
Fully Quantum Modeling of Exciton Diffusion in Mesoscale Light Harvesting Systems



Abstract
1. Introduction
2. Model of Mesoscale LHCs and Methodologies
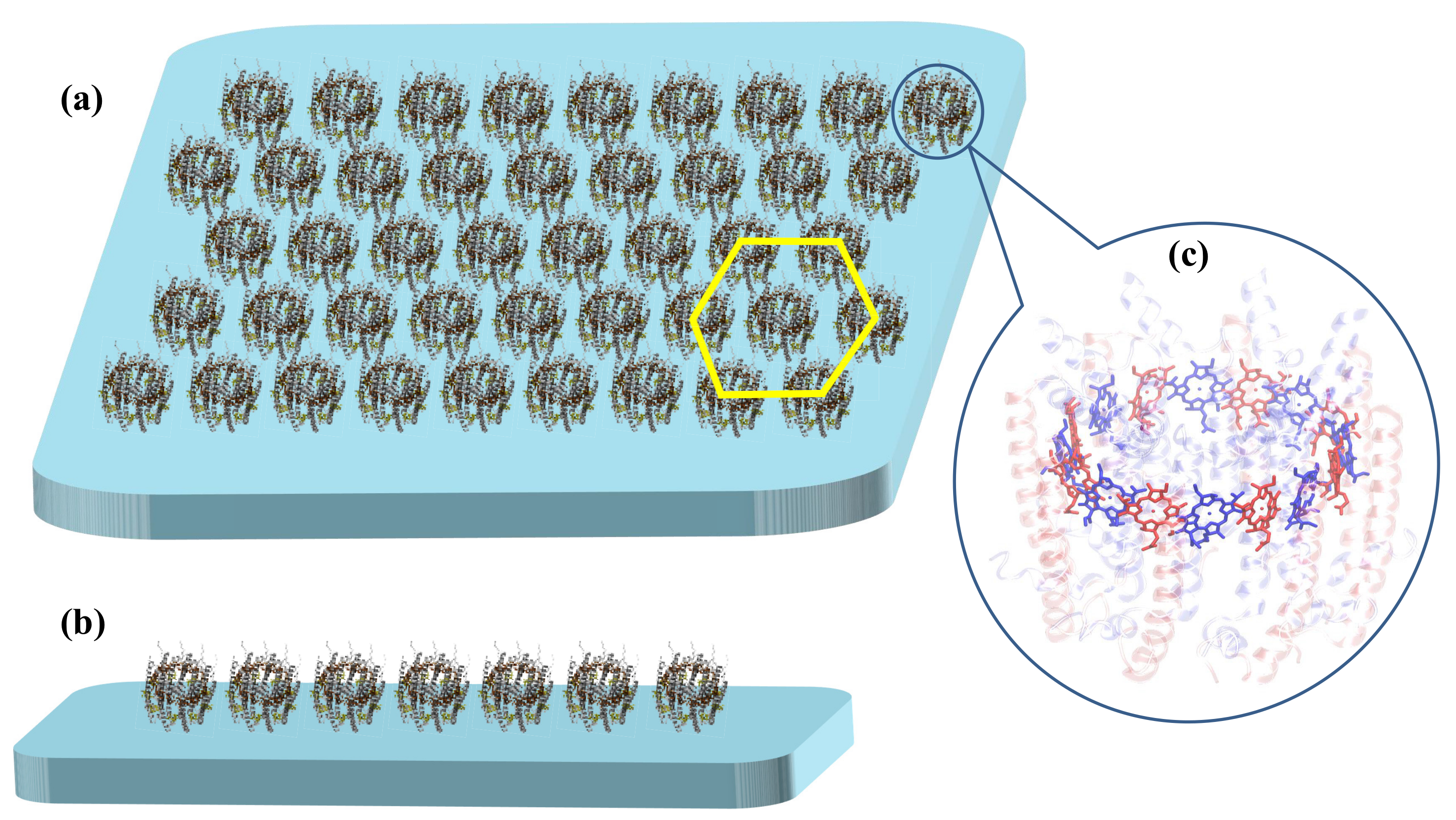
2.1. B850 Nanoarrays
2.2. Quantum Coherent Modeling for Exciton-Phonon Dynamics
2.3. Incoherent Inter-B850 Exciton Hopping
3. Results and Discussion
3.1. Coherent Exciton Transfer in B850 Nanoarrays
3.1.1. Parameterization of the Phonon Bath
3.1.2. Computational Advantage of GPU
3.1.3. Effect of Initial Excitation Conditions
3.1.4. Exciton Delocalization
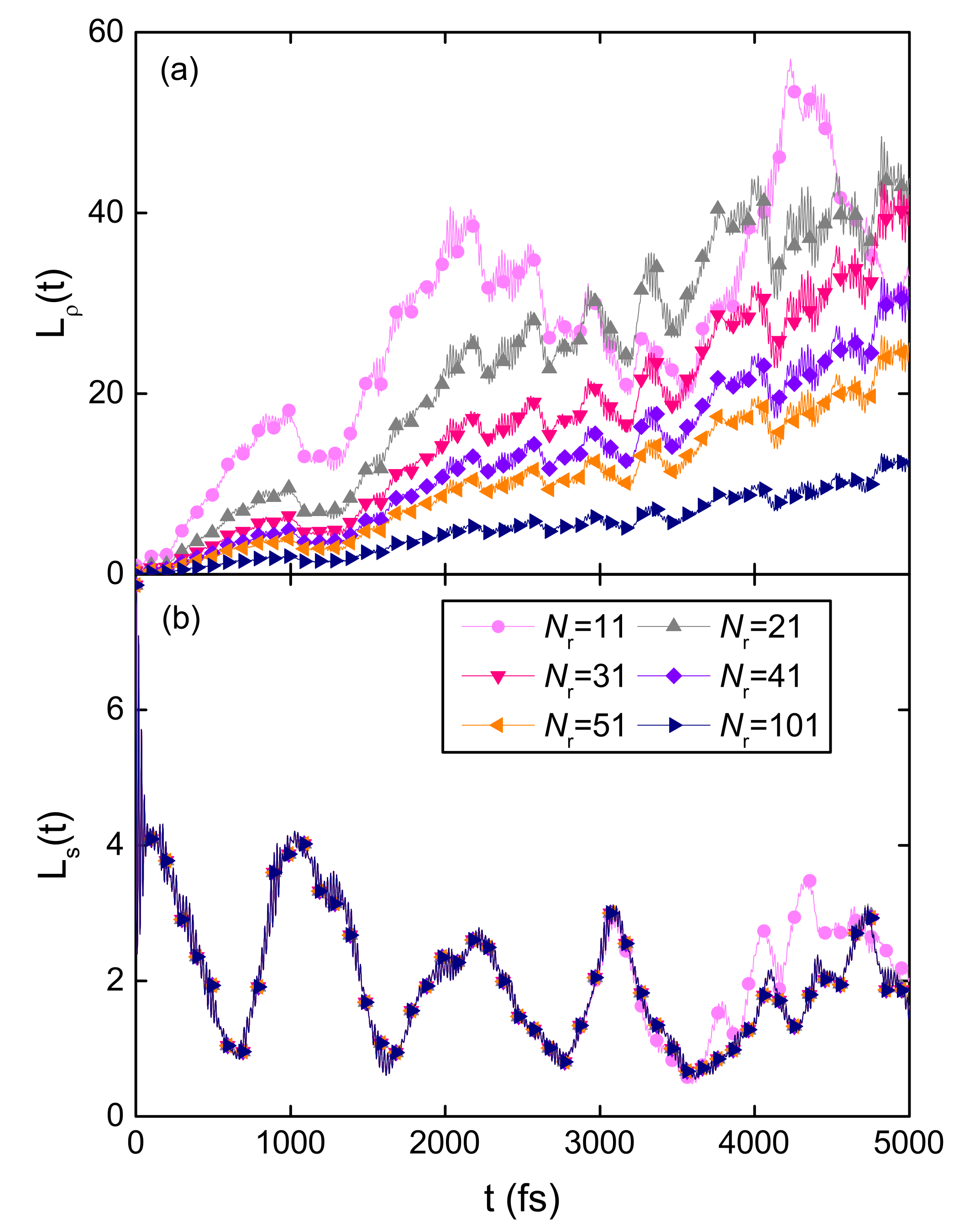
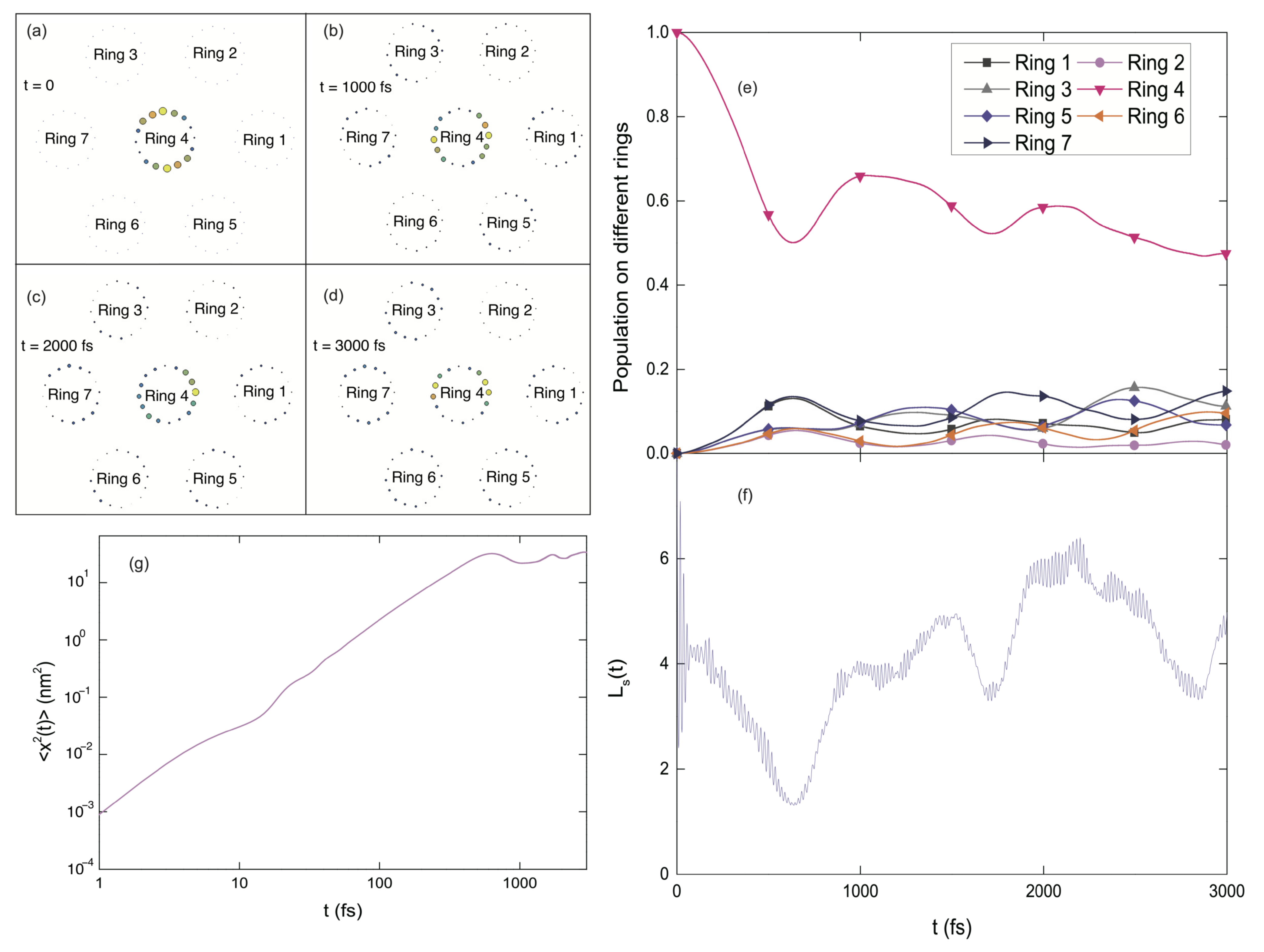
3.1.5. Fractal Analysis of Exciton Dynamics
3.1.6. Exciton Diffusion in 2D B850 Nanoarray
3.2. Incoherent Exciton Hopping in B850 Nanoarrays
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Exciton Hamiltonian of B850 Nanoarrays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 205 | −52 | 14.4 | −7 | 2.9 | −2.2 | 1.2 | −1.4 | 1.2 | −2.2 | 2.9 | −7 | 14.4 | −52 | 144 | |
| 2 | 144 | −31 | 14.4 | −4 | 2.9 | −0.9 | 1.2 | 0.4 | 1.2 | −0.9 | 2.9 | −4 | 14.4 | −31 | ||
| 3 | 205 | −52 | 14.4 | −7 | 2.9 | −2.2 | 1.2 | −1.4 | 1.2 | −2.2 | 2.9 | −7 | 14.4 | |||
| 4 | 144 | −31 | 14.4 | −4 | 2.9 | −0.9 | 1.2 | 0.4 | 1.2 | −0.9 | 2.9 | −4 | ||||
| 5 | 205 | −52 | 14.4 | −7 | 2.9 | −2.2 | 1.2 | −1.4 | 1.2 | −2.2 | 2.9 | |||||
| 6 | 144 | −31 | 14.4 | −4 | 2.9 | −0.9 | 1.2 | 0.4 | 1.2 | −0.9 | ||||||
| 7 | 205 | −52 | 14.4 | −7 | 2.9 | −2.2 | 1.2 | −1.4 | 1.2 | |||||||
| 8 | 144 | −31 | 14.4 | −4 | 2.9 | −0.9 | 1.2 | 0.4 | ||||||||
| 9 | 205 | −52 | 14.4 | −7 | 2.9 | −2.2 | 1.2 | |||||||||
| 10 | 144 | −31 | 14.4 | −4 | 2.9 | -0.9 | ||||||||||
| 11 | 205 | −52 | 14.4 | −7 | 2.9 | |||||||||||
| 12 | 144 | −31 | 14.4 | −4 | ||||||||||||
| 13 | 205 | −52 | 14.4 | |||||||||||||
| 14 | 144 | −31 | ||||||||||||||
| 15 | 205 | |||||||||||||||
| 16 |
Appendix B. Derivation of Exciton-Phonon Dynamics in B850 Nanoarrays
References
- Amerongen, H.V.; Valkunas, L.; van Grondell, R. Photosynthetic Excitons; World Scientific: Singapore, 2000. [Google Scholar]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; World Scientific: London, UK, 2002. [Google Scholar]
- Mirkovic, T.; Ostrumov, E.E.; Anna, J.M.; van Grondelle, R.; Govindjee; Scholes, G.D. Light absorption and energy transfer in the antenna complexes of photosynthetic organisms. Chem. Rev. 2017, 117, 249–293. [Google Scholar] [CrossRef] [PubMed]
- Curutchet, C.; Mennucci, B. Quantum chemical studies of light harvesting. Chem. Rev. 2017, 117, 294–343. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Paleček, D.; Caycedo-Soler, F.; Lincoln, C.N.; Prior, J.; von Berlepsch, H.; Huelga, S.F.; Plenio, M.B.; Zigmantas, D.; Hauer, J. Vibronic origin of long-lived coherence in an artificial molecular light harvester. Nat. Commun. 2015, 6, 7755. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Patra, A. Nanoscale strategies for light harvesting. Chem. Rev. 2017, 117, 712–757. [Google Scholar] [CrossRef] [PubMed]
- Wientjes, E.; Renger, J.; Curto, A.G.; Cogdell, R.; van Hulst, N.F. Strong antenna-enhanced fluorescence of a single light-harvesting complex shows photon antibunching. Nat. Commun. 2014, 5, 4236. [Google Scholar] [CrossRef]
- Driscoll, B.; Lunceford, C.; Lin, S.; Woronowicz, K.; Niederman, R.A.; Woodbury, N.W. Energy transfer properties of rhodobacter sphaeroides chromatophores during adaptation to low light intensity. Phys. Chem. Chem. Phys. 2014, 16, 17133–17141. [Google Scholar] [CrossRef]
- Escalante, M.; Lenferink, A.; Zhao, Y.; Tas, N.; Huskens, J.; Hunter, C.N.; Subramaniam, V.; Otto, C. Long-range energy propagation in nanometer arrays of light harvesting antenna complexes. Nano Lett. 2010, 10, 1450–1457. [Google Scholar] [CrossRef]
- Huang, X.; Vasilev, C.; Hunter, C.N. Excitation energy transfer between monomolecular layers of light harvesting lh2 and lh1-reaction centre complexes printed on a glass substrate. Lab Chip 2020, 20, 2529–2538. [Google Scholar] [CrossRef]
- Lloyd, S.; Mohseni, M. Symmetry-enhanced supertransfer of delocalized quantum states. New J. Phys. 2010, 12, 075020. [Google Scholar] [CrossRef]
- Bredas, J.L.; Sargent, E.H.; Scholes, G.D. The path towards sustainable energy. Nat. Mater. 2016, 16, 35–44. [Google Scholar]
- Singharoy, A.; Maffeo, C.; Delgado-Magnero, K.H.; Swainsbury, D.J.; Sener, M.; Kleinekathöfer, U.; Vant, J.W.; Nguyen, J.; Hitchcock, A.; Isralewitz, B. Atoms to phenotypes: Molecular design principles of cellular energy metabolism. Cell 2019, 179, 1098–1111.e23. [Google Scholar] [CrossRef]
- Föster, T. Zwischenmolekulare energiewanderung und fluoreszenz. Ann. Phys. 1948, 2, 55–75. [Google Scholar] [CrossRef]
- Engel, G.S.; Calhoun, T.R.; Read, R.L.; Ahn, T.K.; Mancal, T.; Cheng, Y.C.; Blankenship, R.E.; Fleming, G.R. Evidence for wavelike energy transfer through quantum coherence in photosynthetic systems. Nature 2007, 446, 782–786. [Google Scholar] [CrossRef]
- Lee, H.; Cheng, Y.C.; Fleming, G.R. Coherence dynamics in photosynthesis: Protein protection of excitonic coherence. Science 2007, 316, 1462–1465. [Google Scholar] [CrossRef]
- Collini, E.; Wong, C.Y.; Wilk, K.E.; Curmi, P.M.G.; Brumer, P.; Scholes, G.D. Coherently wired light-harvesting in photosynthetic marine algae at ambient temperature. Science 2010, 463, 644–647. [Google Scholar] [CrossRef]
- Jang, S.; Newton, M.D.; Silbey, R.J. Multichromophoric Föster resonance energy transfer. Phys. Rev. Lett. 2004, 92, 218301. [Google Scholar] [CrossRef]
- Cleary, L.; Chen, H.; Chuang, C.; Silbey, R.J.; Cao, J. Optimal fold symmetry of LH2 rings on a photosynthetic membrane. Proc. Natl. Acad. Sci. USA 2013, 110, 8537–8542. [Google Scholar] [CrossRef]
- Redfield, A.G. The theory of relaxation processes. Adv. Magn. Reson 1965, 1, 1–32. [Google Scholar]
- Tanimura, Y.; Kubo, R. Time evolution of a quantum system in contact with a nearly gaussian-markoffian noise bath. J. Phys. Soc. Jpn. 1989, 58, 101–114. [Google Scholar] [CrossRef]
- Kondov, I.; Toss, M.; Wang, H. Theoretical study of ultrafast heterogeneous electron transfer reactions at dye-semiconductor interfaces: Coumarin 343 at titanium oxide. J. Phys. Chem. A 2006, 110, 1364–1374. [Google Scholar] [CrossRef]
- Strümpfer, J.; Schulten, K. Light harvesting complex II B850 excitation dynamics. J. Chem. Phys. 2009, 131, 225101. [Google Scholar] [CrossRef]
- Yeh, S.H.; Zhu, J.; Kais, S. Population and coherence dynamics in light harvesting complex II (LH2). J. Chem. Phys. 2012, 137, 084110. [Google Scholar] [CrossRef]
- Suess, D.; Eisfeld, A.; Strunz, W.T. Hierarchy of stochastic pure states for open quantum system dynamics. Phys. Rev. Lett. 2014, 113, 150403. [Google Scholar] [CrossRef]
- Varvelo, L.; Lynd, J.K.; Bennett, D.I.G. Formally exact, arbitrarily scalable simulations of exciton dynamics in molecular materials. Chem. Sci. 2021. [Google Scholar] [CrossRef]
- Davydov, A.S.; Kislukha, N.I. Solitary excitons in one-dimensional molecular chains. Phys. Status Solidi B 1973, 59, 465–470. [Google Scholar] [CrossRef]
- Davydov, A.S. Solitons in Molecular Systems; Springer: Dordrecht, The Netherlands, 1985. [Google Scholar]
- Sun, J.; Luo, B.; Zhao, Y. Dynamics of a one-dimensional holstein polaron with the davydov ansätze. Phys. Rev. B 2010, 82, 014305. [Google Scholar] [CrossRef]
- Luo, B.; Ye, J.; Guan, C.B.; Zhao, Y. Validity of time-dependent trial states for the holstein polaron. Phys. Chem. Chem. Phys. 2010, 12, 15073–15084. [Google Scholar] [CrossRef] [PubMed]
- Sundström, V.; Pullerits, T.; van Grondelle, R. Photosynthetic light-harvesting: Reconciling dynamics and structure of purple bacterial LH2 reveals function of photosynthetic unit. J. Phys. Chem. B 1999, 103, 2327–2346. [Google Scholar] [CrossRef]
- Smyth, C.; Oblinsky, D.G.; Scholes, G.D. B800-B850 coherence correlates with energy transfer rates in the LH2 complex of photosynthetic purple bacteria. Phys. Chem. Chem. Phys. 2015, 17, 30805–30816. [Google Scholar] [CrossRef]
- Van Der Vegte, C.P.; Prajapati, J.D.; Kleinekathöfer, U.; Knoester, J.; Jansen, T.L.C. Atomistic modeling of two-dimensional electronic spectra and excited-state dynamics for a light harvesting 2 complex. J. Phys. Chem. B 2015, 119, 1302–1313. [Google Scholar] [CrossRef]
- Caycedo-Soler, F.; Lim, J.; Oviedo-Casado, S.; van Hulst, N.F.; Huelga, S.F.; Plenio, M.B. Theory of excitonic delocalization for robust vibronic dynamics in lh2. J. Phys. Chem. Lett. 2018, 9, 3446–3453. [Google Scholar] [CrossRef]
- Jang, S.J. Robust and fragile quantum effects in the transfer kinetics of delocalized excitons between b850 units of lh2 complexes. J. Phys. Chem. Lett. 2018, 9, 6576–6583. [Google Scholar] [CrossRef]
- Ye, J.; Sun, K.; Zhao, Y.; Yu, Y.J.; Lee, C.K.; Cao, J.S. Excitonic energy transfer in light-harvesting complexes in purple bacteria. J. Chem. Phys. 2012, 136, 245104. [Google Scholar] [CrossRef]
- Sun, K.; Ye, J.; Zhao, Y. Path induced coherent energy transfer in light-harvesting complexes in purple bacteria. J. Chem. Phys. 2014, 141, 124103. [Google Scholar] [CrossRef]
- Márquez, A.S.; Chen, L.P.; Sun, K.; Zhao, Y. Probing ultrafast excitation energy transfer of the chlorosome with exciton–phonon variational dynamics. Phys. Chem. Chem. Phys. 2016, 18, 20298–20311. [Google Scholar] [CrossRef]
- Cogdell, R.J.; Gall, A.; Kohler, J. The architecture and function of the light-harvesting apparatus of purple bacteria: From single molecules to in vivo membranes. Q. Rev. Biophys. 2006, 39, 227–324. [Google Scholar] [CrossRef]
- McDermott, G.; Prince, S.M.; Freer, A.A.; Hawthormthwaite-Lawless, A.M.; Papiz, M.Z.; Cogdell, R.J.; Issacs, N.W. Crystal structure of an integral membrane light-harvesting complex from photosynthetic bacteria. Nature 1995, 374, 517–521. [Google Scholar] [CrossRef]
- Koepke, J.; Hu, X.; Muenke, C.; Schulten, K.; Michel, H. The crystal structure of the light-harvesting complex II (B800-850) from Rhodospirillum molischianum. Structure 1996, 4, 581–597. [Google Scholar] [CrossRef]
- Fidler, A.F.; Singh, V.P.; Long, P.D.; Dahlberg, P.D.; Engel, G.S. Probing energy transfer events in the light harvesting complex 2 (LH2) of rhodobacter sphaeroides with two-dimensional spectroscopy. J. Chem. Phys. 2013, 139, 155101. [Google Scholar] [CrossRef]
- Van Grondelle, R.; Novoderezhkin, V.I. Energy transfer in photosynthesis: Experimental insights and quantitative models. Phys. Chem. Chem. Phys. 2006, 8, 793–807. [Google Scholar] [CrossRef]
- Fassioli, F.; Dinshaw, R.; Arpin, P.C.; Scholes, G.D. Photosynthetic light harvesting: Excitons and coherence. J. R. Soc. Interface 2014, 11, 20130901. [Google Scholar] [CrossRef]
- Holstein, T. Studies of polaron motion: Part I. the molecular-crystal model. Ann. Phys. 1959, 8, 325–342. [Google Scholar] [CrossRef]
- Mahan, G.D. Many Particle Physics, 3rd ed.; Kluwer Academic: New York, NY, USA, 2000. [Google Scholar]
- Hu, X.; Ritz, T.; Damjanović, A.; Schulten, K. Pigment organization and transfer of electronic excitation in the photosynthetic unit of purple bacteria. J. Phys. Chem. B 1997, 101, 3854–3871. [Google Scholar] [CrossRef]
- Luo, B.; Ye, J.; Zhao, Y. Variational study of polaron dynamics with the davydov ansätze. Phys. Status Solidi C 2011, 8, 70–73. [Google Scholar] [CrossRef]
- Itzykson, C.; Zuber, J.B. Quantum Field Theory; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Meier, T.; Zhao, Y.; Chernyak, V.; Mukamel, S. Polarons, localization, and excitonic coherence in superradiance of biologicalantenna complexes. J. Chem. Phys. 1997, 107, 3876–3893. [Google Scholar] [CrossRef]
- Zhao, Y.; Meier, T.; Zhang, W.M.; Chernyak, V.; Mukamel, S. Superradiance coherence sizes in single molecule spectroscopy of LH2 antenna complexes. J. Phys. Chem. B 1999, 103, 3954–3962. [Google Scholar] [CrossRef]
- Yang, G.; Wu, N.; Chen, T.; Sun, K.; Zhao, Y. Theoretical examination of long-range energy propagation in nano-engineered light-harvesting antenna arrays. J. Phys. Chem. C 2012, 116, 3747–3756. [Google Scholar] [CrossRef]
- Damjanović, A.; Kosztin, I.; Kleinekathöfer, U.; Schulten, K. Excitons in a photosynthetic light-harvesting system: A combined molecular dynamics, quantum chemistry, and polaron model study. Phys. Rev. E 2002, 65, 031919. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.J.; Chen, L.P.; Zhao, Y. Dephasing and dissipation in a source-drain model of light harvesting systems. Chem. Phys. Chem. 2014, 15, 2859–2870. [Google Scholar] [CrossRef] [PubMed]
- Dahlbom, M.; Pullerits, T.; Mukamel, S.; Sundström, V. Exciton delocalization in the B850 light-harvesting complex: Comparison of different measures. J. Phys. Chem. B 2001, 105, 5515–5524. [Google Scholar] [CrossRef]
- Meier, T.; Chernyak, V.; Mukamel, S. Multiple exciton coherence sizes in photosynthetic antenna complexes viewed by pump-probe spectroscopy. J. Phys. Chem. B 1997, 101, 7332–7342. [Google Scholar] [CrossRef]
- Moix, J.M.; Zhao, Y.; Cao, J. Equilibrium-reduced density matirx formulation: Influence of noise, disorder, and temperature on localization in excitonic systems. Phys. Rev. E 2012, 85, 115412. [Google Scholar] [CrossRef]
- Hu, G.Y.; O’Connel, F. Analytical inversion of symmetric tridiagonal matrices. J. Phys. A Math. Gen. 1996, 29, 1511–1513. [Google Scholar] [CrossRef]
- Glauber, R.J. Time-dependent statistics of the ising model. J. Math. Phys 1963, 4, 294–307. [Google Scholar] [CrossRef]
- Wrigley, J.D.; Twigg, M.E.; Ehrlich, G. Lattice walks by long jumps. J. Chem. Phys. 1990, 93, 2885–2902. [Google Scholar] [CrossRef]
- Ferretti, M.; Hendrikx, R.; Romero, E.; Southall, J.; Cogdell, R.J.; Novoderezhkin, V.I.; Scholes, G.D.; van Grondelle, R. Dark states in the light-harvesting complex 2 revealed by two-dimensional electronic spectroscopy. Sci. Rep. 2016, 6, 20834. [Google Scholar] [CrossRef]
- Dahlberg, P.; Fidler, A.; Caram, J.; Long, P.; Engel, G.S. Energy transfer observed in live cells using two-dimensional electronic spectroscopy. J. Phys. Chem. Lett. 2013, 4, 3636–3640. [Google Scholar] [CrossRef]
- Harel, E.; Long, P.D.; Engel, G.S. Single-shot ultrabroadband two-dimensional electronic spectroscopy of the light-harvesting complex lh2. Opt. Lett. 2011, 36, 1665–1667. [Google Scholar] [CrossRef]
- Fidler, A.F.; Singh, V.P.; Long, P.D.; Dahlberg, P.D.; Engel, G.S. Dynamic localization of electronic excitation in photosynthetic complexes revealed with chiral two-dimensional spectroscopy. Nat. Commun. 2014, 5, 3286. [Google Scholar] [CrossRef]
- Chenu, A.; Scholes, G.D. Coherence in energy transfer and photosynthesis. Annu. Rev. Phys. Chem. 2015, 66, 69–96. [Google Scholar] [CrossRef]
- Monshouwer, R.; Abrahamsson, M.; van Mourik, F.; van Grondelle, R. Superradiance and exciton delocalization in bacterial photosynthetic light-harvesting systems. J. Phys. Chem. B 1997, 101, 7241–7248. [Google Scholar] [CrossRef]
- Márquez, A.S.; Sun, K.; Molina, R.; Zhao, Y. Dynamics of coherence, localization and excitation transfer in disordered nanorings. Phys. Chem. Chem. Phys. 2017, 19, 25996–26013. [Google Scholar]
- Zhou, N.; Huang, Z.; Zhu, J.; Chernyak, V.; Zhao, Y. Polaron dynamics with a multitude of davydov D2 trial states. J. Chem. Phys. 2015, 143, 014113. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, L.; Wu, C.; Chen, L.; Grossmann, F.; Zhao, Y. Polaron dynamics with off-diagonal coupling: Beyond the ehrenfest approximation. Phys. Chem. Chem. Phys. 2017, 19, 1655–1668. [Google Scholar] [CrossRef]
- Wang, L.; Fujihashi, Y.; Chen, L.; Zhao, Y. Finite-temperature time-dependent variation with multiple davydov states. J. Chem. Phys. 2017, 146, 124127. [Google Scholar] [CrossRef]
- Zheng, F.; Jin, M.; Mančal, T.; Zhao, Y. Study of electronic structures and pigment–protein interactions in the reaction center of thermochromatium tepidum with a dynamic environment. J. Phys. Chem. B 2016, 120, 10046–10058. [Google Scholar] [CrossRef]
- Novoderezhkin, V.I.; Palacios, M.A.; van Amerongen, H.; van Grondelle, R. Energy-transfer dynamics in the lhcii complex of higher plants: Modified redfield approach. J. Phys. Chem. B 2004, 108, 10363–10375. [Google Scholar] [CrossRef]
- Renger, T.; Marcus, R.A. On the relation of protein dynamics and exciton relaxation in pigment–protein complexes: An estimation of the spectral density and a theory for the calculation of optical spectra. J. Chem. Phys. 2002, 116, 9997–10019. [Google Scholar] [CrossRef]
- Georgakopoulou, S.; Frese, R.N.; Johnson, E.; Koolhaas, C.; Cogdell, R.J.; van Grondelle, R.; van der Zwan, G. Absorption and cd spectroscopy and modeling of various lh2 complexes from purple bacteria. Biophys. J. 2002, 82, 2184–2197. [Google Scholar] [CrossRef]
- Tei, G.; Nakatani, M.; Ishihara, H. The function of geometrical structure of B850 ring in bacterial photosynthetic antenna systems. Phys. Status Solidi B 2011, 248, 399–402. [Google Scholar] [CrossRef]
- Lebovitch, L.S.; Shehadeh, L.A. Introduction to fractals. In Tutorials in Contemporary Nonlinear Methods for the Behavioral Sciences; Riley, M.A., Orden, G.C.V., Eds.; National Science Foundation: Arlington, VA, USA, 2005; Chapter 5; pp. 178–266. Available online: https://www.nsf.gov/pubs/2005/nsf05057/nmbs/nmbs.pdf (accessed on 6 April 2018).
- Riley, M.A.; Bonnette, S.; Kuznetsov, N.; Wallot, S.; Gao, J.B. A tutorial introduction to adaptive fractal analysis. Front. Physiol. 2012, 3, 371. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.B.; Cao, Y.H.; Tung, W.W.; Hu, J. Multiscale Analysis of Complex Time Series: Integration of Chaos and Random Fractal Theory, and Beyond; Wiley Interscience: Hoboken, NJ, USA, 2007. [Google Scholar]
- Hu, J.; Gao, J.B.; Wang, X.S. Multifractal analysis of sunspot time series: The effects of the 11-year cycle and fourier truncation. J. Stat. Mech. 2009, 2009, P02066. [Google Scholar] [CrossRef]
- Gao, J.B.; Hu, J.; Tung, W.W. Facilitating joint chaos and fractal analysis of biosignals through nonlinear adaptive filtering. PLoS ONE 2011, 6, e24331. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.B.; Sultan, H.; Hu, J.; Tung, W.W. Denoising nonlinear time series by adaptive filtering and wavelet shrinkage: A comparison. IEEE Signal Process. Lett. 2010, 17, 237–240. [Google Scholar]
- Kuznetsov, N.; Bonnette, S.; Gao, J.B.; Riley, M.A. Adaptive fractal analysis reveals limits to fractal scaling in center of pressure trajectories. Ann. Biomed. Eng. 2013, 41, 1646–1660. [Google Scholar] [CrossRef]
- Gao, J.B.; Hu, J.; Mao, X.; Perc, M. Culturomics meets random fractal theory: Insights into long-range correlations of social and natural phenomena over the past two centuries. J. R. Soc. Interface 2012, 9, 1956–1964. [Google Scholar] [CrossRef]
- Gao, J.B.; Hu, J.; Tung, W.W.; Cao, Y.H.; Sarshar, N.; Roychowdhury, V.P. Assessment of long-range correlation in time series: How to avoid pitfalls. Phys. Rev. E 2006, 73, 016117. [Google Scholar] [CrossRef]
- Wan, Y.; Guo, Z.; Zhu, T.; Yan, S.; Johnson, J.; Huang, L. Cooperative singlet and triplet exciton transport in tetracene crystals visualized by ultrafast microscopy. Nat. Chem. 2015, 7, 785–792. [Google Scholar] [CrossRef]
- Ritz, T.; Park, S.; Schulten, K. Kinetics of excitation migration and trapping in the photosynthetic unit of purple bacteria. J. Phys. Chem. B 2001, 105, 8259–8267. [Google Scholar] [CrossRef]
- Caycedo, S.F.; Rodrguez, F.J.; Quiroga, L.; Johnson, N.F. Light-harvesting mechanism of bacteria exploits a critical interplay between the dynamics of transport and trapping. Phys. Rev. Lett. 2010, 104, 158302. [Google Scholar] [CrossRef]
- Godik, V.I.; Borisov, A.Y. Excitation trapping by different states of photosynthetic reaction centres. FEBS Lett. 1977, 82, 355–358. [Google Scholar] [CrossRef]
- Campiloo, A.J.; Hyer, R.C.; Monger, T.G.; Parson, W.W.; Shapiro, S.L. Light collection and harvesting processes in bacterial photosynthesis investigated on a picosecond time scale. Proc. Natl. Acad. Sci. USA 1977, 74, 1997–2001. [Google Scholar] [CrossRef]
- Wendling, M.; van Mourik, F.; van Stokkum, I.H.M.; Salverda, J.M.; Michel, H.; van Grondelle, R. Low-intensity pump-probe measurements on the B800 band of rhodospirillum molischianum. Biophys. J. 2003, 84, 440–449. [Google Scholar] [CrossRef][Green Version]
- Akselrod, G.M.; Deotare, P.B.; Thompson, N.J.; Lee, J.; Tisdale, W.A.; Baldo, M.A.; Menon, V.M.; Bulovic, V. Visualization of exciton transport in ordered and disordered molecular solids. Nat. Commun. 2014, 5, 3646. [Google Scholar] [CrossRef]
- Chhowalla, M.; Shin, H.S.; Eda, G.; Li, L.; Loh, K.P.; Zhang, H. The chemistry of two-dimensional layered transition metal dichalcogenide nanosheets. Nat. Chem. 2013, 5, 263–275. [Google Scholar] [CrossRef]
- Madjet, M.E.; Abdurahman, A.; Renger, T. Intermolecular coulomb couplings from ab initio electrostatic potentials: Application to optical transitions of strongly coupled pigments in photosynthetic antennae and reaction centers. J. Phys. Chem. B 2006, 110, 17268–17281. [Google Scholar] [CrossRef]
- Renger, T. Theory of excitation energy transfer: From structure to function. Photosynth. Res. 2009, 102, 471–485. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, F.; Chen, L.; Gao, J.; Zhao, Y. Fully Quantum Modeling of Exciton Diffusion in Mesoscale Light Harvesting Systems. Materials 2021, 14, 3291. https://doi.org/10.3390/ma14123291
Zheng F, Chen L, Gao J, Zhao Y. Fully Quantum Modeling of Exciton Diffusion in Mesoscale Light Harvesting Systems. Materials. 2021; 14(12):3291. https://doi.org/10.3390/ma14123291
Chicago/Turabian StyleZheng, Fulu, Lipeng Chen, Jianbo Gao, and Yang Zhao. 2021. "Fully Quantum Modeling of Exciton Diffusion in Mesoscale Light Harvesting Systems" Materials 14, no. 12: 3291. https://doi.org/10.3390/ma14123291
APA StyleZheng, F., Chen, L., Gao, J., & Zhao, Y. (2021). Fully Quantum Modeling of Exciton Diffusion in Mesoscale Light Harvesting Systems. Materials, 14(12), 3291. https://doi.org/10.3390/ma14123291