
A New Approach to Identifying Sorghum Hybrids Using UAV Imagery Using Multispectral Signature and Machine Learning

 ,
,  , , ,
, , ,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
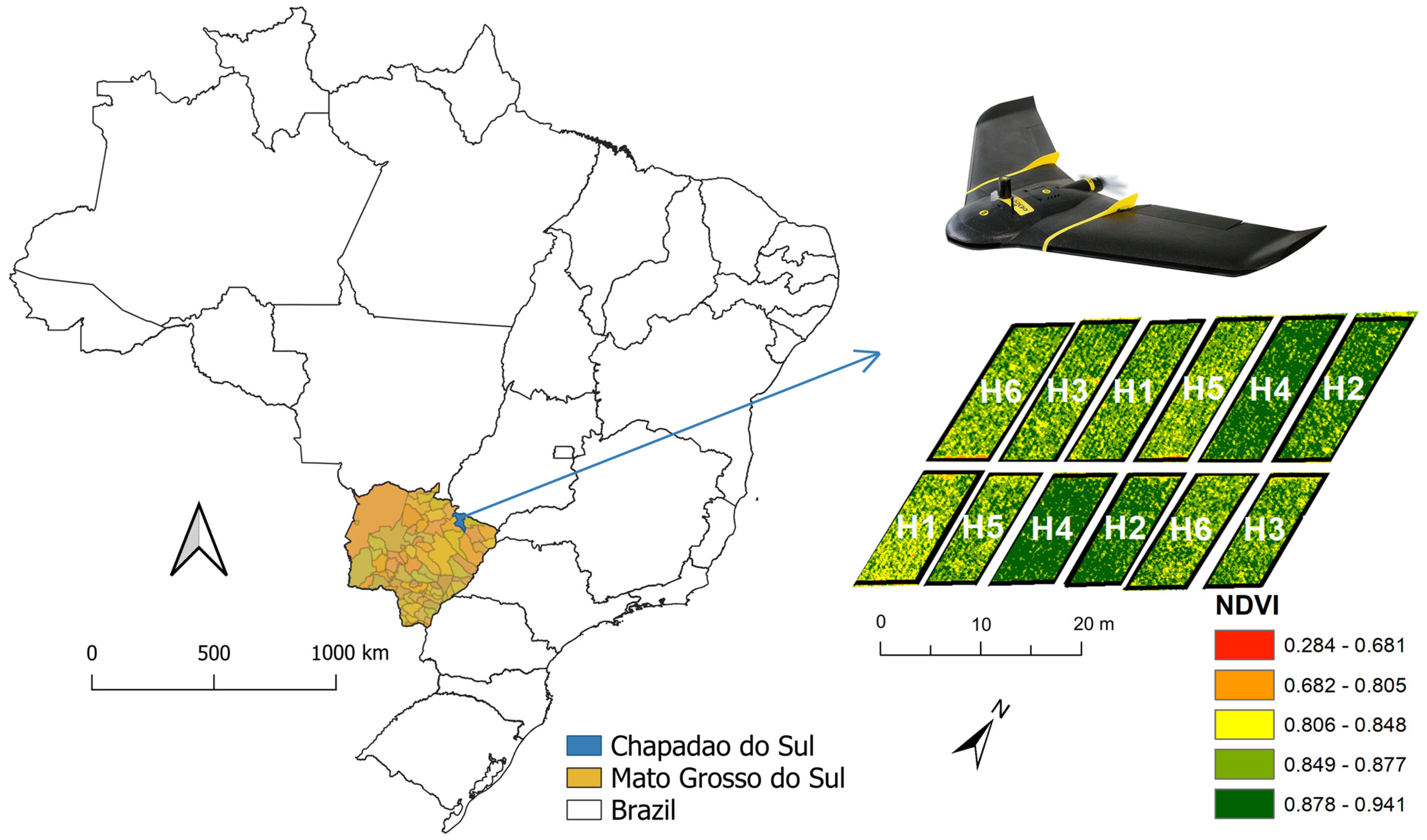
2.1. Field Experiment
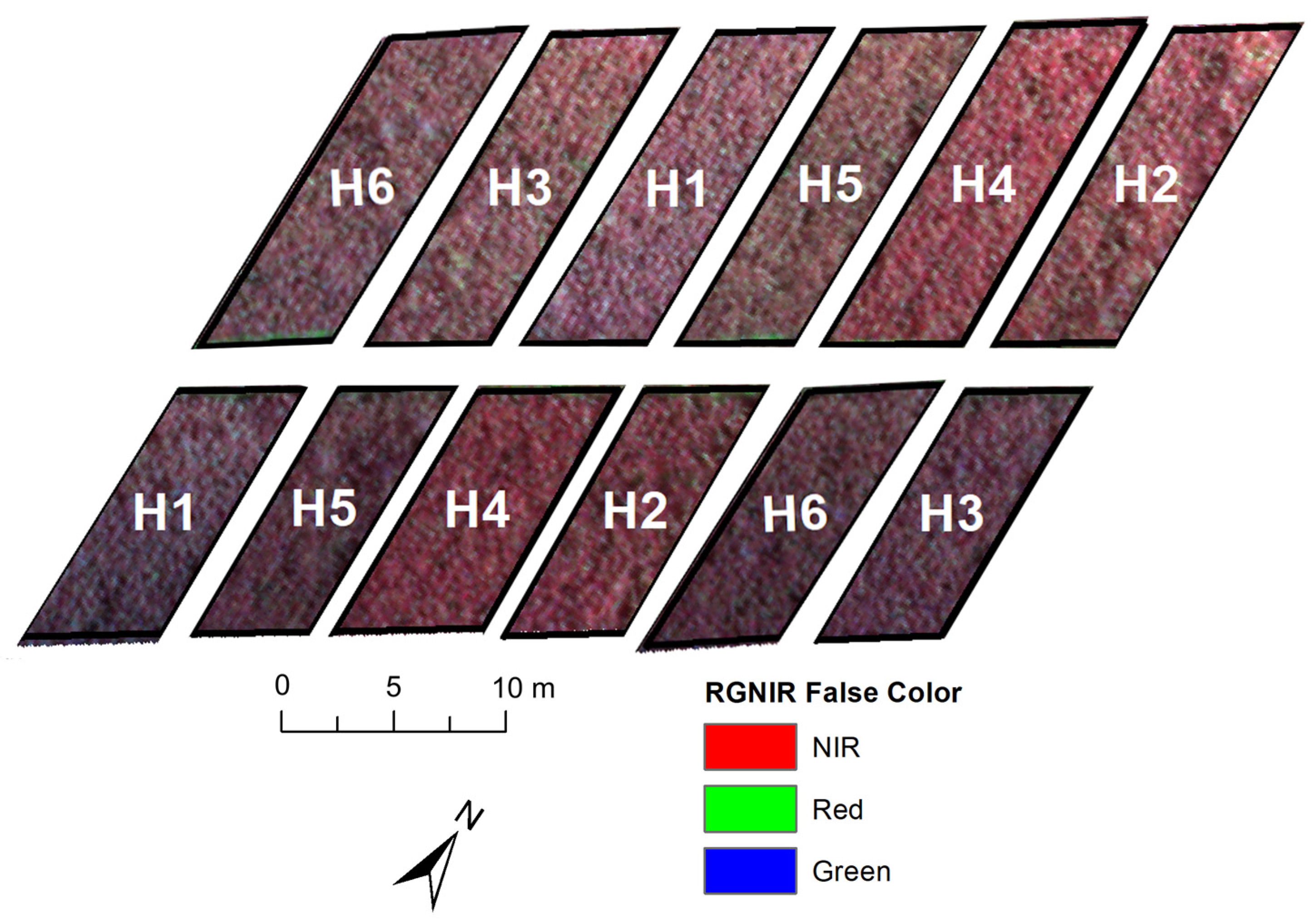
2.2. Collecting and Processing Multispectral Images
2.3. Machine Learning Models and Statistical Analysis
3. Results
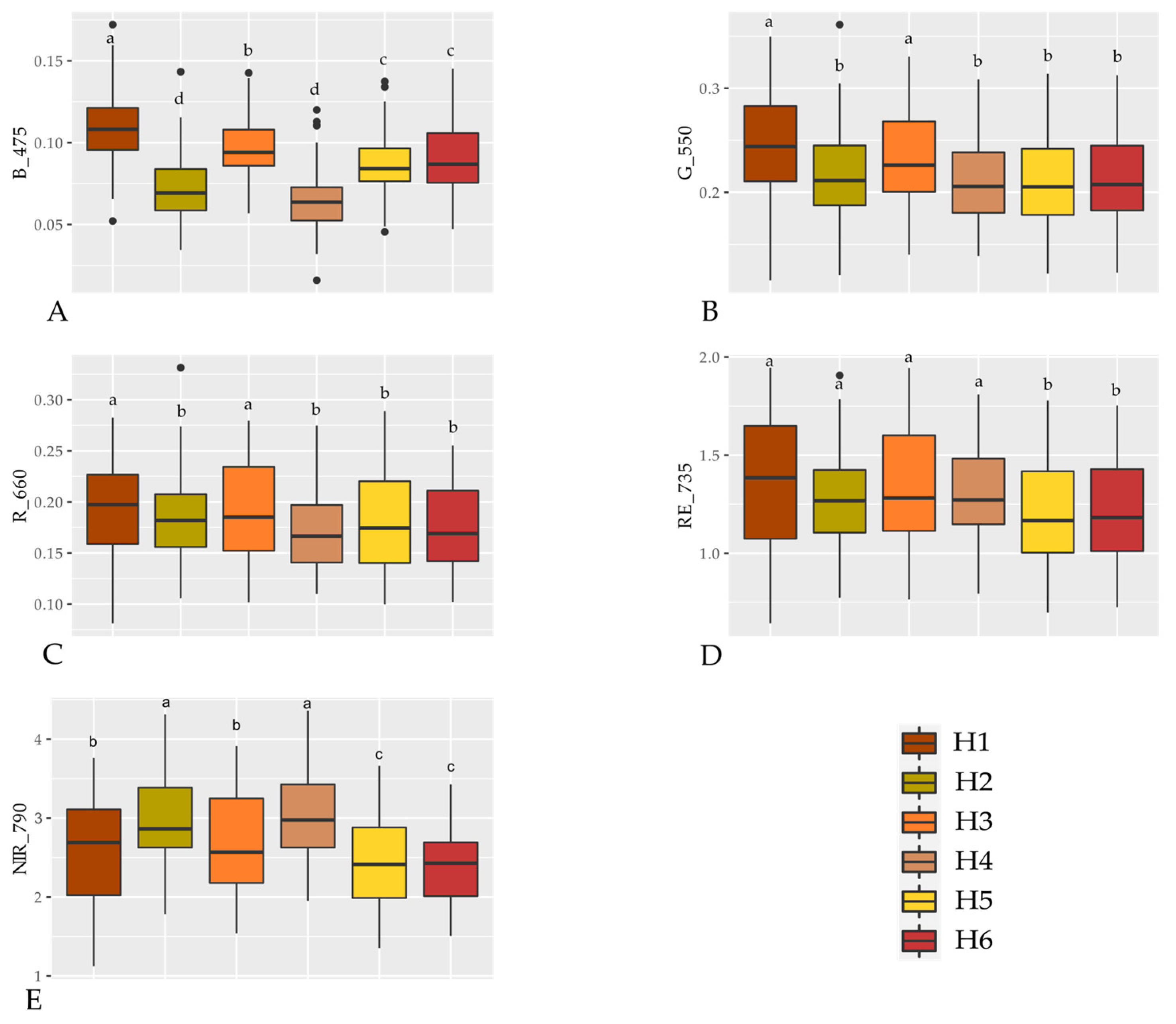
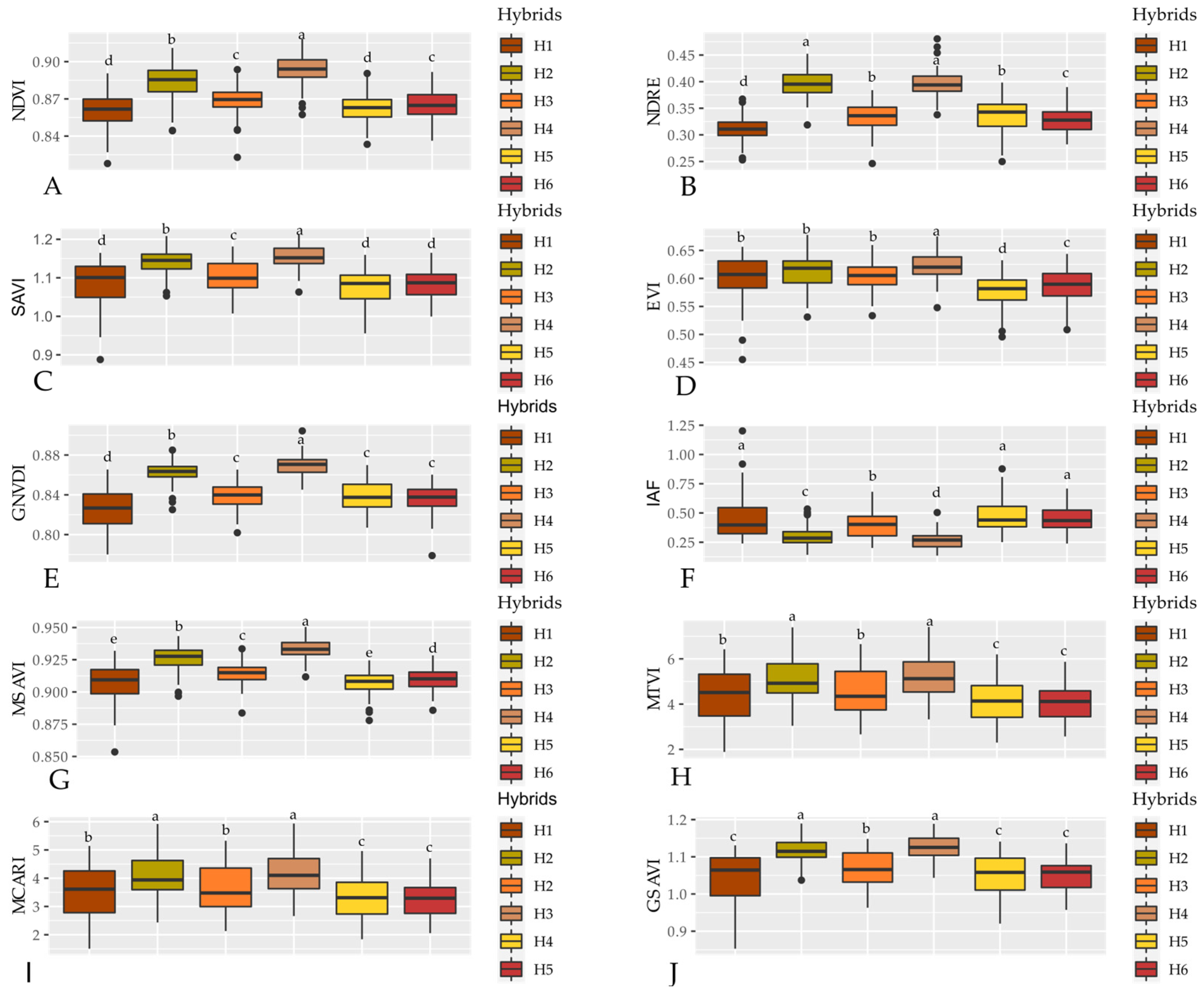
3.1. Spectral Signature of Hybrids
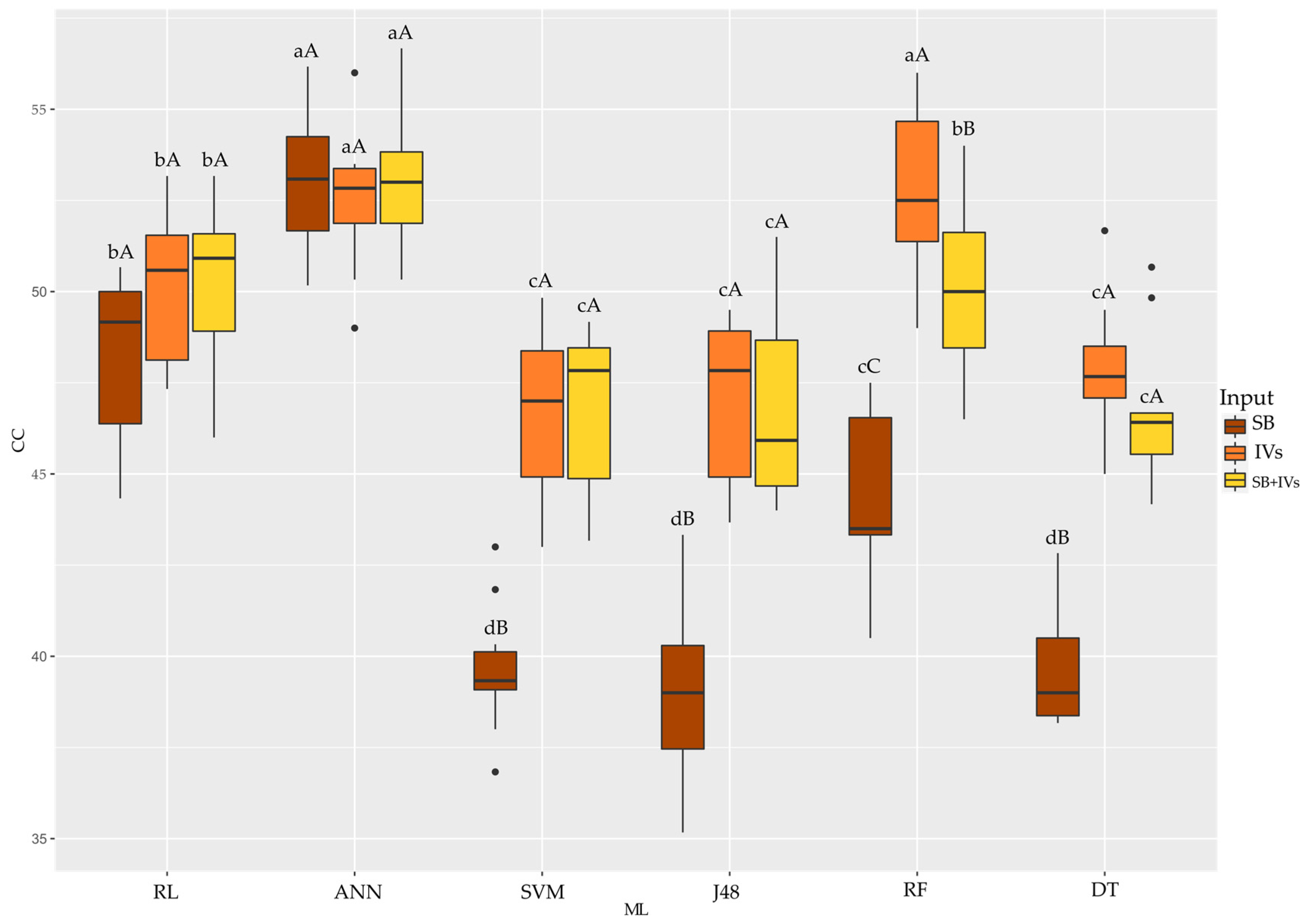
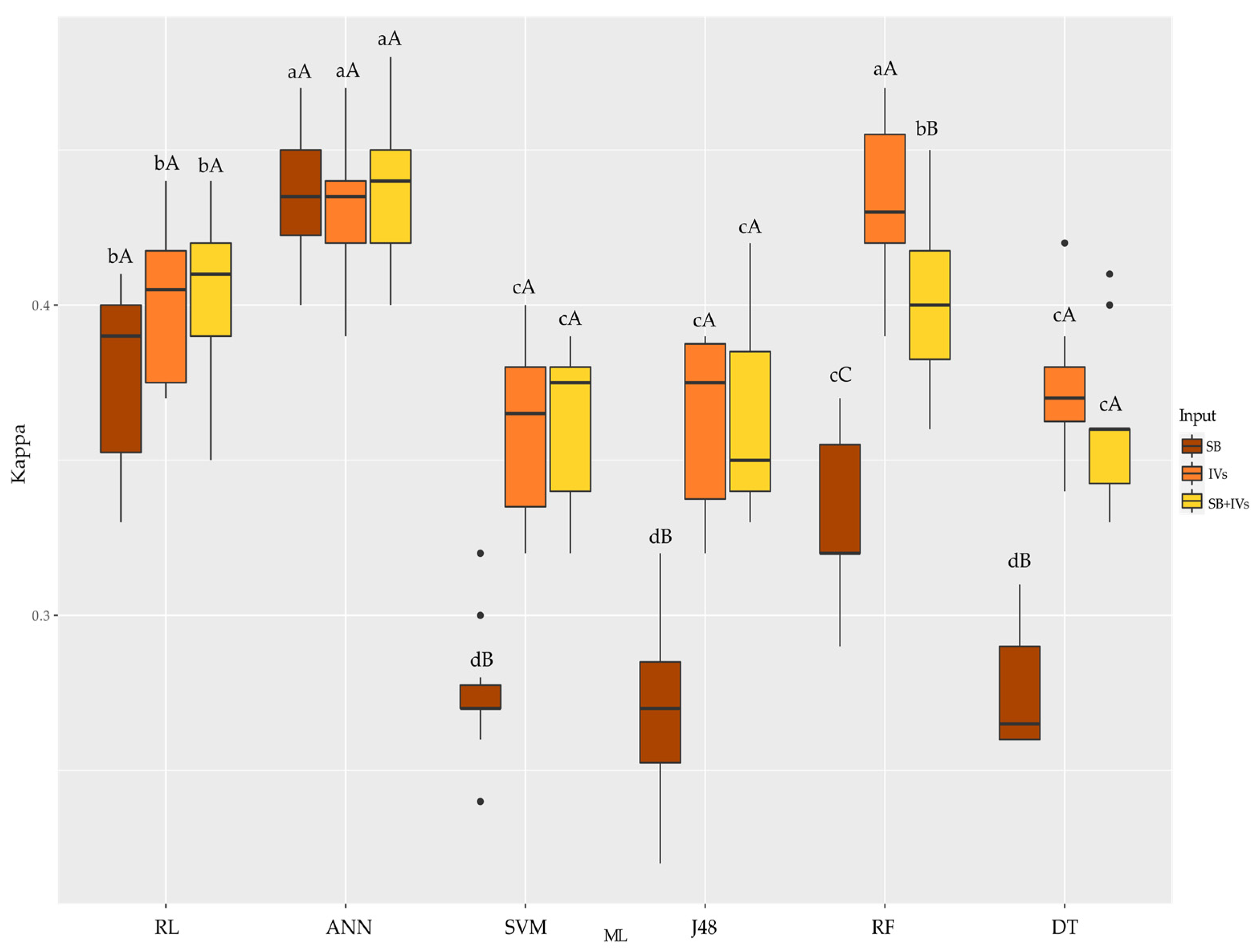
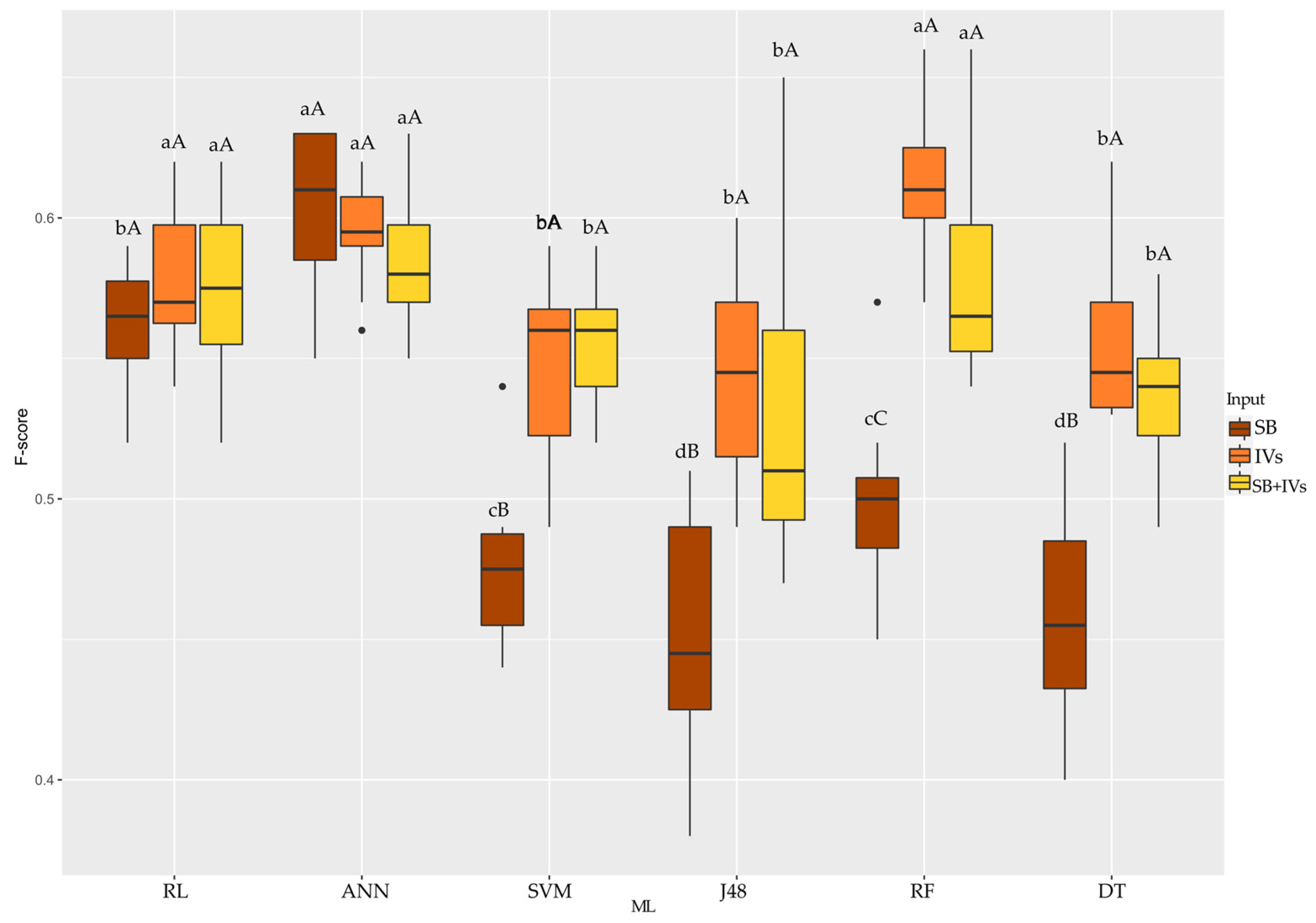
3.2. Classification of Hybrids Using Machine Learning
4. Discussion
4.1. Spectral Signature of Hybrids
4.2. Classification of Hybrids Using Machine Learning
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herzig, P.; Borrmann, P.; Knauer, U.; Klück, H.-C.; Kilias, D.; Seiffert, U.; Pillen, K.; Maurer, A. Evaluation of RGB and multispectral unmanned aerial vehicle (UAV) imagery for high-throughput phenotyping and yield prediction in barley breeding. Remote Sens. 2021, 13, 2670. [Google Scholar] [CrossRef]
- Furbank, R.T.; Tester, M. Phenomics–technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Neumann, K.; Friedel, S.; Kilian, B.; Chen, M.; Altmann, T.; Klukas, C. Dissecting the phenotypic components of crop plant growth and drought responses based on high-throughput image analysis. Plant Cell 2014, 26, 4636–4655. [Google Scholar] [CrossRef] [PubMed]
- Wilke, N.; Siegmann, B.; Postma, J.A.; Muller, O.; Krieger, V.; Pude, R.; Rascher, U. Assessment of plant density for barley and wheat using UAV multispectral imagery for high-throughput field phenotyping. Comput. Electron. Agric. 2021, 189, 106380. [Google Scholar] [CrossRef]
- Candiago, S.; Remondino, F.; De Giglio, M.; Dubbini, M.; Gattelli, M. Evaluating multispectral images and vegetation indices for precision farming applications from UAV images. Remote Sens. 2015, 7, 4026–4047. [Google Scholar] [CrossRef]
- Colomina, I.; Molina, P. Unmanned aerial systems for photogrammetry and remote sensing: A review. ISPRS J. Photogramm. Remote Sens. 2014, 92, 79–97. [Google Scholar] [CrossRef]
- Primicerio, J.; Di Gennaro, S.F.; Fiorillo, E.; Genesio, L.; Lugato, E.; Matese, A.; Vaccari, F.P. A flexible unmanned aerial vehicle for precision agriculture. Precis Agric. 2012, 13, 517–523. [Google Scholar] [CrossRef]
- Kar, S.; Purbey, V.K.; Suradhaniwar, S.; Korbu, L.B.; Kholová, J.; Durbha, S.S.; Adinarayana, J.; Vadez, V. An ensemble machine learning approach for determination of the optimum sampling time for evapotranspiration assessment from high-throughput phenotyping data. Comput. Electron. Agric. 2021, 182, 105992. [Google Scholar] [CrossRef]
- Ren, J.; Shao, Y.; Wan, H.; Xie, Y.; Campos, A. A two-step mapping of irrigated corn with multi-temporal MODIS and Landsat analysis ready data. ISPRS J. Photogramm. Remote Sens. 2021, 176, 69–82. [Google Scholar] [CrossRef]
- Watanabe, K.; Guo, W.; Arai, K.; Takanashi, H.; Kajiya-Kanegae, H.; Kobayashi, M.; Yano, K.; Tokunaga, T.; Fujiwara, T.; Tsutsumi, N.; et al. High-throughput phenotyping of sorghum plant height using an unmanned aerial vehicle and its application to genomic prediction modeling. Front. Plant Sci. 2017, 8, 421. [Google Scholar] [CrossRef]
- Zhang, H.; Ge, Y.; Xie, X.; Atefi, A.; Wijewardane, N.; Thapa, S. High throughput analysis of leaf chlorophyll content in sorghum using RGB, hyperspectral, and fluorescence imaging and sensor fusion. Plant Methods 2022, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, J.D.; Hunt, C.H.; George-Jaeggli, B.; Laws, K.; Watson, J.; Potgieter, A.B.; Cruickshank, A.; Jordan, D.R. High-throughput phenotyping of dynamic canopy traits associated with stay-green in grain sorghum. Plant Phenomics 2020, 2020, 4635153. [Google Scholar] [CrossRef] [PubMed]
- Gill, T.; Gill, S.K.; Saini, D.K.; Chopra, Y.; de Koff, J.P.; Sandhu, K.S. A Comprehensive Review of High Throughput Phenotyping and Machine Learning for Plant Stress Phenotyping. Phenomics 2022, 2, 156–183. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.K.V.; Borges, M.V.V.; Batista, T.S.; da Silva, C.A., Jr.; Furuya, D.E.G.; Osco, L.P.; Ribeiro Teodoro, L.P.; Rojo Baio, F.H.; Ramos, A.P.M.; Gonçalves, W.N.; et al. Predicting eucalyptus diameter at breast height and total height with uav-based spectral indices and machine learning. Forests 2021, 12, 582. [Google Scholar] [CrossRef]
- da Silva, C.A., Jr.; Nanni, M.R.; Shakir, M.; Teodoro, P.E.; de Oliveira-Júnior, J.F.; Cezar, E.; de Gois, G.; Lima, M.; Wojciechowski, J.C.; Shiratsuchi, L.S. Soybean varieties discrimination using non-imaging hyperspectral sensor. Infrared Phys. Technol. 2018, 89, 338–350. [Google Scholar] [CrossRef]
- Yoder, B.J.; Pettigrew-Crosby, R.E. Predicting nitrogen and chlorophyll content and concentrations from reflectance spectra (400–2500 nm) at leaf and canopy scales. Remote Sens. Environ. 1995, 53, 199–211. [Google Scholar] [CrossRef]
- Zhang, J.; Han, M.; Wang, L.; Chen, M.; Chen, C.; Shen, S.; Liu, J.; Zhang, C.; Shang, J.; Yan, X. Study of Genetic Variation in Bermuda Grass along Longitudinal and Latitudinal Gradients Using Spectral Reflectance. Remote Sens. 2023, 15, 896. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Gitelson, A.; Merzlyak, M.N. Spectral Reflectance Changes Associated with Autumn Senescence of Aesculus hippocastanum L. and Acer platanoides L. Leaves. Spectral Features and Relation to Chlorophyll Estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. Available online: https://www.sciencedirect.com/science/article/pii/S0034425702000962 (accessed on 1 November 2023). [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Signature Analysis of Leaf Reflectance Spectra: Algorithm Development for Remote Sensing of Chlorophyll. J. Plant Physiol. 1996, 148, 494–500. Available online: https://www.sciencedirect.com/science/article/pii/S0176161796802847 (accessed on 1 November 2023). [CrossRef]
- Allen, R.G.; Tasumi, M.; Trezza, R.; Waters, R.; Bastiaanssen, W. SEBAL Surface Energy Balance Algorithms for Land: Advanced Training and Users Manual, Idaho Implementation, 1st ed.; Department of Water Resources, University of Idaho: Moscow, ID, USA, 2002. [Google Scholar]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; de Colstoun, E.B.; McMurtrey Iii, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Santana, D.C.; Teixeira Filho, M.C.M.; da Silva, M.R.; das Chagas, P.H.M.; de Oliveira, J.L.G.; Baio, F.H.R.; Campos, C.N.S.; Teodoro, L.P.R.; da Silva, C.A., Jr.; Teodoro, P.E.; et al. Machine Learning in the Classification of Soybean Genotypes for Primary Macronutrients’ Content Using UAV–Multispectral Sensor. Remote Sens. 2023, 15, 1457. [Google Scholar] [CrossRef]
- Gava, R.; Santana, D.C.; Cotrim, M.F.; Rossi, F.S.; Teodoro, L.P.R.; da Silva, C.A., Jr.; Teodoro, P.E. Soybean Cultivars Identification Using Remotely Sensed Image and Machine Learning Models. Sustainability 2022, 14, 7125. [Google Scholar] [CrossRef]
- Baio, F.H.R.; Santana, D.C.; Teodoro, L.P.R.; de Oliveira, I.C.; Gava, R.; de Oliveira, J.L.G.; da Silva, C.A., Jr.; Teodoro, P.E.; Shiratsuchi, L.S. Maize Yield Prediction with Machine Learning, Spectral Variables and Irrigation Management. Remote Sens. 2023, 15, 79. [Google Scholar] [CrossRef]
- Scott, A.J.; Knott, M. A cluster analysis method for grouping means in the analysis of variance. Biometrics 1974, 30, 507–512. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Kycko, M.; Zagajewski, B.; Lavender, S.; Romanowska, E.; Zwijacz-Kozica, M. The impact of tourist traffic on the condition and cell structures of alpine swards. Remote Sens. 2018, 10, 220. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Hobbie, S.E.; Madritch, M.D.; Wang, R.; Tilman, D.; Gamon, J.A. Plant spectral diversity integrates functional and phylogenetic components of biodiversity and predicts ecosystem function. Nat. Ecol. Evol. 2018, 2, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Yoosefzadeh-Najafabadi, M.; Earl, H.J.; Tulpan, D.; Sulik, J.; Eskandari, M. Application of Machine Learning Algorithms in Plant Breeding: Predicting Yield From Hyperspectral Reflectance in Soybean. Front. Plant Sci. 2021, 11, 624273. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.A. Fundamentos Do Sensoriamento Remoto e Metodologias de Aplicação; UFV: Viçosa, Brazil, 2005. [Google Scholar]
- da Silva, C.A., Jr.; Teodoro, P.E.; Teodoro, L.P.R.; Della-Silva, J.L.; Shiratsuchi, L.S.; Baio, F.H.R.; Boechat, C.L.; Capristo-Silva, G.F. Is it possible to detect boron deficiency in eucalyptus using hyper and multispectral sensors? Infrared Phys. Technol. 2021, 116, 103810. [Google Scholar] [CrossRef]
- de Moraes, E.C. Fundamentos de Sensoriamento Remoto; INPE: São José dos Campos, Brazil, 2002; pp. 1–7.
- Bergamaschi, H.; Bergonci, J.I. As Plantas e o Clima: Princípios e Aplicações; Agrolivros: Guaíba, Brazil, 2017. [Google Scholar]
- Liu, L.; Song, B.; Zhang, S.; Liu, X. A novel principal component analysis method for the reconstruction of leaf reflectance spectra and retrieval of leaf biochemical contents. Remote Sens. 2017, 9, 1113. [Google Scholar] [CrossRef]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S.M. Enhancement of the stomatal response to blue light by red light, reduced intercellular concentrations of CO2, and low vapor pressure differences. Plant Physiol. 1988, 87, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, E.; Oiwamoto, K.; Inoue, S.; Shimazaki, K.; Doi, M. Stomatal response to blue light in crassulacean acid metabolism plants Kalanchoe pinnata and Kalanchoe daigremontiana. J. Exp. Bot. 2019, 70, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant Cell Environ. 2000, 23, 539–548. [Google Scholar] [CrossRef]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Trojak, M.; Skowron, E.; Sobala, T.; Kocurek, M.; Pałyga, J. Effects of partial replacement of red by green light in the growth spectrum on photomorphogenesis and photosynthesis in tomato plants. Photosynth. Res. 2021, 151, 295–312. [Google Scholar] [CrossRef]
- Santos, L.B.; Bastos, L.M.; de Oliveira, M.F.; Soares, P.L.M.; Ciampitti, I.A.; da Silva, R.P. Identifying Nematode Damage on Soybean through Remote Sensing and Machine Learning Techniques. Agronomy 2022, 12, 2404. [Google Scholar] [CrossRef]
- Seelig, H.D.; Hoehn, A.; Stodieck, L.S.; Klaus, D.M.; Adams, W.W., III; Emery, W.J. The assessment of leaf water content using leaf reflectance ratios in the visible, near-, and short-wave-infrared. Int. J. Remote Sens. 2008, 29, 3701–3713. [Google Scholar] [CrossRef]
- Kior, A.; Sukhov, V.; Sukhova, E. Application of reflectance indices for remote sensing of plants and revealing actions of stressors. Photonics 2021, 8, 582. [Google Scholar] [CrossRef]
- Motomiya, A.V.D.A.; Molin, J.P.; Motomiya, W.R.; Rojo Baio, F.H. Mapeamento do índice de vegetação da diferença normalizada em lavoura de algodão. Pesqui Agropecu Trop. 2012, 42, 112–118. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H.; Sauer, T.J.; Dold, C.; O’Brien, P.; Wacha, K. Applications of vegetative indices from remote sensing to agriculture: Past and future. Inventions 2019, 4, 71. [Google Scholar] [CrossRef]
- Giovos, R.; Tassopoulos, D.; Kalivas, D.; Lougkos, N.; Priovolou, A. Remote sensing vegetation indices in viticulture: A critical review. Agriculture 2021, 11, 457. [Google Scholar] [CrossRef]
- Santana, D.C.; Teodoro, L.P.R.; Baio, F.H.R.; dos Santos, R.G.; Coradi, P.C.; Biduski, B.; da Silva, C.A., Jr.; Teodoro, P.E.; Shiratsuchi, L.S. Classification of soybean genotypes for industrial traits using UAV multispectral imagery and machine learning. Remote Sens. Appl. 2023, 29, 100919. [Google Scholar] [CrossRef]
- Borges, M.V.V.; Garcia, J.d.O.; Batista, T.S.; Silva, A.N.M.; Baio, F.H.R.; da Silva, C.A., Jr.; de Azevedo, G.B.; Azevedo, G.T.d.O.S.; Teodoro, L.P.R.; Teodoro, P.E. High-throughput phenotyping of two plant-size traits of Eucalyptus species using neural networks. J. For. Res. 2022, 33, 591–599. [Google Scholar] [CrossRef]
- Sedghi, M.; Golian, A.; Soleimani-Roodi, P.; Ahmadi, A.; Aami-Azghadi, M. Relationship between color and tannin content in sorghum grain: Application of image analysis and artificial neural network. Braz. J. Poult. Sci. 2012, 14, 57–62. [Google Scholar] [CrossRef]
- de Oliveira, B.R.; da Silva, A.A.P.; Teodoro, L.P.R.; de Azevedo, G.B.; Azevedo, G.T.D.O.S.; Baio, F.H.R.; Sobrinho, R.L.; da Silva, C.A., Jr.; Teodoro, P.E. Eucalyptus growth recognition using machine learning methods and spectral variables. For. Ecol. Manag. 2021, 497, 119496. [Google Scholar] [CrossRef]
- Mokarram, M.; Hojjati, M.; Roshan, G.; Negahban, S. Modeling the behavior of Vegetation Indices in the salt dome of Korsia in North-East of Darab, Fars, Iran. Model Earth Syst. Environ. 2015, 1, 27. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sigla | Vegetation Index | Equation | Reference |
|---|---|---|---|
| NDVI | Normalized Difference Vegetation Indice | [18] | |
| NDRE | Normalized Difference Red Edge Indice | [19] | |
| SAVI | Soil-Adjusted Vegetation Index | [20] | |
| EVI | Enhanced Vegetation Index (EVI) | [21] | |
| GNDVI | Green Normalized Difference Vegetation | [22] | |
| LAI | Leaf Area Index | [23] | |
| MSAVI | Modified Soil Adjusted Vegetation Index | [24] | |
| MTVI | Modified Triangular Vegetation Index | [25] | |
| MCARI | Modified Chlorophyll Absorption Ratio Index | [26] |
| Sigla | Accuracy | Equation |
|---|---|---|
| CC | Correct classifications | |
| Kappa | Kappa coefficient | |
| F-score | F-score |
| S.V. | D.F. | CC | Kappa | F-Score |
|---|---|---|---|---|
| ML | 5 | 377.88 * | 0.05 * | 0.03 * |
| Input | 2 | 556.7 * | 0.08 * | 0.07 * |
| ML × Input | 10 | 45.06 * | 0.01 * | 0.01 * |
| Residual | 162 | 4.56 | 6.66 | 0 |
| C.V. (%) | 4.5 | 6.99 | 5.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, D.C.; Theodoro, G.d.F.; Gava, R.; de Oliveira, J.L.G.; Teodoro, L.P.R.; de Oliveira, I.C.; Baio, F.H.R.; da Silva Junior, C.A.; de Oliveira, J.T.; Teodoro, P.E. A New Approach to Identifying Sorghum Hybrids Using UAV Imagery Using Multispectral Signature and Machine Learning. Algorithms 2024, 17, 23. https://doi.org/10.3390/a17010023
Santana DC, Theodoro GdF, Gava R, de Oliveira JLG, Teodoro LPR, de Oliveira IC, Baio FHR, da Silva Junior CA, de Oliveira JT, Teodoro PE. A New Approach to Identifying Sorghum Hybrids Using UAV Imagery Using Multispectral Signature and Machine Learning. Algorithms. 2024; 17(1):23. https://doi.org/10.3390/a17010023
Chicago/Turabian StyleSantana, Dthenifer Cordeiro, Gustavo de Faria Theodoro, Ricardo Gava, João Lucas Gouveia de Oliveira, Larissa Pereira Ribeiro Teodoro, Izabela Cristina de Oliveira, Fábio Henrique Rojo Baio, Carlos Antonio da Silva Junior, Job Teixeira de Oliveira, and Paulo Eduardo Teodoro. 2024. "A New Approach to Identifying Sorghum Hybrids Using UAV Imagery Using Multispectral Signature and Machine Learning" Algorithms 17, no. 1: 23. https://doi.org/10.3390/a17010023
APA StyleSantana, D. C., Theodoro, G. d. F., Gava, R., de Oliveira, J. L. G., Teodoro, L. P. R., de Oliveira, I. C., Baio, F. H. R., da Silva Junior, C. A., de Oliveira, J. T., & Teodoro, P. E. (2024). A New Approach to Identifying Sorghum Hybrids Using UAV Imagery Using Multispectral Signature and Machine Learning. Algorithms, 17(1), 23. https://doi.org/10.3390/a17010023