Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains

Abstract
1. Introduction
2. Materials and Methods
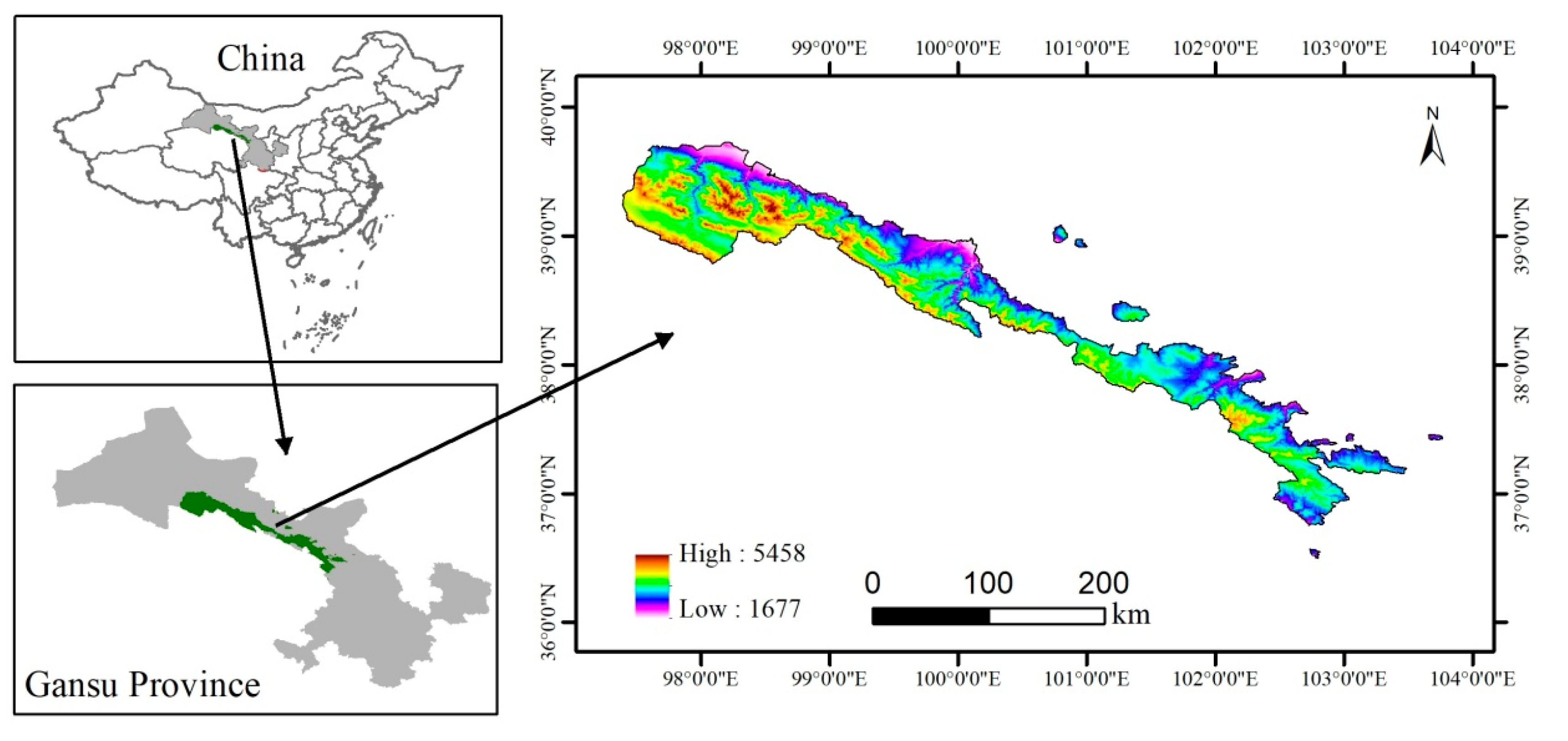
2.1. Study Area
2.2. Qinghai Spruce Distribution Data
2.3. Environmental Data
2.4. Ecological Niche Modeling
3. Results
3.1. Variables Analysis
3.2. Accuracy of the Maxent Model
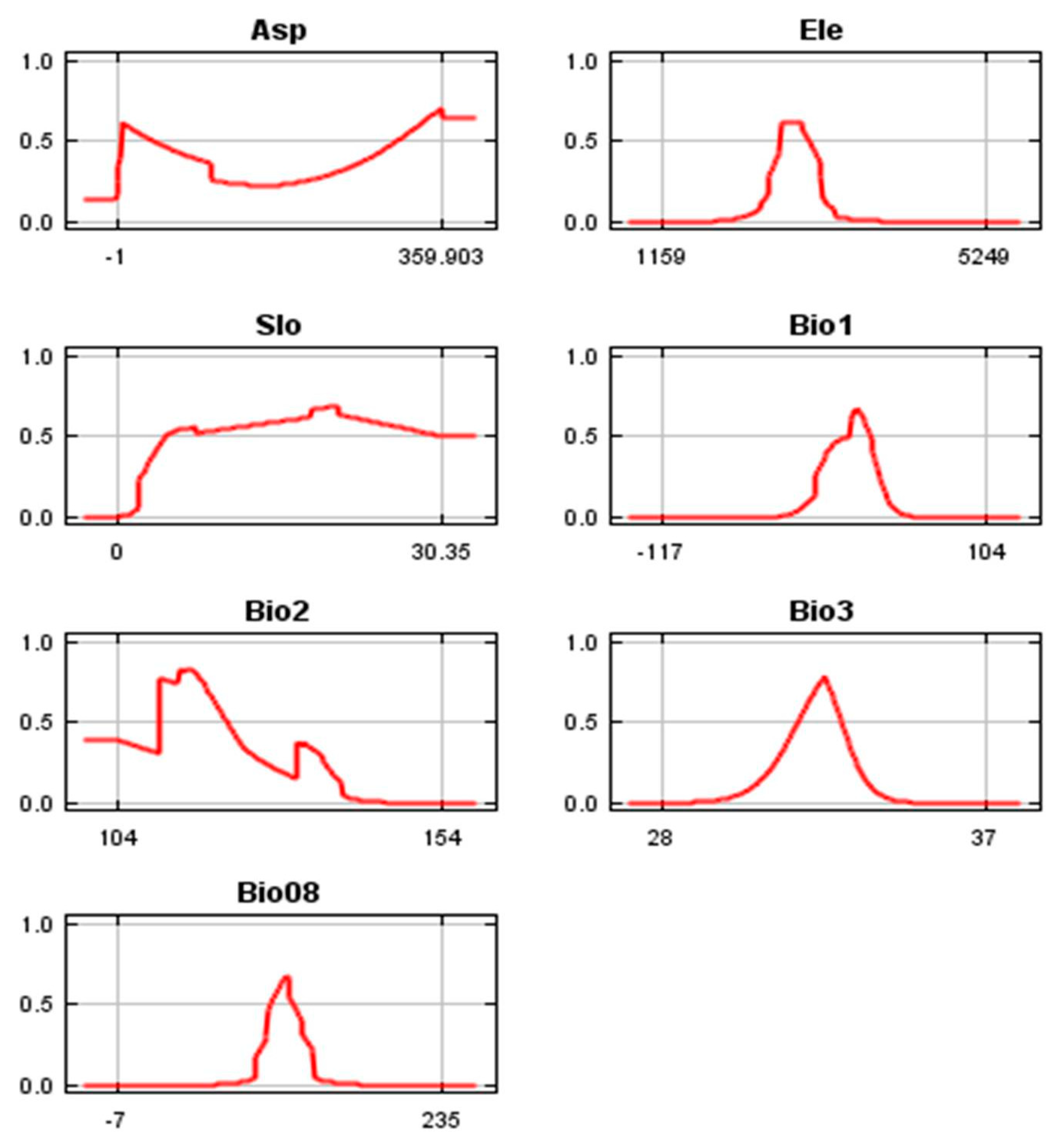
3.3. Variables’ Contribution and Variables’ Response to Suitability
3.4. Potential Distribution of Qinghai Spruce under Different Climate Scenario
3.5. The Transfer Matrix of Suitable Distribution Areas under the Current and Future Scenarios
3.6. The Relationship Between Qinghai Spruce’ Actual Distribution and Potential Distribution Under Current and Four Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Annex I: Atlas of Global and Regional Climate Projections. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Cambridge, UK; New York, NY, USA, 2013; p. 1311. [Google Scholar]
- Alberto, F.J.; Aitken, S.N.; Alia, R.; Gonzalez-Martinez, S.C.; Hanninen, H.; Kremer, A.; Lefevre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Bertelsmeier, C.; Paul, L.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Beltramino, A.A.; Vogler, R.E.; Gregoric, D.E.G.; Rumi, A. Impact of climate change on the distribution of a giant land snail from South America: Predicting future trends for setting conservation priorities on native malacofauna. Clim. Chang. 2015, 131, 621–633. [Google Scholar] [CrossRef]
- Davies, N.; Gramotnev, G.; Seabrook, L.; McAlpine, C.; Baxter, G.; Lunney, D.; Bradley, A. Climate-driven changes in diet composition and physiological stress in an arboreal folivore at the semi-arid edge of its distribution. Biol. Conserv. 2014, 172, 80–88. [Google Scholar] [CrossRef]
- McKenny, W.D.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchinson, M.F. Potential impacts of climate change on the distribution of north American trees. Bioscience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Willimas, S.E. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W. Climate change and the ecologist. Nature 2007, 448, 550–552. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Zhao, C.Y. Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agric. For. Meteorol. 2018, 262, 1–13. [Google Scholar] [CrossRef]
- Adhikari, D.; Barik, S.K.; Upadhya, K. Habitat distribution modeling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of Maximum Entropy Modeling in Wildlife Research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Kamilar, J.M.; Beaudrot, L. Understanding primate communities: Recent developments and future directions. Evol. Anthropol. 2013, 22, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Bakkenes, M.; Alkemade, J.R.M.; Ihle, F.; Leemans, R.; Latour, J.B. Assessing effects of forecasted climate change on the diversity and distribution of European higher plants for 2050. Glob. Chang. Biol. 2002, 8, 390–407. [Google Scholar] [CrossRef]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface 2013, 9, 2708–2717. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H.; Gao, B.; Gu, W. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom tricholoma matsutake under multiple climate change scenarios. Sci. Rep. 2017, 7, 46221. [Google Scholar] [CrossRef]
- Liu, X.; Guo, Z.; Ke, Z.; Wang, S.; Li, Y. Increasing potential risk of a global aquatic invader in Europe in contrast to other continents under future climate change. PLoS ONE 2011, 6, e18429. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T. Ecologic niche modeling and spatial patterns of disease transmission. Emerg. Infect. Dis. 2006, 12, 1822–1826. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, N.E., Jr.; Edwards, T.C.; Graham, C.H.; Pearman, P.B.; Svenning, J.C. New trends in species distribution modelling. Ecography 2010, 33, 985–989. [Google Scholar] [CrossRef]
- Sánchez, F.E. GARP modeling of natural and human factors affecting the potential distribution of the invasives Schismus arabicus and Brassica tournefortii, in ‘El Pinacate y Gran Desierto de Altar’ Biosphere Reserve. Ecol. Model. 2007, 204, 457–474. [Google Scholar] [CrossRef]
- Stockwell, D.R.; Beach, J.H.; Stewart, A. The use of the GARP genetic algorithm and internet grid computing in the Lifemapper world atlas of species biodiversity. Ecol. Model. 2006, 195, 139–145. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 251–270. [Google Scholar] [CrossRef]
- Carpentar, G.; Gillison, A.N.; Winter, J. DOMAIN: A flexible modelling procedure for mapping potential distributions of plants and animals. Biodivers. Conserv. 1993, 2, 667–680. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Hausser, J.; Chessel, D.; Perrin, N. Ecological-niche factor analysis: How to compute habitat-suitability maps without absence data? Ecology 2002, 83, 2027–2036. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Kumar, S.; Stohlgren, T.J. Maxent modelling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Costa, G.C.; Nogueira, C.; Machado, R.B.; Colli, G.R. Sampling bias and the use of ecological niche modeling in conservation planning: A field evaluation in a biodiversity hotspot. Biodivers. Conserv. 2009, 19, 883–899. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Papeş, M.; Gaubert, P. Modelling ecological niches from low numbers of occurrences: Assessment of the conservation status of poorly known viverrids (Mammalia, Carnivora) across two continents. Divers. Distrib. 2007, 13, 890–902. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Original article: Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in madagascar. J. Biogeogr. 2007, 34, 16. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance ofspecies distribution models. Biodivers. Lett. 2008, 14, 763–773. [Google Scholar]
- Phillips, S.J.; Miroslav, D.; Schapire, R.E. Maxent Software for Species Distribution Modeling. 2004. Available online: http://cs.princeton.edu/∼schapire/Maxent/ (accessed on 11 December 2015).
- Scheldeman, X.; Zonneveld, M.V. Training Manual on Spatial Analysis of Plant Diversity and Distribution; Biodiversity International: Rome, Italy, 2010; pp. 139–158. [Google Scholar]
- Tognelli, M.F.; Roig-Juñent, S.A.; Marvaldi, A.E.; Flores, G.E.; Lobo, J.M. An evaluation for methods on modelling distribution of Patagonian insects. Rev. Chil. Hist. Nat. 2009, 82, 347–360. [Google Scholar] [CrossRef]
- Fuller, D.O.; Ahumada, M.L.; Quiñones, M.L.; Herrera, S.; Beier, J.C. Near-present and future distribution of Anopheles albimanus in Mesoamerica and the Caribbean Basin modeled with climate and topographic data. Int. J. Health Geogr. 2012, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Reiss, H.; Cunze, S.; König, K.; Neumann, H.; Kröncke, I. Species distribution modelling of marine benthos: A North Sea case study. Mar. Ecol. Prog. Ser. 2011, 442, 71–86. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, C.; Feng, Z. Species distribution models to estimate the deforested area of picea crassifolia in arid region recently protected: Qilian Mts. National natural reserve (China). Pol. J. Ecol. 2012, 60, 515–524. [Google Scholar]
- Zhao, C.; Jia, Y.; Cheng, G.; Li, S.B. Estimation of leaf area index of Qinghai spruce (Picea crassifolia) forest using remote sensing in Qilian Mountains, Northwest China. In Proceedings of the 2007 IEEE International Geoscience and Remote Sensing Symposium, Barcelona, Spain, 23–27 July 2007; pp. 23–28. [Google Scholar]
- Irena, F.; Creed, I.F.; Noordwijk, M.V. Forest and Water on a Changing Planet: Vulnerability, Adaptation and Governance Opportunities; A Global Assessment Report; IUFRO World Series: Vienna, Austria, 2018; p. 192. [Google Scholar]
- Moritz, C.; Patton, J.L.; Conroy, C.J. Impact of a century of climate change on small-mammal communities in Yosemite National Park, USA. Science 2008, 322, 261–264. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, C.; Feng, Z.; Zhang, F.; Sher, H.; Wang, C.; Peng, H.; Wang, Y.; Zhao, Y.; Wang, Y. Estimating realized and potential carbon storage benefits from reforestation and afforestation under climate change: A case study of the Qinghai spruce forests in the Qilian Mountains northwestern China. Mitig. Adapt. Strateg. Glob. Chang. 2012, 18, 1257–1268. [Google Scholar] [CrossRef]
- Zhao, C.; Bie, Q.; Peng, H. Analysis of the Niche Space of Picea crassifolia on the Northern Slope of Qilian Mountains. Acta Ecol. Sin. 2010, 65, 113–121. [Google Scholar] [CrossRef]
- Weyant, J.; Azar, C.; Kainuma, M.; Kejun, J.; Nakicenovic, N.; Shukla, P.; Rovere, E.L.; Yohe, G. Report of 2.6Versus 2.9 Watts/m2 RCP Evaluation Panel; IPCC Secretariat: Geneva, Switzerland, 2009. [Google Scholar]
- Yang, X.Q.; Kushwaha, S.P.; Saran, S.; Xu, J.; Roy, P.S. MaxEnt modeling for predicting the potential distribution of medicinal plant Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 2002, 24, 38–49. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Chang. Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Khafaga, O.; Hatab, E.E.; Omar, K. Predicting the potential geographical distribution of Nepeta septemcrenata in Saint Katherine Protectorate, South Sinai, Egypt using Maxent. Acad. Arena 2011, 3, 45–50. [Google Scholar]
- Remya, K.; Ramachandran, A.; Jayakumar, S. Predicting the current and future suitable habitat distribution of myristica dactyloides, gaertn. using maxent model in the eastern ghats, india. Ecol. Eng. 2015, 82, 184–188. [Google Scholar] [CrossRef]
- Cardillo, M.; Mace, G.M.; Gittleman, J.L.; Jones, K.E.; Bielby, J.; Purvis, A. The predictability of extinction: Biological and external correlates of decline in mammals. Proc. Biol. Sci. 2008, 275, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Purvis, A.; Gittleman, J.L.; Cowlishaw, G.; Mace, G.M. Predicting extinction risk in declining species. Proc. Biol. Sci. 2000, 267, 1947–1952. [Google Scholar] [CrossRef]
- Profirio, L.L.; Harris, R.M.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef]
- Sinclair, S.J.; White, M.D.; Newell, G.R. How useful are species distribution models for managing biodiversity under future climates? Ecol. Soc. 2010, 15, 8. [Google Scholar] [CrossRef]
- Lozier, J.D.; Aniello, P.; Hickerson, M.J. Predicting the distribution of Sasquatch in western North America: Anything goes with ecological niche modelling. J. Biogeogr. 2009, 36, 1623–1627. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of Maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Xu, J.; Shrestha, K.K.; Kindt, R. Ensemble forecast of climate suitability for the Trans-Himalayan Nyctaginaceae species. Ecol. Modell. 2014, 282, 18–24. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le, L.G. Habitat suitability modelling and niche theory. J. Appl Ecol. 2008, 45, 1372–1381. [Google Scholar] [CrossRef]
- Guisan, A.; Graham, C.H.; Elith, J.; Huettmann, F. Sensitivity of predictive species distribution models to change in grain size. Divers Distrib. 2007, 13, 332–340. [Google Scholar] [CrossRef]
- Peng, S.; Zhao, C.; Xu, Z.; Ashiq, M.W. Restoration and conservation potential of destroyed Qinghai spruce (Picea crassifolia) forests in the Qilian Mountains of northwest China. Mitig. Adapt. Strateg. Glob. Chang. 2014, 21, 153–165. [Google Scholar] [CrossRef]
- Kosaka, Y.; Nakamura, H.; Watanabe, M.; Kimoto, M. Analysis on the dynamics of a wave-like tele- connection pattern along the summertime Asian jet based on a reanalysis dataset and climate model simulations. J. Meteor. Soc. Jpn. 2009, 87, 561–580. [Google Scholar] [CrossRef]
- Song, F.; Zhou, T.; Wang, L. Two modes of the Silk Road pattern and their interannual variability simulated by LASG/IAP AGCM SAMIL2.0. Adv. Atmos. Sci. 2013, 30, 908–921. [Google Scholar] [CrossRef]
- Song, F.; Zhou, T. FGOALS-s2 simulation of upper-level jet streams over East Asia: Mean state bias and synoptic-scale transient eddy activity. Adv. Atmos. Sci. 2013, 30, 739–753. [Google Scholar] [CrossRef]
- Monteith, K.L.; Klaver, R.W.; Hersey, K.R.; Holland, A.A.; Thomas, T.P.; Kauffman, M.J. Effects of climate and plant phenology on recruitment of moose at the southern extent of their range. Oecologia 2015, 178, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhao, C.; Feng, Z. A study of the impact of climate change on the potential distribution of Qinghai spruce (Picea crassifolia) in Qilian Mountains. Acta Ecol. Sin. 2009, 29, 278–285. [Google Scholar] [CrossRef]
- Zhou, T.; Song, F.; Chen, X. Historical evolution of global and regional surface air temperature simulated by FGOALS-s2 and FGOALS-g2: How reliable are the model results? Adv. Atmos. Sci. 2013, 30, 638–657. [Google Scholar] [CrossRef]
- Al-Qaddi, N.; Vessella, F.; Stephan, J.; Al-Eisawi, D.; Schirone, B. Current and future suitability areas of kermes oak (Quercuscoccifera L.) in the Levant under climate change. Reg. Environ. Chang. 2016, 17, 143–156. [Google Scholar] [CrossRef]
- Gong, M.; Guan, T.; Hou, M.; Liu, G.; Zhou, T. Hopes and challenges for giant panda conservation under climate change in the Qinling Mountains of China. Ecol. Evol. 2016, 7, 596–605. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecol. Lett. 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.E.; Ditommaso, A.; Wang, R.L.; Liang, K.M. Predicting the potential distribution of lantana camara l. under rcp scenarios using ISI-MIP models. Clim. Chang. 2016, 134, 193–208. [Google Scholar] [CrossRef]
- Cruz-Cárdenas, G.; López-Mata, L.; Silva, J.T.; Bernal-Santana, N.; Estrada-Godoy, F.; López-Sandoval, J.A. Potential distribution model of Pinaceae species under climate change scenarios in Michoacán. Revista Chapingo Serie Ciencias Forestales y del Ambiente 2016, 22, 135–148. [Google Scholar] [CrossRef]
- Zhang, Q.; Wen, J.; Chang, Z.Q.; Xie, C.X.; Song, J.Y. Evaluation and prediction of ecological suitability of medicinal plant American ginseng (panax quinquefolius). Chin. Herb. Med. 2018, 10, 80–85. [Google Scholar] [CrossRef]
- Hu, J.; Jiang, Z. Predicting the potential distribution of the endangered Przewalski’s gazelle. J. Zool. 2010, 282, 54–63. [Google Scholar] [CrossRef]
- Hu, J.; Jiang, Z. Climate Change Hastens the Conservation Urgency of an Endangered Ungulate. PLoS ONE 2011, 6, e22873. [Google Scholar] [CrossRef]
- Ren, Z.; Wang, D.; Ma, A.; Hwang, J.; Bennett, A.; Sturrock, H.J.W.; Fan, J.; Zhang, W.; Yang, D.; Feng, X.; et al. Predicting malaria vector distribution under climate change scenarios in China: Challenges for malaria elimination. Sci. Rep. 2016, 6, 20604. [Google Scholar] [CrossRef]
- Tuanmu, M.N.; Viña, A.; Winkler, J.A.; Li, Y.; Xu, W.; Ouyang, Z.; Liu, J. Climate-change impacts on understorey bamboo species and giant pandas in China’s Qinling Mountains. Nat. Clim. Chang. 2012, 3, 249–253. [Google Scholar] [CrossRef]
- Araújo, M.B.; Luoto, M. The importance of biotic interactions for modelling species distributions under climate change. Glob. Ecol. Biogeogr. 2007, 16, 743–753. [Google Scholar] [CrossRef]
- Boulangeat, I.; Gravel, D.; Thuiller, W. Accounting for dispersal and biotic interactions to disentangle the drivers of species distributions and their abundances. Ecol. Lett. 2012, 15, 584–593. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variables | Unit | Current | RCP2.6 | RCP4.5 | RCP6.0 | RCP8.5 |
|---|---|---|---|---|---|---|---|
| Bio1 | Annual mean temperature | °C | 16.0 | 15.5 | 22.3 | 23.7 | 27.0 |
| Bio2 | Mean diurnal range (mean of monthly (max temp-min temp)) | °C | 10.4 | 8.0 | 13.4 | 13.5 | 8.7 |
| Bio3 | Isothermality (Bio2/Bio7) (*100) | 6.0 | 6.1 | 5.0 | 4.3 | 8.5 | |
| Bio4 | Temperature seasonality (standard deviation×100) | C of V | |||||
| Bio5 | Maximum temperature of warmest month | °C | |||||
| Bio6 | Minimum temperature of coldest month | °C | |||||
| Bio7 | Temperature annual range (Bio5–Bio6) | °C | |||||
| Bio8 | Mean temperature of wettest quarter | °C | 24.4 | 24.0 | 19.9 | 17.9 | 17.5 |
| Bio9 | Mean temperature of driest season | °C | |||||
| Bio10 | Mean temperature of warmest season | °C | |||||
| Bio11 | Mean temperature of coldest season | °C | |||||
| Bio12 | Annual precipitation | mm | |||||
| Bio13 | Precipitation of wettest period | mm | |||||
| Bio14 | Precipitation of driest period | mm | |||||
| Bio15 | Precipitation seasonality | C of V | |||||
| Bio16 | Precipitation of wettest season | mm | |||||
| Bio17 | Precipitation of driest season | mm | |||||
| Bio18 | Precipitation of warmest season | mm | |||||
| Bio19 | Precipitation of coldest season | mm | |||||
| Ele | Elevation | m | 29.2 | 30.1 | 23.5 | 23.7 | 21.3 |
| Slo | Slope | º | 11.5 | 14.1 | 13.8 | 14.7 | 14.7 |
| Asp | Aspect | º | 2.5 | 2.3 | 2.1 | 2.2 | 2.4 |
| Variables | Current | RCP2.6 | RCP4.5 | RCP6.0 | RCP8.5 | ||||
|---|---|---|---|---|---|---|---|---|---|
| mean | mean | Change rate (%) | mean | Change rate (%) | mean | Change rate (%) | mean | Change rate (%) | |
| Bio1 | −16.20 | 3.00 | −118.52 | 10.50 | −164.81 | 11.50 | −170.99 | 24.8 | −253.09 |
| Bio2 | 126.51 | 126.65 | 0.11 | 126.25 | −0.21 | 125.77 | −0.58 | 125.11 | −1.11 |
| Bio3 | 32.30 | 32.02 | −0.87 | 31.74 | −1.73 | 31.56 | −2.29 | 31.25 | −3.25 |
| Bio8 | 94.59 | 112.13 | 18.54 | 122.20 | 29.19 | 123.16 | 30.20 | 137.40 | 45.26 |
| AUC | Current | RCP2.6 | RCP4.5 | RCP6.0 | RCP8.5 |
|---|---|---|---|---|---|
| AUC (Training data) | 0.980 | 0.981 | 0.982 | 0.981 | 0.981 |
| AUC (Test data) | 0.968 | 0.968 | 0.969 | 0.968 | 0.964 |
| Current | RCP2.6 | RCP4.5 | RCP6.0 | RCP8.0 | |
|---|---|---|---|---|---|
| Unsuitable area (km2) | 16,362.6 | 16,446.4 | 16,481.0 | 16,498.9 | 17,315.2 |
| Low suitability area (km2) | 2994.1 | 3191.0 | 3189.8 | 3117.8 | 3191.4 |
| Moderate suitability area (km2) | 4250.7 | 4126.5 | 4040.8 | 3952.9 | 31913.6 |
| High suitability area (km2) | 1312.0 | 1155.6 | 1207.9 | 1349.9 | 1221.3 |
| Total suitability area (km2) | 8556.8 | 8473.1 | 8438.5 | 8420.6 | 7604.2 |
| Change area (km2) | −83.8 | −118.4 | −136.3 | −857.6 | |
| Percent (%) | 34.3 | 34.0 | 33.9 | 33.8 | 30.5 |
| Classes | Current | RCP2.6 | RCP4.5 | RCP6.0 | RCP8.5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Area (km2) | Percent (%) | Area (km2) | Percent (%) | Area (km2) | Percent (%) | Area (km2) | Percent (%) | Area (km2) | Percent (%) | |
| Unsuitable | 60.7 | 4.3 | 138.2 | 9.7 | 137.7 | 9.7 | 138.8 | 9.8 | 185.0 | 13.0 |
| Moderate suitability | 323.5 | 22.7 | 251.6 | 17.7 | 262.5 | 18.5 | 259.0 | 18.2 | 199.4 | 14.4 |
| High suitability | 654.2 | 46.0 | 679.5 | 47.8 | 647.4 | 45.5 | 633.5 | 44.5 | 644.3 | 45.3 |
| Total suitability area | 383.7 | 27.0 | 352.8 | 24.8 | 374.5 | 26.3 | 390.7 | 27.5 | 388.7 | 27.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, Z.; Zhao, C.; Liu, J.; Gao, Y.; Zang, F.; Guo, Z.; Mao, Y.; Wang, L. Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains. Forests 2019, 10, 62. https://doi.org/10.3390/f10010062
Rong Z, Zhao C, Liu J, Gao Y, Zang F, Guo Z, Mao Y, Wang L. Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains. Forests. 2019; 10(1):62. https://doi.org/10.3390/f10010062
Chicago/Turabian StyleRong, Zhanlei, Chuanyan Zhao, Junjie Liu, Yunfei Gao, Fei Zang, Zhaoxia Guo, Yahua Mao, and Ling Wang. 2019. "Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains" Forests 10, no. 1: 62. https://doi.org/10.3390/f10010062
APA StyleRong, Z., Zhao, C., Liu, J., Gao, Y., Zang, F., Guo, Z., Mao, Y., & Wang, L. (2019). Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains. Forests, 10(1), 62. https://doi.org/10.3390/f10010062