
Investigating Shared Genetic Bases between Psychiatric Disorders, Cardiometabolic and Sleep Traits Using K-Means Clustering and Local Genetic Correlation Analysis

Abstract
1. Introduction
2. Materials and Methods
2.1. GWAS Datasets
2.2. Linkage Disequilibrium Score Regression (LDSC) and K-Means Clustering
2.3. Mendelian Randomization
2.4. Functional Effect of SNPs Identified with MR
2.5. Local Genetic Correlation Analysis
3. Results
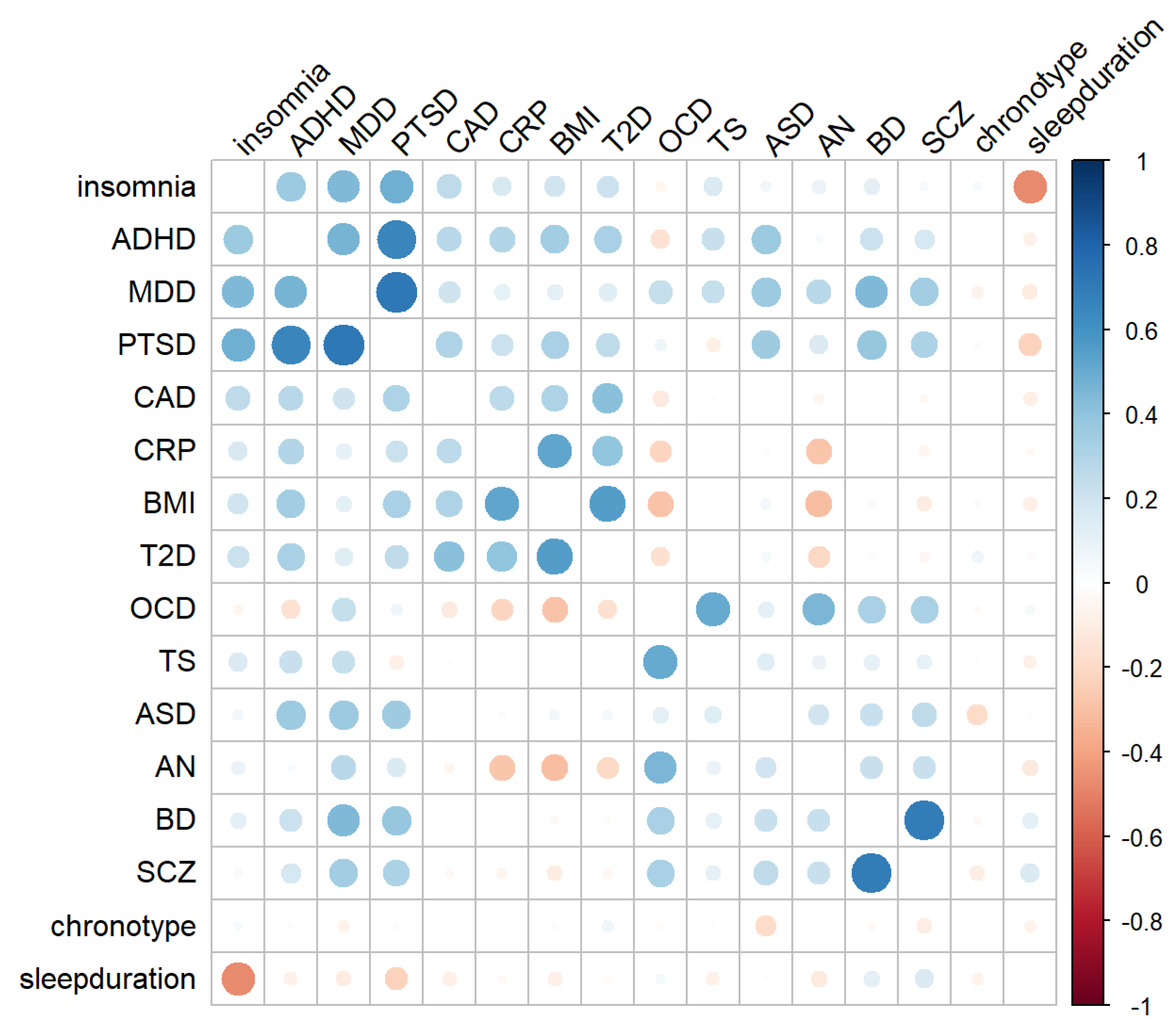
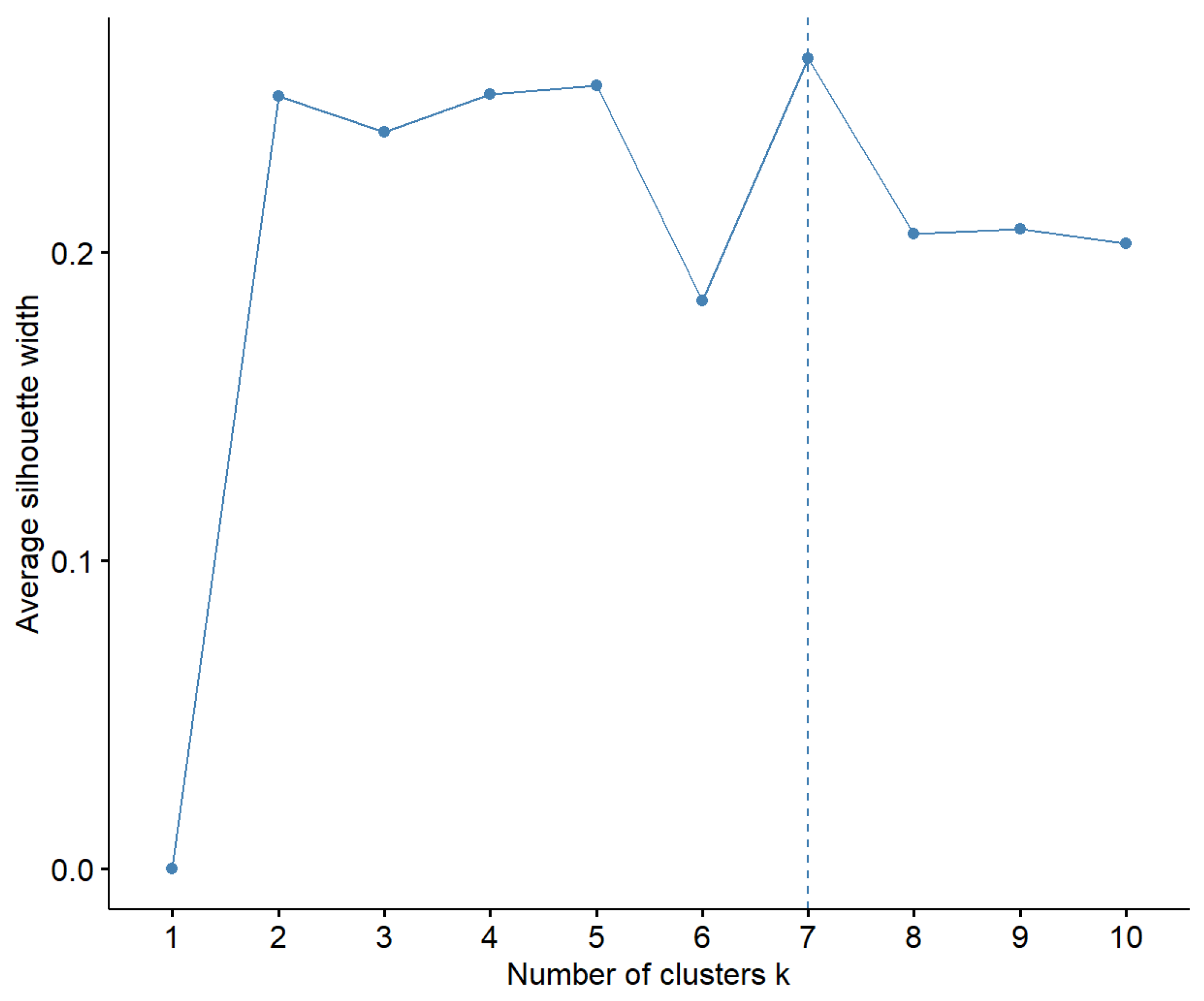
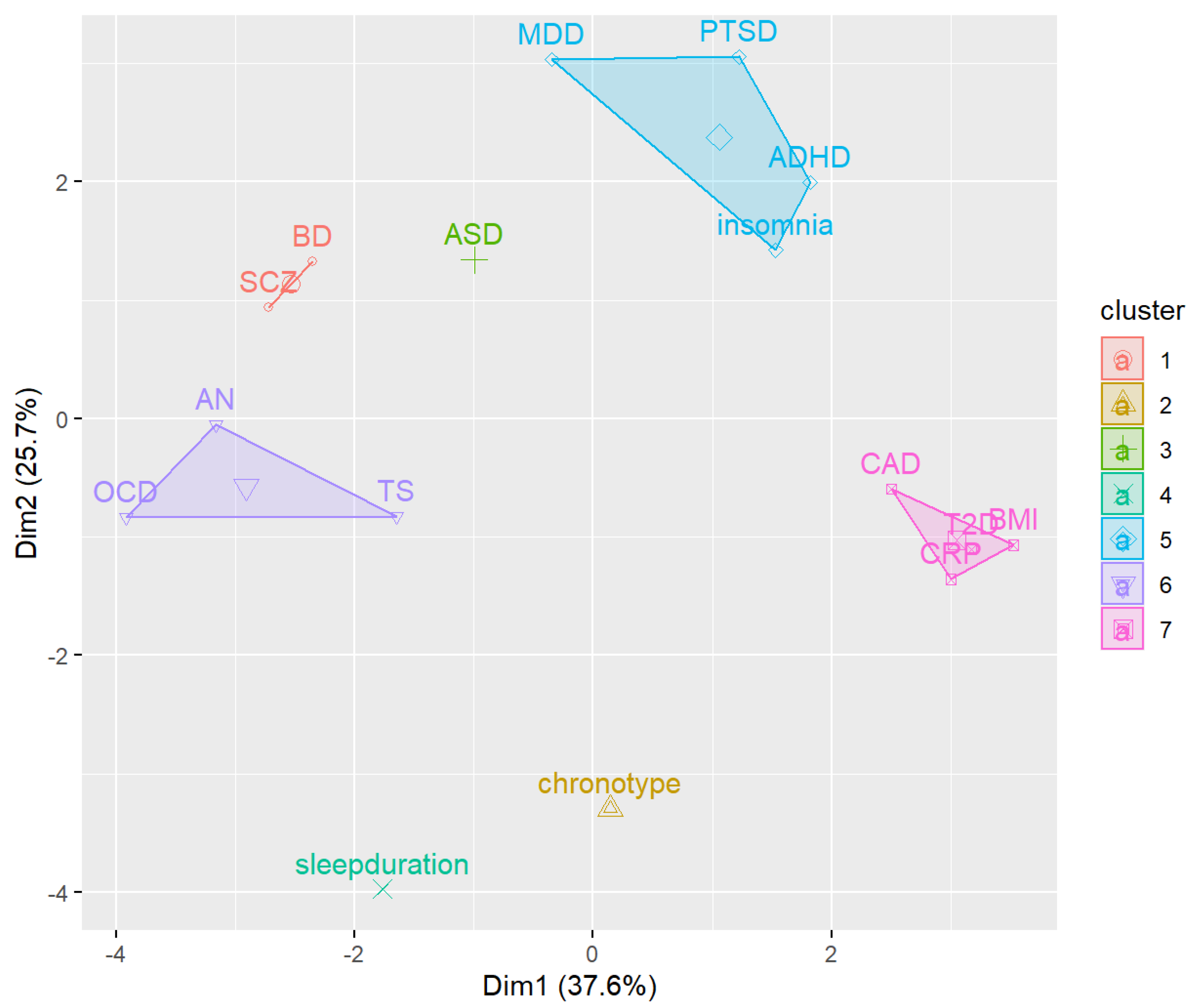
3.1. Linkage Disequilibrium Score Regression (LDSC) and K-Means Clustering
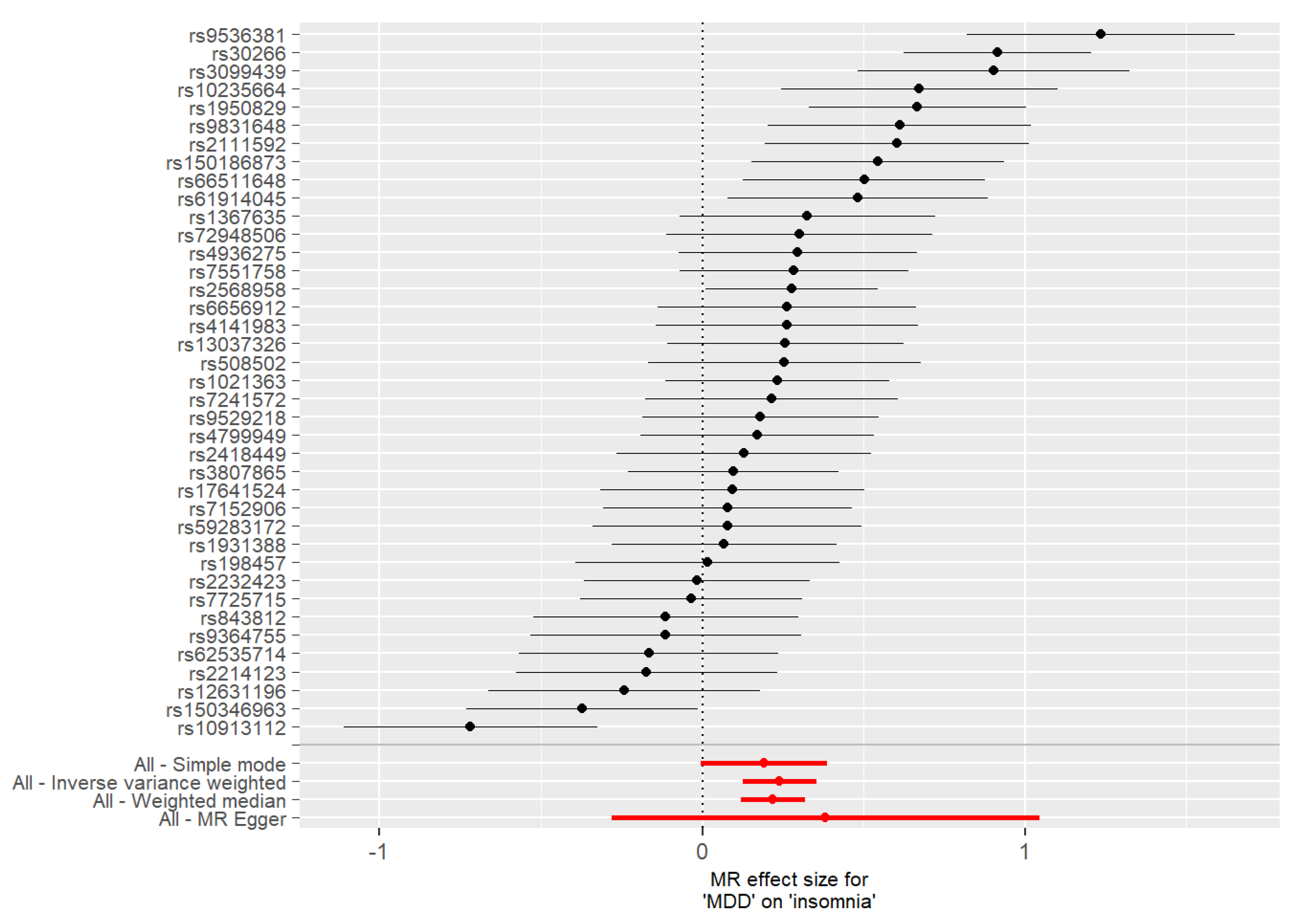
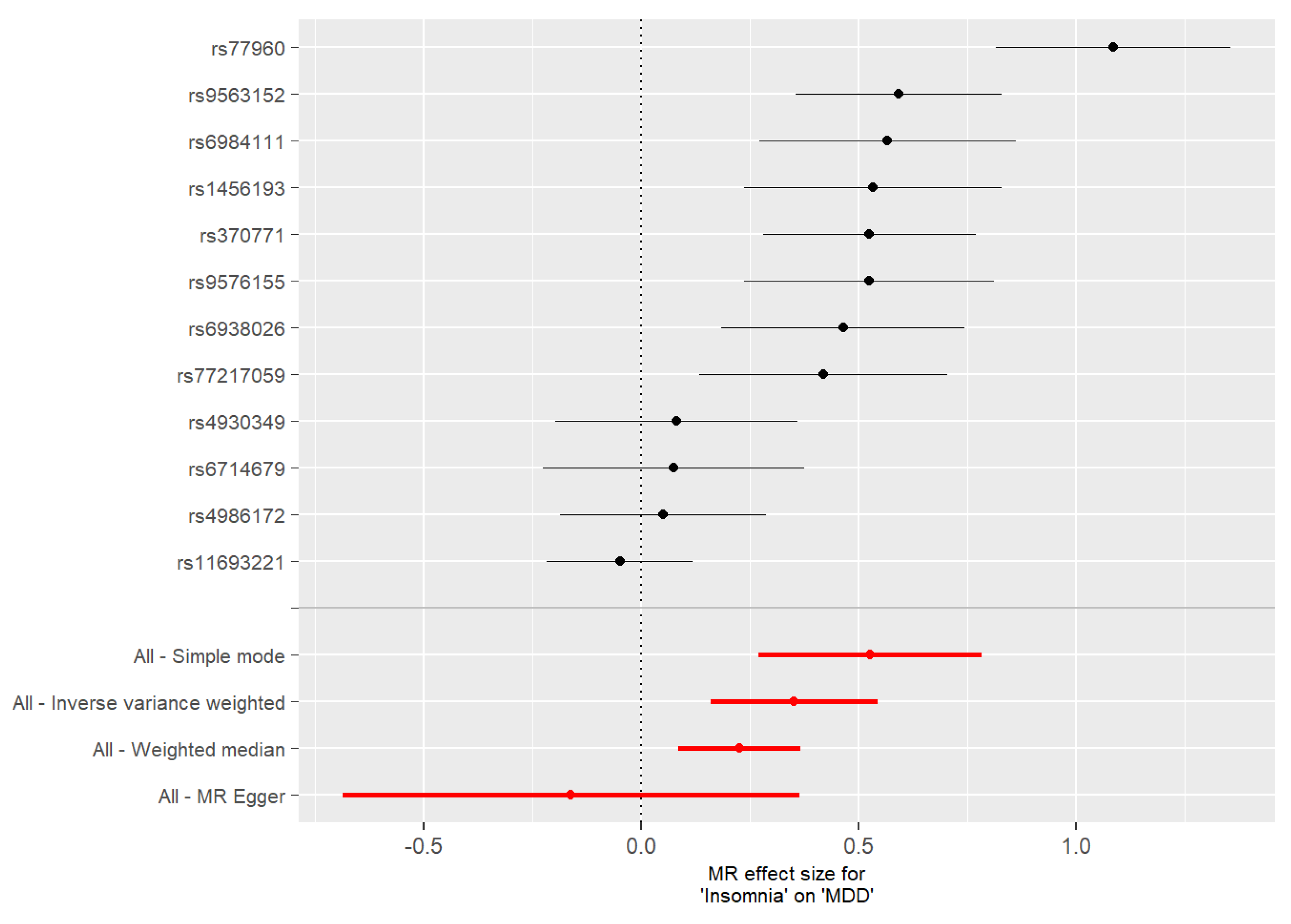
3.2. Mendelian Randomization
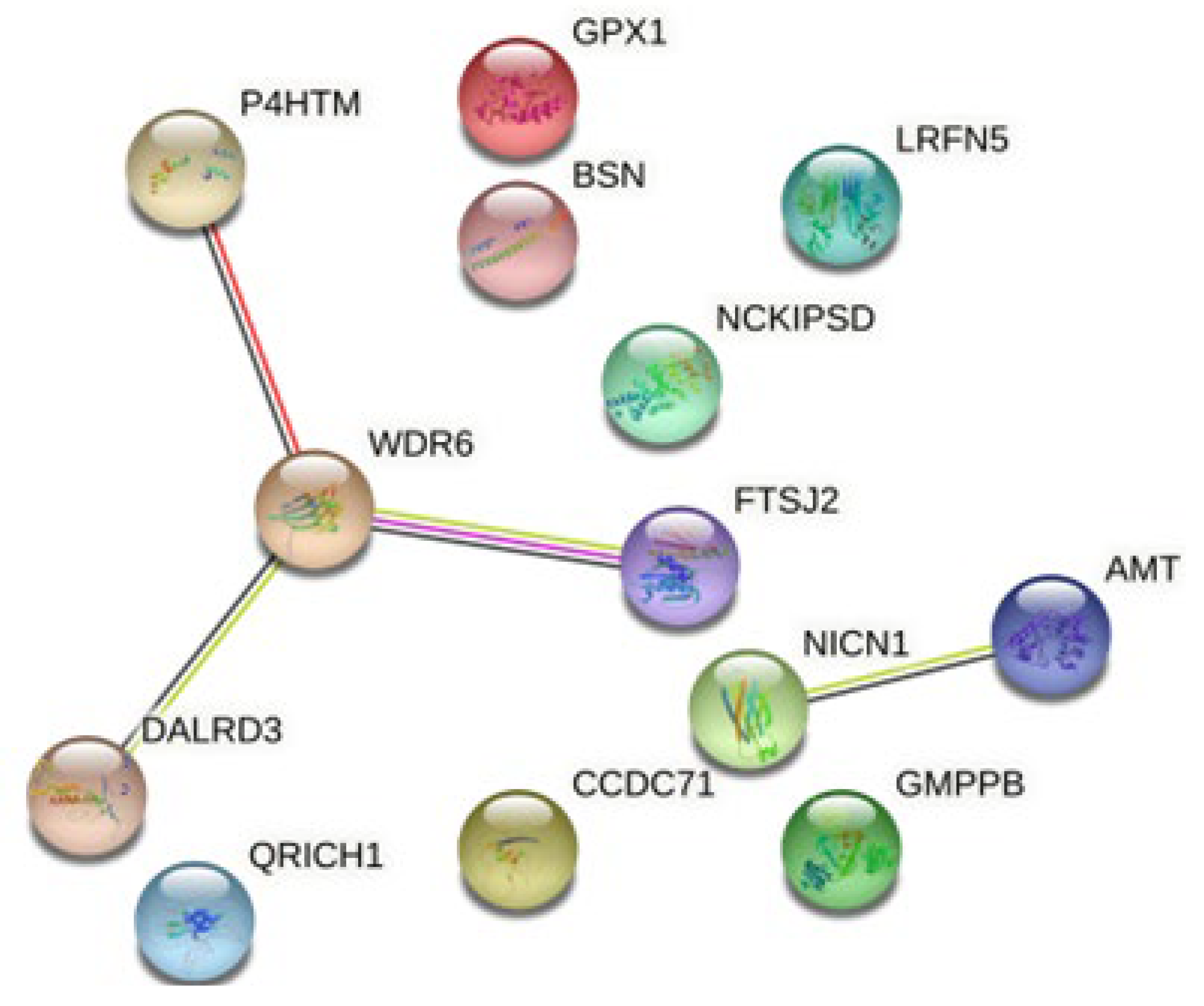
3.3. Functional Effect of SNPs Identified with MR in the Analysis with MDD and Insomnia
3.4. Local Genetic Correlation Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Who Mental Health. Available online: https://www.who.int/mental_health/management/en (accessed on 8 September 2022).
- WHO Mental Disorders. Available online: https://www.who.int/news-room/fact-sheets/detail/mental-disorders (accessed on 8 September 2022).
- GBD Compare Viz Hub. Available online: https://vizhub.healthdata.org/gbd-compare/# (accessed on 8 September 2022).
- Collaborators, C.-M.D. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef]
- Mullins, N.; Forstner, A.J.; O’Connell, K.S.; Coombes, B.; Coleman, J.R.I.; Qiao, Z.; Als, T.D.; Bigdeli, T.B.; Borte, S.; Bryois, J.; et al. Genome-wide association study of more than 40,000 bipolar disorder cases provides new insights into the underlying biology. Nat. Genet. 2021, 53, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Trubetskoy, V.; Pardinas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Howard, D.M.; Adams, M.J.; Clarke, T.K.; Hafferty, J.D.; Gibson, J.; Shirali, M.; Coleman, J.R.I.; Hagenaars, S.P.; Ward, J.; Wigmore, E.M.; et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat. Neurosci. 2019, 22, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.H.; Daumit, G.L.; Dua, T.; Aquila, R.; Charlson, F.; Cuijpers, P.; Druss, B.; Dudek, K.; Freeman, M.; Fujii, C.; et al. Excess mortality in persons with severe mental disorders: A multilevel intervention framework and priorities for clinical practice, policy and research agendas. World Psychiatry 2017, 16, 30–40. [Google Scholar] [CrossRef]
- Nordentoft, M.; Wahlbeck, K.; Hallgren, J.; Westman, J.; Osby, U.; Alinaghizadeh, H.; Gissler, M.; Laursen, T.M. Excess mortality, causes of death and life expectancy in 270,770 patients with recent onset of mental disorders in Denmark, Finland and Sweden. PLoS ONE 2013, 8, e55176. [Google Scholar] [CrossRef]
- Lawrence, D.; Kisely, S.; Pais, J. The epidemiology of excess mortality in people with mental illness. Can. J. Psychiatry 2010, 55, 752–760. [Google Scholar] [CrossRef]
- Newcomer, J.W.; Hennekens, C.H. Severe mental illness and risk of cardiovascular disease. JAMA 2007, 298, 1794–1796. [Google Scholar] [CrossRef]
- De Hert, M.; Dekker, J.M.; Wood, D.; Kahl, K.G.; Holt, R.I.; Moller, H.J. Cardiovascular disease and diabetes in people with severe mental illness position statement from the European Psychiatric Association (EPA), supported by the European Association for the Study of Diabetes (EASD) and the European Society of Cardiology (ESC). Eur. Psychiatry 2009, 24, 412–424. [Google Scholar] [CrossRef]
- Osby, U.; Brandt, L.; Correia, N.; Ekbom, A.; Sparen, P. Excess mortality in bipolar and unipolar disorder in Sweden. Arch. Gen. Psychiatry 2001, 58, 844–850. [Google Scholar] [CrossRef]
- Calkin, C.V.; Gardner, D.M.; Ransom, T.; Alda, M. The relationship between bipolar disorder and type 2 diabetes: More than just co-morbid disorders. Ann. Med. 2013, 45, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Pisanu, C.; Williams, M.J.; Ciuculete, D.M.; Olivo, G.; Del Zompo, M.; Squassina, A.; Schioth, H.B. Evidence that genes involved in hedgehog signaling are associated with both bipolar disorder and high BMI. Transl. Psychiatry 2019, 9, 315. [Google Scholar] [CrossRef] [PubMed]
- Maina, G.; Salvi, V.; Vitalucci, A.; D’Ambrosio, V.; Bogetto, F. Prevalence and correlates of overweight in drug-naive patients with bipolar disorder. J. Affect. Disord. 2008, 110, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.M.; Barry, D.; Pietrzak, R.H.; Wagner, J.A. Overweight and obesity are associated with psychiatric disorders: Results from the National Epidemiologic Survey on Alcohol and Related Conditions. Psychosom. Med. 2008, 70, 288–297. [Google Scholar] [CrossRef]
- Zou, H.; Zhou, H.; Yan, R.; Yao, Z.; Lu, Q. Chronotype, circadian rhythm, and psychiatric disorders: Recent evidence and potential mechanisms. Front. Neurosci. 2022, 16, 811771. [Google Scholar] [CrossRef]
- Harvey, A.G. A transdiagnostic approach to treating sleep disturbance in psychiatric disorders. Cogn. Behav. Ther. 2009, 38 (Suppl. 1), 35–42. [Google Scholar] [CrossRef]
- Taylor, B.J.; Hasler, B.P. Chronotype and Mental Health: Recent Advances. Curr. Psychiatry Rep. 2018, 20, 59. [Google Scholar] [CrossRef]
- O’Connell, K.S.; Frei, O.; Bahrami, S.; Smeland, O.B.; Bettella, F.; Cheng, W.; Chu, Y.; Hindley, G.; Lin, A.; Shadrin, A.; et al. Characterizing the Genetic Overlap Between Psychiatric Disorders and Sleep-Related Phenotypes. Biol. Psychiatry 2021, 90, 621–631. [Google Scholar] [CrossRef]
- Sun, X.; Liu, B.; Liu, S.; Wu, D.J.H.; Wang, J.; Qian, Y.; Ye, D.; Mao, Y. Sleep disturbance and psychiatric disorders: A bidirectional Mendelian randomisation study. Epidemiol. Psychiatr. Sci. 2022, 31, e26. [Google Scholar] [CrossRef]
- Demontis, D.; Walters, R.K.; Martin, J.; Mattheisen, M.; Als, T.D.; Agerbo, E.; Baldursson, G.; Belliveau, R.; Bybjerg-Grauholm, J.; Baekvad-Hansen, M.; et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat. Genet. 2019, 51, 63–75. [Google Scholar] [CrossRef]
- Grove, J.; Ripke, S.; Als, T.D.; Mattheisen, M.; Walters, R.K.; Won, H.; Pallesen, J.; Agerbo, E.; Andreassen, O.A.; Anney, R.; et al. Identification of common genetic risk variants for autism spectrum disorder. Nat. Genet. 2019, 51, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Nievergelt, C.M.; Maihofer, A.X.; Klengel, T.; Atkinson, E.G.; Chen, C.Y.; Choi, K.W.; Coleman, J.R.I.; Dalvie, S.; Duncan, L.E.; Gelernter, J.; et al. International meta-analysis of PTSD genome-wide association studies identifies sex- and ancestry-specific genetic risk loci. Nat. Commun. 2019, 10, 4558. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.D.; Askland, K.D.; Barlassina, C.; Bellodi, L.; Bienvenu, O.J.; Black, D.; Bloch, M.; Brentani, H.; Burton, C.L.; Camarena, B.; et al. Revealing the complex genetic architecture of obsessive-compulsive disorder using meta-analysis. Mol. Psychiatry 2018, 23, 1181–1188. [Google Scholar] [CrossRef]
- Yu, D.; Sul, J.H.; Tsetsos, F.; Nawaz, M.S.; Huang, A.Y.; Zelaya, I.; Illmann, C.; Osiecki, L.; Darrow, S.M.; Hirschtritt, M.E.; et al. Interrogating the Genetic Determinants of Tourette’s Syndrome and Other Tic Disorders Through Genome-Wide Association Studies. Am. J. Psychiatry 2019, 176, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.J.; Yilmaz, Z.; Thornton, L.M.; Hubel, C.; Coleman, J.R.I.; Gaspar, H.A.; Bryois, J.; Hinney, A.; Leppa, V.M.; Mattheisen, M.; et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 2019, 51, 1207–1214. [Google Scholar] [CrossRef]
- Pulit, S.L.; Stoneman, C.; Morris, A.P.; Wood, A.R.; Glastonbury, C.A.; Tyrrell, J.; Yengo, L.; Ferreira, T.; Marouli, E.; Ji, Y.; et al. Meta-analysis of genome-wide association studies for body fat distribution in 694 649 individuals of European ancestry. Hum. Mol. Genet. 2019, 28, 166–174. [Google Scholar] [CrossRef]
- Mahajan, A.; Spracklen, C.N.; Zhang, W.; Ng, M.C.Y.; Petty, L.E.; Kitajima, H.; Yu, G.Z.; Rueger, S.; Speidel, L.; Kim, Y.J.; et al. Multi-ancestry genetic study of type 2 diabetes highlights the power of diverse populations for discovery and translation. Nat. Genet. 2022, 54, 560–572. [Google Scholar] [CrossRef] [PubMed]
- van der Harst, P.; Verweij, N. Identification of 64 Novel Genetic Loci Provides an Expanded View on the Genetic Architecture of Coronary Artery Disease. Circ. Res. 2018, 122, 433–443. [Google Scholar] [CrossRef]
- Said, S.; Pazoki, R.; Karhunen, V.; Vosa, U.; Ligthart, S.; Bodinier, B.; Koskeridis, F.; Welsh, P.; Alizadeh, B.Z.; Chasman, D.I.; et al. Genetic analysis of over half a million people characterises C-reactive protein loci. Nat. Commun. 2022, 13, 2198. [Google Scholar] [CrossRef]
- Watanabe, K.; Jansen, P.R.; Savage, J.E.; Nandakumar, P.; Wang, X.; 23andMe Research Team; Hinds, D.A.; Gelernter, J.; Levey, D.F.; Polimanti, R.; et al. Genome-wide meta-analysis of insomnia prioritizes genes associated with metabolic and psychiatric pathways. Nat. Genet. 2022, 54, 1125–1132. [Google Scholar] [CrossRef]
- Jones, S.E.; Lane, J.M.; Wood, A.R.; van Hees, V.T.; Tyrrell, J.; Beaumont, R.N.; Jeffries, A.R.; Dashti, H.S.; Hillsdon, M.; Ruth, K.S.; et al. Genome-wide association analyses of chronotype in 697,828 individuals provides insights into circadian rhythms. Nat. Commun. 2019, 10, 343. [Google Scholar] [CrossRef]
- Dashti, H.S.; Jones, S.E.; Wood, A.R.; Lane, J.M.; van Hees, V.T.; Wang, H.; Rhodes, J.A.; Song, Y.; Patel, K.; Anderson, S.G.; et al. Genome-wide association study identifies genetic loci for self-reported habitual sleep duration supported by accelerometer-derived estimates. Nat. Commun. 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Li, Y.; Abecasis, G.R. METAL: Fast and efficient meta-analysis of genomewide association scans. Bioinformatics 2010, 26, 2190–2191. [Google Scholar] [CrossRef] [PubMed]
- Bulik-Sullivan, B.; Finucane, H.K.; Anttila, V.; Gusev, A.; Day, F.R.; Loh, P.R.; ReproGen Consortium; Psychiatric Genomics Consortium; Genetic Consortium for Anorexia Nervosa of the Wellcome Trust Case Control Consortium 3; Duncan, L.; et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Bulik-Sullivan, B.K.; Loh, P.R.; Finucane, H.K.; Ripke, S.; Yang, J.; Schizophrenia Working Group of the Psychiatric Genomics Consortium; Patterson, N.; Daly, M.J.; Price, A.L.; Neale, B.M. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat. Genet. 2015, 47, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. cluster: Cluster Analysis Basics and Extensions. R package version 2.1.4. 2019. Available online: https://cran.r-project.org/web/packages/cluster/index.html (accessed on 8 September 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Franti, P.; Sieranoja, S. K-means properties on six clustering benchmark datasets. Appl. Intell. 2018, 48, 4743–4759. [Google Scholar] [CrossRef]
- Fave, M.J.; Lamaze, F.C.; Soave, D.; Hodgkinson, A.; Gauvin, H.; Bruat, V.; Grenier, J.C.; Gbeha, E.; Skead, K.; Smargiassi, A.; et al. Gene-by-environment interactions in urban populations modulate risk phenotypes. Nat. Commun. 2018, 9, 827. [Google Scholar] [CrossRef]
- Hemani, G.; Tilling, K.; Smith, G.D. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017, 13, e1007081. [Google Scholar] [CrossRef]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Sanderson, E.; Richardson, T.G.; Morris, T.T.; Tilling, K.; Smith, G.D. Estimation of causal effects of a time-varying exposure at multiple time points through multivariable mendelian randomization. PLoS Genet. 2022, 18, e1010290. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Thompson, S.G. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur. J. Epidemiol. 2017, 32, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Verbanck, M.; Chen, C.Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Boyle, A.P. Predicting functional variants in enhancer and promoter elements using RegulomeDB. Hum. Mutat. 2019, 40, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.T. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Werme, J.; van der Sluis, S.; Posthuma, D.; de Leeuw, C.A. An integrated framework for local genetic correlation analysis. Nat. Genet. 2022, 54, 274–282. [Google Scholar] [CrossRef]
- Cai, L.; Bao, Y.; Fu, X.; Cao, H.; Baranova, A.; Zhang, X.; Sun, J.; Zhang, F. Causal links between major depressive disorder and insomnia: A Mendelian randomisation study. Gene 2021, 768, 145271. [Google Scholar] [CrossRef]
- Hao, W.Z.; Chen, Q.; Wang, L.; Tao, G.; Gan, H.; Deng, L.J.; Huang, J.Q.; Chen, J.X. Emerging roles of long non-coding RNA in depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 115, 110515. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, R.; She, Y.; Liu, X.; Zhao, H.; Li, C.; Jia, Y. A new perspective on depression and neuroinflammation: Non-coding RNA. J. Psychiatr. Res. 2022, 148, 293–306. [Google Scholar] [CrossRef]
- Ballesio, A.; Zagaria, A.; Ottaviani, C.; Steptoe, A.; Lombardo, C. Sleep disturbance, neuro-immune markers, and depressive symptoms in older age: Conditional process analysis from the English Longitudinal Study of Aging (ELSA). Psychoneuroendocrinology 2022, 142, 105770. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Tu, S.; Sheng, J.; Shao, A. Depression in sleep disturbance: A review on a bidirectional relationship, mechanisms and treatment. J. Cell. Mol. Med. 2019, 23, 2324–2332. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Psychiatric Trait | Cardiometabolic Trait | rg | se | Z | p | adj p |
|---|---|---|---|---|---|---|
| SCZ | BMI | −0.10 | 0.01 | −7.07 | 1.6 × 10−12 | 1.9 × 10−10 |
| SCZ | CRP | −0.06 | 0.02 | −3.75 | 0.0002 | 0.02 |
| MDD | BMI | 0.11 | 0.02 | 6.55 | 5.6 × 10−11 | 6.8 × 10−9 |
| MDD | CAD | 0.21 | 0.02 | 10.33 | 5.1 × 10−25 | 6.1 × 10−23 |
| MDD | CRP | 0.11 | 0.02 | 5.50 | 3.8 × 10−8 | 4.6 × 10−6 |
| MDD | T2D | 0.14 | 0.02 | 6.58 | 4.8 × 10−11 | 5.8 × 10−9 |
| PTSD | BMI | 0.32 | 0.04 | 7.77 | 8.1 × 10−15 | 9.7 × 10−13 |
| PTSD | CAD | 0.30 | 0.05 | 6.06 | 6.5 × 10−7 | 7.7 × 10−5 |
| PTSD | CRP | 0.21 | 0.04 | 4.98 | 1.4 × 10−9 | 1.7 × 10−7 |
| PTSD | T2D | 0.25 | 0.05 | 5.21 | 1.9 × 10−7 | 2.2 × 10−5 |
| AN | BMI | −0.31 | 0.02 | −13.52 | 1.3 × 10−41 | 1.5 × 10−39 |
| AN | CRP | −0.28 | 0.03 | −9.28 | 1.7 × 10−20 | 2.1 × 10−18 |
| AN | T2D | −0.20 | 0.03 | −7.37 | 1.8 × 10−13 | 2.1 × 10−11 |
| ADHD | BMI | 0.35 | 0.02 | 14.65 | 1.4 × 10−48 | 1.7 × 10−46 |
| ADHD | CAD | 0.27 | 0.03 | 9.86 | 1.4 × 10−16 | 1.6 × 10−14 |
| ADHD | CRP | 0.30 | 0.04 | 8.27 | 5.9 × 10−23 | 7.1 × 10−21 |
| ADHD | T2D | 0.32 | 0.03 | 12.16 | 5.0 × 10−34 | 6.0 × 10−32 |
| OCD | BMI | −0.29 | 0.04 | −6.97 | 3.2 × 10−12 | 3.8 × 10−10 |
| OCD | CRP | −0.22 | 0.04 | −5.39 | 6.9 × 10−8 | 8.3 × 10−6 |
| OCD | T2D | −0.17 | 0.04 | −3.81 | 0.0001 | 0.02 |
| Psychiatric Trait | Sleep Trait | rg | se | Z | p | adj p |
|---|---|---|---|---|---|---|
| BD | Insomnia | 0.11 | 0.03 | 4.23 | 2.4 × 10−5 | 0.0028 |
| BD | Sleep duration | 0.11 | 0.02 | 4.91 | 9.0 × 10−7 | 0.0011 |
| SCZ | Chronotype | −0.10 | 0.02 | −5.38 | 7.6 × 10−8 | 9.1 × 10−6 |
| SCZ | Sleep duration | 0.15 | 0.02 | 7.37 | 1.7 × 10−13 | 2.1 × 10−11 |
| MDD | Insomnia | 0.44 | 0.03 | 17.60 | 2.3 × 10−69 | 2.8 × 10−67 |
| MDD | Sleep duration | −0.11 | 0.02 | −4.43 | 9.4 × 10−6 | 0.0011 |
| ADHD | Insomnia | 0.37 | 0.03 | 10.67 | 1.4 × 10−26 | 1.7 × 10−24 |
| PTSD | Insomnia | 0.48 | 0.07 | 7.33 | 2.3 × 10−13 | 2.7 × 10−11 |
| PTSD | Sleep duration | −0.23 | 0.06 | −3.86 | 0.0001 | 0.013 |
| ASD | Chronotype | −0.18 | 0.03 | −5.45 | 5.1 × 10−8 | 6.1 × 10−6 |
| AN | Sleep duration | −0.12 | 0.03 | −3.83 | 0.0001 | 0.015 |
| Outcome | Exposure | Method | beta | se | p |
|---|---|---|---|---|---|
| Insomnia | MDD | MR Egger | 0.40 | 0.34 | 0.27 |
| Insomnia | MDD | Weighted median | 0.22 | 0.05 | 2.1 × 10−5 |
| Insomnia | MDD | Inverse variance weighted | 0.24 | 0.06 | 4.1 × 10−5 |
| Insomnia | MDD | Simple mode | 0.19 | 0.10 | 0.06 |
| Insomnia | MDD | MR-PRESSO raw | 0.26 | 0.05 | 6.6 × 10−6 |
| Insomnia | MDD | MR-PRESSO outlier corrected | 0.24 | 0.04 | 2.7 × 10−7 |
| MDD | Insomnia | MR Egger | −0.16 | 0.27 | 0.56 |
| MDD | Insomnia | Weighted median | 0.23 | 0.08 | 0.0027 |
| MDD | Insomnia | Inverse variance weighted | 0.35 | 0.10 | 0.0003 |
| MDD | Insomnia | Simple mode | 0.53 | 0.13 | 0.0018 |
| MDD | Insomnia | MR-PRESSO raw | 0.38 | 0.10 | 0.0020 |
| MDD | Insomnia | MR-PRESSO outlier corrected | 0.38 | 0.07 | 0.0004 |
| SNP | Chr | Gene | EA | OA | b exp | b out | eQTL for Gene (Tissue) | RDB Score |
|---|---|---|---|---|---|---|---|---|
| rs2111592 | 2 | AC007879.1 | A | G | 0.03 | 0.02 | GMPPB (Amygdala, anterior cingulate, caudate, cerebellum, cortex, frontal cortex, hippocampus, hypothalamus, nucleus accumbens, putamen, spinal cord, substantia nigra); GPX1 (Caudate, cerebellum, cortex, frontal cortex, accumbens, putamen); NCKIPSD (Amygdala, anterior cingulate, caudate, cerebellum, cortex, frontal cortex, hippocampus, hypothalamus, nucleus accumbens, putamen, spinal cord); NICN1 (Nucleus accumbens); P4HTM (Cerebellum, cortex, frontal cortex, nucleus accumbens, putamen, spinal cord); QRICH1 (Caudate, cerebellum, nucleus accumbens); RP11-3B7.1 (Anterior cingulate); RP11-694I15.7 (Cerebellum); WDR6 (Cerebellum, nucleus accumbens, putamen) | 0.03 |
| rs66511648 | 3 | RP11-384F7.2 | T | C | 0.03 | 0.02 | - | 0.65 |
| rs9831648 | 3 | Intergenic | T | G | −0.03 | −0.02 | AMT (Anterior cingulate, caudate, cerebellum, cortex, frontal cortex, hippocampus, hypothalamus, nucleus accumbens, putamen, spinal cord, substantia nigra); BSN (Cerebellum); BSN-AS2 (Putamen); CCDC71 (Amygdala, caudate, cerebellum, frontal cortex, putamen); DALRD3 (Cerebellum, cortex) | 0.92 |
| rs30266 | 5 | RP11-6N13.1 | A | G | 0.04 | 0.03 | - | 0.13 |
| rs3099439 | 5 | TMEM161B | T | C | −0.02 | −0.02 | CTC-467M3.3 (Anterior cingulate); CTC-498M16.4 (Amygdala, anterior cingulate, caudate, cerebellum, cortex, frontal cortex, hippocampus, hypothalamus, nucleus accumbens, putamen); TMEM161B-AS1 (Anterior cingulate, caudate, cerebellum, cortex, frontal cortex, hippocampus, hypothalamus, nucleus accumbens, putamen, spinal cord, substantia nigra) | 0.18 |
| rs150186873 | 6 | Intergenic | A | C | −0.07 | −0.04 | BTN2A3P (Cortex) | 0.48 |
| rs10235664 | 7 | MAD1L1 | T | C | 0.03 | 0.02 | FTSJ2 (Cerebellum, caudate); AC110781.3 (Nucleus accumbens) | 0.13 |
| rs61914045 | 12 | ACVR1B | A | G | 0.03 | 0.02 | - | 0.18 |
| rs9536381 | 13 | Intergenic | T | C | 0.03 | 0.03 | - | 0.08 |
| rs1950829 | 14 | LRFN5 | A | G | 0.03 | 0.02 | LRFN5 (Cerebellum) | 0.18 |
| SNP | Chr | Gene | EA | OA | b exp | b out | eQTL for Gene | RDB Score |
|---|---|---|---|---|---|---|---|---|
| rs77960 | 5 | Intergenic | A | G | 0.03 | 0.04 | - | 0.99 |
| rs9563152 | 13 | Intergenic | T | C | 0.04 | 0.02 | RP11-24H2.3 (Anterior cingulate) | 0.18 |
| rs6984111 | 8 | MSRA | C | T | 0.04 | 0.02 | - | 0.14 |
| rs1456193 | 3 | RP11-384F7.2 | T | C | −0.04 | −0.02 | - | 0.18 |
| rs370771 | 6 | LIN28B | G | T | −0.04 | −0.02 | LIN28B-AS1 (Caudate, putamen) HACE1 (Cortex) | 0.59 |
| rs9576155 | 13 | SUPT20H | A | G | 0.03 | 0.02 | ALG5 (Caudate, cortex) | 0.18 |
| rs6938026 | 6 | CUL9 | G | A | 0.04 | 0.02 | CUL9 (Caudate, cortex, frontal cortex, nucleus accumbens, spinal cord) | 0.61 |
| rs77217059 | 2 | LINC01122 | A | T | 0.03 | 0.03 | - | 0.73 |
| MDD | Insomnia | ||||||
|---|---|---|---|---|---|---|---|
| SNP(s) | Chr | Start Locus | Stop Locus | p | adj p | p | adj p |
| rs77217059 | 2 | 57952946 | 59251996 | 3.6 × 10−7 | 2.2 × 10−5 | 6.8 × 10−11 | 4.1 × 10−9 |
| rs2111592 | 2 | 207726595 | 208674588 | 4.7 × 10−12 | 2.8 × 10−10 | 0.046 | 1 |
| rs9831648 | 3 | 47588462 | 50387742 | 4.1 × 10−5 | 0.002 | 7.3 × 10−10 | 4.4 × 10−8 |
| rs66511648, rs1456193 | 3 | 117241645 | 118086929 | 0.068 | 1 | 2.7 × 10−5 | 0.002 |
| rs3099439 | 5 | 87943483 | 89584466 | 2.3 × 10−7 | 1.4 × 10−5 | 1.2 × 10−11 | 7.2 × 10−10 |
| rs30266, rs77960 | 5 | 103788461 | 104850490 | 4.0 × 10−6 | 0.0002 | 0.01 | 0.600 |
| rs150186873 | 6 | 26396201 | 27261035 | 6.8 × 10−12 | 4.1 × 10−10 | 0.09 | 1 |
| rs6938026 | 6 | 42103739 | 43770626 | 0.006 | 0.360 | 1.1 × 10−10 | 6.6 × 10−9 |
| rs370771 | 6 | 104951345 | 106053915 | 0.017 | 1 | 0.003 | 0.180 |
| rs10235664 | 7 | 1366973 | 2473749 | 0.0001 | 0.006 | 0.005 | 0.300 |
| rs6984111 | 8 | 9835864 | 10478851 | 0.003 | 0.180 | 0.057 | 3.420 |
| rs61914045 | 12 | 51769420 | 53039987 | 1.6 × 10−7 | 9.6 × 10−6 | 0.007 | 0.420 |
| rs9576155 | 13 | 37499811 | 38290689 | 0.0084 | 0.504 | 0.0022 | 0.132 |
| rs9536381, rs9563152 | 13 | 53336572 | 54684856 | 0.0001 | 0.008 | 6.6 × 10−5 | 0.004 |
| rs1950829 | 14 | 41614834 | 42562550 | 7.5 × 10−7 | 4.5 × 10−5 | 0.0021 | 0.126 |
| Trait 1 | Trait 2 | rho | r2 | p | adj p |
|---|---|---|---|---|---|
| Locus chr2:57952946-59251996 (SNP: rs77217059) | |||||
| MDD | Insomnia | 0.64 | 0.41 | 8.0 × 10−5 | 0.002 |
| MDD | BMI | 0.09 | 0.01 | 0.43 | 1 |
| MDD | CRP | 0.20 | 0.04 | 0.31 | 1 |
| MDD | T2D | 0.02 | 0.00 | 0.87 | 1 |
| Insomnia | BMI | −0.28 | 0.08 | 0.009 | 1 |
| Insomnia | CRP | 0.07 | 0.00 | 0.70 | 1 |
| Insomnia | T2D | −0.20 | 0.04 | 0.19 | 1 |
| Locus chr3:47588462-50387742 (SNP: rs9831648) | |||||
| MDD | Insomnia | 0.60 | 0.36 | 0.0015 | 0.045 |
| MDD | BMI | 0.48 | 0.23 | 0.0002 | 0.006 |
| MDD | CRP | 0.40 | 0.16 | 0.015 | 0.44 |
| MDD | CAD | 0.36 | 0.12 | 0.08 | 1 |
| Insomnia | BMI | 0.69 | 0.47 | 5.6 × 10−11 | 1.7 × 10−9 |
| Insomnia | CRP | 0.66 | 0.44 | 1.4 × 10−6 | 4.2 × 10−5 |
| Insomnia | CAD | 0.54 | 0.29 | 0.001 | 0.033 |
| Locus chr5:87943483-89584466 (SNP: rs3099439) | |||||
| MDD | Insomnia | −0.05 | 0.00 | 0.77 | 1 |
| MDD | BMI | 0.23 | 0.05 | 0.048 | 1 |
| MDD | CRP | −0.07 | 0.01 | 0.71 | 1 |
| MDD | T2D | −0.16 | 0.02 | 0.46 | 1 |
| MDD | CAD | 0.35 | 0.12 | 0.03 | 0.9 |
| Insomnia | BMI | −0.14 | 0.02 | 0.17 | 1 |
| insomnia | CAD | −0.13 | 0.02 | 0.35 | 1 |
| Insomnia | CRP | 0.02 | 0.00 | 0.91 | 1 |
| Insomnia | T2D | −0.25 | 0.06 | 0.19 | 1 |
| Locus chr13:53336572-54684856 (SNPs: rs9536381, rs9563152) | |||||
| MDD | Insomnia | 1.00 | 1.00 | 3.3 × 10−6 | 9.9 × 10−5 |
| MDD | BMI | 0.48 | 0.23 | 0.0004 | 0.012 |
| MDD | CAD | 0.28 | 0.07 | 0.208 | 1 |
| MDD | CRP | 0.54 | 0.30 | 0.016 | 0.47 |
| Insomnia | BMI | 0.55 | 0.31 | 3.9 × 10−5 | 0.001 |
| Insomnia | CAD | 0.43 | 0.18 | 0.042 | 1 |
| Insomnia | CRP | 1.00 | 1.00 | 7.3 × 10−6 | 0.0002 |
| Trait 1 | Trait 2 | Z | r2_Trait 1_Z | r2_Trait 2_Z | rho Partial Correlation | p Partial Correlation |
|---|---|---|---|---|---|---|
| Locus chr3:47588462-50387742 (SNP: rs9831648) | ||||||
| MDD | Insomnia | BMI | 0.23 | 0.47 | 0.41 | 0.11 |
| MDD | Insomnia | CRP | 0.16 | 0.44 | 0.48 | 0.08 |
| MDD | Insomnia | BMI, CRP | 0.25 | 0.57 | 0.39 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zammarchi, G.; Conversano, C.; Pisanu, C. Investigating Shared Genetic Bases between Psychiatric Disorders, Cardiometabolic and Sleep Traits Using K-Means Clustering and Local Genetic Correlation Analysis. Algorithms 2022, 15, 409. https://doi.org/10.3390/a15110409
Zammarchi G, Conversano C, Pisanu C. Investigating Shared Genetic Bases between Psychiatric Disorders, Cardiometabolic and Sleep Traits Using K-Means Clustering and Local Genetic Correlation Analysis. Algorithms. 2022; 15(11):409. https://doi.org/10.3390/a15110409
Chicago/Turabian StyleZammarchi, Gianpaolo, Claudio Conversano, and Claudia Pisanu. 2022. "Investigating Shared Genetic Bases between Psychiatric Disorders, Cardiometabolic and Sleep Traits Using K-Means Clustering and Local Genetic Correlation Analysis" Algorithms 15, no. 11: 409. https://doi.org/10.3390/a15110409
APA StyleZammarchi, G., Conversano, C., & Pisanu, C. (2022). Investigating Shared Genetic Bases between Psychiatric Disorders, Cardiometabolic and Sleep Traits Using K-Means Clustering and Local Genetic Correlation Analysis. Algorithms, 15(11), 409. https://doi.org/10.3390/a15110409