An Algorithm for Creating a Synaptic Cleft Digital Phantom Suitable for Further Numerical Modeling



Abstract
1. Introduction
2. Materials and Methods
2.1. Forming of Pre- and Postsynaptic Endings and an Auxiliary Plane
2.2. Synapse Phantom
2.3. Synaptic Cleft
2.4. Astrocytic Leaflets
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, Y.; Yang, X. A time two-grid difference method for nonlinear generalized viscous Burgers’ equation. J. Math. Chem. 2024, 62, 1323–1356. [Google Scholar] [CrossRef]
- Shi, Y.; Yang, X.; Zhang, Z. Construction of a new time-space two-grid method and its solution for the generalized Burgers’ equation. Appl. Math. Lett. 2024, 158, 109244. [Google Scholar] [CrossRef]
- Shi, Y.; Yang, X. Pointwise error estimate of conservative difference scheme for supergeneralized viscous Burgers’ equation. Electron. Res. Arch. 2024, 32, 1471–1497. [Google Scholar] [CrossRef]
- Li, C.; Zhang, H.; Yang, X. A new nonlinear compact difference scheme for a fourth-order nonlinear Burgers type equation with a weakly singular kernel. J. Appl. Math. Comp. 2024, 70, 2045–2077. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, H.; Yang, X.; Wang, F. A second-order finite difference method for the multi-term fourth-order integral–differential equations on graded meshes. Comput. Appl. Math. 2022, 41, 313. [Google Scholar] [CrossRef]
- Yang, X.; Qiu, W.; Chen, H.; Zhang, H. Second-order BDF ADI Galerkin finite element method for the evolutionary equation with a nonlocal term in three-dimensional space. Appl. Numer. Math. 2022, 172, 497–513. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, H.; Yang, X. CN ADI fast algorithm on non-uniform meshes for the three-dimensional nonlocal evolution equation with multi-memory kernels in viscoelastic dynamics. Appl. Math. Comput. 2024, 474, 128680. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z. Superconvergence Analysis of a Robust Orthogonal Gauss Collocation Method for 2D Fourth-Order Subdiffusion Equations. J. Sci. Comput. 2024, 100, 62. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, X.; Xu, D. Unconditional convergence of linearized orthogonal spline collocation algorithm for semilinear subdiffusion equation with nonsmooth solution. Numer. Methods Partial. Differ. Equ. 2021, 37, 1361–1373. [Google Scholar] [CrossRef]
- Yang, X.; Qiu, W.; Zhang, H.; Tang, L. An efficient alternating direction implicit finite difference scheme for the three-dimensional time-fractional telegraph equation. Comput. Math. Appl. 2021, 102, 233–247. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, H.; Zhou, Z.; Yang, X. A fast compact finite difference scheme for the fourth-order diffusion-wave equation. Int. J. Comput. Math. 2024, 101, 170–193. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z. On conservative, positivity preserving, nonlinear FV scheme on distorted meshes for the multi-term nonlocal Nagumo-type equations. Appl. Math. Lett. 2024, 150, 108972. [Google Scholar] [CrossRef]
- Wang, F.; Yang, X.; Zhang, H.; Wu, L. A time two-grid algorithm for the two dimensional nonlinear fractional PIDE with a weakly singular kernel. Math. Comput. Simul. 2022, 199, 38–59. [Google Scholar] [CrossRef]
- Yang, X.; Wu, L.; Zhang, H. A space-time spectral order sinc-collocation method for the fourth-order nonlocal heat model arising in viscoelasticity. Appl. Math. Comput. 2023, 457, 128192. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, X.; Wang, F.; Yang, X. The time two-grid algorithm combined with difference scheme for 2D nonlocal nonlinear wave equation. J. Appl. Math. Comp. 2024, 70, 1127–1151. [Google Scholar] [CrossRef]
- Appadu, A.R.; Lebelo, R.S.; Gidey, H.H.; Inan, B. Editorial: Modeling and numerical simulations with differential equations in mathematical biology, medicine, and the environment. Front. Appl. Math. Stat. 2023, 9, 1150552. [Google Scholar] [CrossRef]
- Ehrhardt, M.; Sánchez, L.J.; Villanueva Micó, R.J. Numerical methods and mathematical modelling in biology, medicine and social sciences. Int. J. Comput. Math. 2014, 91, 176–178. [Google Scholar] [CrossRef]
- Huppert, A.; Katriel, G. Mathematical modelling and prediction in infectious disease epidemiology. Clin. Microbiol. Infect. 2013, 19, 999–1005. [Google Scholar] [CrossRef]
- Kucharski, A.J.; Russell, T.W.; Diamond, C.; Liu, Y.; Edmunds, J.; Funk, S.; Eggo, R.M.; Sun, F.; Jit, M.; Munday, J.D.; et al. Early dynamics of transmission and control of COVID-19: A mathematical modelling study. Lancet Infect. Dis. 2020, 20, 553–558. [Google Scholar] [CrossRef]
- Saldaña, F.; Velasco-Hernández, J.X. Modeling the COVID-19 pandemic: A primer and overview of mathematical epidemiology. SeMA J. 2022, 79, 225–251. [Google Scholar] [CrossRef]
- Deplazes, E. Molecular simulations of disulfide-rich venom peptides with ion channels and membranes. Molecules 2017, 22, 362. [Google Scholar] [CrossRef] [PubMed]
- Cusimano, N.; Gizzi, A.; Fenton, F.H.; Filippi, S.; Gerardo-Giorda, L. Key aspects for effective mathematical modelling of fractional-diffusion in cardiac electrophysiology: A quantitative study. Commun. Nonlinear Sci. Numer. Simul. 2020, 84, 105152. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, D.E.; Castro, S.; Gizzi, A. Computational modeling of non-linear diffusion in cardiac electrophysiology: A novel porous-medium approach. Comput. Methods Appl. Mech. Eng. 2016, 300, 70–83. [Google Scholar] [CrossRef]
- Nartsissov, Y.R. The Effect of Flux Dysconnectivity Functions on Concentration Gradients Changes in a Multicomponent Model of Convectional Reaction-Diffusion by the Example of a Neurovascular Unit. Defect Diffus. Forum 2021, 413, 19–28. [Google Scholar] [CrossRef]
- Nartsissov, Y.R. Application of a multicomponent model of convectional reaction-diffusion to description of glucose gradients in a neurovascular unit. Front. Physiol. 2022, 13, 843473. [Google Scholar] [CrossRef]
- Ulman, V.; Svoboda, D.; Nykter, M.; Kozubek, M.; Ruusuvuori, P. Virtual cell imaging: A review on simulation methods employed in image cytometry. Cytom. Part A 2016, 89, 1057–1072. [Google Scholar] [CrossRef] [PubMed]
- Mishchenko, Y.; Hu, T.; Spacek, J.; Mendenhall, J.; Harris, K.M.; Chklovskii, D.B. Ultrastructural analysis of hippocampal neuropil from the connectomics perspective. Neuron 2010, 67, 1009–1020. [Google Scholar] [CrossRef]
- Radojević, M.; Meijering, E. Automated Neuron Reconstruction from 3D Fluorescence Microscopy Images Using Sequential Monte Carlo Estimation. Neuroinformatics 2019, 17, 423–442. [Google Scholar] [CrossRef]
- Nartsissov, Y.R. A novel algorithm of the digital nervous tissue phantom creation based on 3D Voronoi diagram application. J. Phys. Conf. Ser. 2021, 2090, 012009. [Google Scholar] [CrossRef]
- Nartsissov, Y.R.; Ivontsin, L.A. Mathematical Modelling of Physiological Effects Caused by a Glycine Receptors Post-Synaptic Density Spatial Polymorphism. Mathematics 2023, 11, 2499. [Google Scholar] [CrossRef]
- Nartsissov, Y.R.; Zagubnaya, O.A. A digital 3D reconstruction of a synaptic cleft which can be used for further modeling of neuromediators convectional diffusion in a nervous tissue. AIP Conf. Proc. 2023, 2872, 120003. [Google Scholar] [CrossRef]
- Südhof, T.C. The cell biology of synapse formation. J. Cell Biol. 2021, 220, e202103052. [Google Scholar] [CrossRef] [PubMed]
- Grewer, C.; Gameiro, A.; Rauen, T. SLC1 glutamate transporters. Pflug. Arch. Eur. J. Physiol. 2014, 466, 3–24. [Google Scholar] [CrossRef]
- Armbruster, M.; Hanson, E.; Dulla, C.G. Glutamate clearance is locally modulated by presynaptic neuronal activity in the cerebral cortex. J. Neurosci. 2016, 36, 10404–10415. [Google Scholar] [CrossRef]
- Belov Kirdajova, D.; Kriska, J.; Tureckova, J.; Anderova, M. Ischemia-Triggered Glutamate Excitotoxicity from the Perspective of Glial Cells. Front. Cell. Neurosci. 2020, 14, 51. [Google Scholar] [CrossRef] [PubMed]
- Calvetti, D.; Cheng, Y.; Somersalo, E. A spatially distributed computational model of brain cellular metabolism. J. Theor. Biol. 2015, 376, 48–65. [Google Scholar] [CrossRef]
- Idumah, G.; Somersalo, E.; Calvetti, D. A Spatially Distributed Model of Brain Metabolism Highlights the Role of Diffusion in Brain Energy Metabolism. J. Theor. Biol. 2022, 572, 111567. [Google Scholar] [CrossRef]
- Kinney, J.P.; Spacek, J.; Bartol, T.M.; Bajaj, C.L.; Harris, K.M.; Sejnowski, T.J. Extracellular sheets and tunnels modulate glutamate diffusion in hippocampal neuropil. J. Comp. Neurol. 2013, 521, 448–464. [Google Scholar] [CrossRef]
- Rusakov, D.A.; Kullmann, D.M. Geometric and viscous components of the tortuosity of the extracellular space in the brain. Proc. Natl. Acad. Sci. USA 1998, 95, 8975–8980. [Google Scholar] [CrossRef]
- Borges, R.; Gu, C.; Machado, J.D.; Ewing, A.G. The dynamic nature of exocytosis from large secretory vesicles. A view from electrochemistry and imaging. Cell Calcium 2023, 110, 102699. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, Q.; Liu, B.; Li, Y.; Wu, X.; Kuo, S.; Zheng, L.; Wang, C.; Zhu, F.; Zhou, Z. Dynamin 1 restrains vesicular release to a subquantal mode in mammalian adrenal chromaffin cells. J. Neurosci. 2019, 39, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Limbach, C.; Laue, M.M.; Wang, X.; Hu, B.; Thiede, N.; Hultqvist, G.; Kilimann, M.W. Molecular in situ topology of Aczonin/Piccolo and associated proteins at the mammalian neurotransmitter release site. Proc. Natl. Acad. Sci. USA 2011, 108, E392–E401. [Google Scholar] [CrossRef]
- Colombo, M.N.; Maiellano, G.; Putignano, S.; Scandella, L.; Francolini, M. Comparative 2D and 3D ultrastructural analyses of dendritic spines from CA1 pyramidal neurons in the mouse hippocampus. Int. J. Mol. Sci. 2021, 22, 1188. [Google Scholar] [CrossRef] [PubMed]
- Aina, B.-S.; Tim, S.H.; Mandy, S.J.K.; Adrian, N.; Huibert, D.M.; Baljit, S.K.; Rogier, M.; August, B.S.; Mark, H.G.V. Proximity of astrocyte leaflets to the synapse determines memory strength. bioRxiv 2022. [CrossRef]
- Toman, M.; Wade, J.J.; Verkhratsky, A.; Dallas, M.; Bithell, A.; Flanagan, B.; Harkin, J.; McDaid, L. The influence of astrocytic leaflet motility on ionic signalling and homeostasis at active synapses. Sci. Rep. 2023, 13, 3050. [Google Scholar] [CrossRef]
- Badia-Soteras, A.; Heistek, T.S.; Kater, M.S.J.; Mak, A.; Negrean, A.; van den Oever, M.C.; Mansvelder, H.D.; Khakh, B.S.; Min, R.; Smit, A.B.; et al. Retraction of Astrocyte Leaflets from the Synapse Enhances Fear Memory. Biol. Psychiatry 2023, 94, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Gavrilov, N.; Golyagina, I.; Brazhe, A.; Scimemi, A.; Turlapov, V.; Semyanov, A. Astrocytic coverage of dendritic spines, dendritic shafts, and axonal boutons in hippocampal neuropil. Front. Cell. Neurosci. 2018, 12, 248. [Google Scholar] [CrossRef]
- Ventura, R.; Harris, K.M. Three-dimensional relationships between hippocampal synapses and astrocytes. J. Neurosci. 1999, 19, 6897–6906. [Google Scholar] [CrossRef]
- Ponuwei, G.A. A glimpse of the ERM proteins. J. Biomed. Sci. 2016, 23, 35. [Google Scholar] [CrossRef]
- Schacke, S.; Kirkpatrick, J.; Stocksdale, A.; Bauer, R.; Hagel, C.; Riecken, L.B.; Morrison, H. Ezrin deficiency triggers glial fibrillary acidic protein upregulation and a distinct reactive astrocyte phenotype. GLIA 2022, 70, 2309–2329. [Google Scholar] [CrossRef]
- Popov, A.; Brazhe, N.; Morozova, K.; Yashin, K.; Bychkov, M.; Nosova, O.; Sutyagina, O.; Brazhe, A.; Parshina, E.; Li, L.; et al. Mitochondrial malfunction and atrophy of astrocytes in the aged human cerebral cortex. Nat. Commun. 2023, 14, 8380. [Google Scholar] [CrossRef] [PubMed]
- Hamada, K.; Shimizu, T.; Matsui, T.; Tsukita, S.; Hakoshima, T. Structural basis of the membrane-targeting and unmasking mechanisms of the radixin FERM domain. EMBO J. 2000, 19, 4449–4462. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef]
- Fehon, R.G.; McClatchey, A.I.; Bretscher, A. Organizing the cell cortex: The role of ERM proteins. Nat. Rev. Mol. Cell Biol. 2010, 11, 276–287. [Google Scholar] [CrossRef]
- Fomitcheva, I.V.; Sword, J.; Shi, Y.; Kirov, S.A. Plasticity of perisynaptic astroglia during ischemia-induced spreading depolarization. Cereb. Cortex 2023, 33, 5469–5483. [Google Scholar] [CrossRef]
- Rusakov, D.A. The role of perisynaptic glial sheaths in glutamate spillover and extracellular Ca2+ depletion. Biophys. J. 2001, 81, 1947–1959. [Google Scholar] [CrossRef]
- Zheng, K.; Scimemi, A.; Rusakov, D.A. Receptor actions of synaptically released glutamate: The role of transporters on the scale from nanometers to microns. Biophys. J. 2008, 95, 4584–4596. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.S. Neuronal glutamate transporters limit activation of NMDA receptors by neurotransmitter spillover on CA1 pyramidal cells. J. Neurosci. 2001, 21, 8328–8338. [Google Scholar] [CrossRef]
- Freche, D.; Pannasch, U.; Rouach, N.; Holcman, D. Synapse geometry and receptor dynamics modulate synaptic strength. PLoS ONE 2011, 6, e25122. [Google Scholar] [CrossRef] [PubMed]
- Herde, M.K.; Bohmbach, K.; Domingos, C.; Vana, N.; Komorowska-Müller, J.A.; Passlick, S.; Schwarz, I.; Jackson, C.J.; Dietrich, D.; Schwarz, M.K.; et al. Local efficacy of glutamate uptake decreases with synapse size. Cell Rep. 2020, 32, 108182. [Google Scholar] [CrossRef]
- Rǎdulescu, A.R.; Todd, G.C.; Williams, C.L.; Bennink, B.A.; Lemus, A.A.; Chesbro, H.E.; Bourgeois, J.R.; Kopec, A.M.; Zuloaga, D.G.; Scimemi, A. Estimating the glutamate transporter surface density in distinct sub-cellular compartments of mouse hippocampal astrocytes. PLoS Comput. Biol. 2022, 18, e1009845. [Google Scholar] [CrossRef] [PubMed]
- Feghhi, T.; Hernandez, R.X.; Stawarski, M.; Thomas, C.I.; Kamasawa, N.; Lau, A.W.C.; Macleod, G.T. Computational modeling predicts ephemeral acidic microdomains in the glutamatergic synaptic cleft. Biophys. J. 2021, 120, 5575–5591. [Google Scholar] [CrossRef]
- Kleinle, J.; Vogt, K.; Lüscher, H.R.; Müller, L.; Senn, W.; Wyler, K.; Streit, J. Transmitter concentration profiles in the synaptic cleft: An analytical model of release and diffusion. Biophys. J. 1996, 71, 2413–2426. [Google Scholar] [CrossRef]
- Nielsen, T.A.; DiGregorio, D.A.; Silver, R.A. Modulation of glutamate mobility reveals the mechanism underlying slow-rising AMPAR EPSCs and the diffusion coefficient in the synaptic cleft. Neuron 2004, 42, 757–771. [Google Scholar] [CrossRef]
- McCauley, J.P.; Petroccione, M.A.; D’Brant, L.Y.; Todd, G.C.; Affinnih, N.; Wisnoski, J.J.; Zahid, S.; Shree, S.; Sousa, A.A.; De Guzman, R.M.; et al. Circadian Modulation of Neurons and Astrocytes Controls Synaptic Plasticity in Hippocampal Area CA1. Cell Rep. 2020, 33, 108255. [Google Scholar] [CrossRef]
- Sweeney, A.M.; Fleming, K.E.; McCauley, J.P.; Rodriguez, M.F.; Martin, E.T.; Sousa, A.A.; Leapman, R.D.; Scimemi, A. PAR1 activation induces rapid changes in glutamate uptake and astrocyte morphology. Sci. Rep. 2017, 7, srep43606. [Google Scholar] [CrossRef]
- Antunes, G.; de Souza, F.M.S. 3d modeling of dendritic spines with synaptic plasticity. J. Vis. Exp. 2020, 2020, e60896. [Google Scholar] [CrossRef]
- Héja, L.; Szabó, Z.; Péter, M.; Kardos, J. Spontaneous Ca2+ Fluctuations Arise in Thin Astrocytic Processes With Real 3D Geometry. Front. Cell. Neurosci. 2021, 15, 617989. [Google Scholar] [CrossRef] [PubMed]
- Savtchenko, L.P.; Rusakov, D.A. Increased Extrasynaptic Glutamate Escape in Stochastically Shaped Probabilistic Synaptic Environment. Biomedicines 2022, 10, 2406. [Google Scholar] [CrossRef]
- Nartsissov, Y.R. A spatial classification applied to convectional reaction-diffusion boundary problems basing on a geometrical polymorphism of biological objects. J. Phys. Conf. Ser. 2024, 2701, 012084. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, H.; Zhang, Q.; Yuan, G.; Sheng, Z. The finite volume scheme preserving maximum principle for two-dimensional time-fractional Fokker–Planck equations on distorted meshes. Appl. Math. Lett. 2019, 97, 99–106. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, H.; Zhang, Q.; Yuan, G. Simple positivity-preserving nonlinear finite volume scheme for subdiffusion equations on general non-conforming distorted meshes. Nonlinear Dyn. 2022, 108, 3859–3886. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, H. The uniform l1 long-time behavior of time discretization for time-fractional partial differential equations with nonsmooth data. Appl. Math. Lett. 2022, 124, 107644. [Google Scholar] [CrossRef]







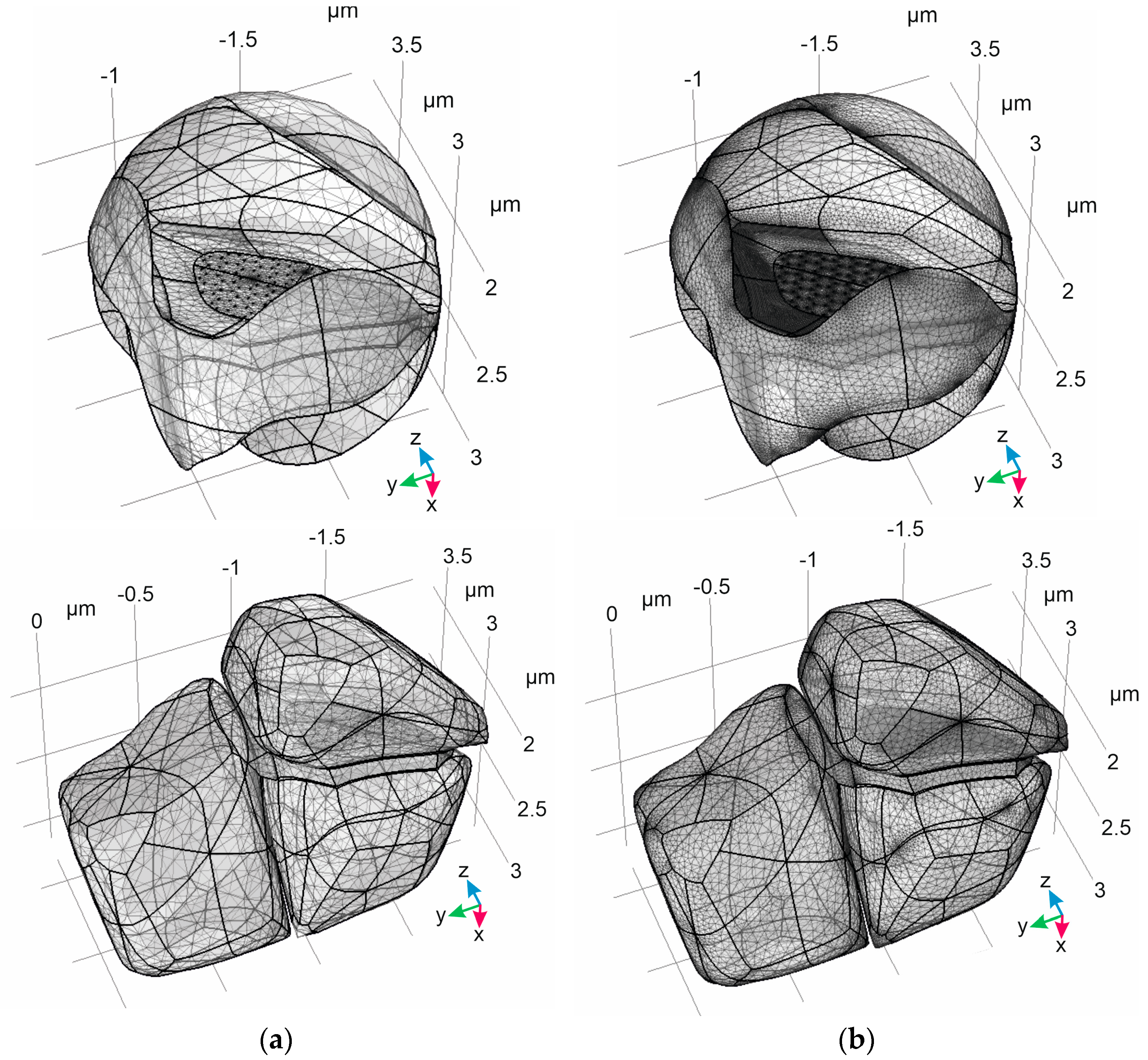

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
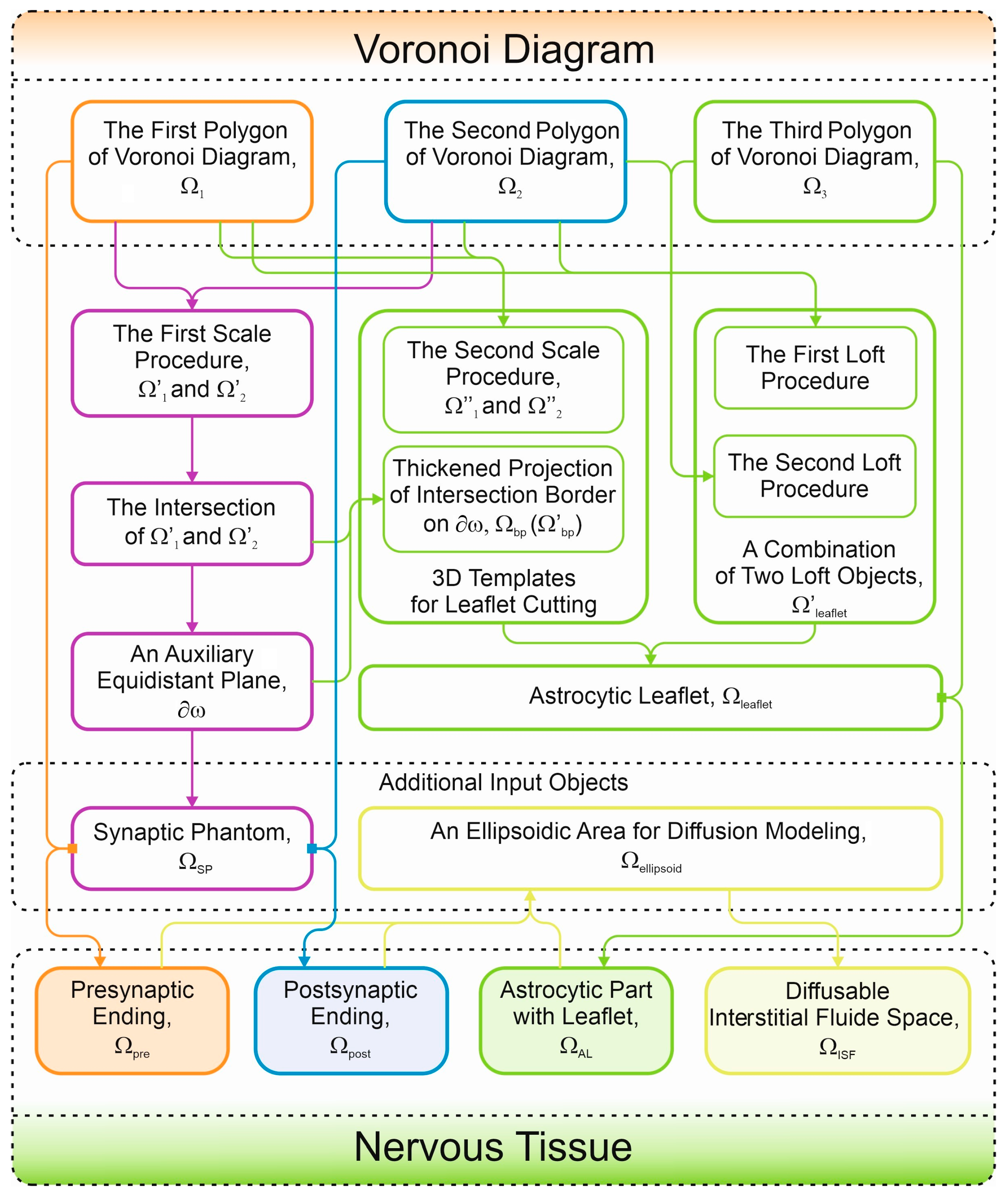
| The Object | Description | Comments |
|---|---|---|
| Input Voronoi diagram bodies | ||
| Ω1 | The first Voronoi diagram body | This object can be chosen fluently |
| Ω2 | The second Voronoi diagram body | This object is selected as a close neighbor of Ω1 |
| Ω3 | The third Voronoi diagram body | The third object lies near the contact area of Ω1 and Ω2 |
| Auxiliary objects, planes, and surfaces | ||
| Ω′1 and Ω′2 | The auxiliary scaled Ω1 and Ω2 for astrocytic leaflet tip cutting | Ω′i = γ·Ωi |
| Ω′1–2 | Ω1–2 edge projection on auxiliary plane is used for leaflet tip cutting | Ω′1–2 = Ω′1 Ո Ω′2 |
| 𝜕ω | An auxiliary plane | 𝜕Ωbp ⊂ 𝜕ω; 𝜕Ω′bp ⊂ 𝜕ω |
| Ω′leaflet | A combination of two loft objects for leaflet creation | An intermediate leaflet object (see in text) |
| Ω″1 and Ω″2 | The auxiliary scaled Ω1 and Ω2 for astrocytic leaflet cutting | Ω″i = λ·Ωi |
| Ωbp | A 3D template for leaflet cutting | Ωbp = 𝜕h·𝜕Ωbp |
| Ω′bp | A 3D template for leaflet cutting in the case of bigger synaptic phantoms | Ω′bp = 𝜕h·𝜕Ω′bp |
| ΩSP | Synapse phantom | Determined by the experimental data object composed of upper and lower hemispheres joined by a cylindrical linker |
| Ωellipsoid | An ellipsoid used for indication of considered ISF borders | The geometric parameters of ellipsoid have a common restriction (see in text) |
| 𝜕Ωbp | A solid type of interpolation curve of Ω1–2 edge projection on auxiliary plane 𝜕ω | 𝜕Ωbp ⊂ 𝜕ω |
| 𝜕Ω′bp | A solid type of combined interpolation curve on auxiliary plane 𝜕ω | 𝜕Ω′bp ⊂ 𝜕ω |
| 𝜕Ω1–2 | The presynaptic membrane | Enclosing first two Voronoi bodies contact area which faces Ω1 |
| 𝜕Ω2–1 | The postsynaptic membrane | Enclosing first two Voronoi bodies contact area which faces Ω2 |
| Digital domains corresponding to nervous tissue’s cells and ISF | ||
| Ωpre | The first body with embedded synapse phantom track for presynaptic modeling | Ωpre = Ω1\ΩSP |
| Ωpost | The second body with embedded synapse phantom track for postsynaptic modeling | Ωpost = Ω2\ΩSP |
| Ωleaflet | A small protrusion of Ω3 toward the synaptic cleft for astrocytic leaflet modeling | Ωleaflet = Ω′leaflet\(Ω′1 U Ω′2 U Ωbp), or Ωleaflet = Ω′leaflet\(Ω′1 U Ω′2 U Ωbp U Ω′synapse) (see in the text) |
| ΩAL | A complex body for astrocyte modeling | ΩAL = Ω3 U Ωleaflet |
| ΩISF | The final complex body which limits neurotransmitter diffusion area includes synaptic cleft space and marked 𝜕ΩAZ, 𝜕Ωifusion, and 𝜕ΩPSD | ΩISF = Ωellipsoid\(Ωpre U Ωpost U ΩAL) |
| Digital surfaces coinciding with essential biological areas | ||
| 𝜕ΩAZ | The presynaptic membrane area on Ωpre, which contains Ca2+-channels and vesicular emitting protein machinery | The track after Ω1\ΩSP Boolean operation |
| 𝜕Ωifusion | The places of plausible vesicular fusion pores on 𝜕ΩAZ | 𝜕Ωifusion diameter is approximately 9 nm and has consistent pattern on 𝜕ΩAZ (see in the text) |
| 𝜕ΩPSD | The PSD area on Ωpost | The track after Ω2\ΩSP Boolean operation |
| 𝜕ΩAZ Properties | RSP = 50 nm | RSP = 130 nm | RSP = 200 nm | RSP = 300 nm | RSP = 450 nm | |
|---|---|---|---|---|---|---|
| HSP = 15 nm | 𝜕ΩAZ, µm2 | 0.007 | 0.045 | 0.090 | 0.149 | 0.200 |
| 𝜕ΩPSD, µm2 | ||||||
| HSP = 20 nm | 𝜕ΩAZ, µm2 | 0.008 | 0.049 | 0.104 | 0.184 | 0.259 |
| 𝜕ΩPSD, µm2 | ||||||
| HSP = 25 nm | 𝜕ΩAZ, µm2 | 0.008 | 0.051 | 0.112 | 0.206 | 0.302 |
| 𝜕ΩPSD, µm2 | ||||||
| HSP = 30 nm | 𝜕ΩAZ, µm2 | 0.009 | 0.052 | 0.116 | 0.221 | 0.335 |
| 𝜕ΩPSD, µm2 | ||||||
| Ωleaflet Properties | HSP = 15 nm | HSP = 20 nm | HSP = 25 nm | HSP = 30 nm | |
|---|---|---|---|---|---|
| RSP = 50 nm RSP = 130 nm RSP = 200 nm | SVR24, µm−1 | 47.2 | |||
| SVR38, µm−1 | 50.9 | ||||
| SVR48, µm−1 | 54.2 | ||||
| SVR62, µm−1 | 57.0 | ||||
| RSP = 300 nm | SVR24, µm−1 | 44.3 | 44.3 | ||
| SVR38, µm−1 | 47.3 | 47.3 | |||
| SVR48, µm−1 | 49.8 | 49.7 | |||
| SVR62, µm−1 | 52.1 | 52.7 | |||
| RSP = 450 nm | SVR24, µm−1 | 43.8 | 43.1 | 42.8 | 42.6 |
| SVR38, µm−1 | 46.9 | 46.4 | 45.98 | 45.7 | |
| SVR48, µm−1 | 49.4 | 48.9 | 48.5 | 48.4 | |
| SVR62, µm−1 | 51.7 | 51.3 | 50.8 | 50.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zagubnaya, O.A.; Nartsissov, Y.R. An Algorithm for Creating a Synaptic Cleft Digital Phantom Suitable for Further Numerical Modeling. Algorithms 2024, 17, 451. https://doi.org/10.3390/a17100451
Zagubnaya OA, Nartsissov YR. An Algorithm for Creating a Synaptic Cleft Digital Phantom Suitable for Further Numerical Modeling. Algorithms. 2024; 17(10):451. https://doi.org/10.3390/a17100451
Chicago/Turabian StyleZagubnaya, Olga A., and Yaroslav R. Nartsissov. 2024. "An Algorithm for Creating a Synaptic Cleft Digital Phantom Suitable for Further Numerical Modeling" Algorithms 17, no. 10: 451. https://doi.org/10.3390/a17100451
APA StyleZagubnaya, O. A., & Nartsissov, Y. R. (2024). An Algorithm for Creating a Synaptic Cleft Digital Phantom Suitable for Further Numerical Modeling. Algorithms, 17(10), 451. https://doi.org/10.3390/a17100451