A.; Wu, H.X. Age trend of heritability, genetic correlation and efficiency of early selection for wood quality traits in Scots pine. Can. J. For. Res. 2015, 45, 817–825. [Google Scholar]
- Lai, M.; Sun, X.; Chen, D.; Xie, Y.; Zhang, S. Age-related trends in genetic parameters for Larix kaempferiand their implications for early selection. Bmc Genet. 2014, 15 (Suppl. S1), S10. [Google Scholar] [CrossRef]
- Ye, P.Z.; Chen, Y.W.; Ruan, Y.C.; Chen, S.B.; Liu, D.L.; Guo, M.C. Study on early selection of Cunninghamia lanceolata. J. Nanjing For. Univ. 1981, 5, 106–116. [Google Scholar]
- Zhao, C.K. A study on optimum age and gain for early selection of superior clone in Cunninghamia lanceolata. Sci. Silvae Sin. 2002, 38, 53–60. [Google Scholar]
- Cotterill, P.P.; Dean, C.A. Changes in the genetic control of growth of radiata pine to 16 years and efficiencies of early selection. Silvae Genet. 1987, 37, 138–146. [Google Scholar]
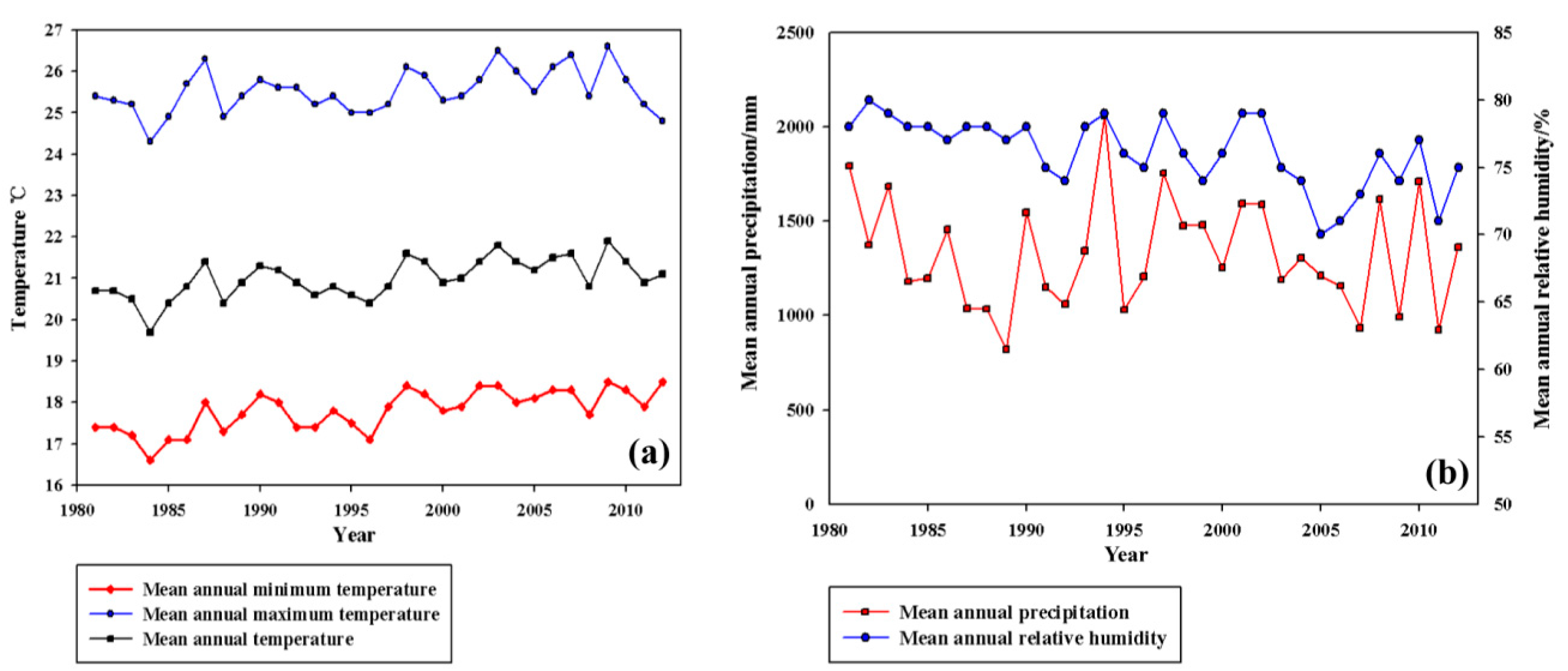
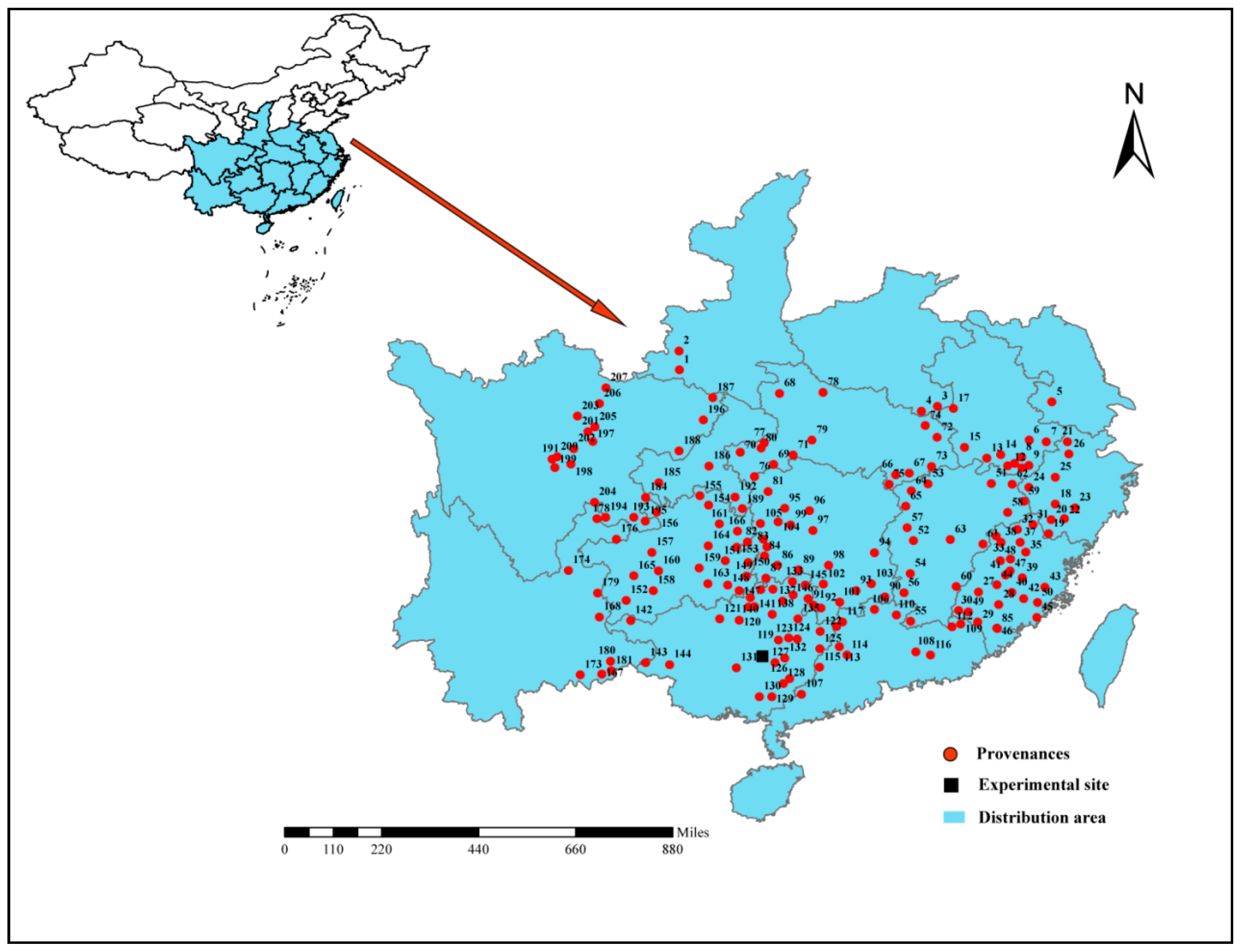
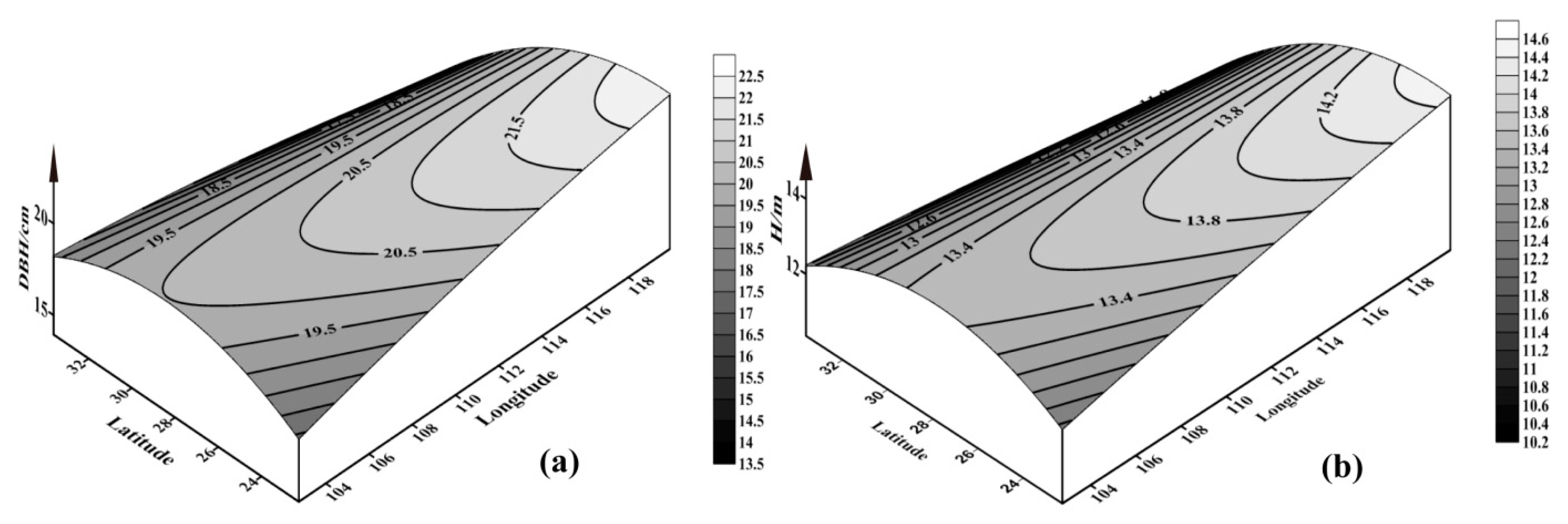
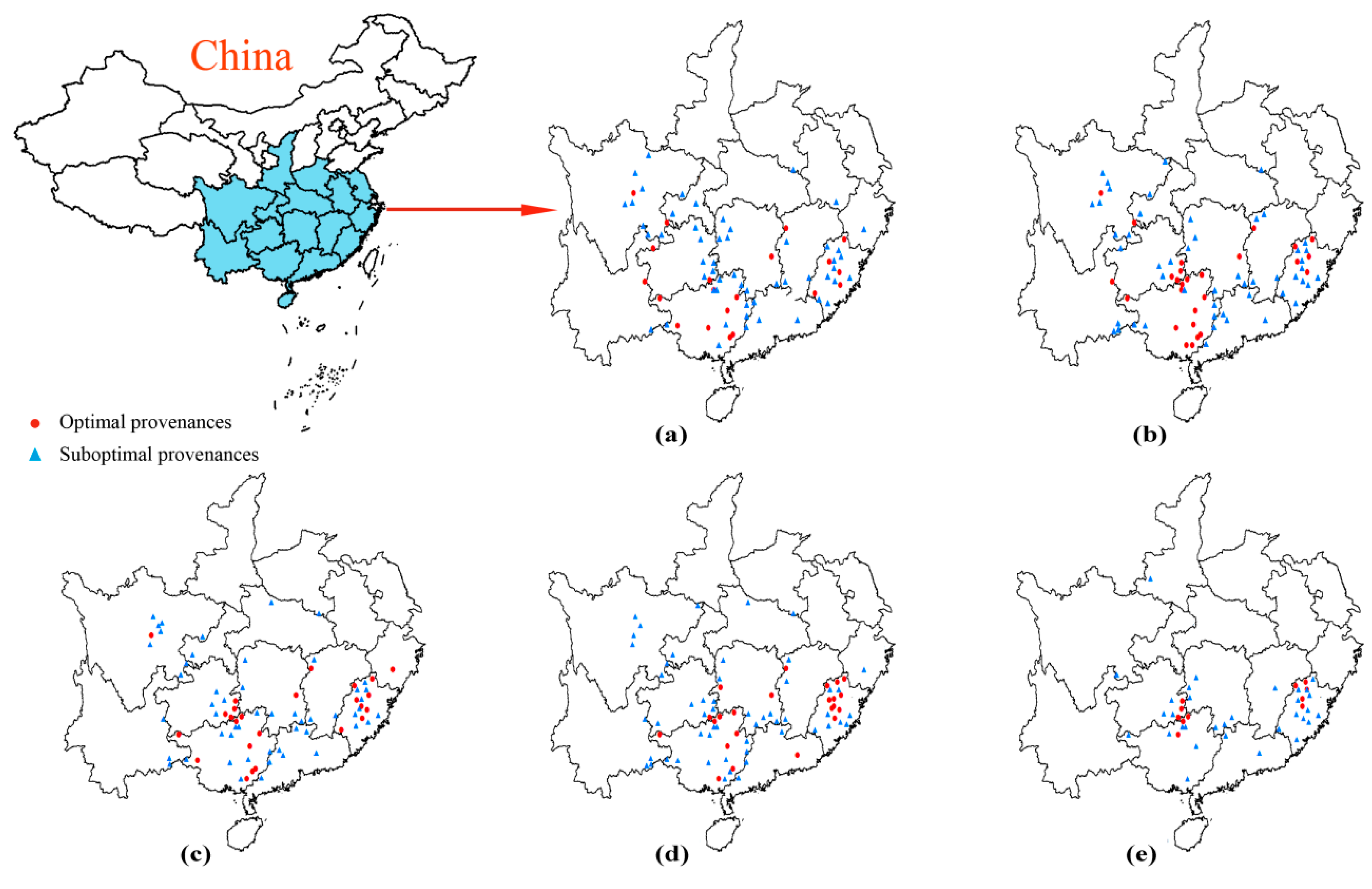

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forests Age (Years) | Source of Variation | DF | DBH/cm | Height/m | Volume/m3 | |||
|---|---|---|---|---|---|---|---|---|
| Mean Square | F Value | Mean Square | F Value | Mean Square | F Value | |||
| 5 | Provenance | 194 | 3.60 | 2.34 ** | 0.97 | 2.23 ** | 1.93 × 10−5 | 2.16 ** |
| Block | 4 | 316.57 | 205.63 ** | 68.19 | 156.08 ** | 1.72 × 10−3 | 192.88 ** | |
| Experimental error | 776 | 1.54 | 0.44 | 8.93 × 10−6 | ||||
| 6 | Provenance | 194 | 8.04 | 4.05 ** | 2.35 | 4.04 ** | 1.15 × 10−4 | 4.40 ** |
| Block | 9 | 227.48 | 114.65 ** | 65.22 | 111.98 ** | 2.62 × 10−3 | 100.07 ** | |
| Experimental error | 1746 | 1.98 | 0.58 | 2.67 × 10−5 | ||||
| 8 | Provenance | 194 | 11.86 | 4.68 ** | 4.12 | 4.54 ** | 5.35 × 10−4 | 4.88 ** |
| Block | 9 | 163.74 | 64.57 ** | 72.62 | 80.08 ** | 7.29 × 10−3 | 66.53 ** | |
| Experimental error | 1746 | 2.54 | 0.91 | 1.10 × 10−4 | ||||
| 12 | Provenance | 194 | 11.16 | 5.15 ** | 4.49 | 3.90 ** | 1.03 × 10−3 | 4.97 ** |
| Block | 9 | 34.75 | 16.03 ** | 51.82 | 45.03 ** | 4.01 × 10−3 | 19.42 ** | |
| Experimental error | 1746 | 2.17 | 1.15 | 2.07 × 10−4 | ||||
| 33 | Provenance | 194 | 47.61 | 4.08 ** | 11.12 | 4.00 ** | 3.11 × 10−2 | 4.61 ** |
| Block | 6 | 119.63 | 10.26 ** | 27.23 | 9.79 ** | 8.15 × 10−2 | 10.66 ** | |
| Experimental error | 1164 | 11.66 | 2.78 | 7.65 × 10−3 | ||||
| Forest Age (Years) | Trait | Min. | Max. | Mean | Standard Deviation (SD) | Coefficient of Variation (CV)/% | Heritability |
|---|---|---|---|---|---|---|---|
| 5 | DBH (cm) | 0.8 | 9.7 | 4.49 | 1.8 | 38.95 | 0.57 |
| Height (m) | 0.76 | 5.8 | 3.38 | 0.9 | 25.67 | 0.55 | |
| Volume(m3) | 9.24 × 10−5 | 0.022 | 4.94 × 10−3 | 4.20 × 10−3 | 81.95 | 0.54 | |
| 6 | DBH (cm) | 1 | 12.8 | 6.69 | 1.91 | 28.55 | 0.75 |
| Height (m) | 1 | 7.65 | 4.47 | 1.03 | 23.06 | 0.75 | |
| Volume (m3) | 2.10 × 10−4 | 0.043 | 0.012 | 6.86 × 10−3 | 59.65 | 0.77 | |
| 8 | DBH (cm) | 1.5 | 15.1 | 9.34 | 2.05 | 21.98 | 0.79 |
| Height (m) | 1.5 | 10.8 | 6.42 | 1.25 | 19.52 | 0.78 | |
| Volume (m3) | 3.94 × 10−4 | 0.086 | 0.028 | 0.0136 | 47.99 | 0.80 | |
| 12 | DBH (cm) | 2.8 | 17 | 10.93 | 1.93 | 17.67 | 0.81 |
| Height (m) | 3 | 17 | 8.34 | 1.34 | 16.08 | 0.74 | |
| Volume (m3) | 2.70 × 10−3 | 0.126 | 0.045 | 0.018 | 40.56 | 0.80 | |
| 33 | DBH (cm) | 7.2 | 40.7 | 20.08 | 5.4 | 26.91 | 0.76 |
| Height (m) | 6.07 | 21.37 | 13.35 | 2.62 | 19.63 | 0.75 | |
| Volume (m3) | 0.015 | 0.969 | 0.231 | 0.102 | 44.12 | 0.75 |
| Trait | Regression Equation of Trend Surface Analysis | Fitting Coefficient | p Value |
|---|---|---|---|
| DBH (cm) | Z = 1.804x + 8.484y − 0.003x2 − 0.076y2 − 0.041xy − 194.221 | 0.175 | <0.01 |
| Height (m) | Z = 0.905x + 4.151y − 0.001x2 − 0.037y2 − 0.021xy − 92.592 | 0.182 | <0.01 |
| Bioclimatic Variables | Abbreviation | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|---|
| Altitude | Alt | −0.77 | 0.42 | 0.35 | 0.12 |
| Annual mean temperature | bio1 | 0.84 | 0.39 | −0.36 | 0.08 |
| Mean diurnal range (mean of monthly temperatures (max temp-min temp)) | bio2 | −0.02 | 0.00 | 0.25 | 0.81 |
| Isothermality (BIO2/BIO7) (* 100) | bio3 | 0.01 | 0.83 | 0.12 | 0.52 |
| Temperature seasonality (standard deviation * 100) | bio4 | −0.05 | −0.96 | 0.03 | −0.16 |
| Max temperature of warmest month | bio5 | 0.77 | −0.44 | −0.34 | 0.01 |
| Min temperature of coldest month | bio6 | 0.62 | 0.67 | −0.35 | −0.03 |
| Temperature annual range (BIO5-BIO6) | bio7 | −0.04 | −0.94 | 0.09 | 0.04 |
| Mean temperature of wettest quarter | bio8 | 0.26 | 0.37 | −0.61 | −0.24 |
| Mean temperature of driest quarter | bio9 | 0.79 | 0.48 | −0.18 | 0.13 |
| Mean temperature of warmest quarter | bio10 | 0.84 | −0.30 | −0.38 | −0.05 |
| Mean temperature of coldest quarter | bio11 | 0.65 | 0.69 | −0.28 | 0.12 |
| Annual precipitation | bio12 | 0.82 | 0.03 | 0.52 | −0.08 |
| Precipitation of wettest month | bio13 | 0.59 | 0.34 | 0.63 | −0.23 |
| Precipitation of driest month | bio14 | 0.75 | −0.52 | 0.26 | 0.09 |
| Precipitation seasonality (coefficient of variation) | bio15 | −0.44 | 0.73 | 0.15 | −0.16 |
| Precipitation of wettest quarter | bio16 | 0.62 | 0.36 | 0.64 | −0.16 |
| Precipitation of driest quarter | bio17 | 0.75 | −0.53 | 0.28 | 0.10 |
| Precipitation of warmest quarter | bio18 | 0.20 | 0.69 | 0.45 | −0.37 |
| Precipitation of coldest quarter | bio19 | 0.82 | −0.39 | 0.29 | 0.15 |
| Eigenvalue | 7.56 | 6.33 | 2.74 | 1.36 | |
| Variance explained (%) | 37.78 | 31.66 | 13.68 | 6.78 |
| Trait | Forest Age | 5 | 6 | 8 | 12 | 33 |
|---|---|---|---|---|---|---|
| DBH | 5 | 1 | 0.815 ** | 0.776 ** | 0.700 ** | 0.323 ** |
| 6 | 0.836 ** | 1 | 0.939 ** | 0.881 ** | 0.516 ** | |
| 8 | 0.802 ** | 0.941 ** | 1 | 0.922 ** | 0.557 ** | |
| 12 | 0.76 ** | 0.898 ** | 0.935 ** | 1 | 0.601 ** | |
| 33 | 0.426 ** | 0.591 ** | 0.621 ** | 0.672 ** | 1 | |
| Height | 5 | 1 | 0.826 ** | 0.797 ** | 0.740 ** | 0.342 ** |
| 6 | 0.852 ** | 1 | 0.952 ** | 0.876 ** | 0.464 ** | |
| 8 | 0.811 ** | 0.934 ** | 1 | 0.901 ** | 0.529 ** | |
| 12 | 0.771 ** | 0.895 ** | 0.894 ** | 1 | 0.508 ** | |
| 33 | 0.392 ** | 0.516 ** | 0.551 ** | 0.543 ** | 1 | |
| Volume | 5 | 1 | 0.803 ** | 0.771 ** | 0.707 ** | 0.334 ** |
| 6 | 0.831 ** | 1 | 0.954 ** | 0.902 ** | 0.511 ** | |
| 8 | 0.816 ** | 0.946 ** | 1 | 0.939 ** | 0.565 ** | |
| 12 | 0.725 ** | 0.731 ** | 0.76 ** | 1 | 0.592 ** | |
| 33 | 0.395 ** | 0.536 ** | 0.582 ** | 0.602 ** | 1 |
| Cluster | Forest Age | Number of Selections | Number of Correct Selections | Correct Selection Rate (%) | Number of Incorrect Selections | Incorrect Selection Rate (%) | Number of Missed Selections | Missed Selection Rate (%) |
|---|---|---|---|---|---|---|---|---|
| Ⅰ | 5 | 20 | 1 | 5 | 19 | 95 | 10 | 90.91 |
| 6 | 27 | 6 | 22.22 | 21 | 77.78 | 5 | 45.45 | |
| 8 | 25 | 7 | 28 | 18 | 72 | 4 | 36.36 | |
| 12 | 23 | 7 | 30.43 | 16 | 69.57 | 4 | 36.36 | |
| 33 | 11 | 11 | 100 | 0 | 0 | 0 | 0 | |
| Ⅰ+Ⅱ | 5 | 73 | 24 | 32.88 | 49 | 67.12 | 19 | 45.24 |
| 6 | 77 | 30 | 38.96 | 47 | 61.04 | 13 | 30.95 | |
| 8 | 79 | 31 | 39.24 | 48 | 60.76 | 12 | 28.57 | |
| 12 | 77 | 29 | 37.66 | 48 | 62.34 | 14 | 33.33 | |
| 33 | 42 | 42 | 100 | 0 | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Duan, A.; Zhang, J. Long-term Growth Variation and Selection of Geographical Provenances of Cunninghamialanceolata (Lamb.) Hook. Forests 2019, 10, 876. https://doi.org/10.3390/f10100876
Wu H, Duan A, Zhang J. Long-term Growth Variation and Selection of Geographical Provenances of Cunninghamialanceolata (Lamb.) Hook. Forests. 2019; 10(10):876. https://doi.org/10.3390/f10100876
Chicago/Turabian StyleWu, Hanbin, Aiguo Duan, and Jianguo Zhang. 2019. "Long-term Growth Variation and Selection of Geographical Provenances of Cunninghamialanceolata (Lamb.) Hook" Forests 10, no. 10: 876. https://doi.org/10.3390/f10100876
APA StyleWu, H., Duan, A., & Zhang, J. (2019). Long-term Growth Variation and Selection of Geographical Provenances of Cunninghamialanceolata (Lamb.) Hook. Forests, 10(10), 876. https://doi.org/10.3390/f10100876