From a Basic Microalga and an Acetic Acid Bacterium Cellulose Producer to a Living Symbiotic Biofilm

 ,
,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
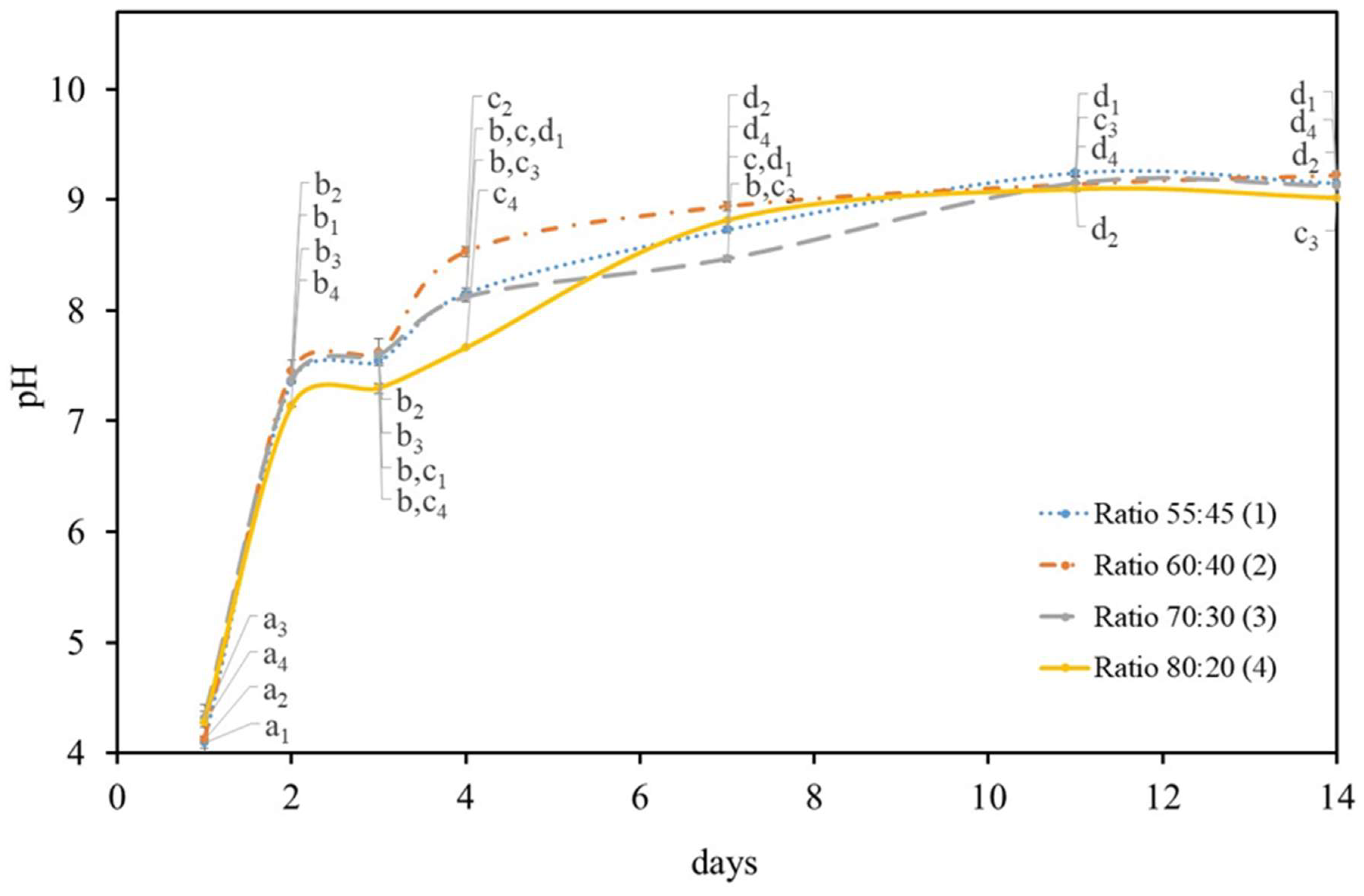
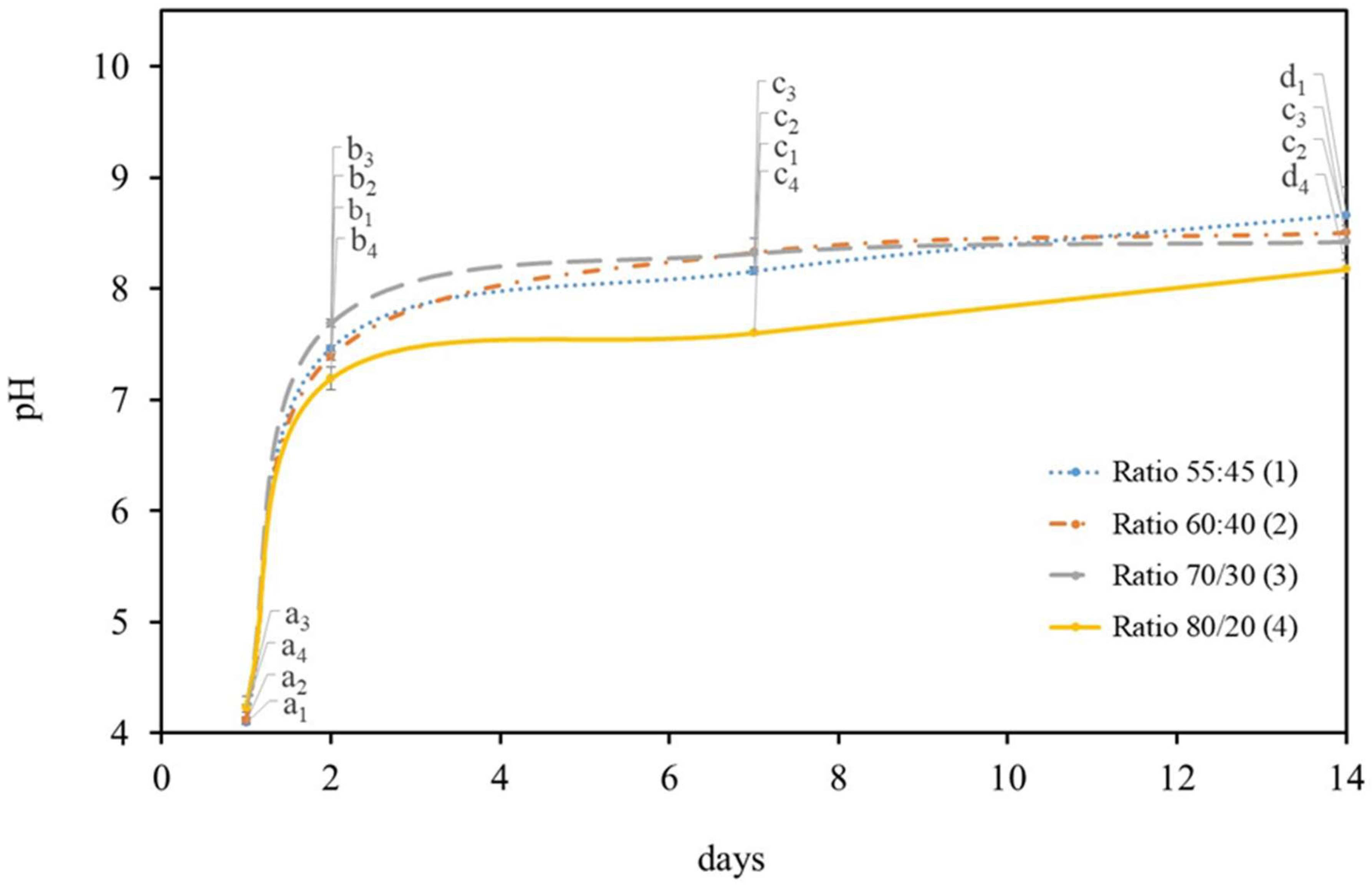
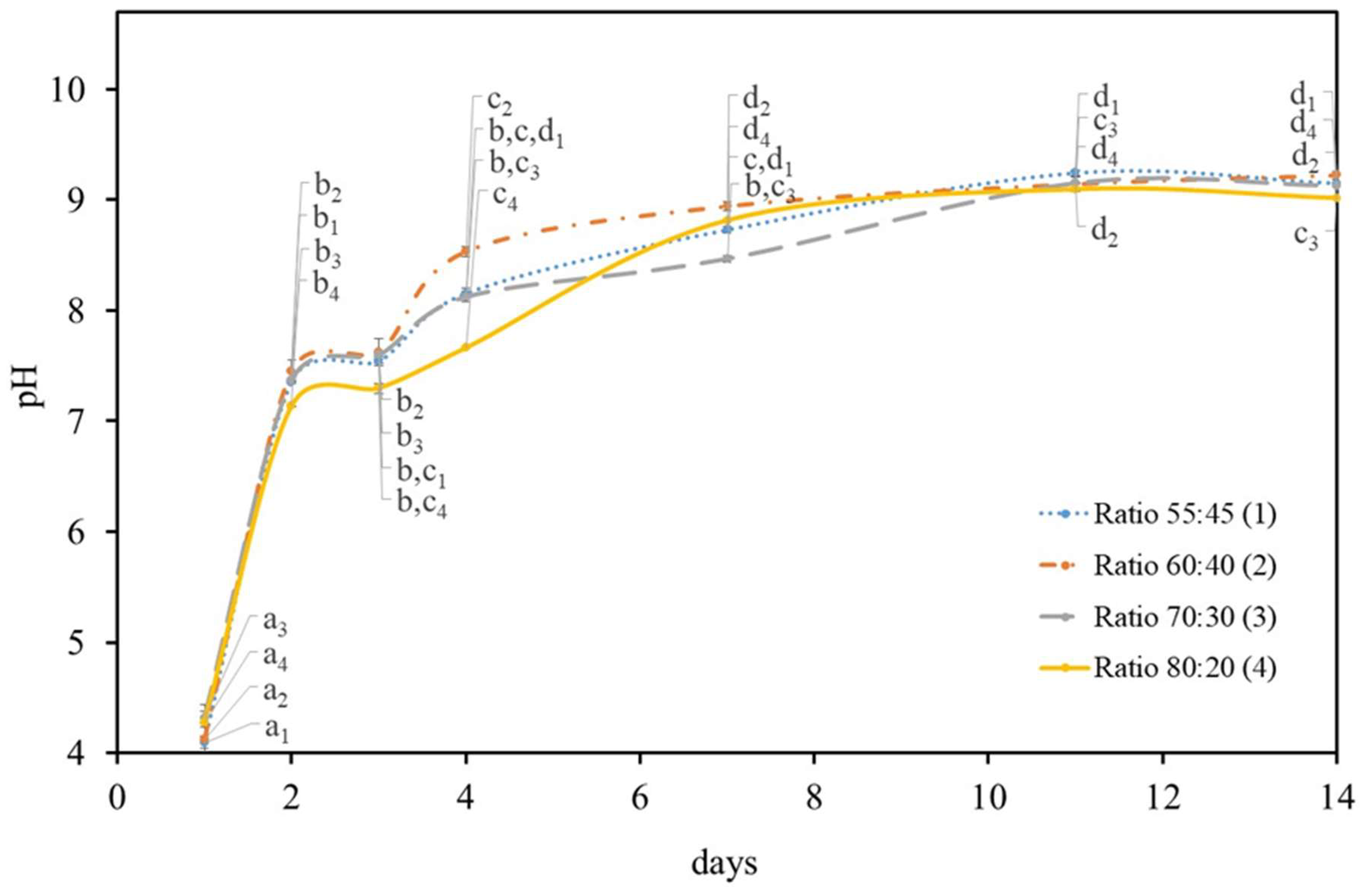
2.1. Growth Symbiotic Media Optimization
2.1.1. Carbon Source
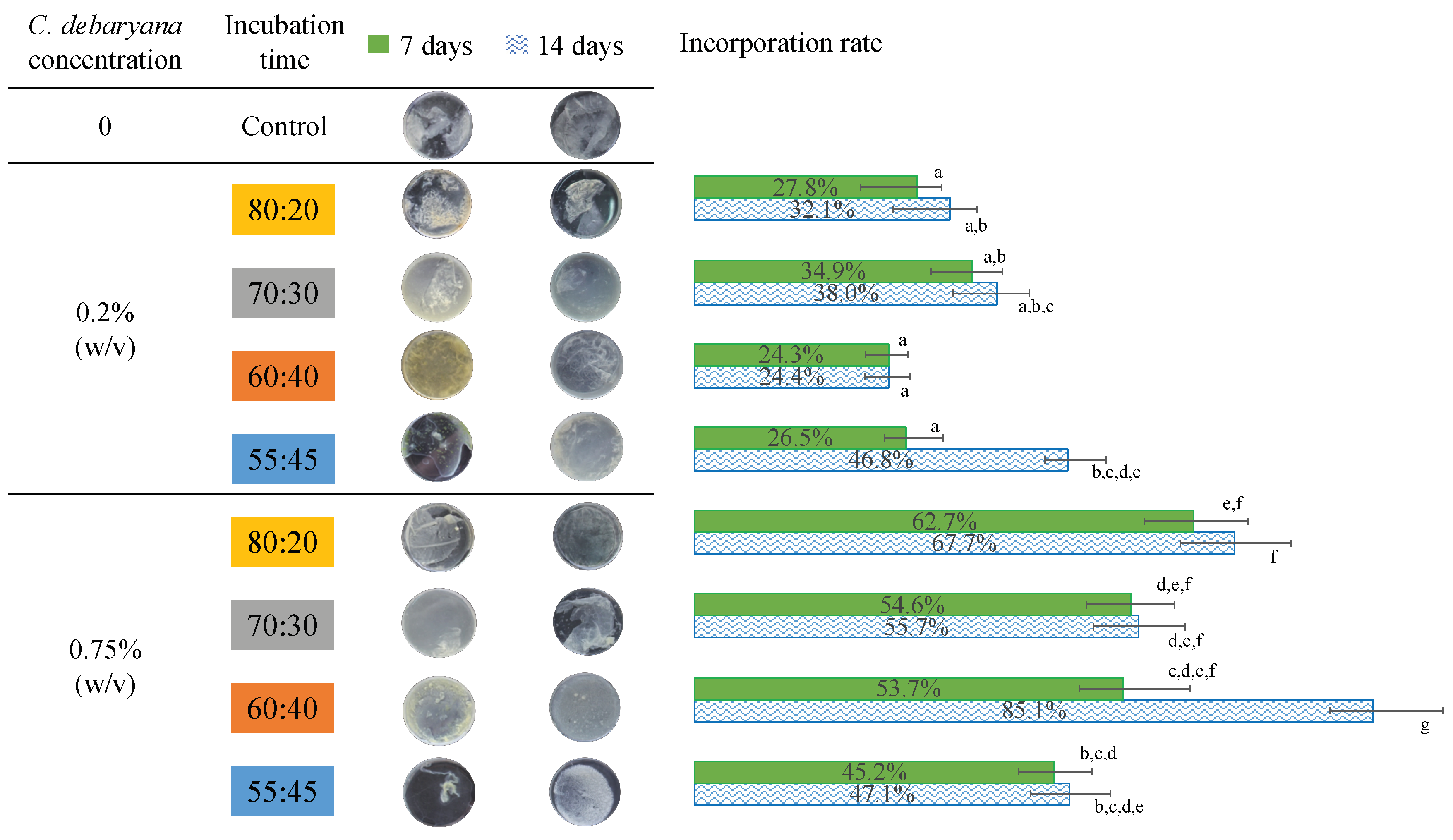
2.1.2. Symbiotic Media Constitution
2.2. Growth Temperature
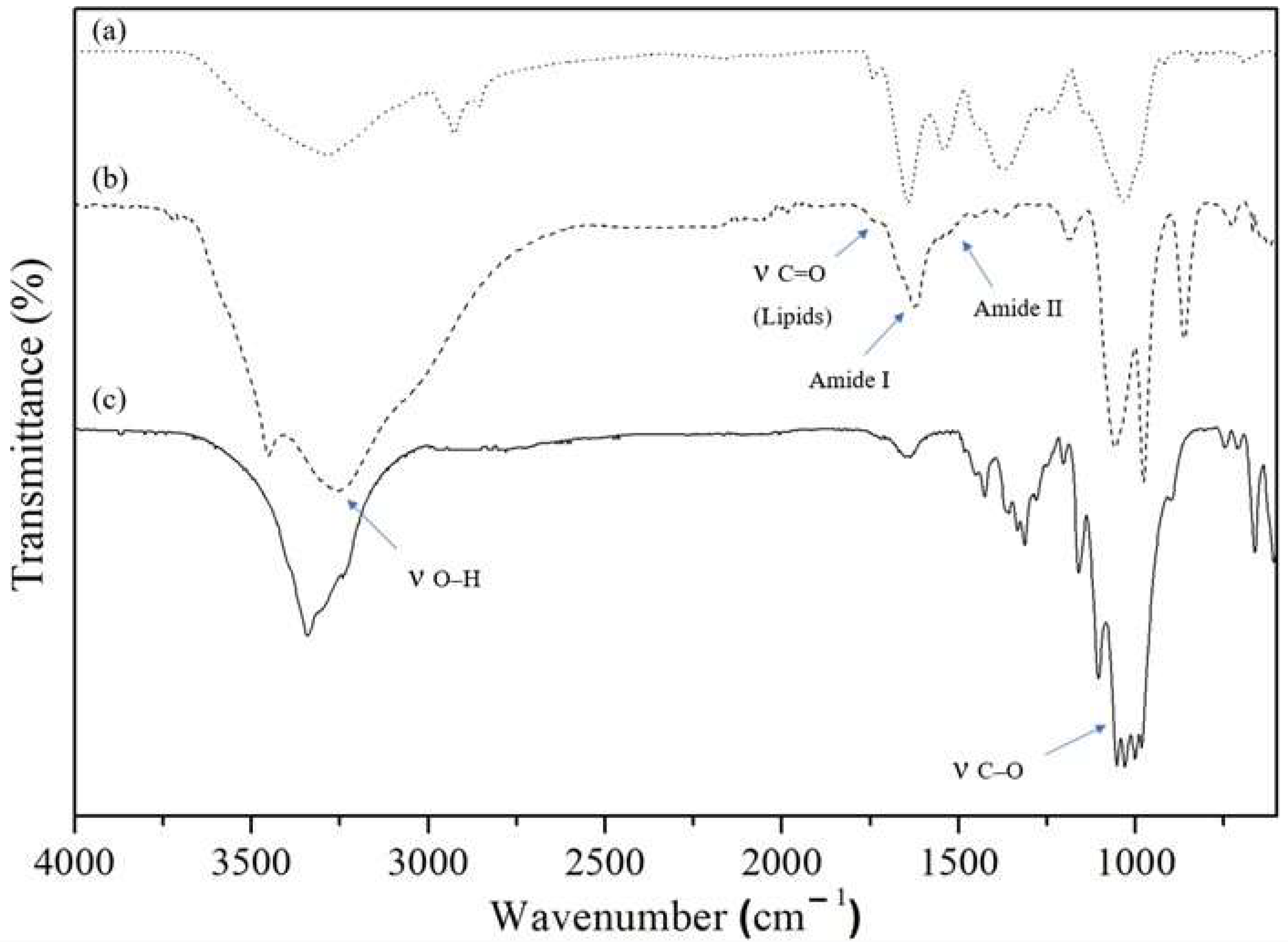
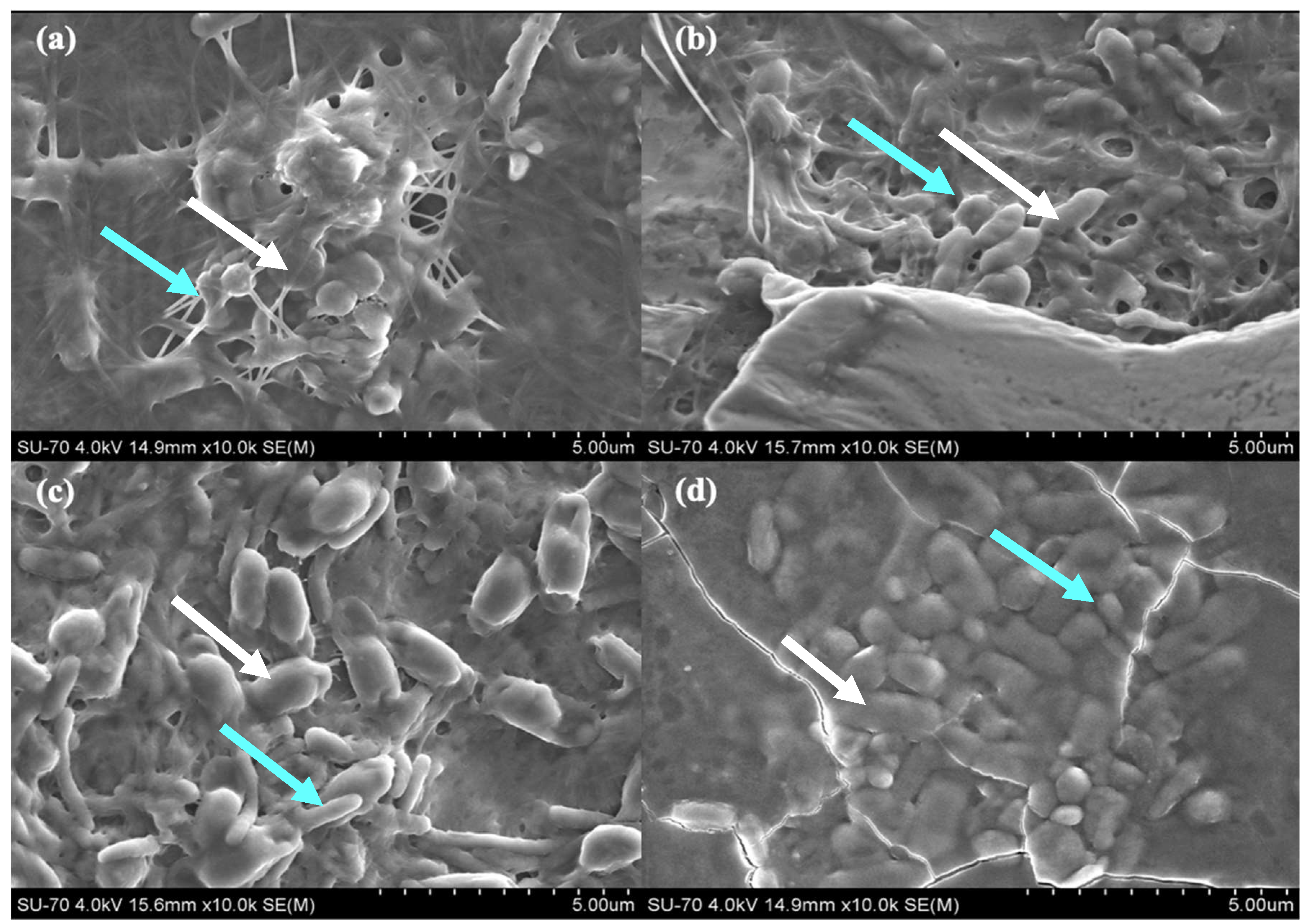
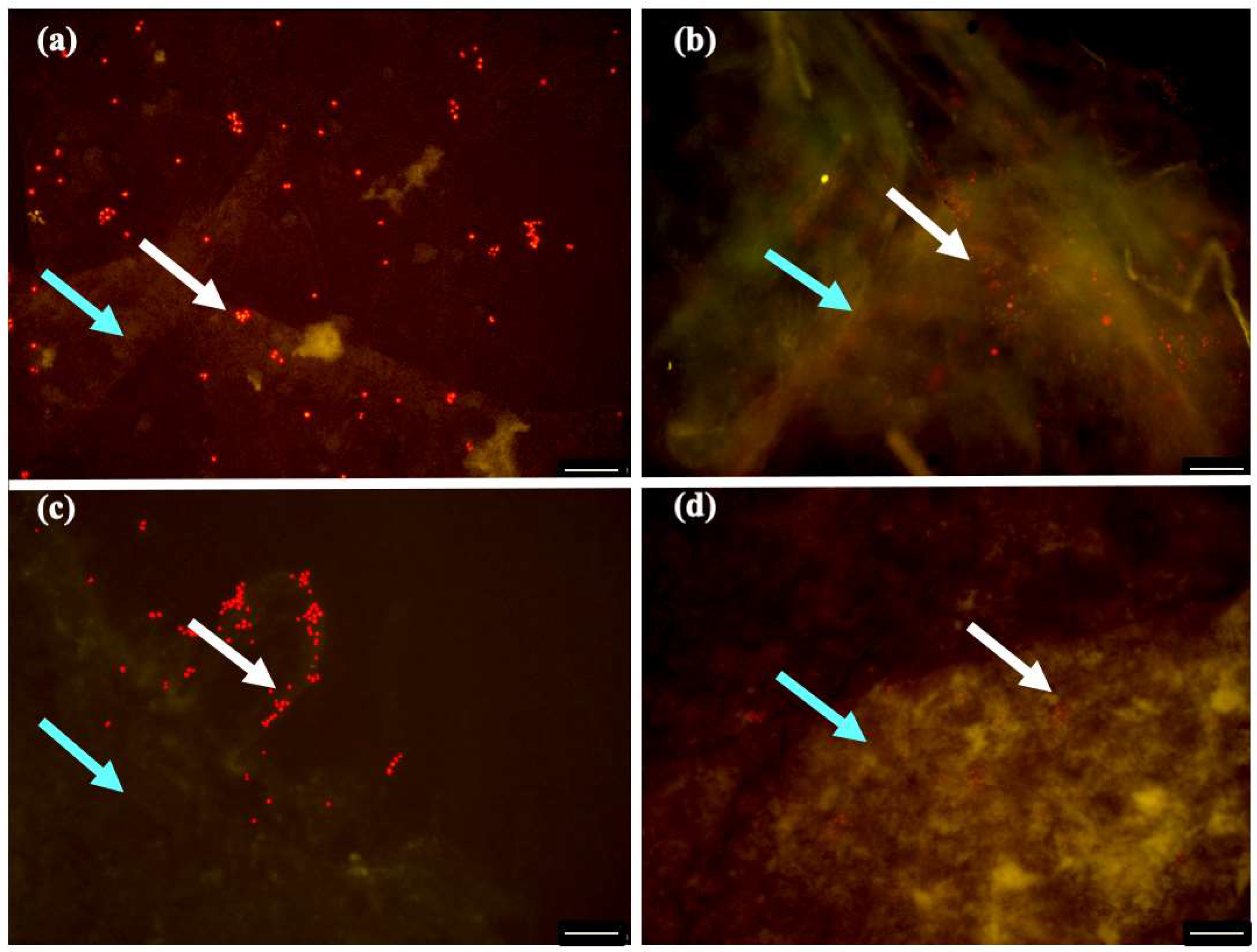
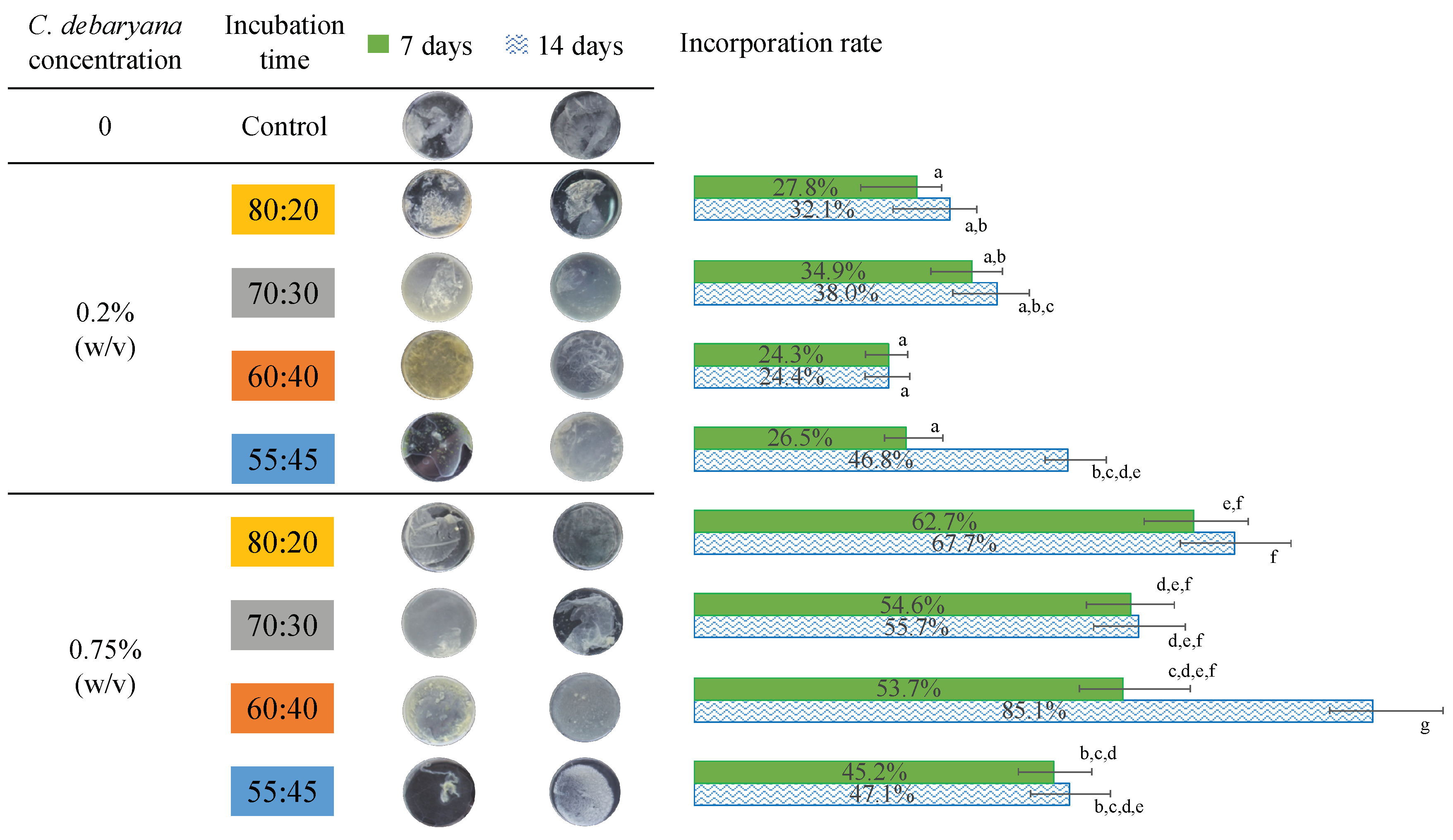
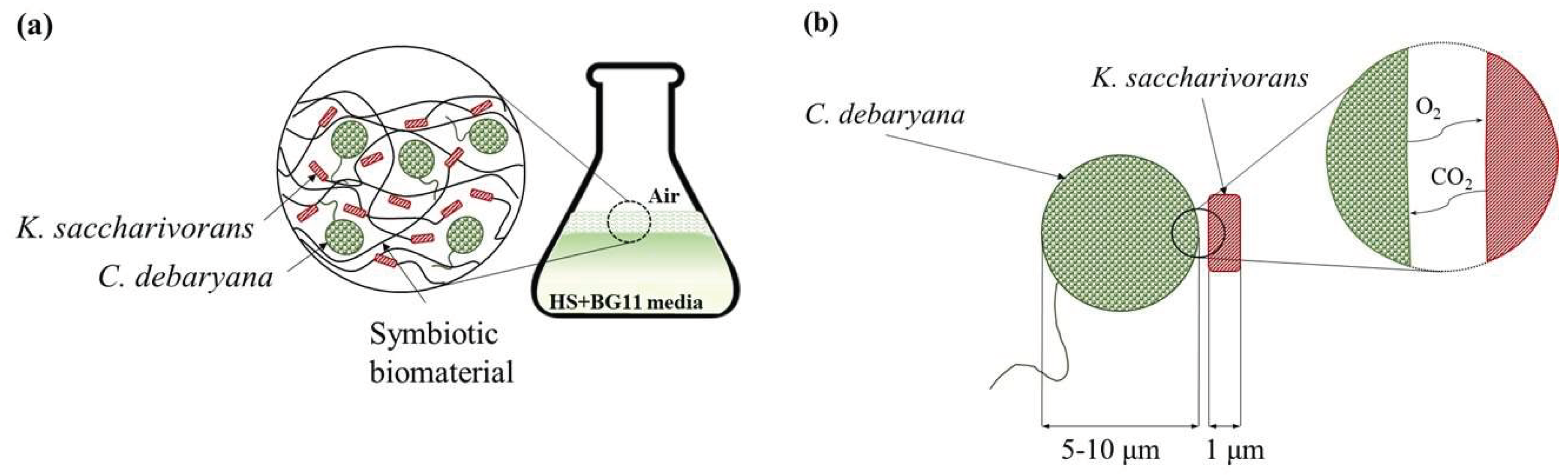
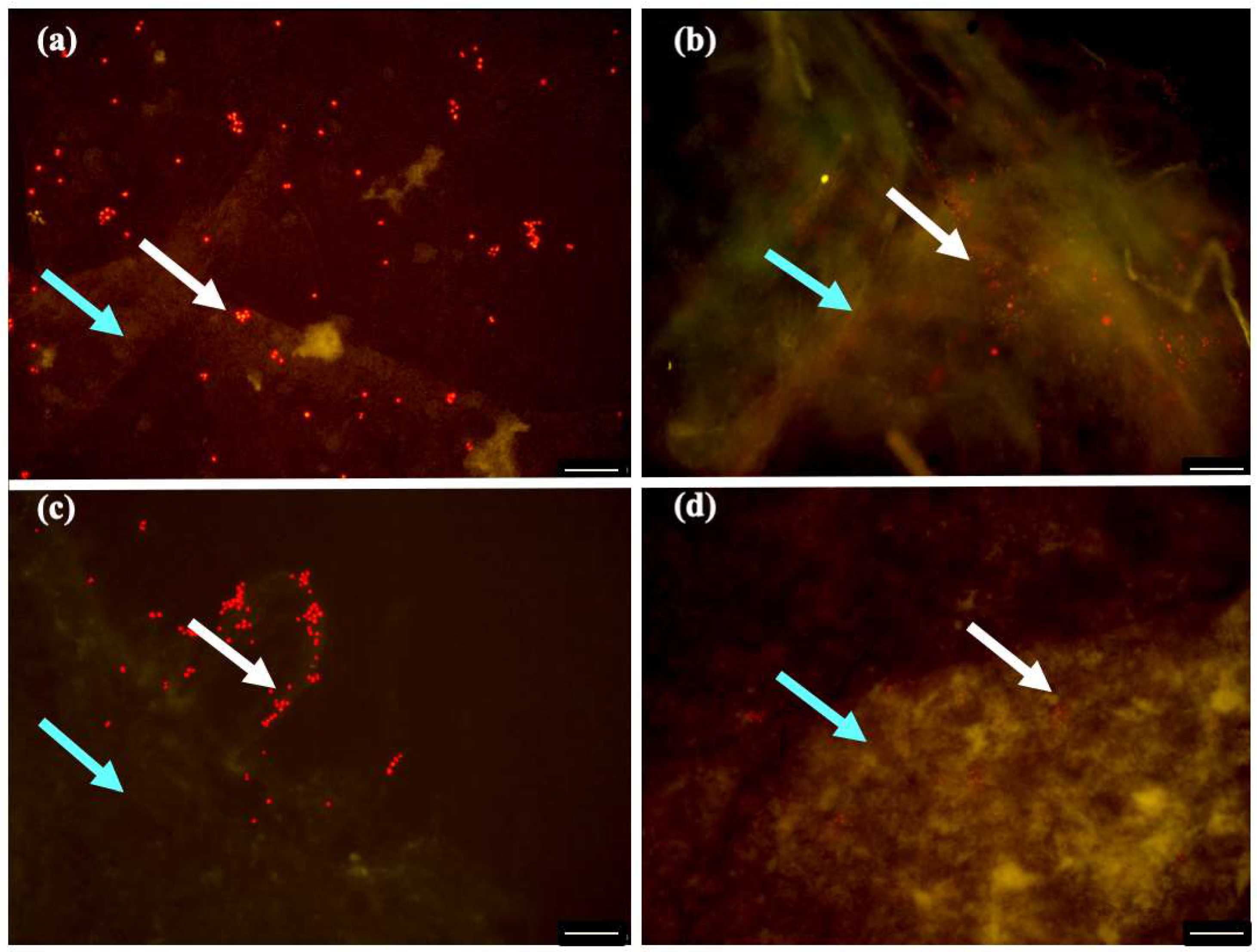
2.3. Biofilm Characterization
3. Materials and Methods
3.1. Microorganisms and Grown Medium Conditions
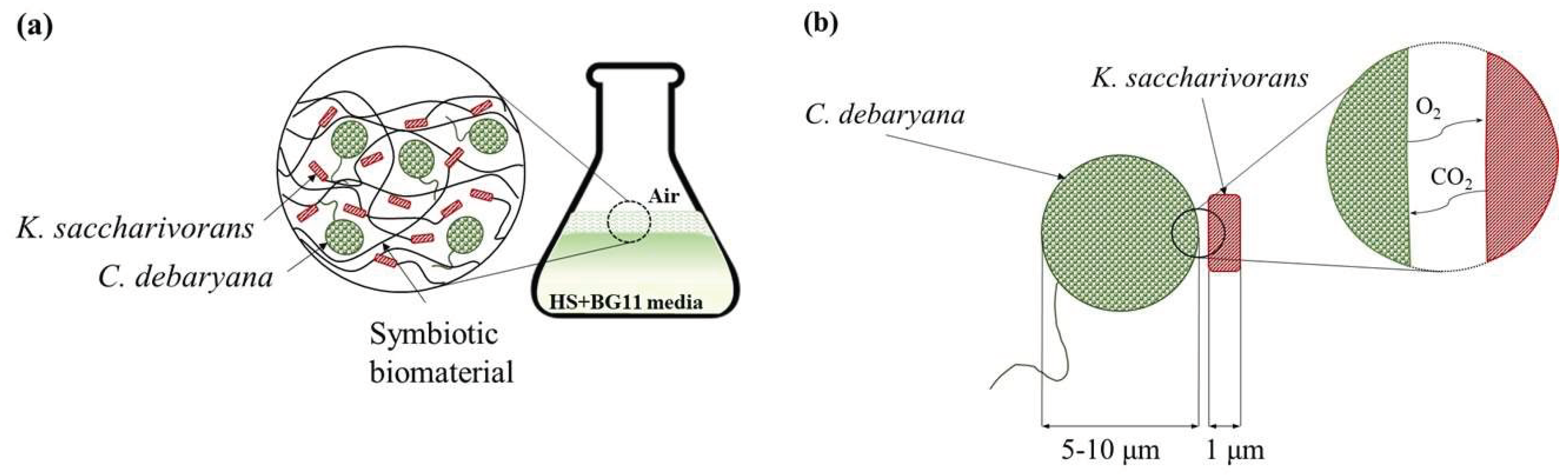
3.2. Symbiotic Process
3.3. High Performance Liquid Chromatography (HPLC)
3.4. Determination of Incorporation Rate
3.5. Attenuated Total Reflection Fourier Transform Infrared Spectroscopy (ATR-FTIR) Analysis
3.6. Scanning Electron Microscopy (SEM) Analysis
3.7. Fluorescence Microscopic Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ton, N.; Le, V. Application of immobilized yeast in bacterial cellulose to the repeated batch fermentation in wine-making. Int. Food Res. J. 2011, 18, 983–987. [Google Scholar]
- Picheth, G.F.; Pirich, C.L.; Sierakowski, M.R.; Woehl, M.A.; Sakakibara, C.N.; Souza, C.F.; Martin, A.A.; Silva, R.; Freitas, R.A. Bacterial cellulose in biomedical applications: A review. Int. J. Biol. Macromol. 2017, 104, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Sulaeva, I.; Henniges, U.; Rosenau, T.; Potthast, A. Bacterial cellulose as a material for wound treatment: Properties and modifications. A review. Biotechnol. Adv. 2017, 33, 1547–1571. [Google Scholar] [CrossRef]
- Tayeb, A.H.; Amini, E.; Ghasemi, S.; Tajvidi, M. Cellulose Nanomaterials-Binding Properties and Applications: A Review. Molecules 2018, 23, 2684. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, H.; Zheng, W.; Gong, N.; Chen, L.; Jiang, X.; Yang, G. Bacterial cellulose—hyaluronan nanocomposite biomaterials as wound dressings for severe skin injury repair. J. Mater. Chem. B 2015, 3, 3498–3507. [Google Scholar] [CrossRef]
- Dórame-Miranda, R.F.; Gámez-Meza, N.; Medina-Juárez, L.Á.; Ezquerra-Brauer, J.M.; Ovando-Martínez, M.; Lizardi-Mendoza, J. Bacterial cellulose production by Gluconacetobacter entanii using pecan nutshell as carbon source and its chemical functionalization. Carbohydr. Polym. 2019, 207, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Catchmark, J.M.; Vogler, E.A. Factors Impacting the Formation of Sphere-Like Bacterial Cellulose Particles and Their Biocompatibility for Human Osteoblast Growth. Bio. Macromol. 2013, 14, 3444–3452. [Google Scholar] [CrossRef] [PubMed]
- Czaja, W.; Krystynowicza, A.; Bieleckia, S.; Brown, R.M.Jr. Microbial cellulose—the natural power to heal wounds. Biomater. 2006, 27, 145–151. [Google Scholar] [CrossRef]
- Divakar, P.K.; Crespo, A.; Wedin, M.; Leavitt, S.D.; Hawksworth, D.L.; Myllys, L.; McCune, B. Evolution of complex symbiotic relationships in a morphologically derived family of lichen-forming fungi. New Phytol. 2015, 208, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Hom, E.F.; Murray, A.W. Niche Engineering Demonstrates a Latent Capacity for Fungal-Algal Mutualism. Science 2014, 345, 94–98. [Google Scholar] [CrossRef]
- Aschenbrenner, I.A.; Cernava, T.; Berg, G.; Grube, M. Understanding Microbial Multi-Species Symbioses. Front. Microbiol. 2016, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Grube, M.; Koberl, M.; Lackner, S.; Berg, C.; Berg, G. Host-parasite interaction and microbiome response: Effects of fungal infections on the bacterial community of the Alpine lichen Solorina crocea. FEMS Microbiol. Ecol. 2012, 82, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.T.; Cropsey, G.W.; Caporaso, J.G.; Knight, R.; Fierer, N. Bacterial Communities Associated with the Lichen Symbiosis. Appl. Environ. Microbiol. 2011, 77, 1309–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Tong, Y.W. The interactions between Chlorella vulgaris and algal symbiotic bacteria under photoautotrophic and photoheterotrophic conditions. J. Appl. Phycol. 2014, 26, 1483–1492. [Google Scholar] [CrossRef]
- Das, A.A.; Bovill, J.; Ayesh, M.; Stoyanov, S.D.; Paunov, V.N. Fabrication of living soft matter by symbiotic growth of unicellular microorganisms. J. Mater. Chem. B 2016, 4, 3685–3694. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Fan, Q.; Zhang, Z.; Mei, Y.; Wang, H. Effective in situ harvest of microalgae with bacterial cellulose produced by Gluconacetobacter xylinus. Algal Res. 2018, 35, 349–354. [Google Scholar] [CrossRef]
- Nélson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 5th ed.; W.H. Freeman and Company: New York, NY, USA, 2008; Chapter 19. [Google Scholar]
- Augimeri, R.V.; Varley, A.J.; Strap, J.L. Establishing a Role for Bacterial Cellulose in Environmental Interactions: Lessons Learned from Diverse Biofilm-Producing Proteobacteria. Front. Microbiol. 2015, 6, 1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullo, M.; La China, S.; Falcone, P.M.; Giudici, P. Biotechnological production of cellulose by acetic acid bacteria: Current state and perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 6885–6898. [Google Scholar] [CrossRef]
- Fang, L.; Catchmark, J.M. Characterization of cellulose and other exopolysaccharides produced from Gluconacetobacter strains. Carbohydr. Polym. 2015, 115, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Embuscado, M.E.; Marks, J.S.; BeMiller, J.N. Bacterial cellulose. I. Factors affecting the production of celulose by Acetobacter xylinum. Food Hydrocoll 1994, 8, 407–418. [Google Scholar] [CrossRef]
- Vandamme, E.J.; Baets, S.D.; Vanbaelen, A.; Joris, K.; Wulf, P.D. Improved production of bacterial cellulose and its application potential. Polym. Degrad. Stab. 1998, 59, 93–99. [Google Scholar] [CrossRef]
- Klemm, D.; Schumann, D.; Udhardt, U.; Marsch, S. Bacterial synthesized cellulose—artificial blood vessels for microsurgery. Prog. Polym. Sci. 2001, 26, 1561–1603. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, D.; Flanagan, B.; Dykes, G.; Gidley, M. Influence of different carbon sources on bacterial cellulose production by Gluconacetobacter xylinus strain ATCC 53524. J. Appl. Microbiol. 2009, 107, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Kornmann, H.; Duboc, P.; Marison, I.; von Stockar, U. Influence of nutritional factors on the nature, yield, and composition of exopolysaccharides produced by Gluconacetobacter xylinus I-2281. Appl. Environ. Microbiol. 2003, 69, 6091–6098. [Google Scholar] [CrossRef] [PubMed]
- Mamlouk, D.; Gullo, M. Acetic Acid Bacteria: Physiology and Carbon Sources Oxidation. Indian, J. Microbiol. 2013, 53, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Nieto-Peñalver, C.G.; Savino, M.J.; Bertini, E.V.; Sánchez, L.A.; Figueroa, L.I. Gluconic acid produced by Gluconacetobacter diazotrophicus Pal5 possesses antimicrobial properties. Res. Microbiol. 2012, 165, 549–558. [Google Scholar] [CrossRef]
- Keshk, S.; Sameshima, K. Influence of lignosulfonate on crystal structure and productivity of bacterial cellulose in a static culture. Enzyme Microb. Technol. 2006, 40, 4–8. [Google Scholar] [CrossRef]
- Rangaswamy, B.E.; Vanitha, K.P.; Hungund, B.S. Microbial cellulose production from bacteria isolated from rotten fruit. Int. J. Polym. Sci. 2015, 1–8. [Google Scholar] [CrossRef]
- Giordano, M.; Kansiz, M.; Heraud, P.; Beardall, J.; Wood, B.; McNaughton, D. Fourier transform infrared spectroscopy as a novel tool to investigate changes in intracellular macromolecular pools in the marine microalga Chaetoceros muellerii (bacillariophyceae). J. Phycol. 2001, 37, 271–279. [Google Scholar] [CrossRef]
- Rudie, A. Commercialization of cellulose nanofibril (CNF) and cellulose nanocrystal (CNC): Pathway and challenges. In Handbook of Nanocellulose and Cellulose Nanocomposites; Kargarzadeh, H., Ahmad, I., Thomas, S., Duiresne, A., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2017. [Google Scholar]
- Castro, C.; Zuluaga, R.; Álvarez, C.; Putaux, J.; Caro, G.; Rojas, O.J.; Mondragon, I.; Gañána, P. Bacterial cellulose produced by a new acid-resistant strain of Gluconacetobacter genus. Carbohydr. Polym. 2012, 89, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Horii, F. In Situ crystallization of bacterial cellulose I. Influences of polymeric additives, stirring and temperature on the formation celluloses Iα and Iβ as revealed by cross polarization/magic angle spinning (CP/MAS) 13C NMR spectroscopy. Cellulose 1994, 1, 57–66. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source | wt % | pH Final |
|---|---|---|
| Glucose | 1 | 3.3 ± 0.09 a |
| 2 | 2.9 ± 0.01 a | |
| 4 | 3.0 ± 0.01 a | |
| Fructose | 1 | 5.4 ± 0.16 b |
| 2 | 7.2 ± 0.02 c | |
| 4 | 7.1 ±0.19 c | |
| Sucrose | 1 | 8.4 ± 0.08 d |
| 2 | 8.3 ± 0.01 d | |
| 4 | 8.2 ± 0.10 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nóbrega, V.; Faria, M.; Quintana, A.; Kaufmann, M.; Ferreira, A.; Cordeiro, N. From a Basic Microalga and an Acetic Acid Bacterium Cellulose Producer to a Living Symbiotic Biofilm. Materials 2019, 12, 2275. https://doi.org/10.3390/ma12142275
Nóbrega V, Faria M, Quintana A, Kaufmann M, Ferreira A, Cordeiro N. From a Basic Microalga and an Acetic Acid Bacterium Cellulose Producer to a Living Symbiotic Biofilm. Materials. 2019; 12(14):2275. https://doi.org/10.3390/ma12142275
Chicago/Turabian StyleNóbrega, Vítor, Marisa Faria, Antera Quintana, Manfred Kaufmann, Artur Ferreira, and Nereida Cordeiro. 2019. "From a Basic Microalga and an Acetic Acid Bacterium Cellulose Producer to a Living Symbiotic Biofilm" Materials 12, no. 14: 2275. https://doi.org/10.3390/ma12142275
APA StyleNóbrega, V., Faria, M., Quintana, A., Kaufmann, M., Ferreira, A., & Cordeiro, N. (2019). From a Basic Microalga and an Acetic Acid Bacterium Cellulose Producer to a Living Symbiotic Biofilm. Materials, 12(14), 2275. https://doi.org/10.3390/ma12142275