A Facile Measurement for Monitoring Dragline Silk Dope Concentration in Nephila pilipes upon Spinning



Abstract
1. Introduction
2. Materials and Methods
2.1. Spider Silk Collection and Measurement of Mechanical Properties
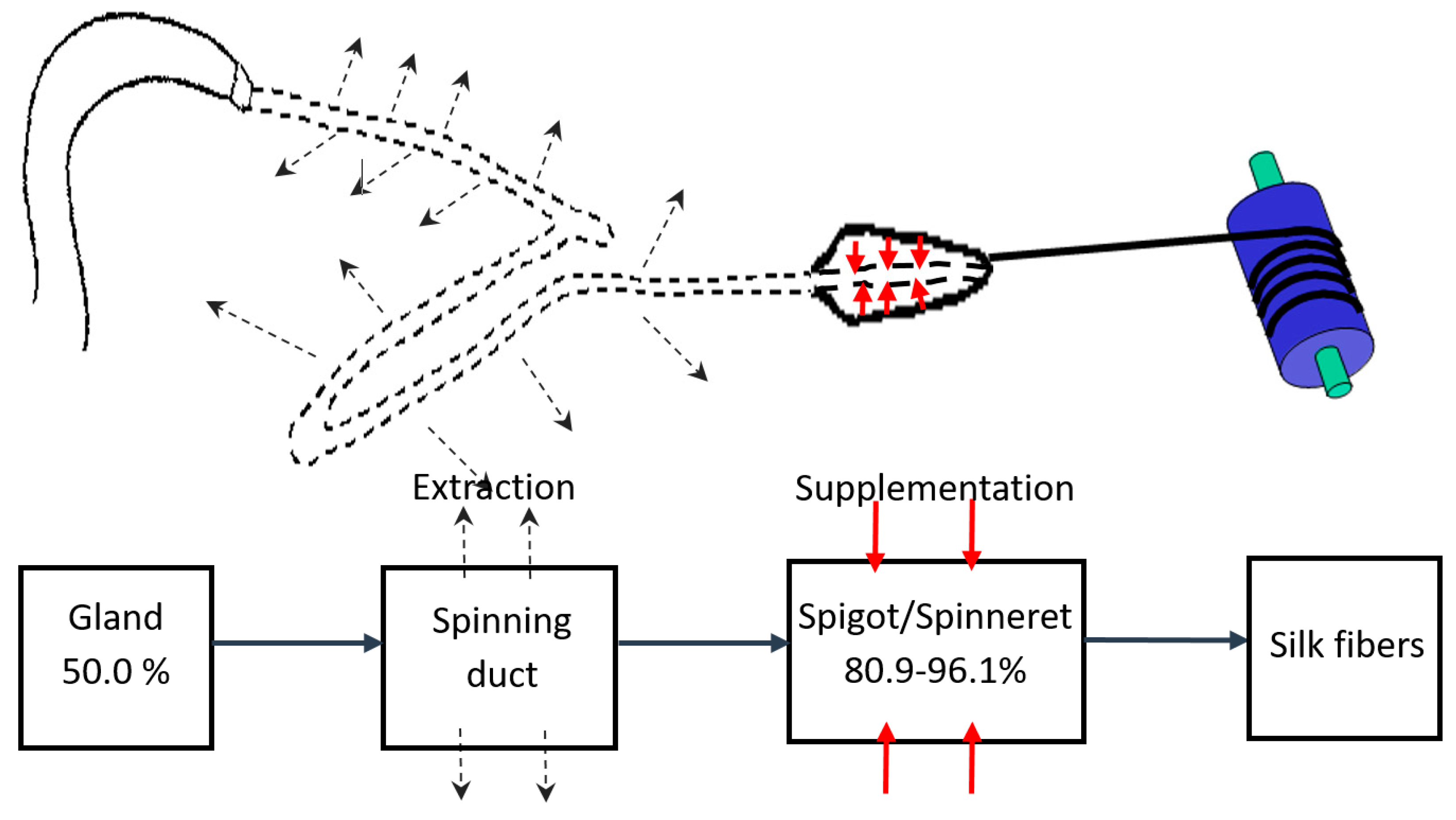
2.2. Measurement of Liquid Content of Silk Fibers Exiting the Spinneret
2.3. Measurement of Gland Silk Liquid Content
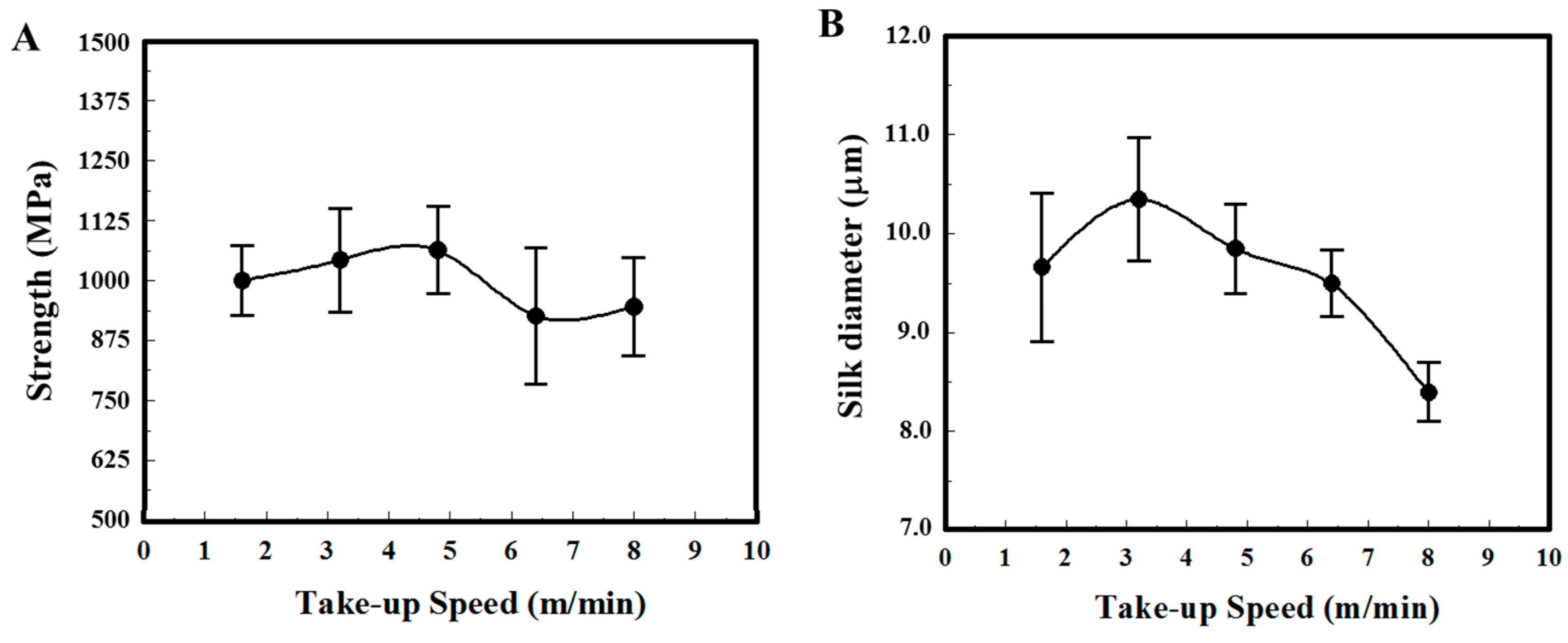
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knight, D.P.; Vollrath, F. Spinning an elastic ribbon of spider silk. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Lazaris, A.; Arcidiacono, S.; Huang, Y.; Zhou, J.-F.; Duguay, F.; Chretien, N.; Welsh, E.A.; Soares, J.W.; Karatzas, C.N. Spider silk fibers spun from soluble recombinant silk produced in mammalian cells. Science 2002, 295, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Liivak, O.; Blye, A.; Shah, N.; Jelinski, L.W. A microfabricated wet-spinning apparatus to spin fibers of silk proteins. Structure-property correlations. Macromolecules 1998, 31, 2947–2951. [Google Scholar]
- Seidel, A.; Liivak, O.; Jelinski, L.W. Artificial spinning of spider silk. Macromolecules 1998, 31, 6733–6736. [Google Scholar] [CrossRef]
- Shao, Z.; Vollrath, F. Surprising strength of silkworm silk. Nature 2002, 418, 741. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, F.; Knight, D.P. Structure and function of the silk production pathway in the spider Nephila edulis. Int. J. Biol. Macromol. 1999, 24, 243–249. [Google Scholar] [CrossRef]
- Vollrath, F. Strength and structure of spiders’ silks. J. Biotechnol. 2000, 74, 67–83. [Google Scholar] [CrossRef]
- Vollrath, F.; Madsen, B.; Shao, Z. The effect of spinning conditions on the mechanics of a spider’s dragline silk. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, F.; Knight, D.P. Liquid crystalline spinning of spider silk. Nature 2001, 410, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Masuda, H.; Zhao, C.; Asakura, T. Artificial spinning and characterization of silk fiber from bombxy mori silk fibroin in hexafluoroacetone hydrate. Macromolecules 2002, 35, 6–9. [Google Scholar] [CrossRef]
- Xia, X.-X.; Qian, Z.-G.; Ki, C.S.; Park, Y.H.; Kaplan, D.L.; Lee, S.Y. Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber. Proc. Natl. Acad. Sci. USA 2010, 107, 14059–14063. [Google Scholar] [CrossRef] [PubMed]
- Hauptmann, V.; Weichert, N.; Menzel, M.; Knoch, D.; Paege, N.; Scheller, J.; Spohn, U.; Conrad, U.; Gils, M. Native-sized spider silk proteins synthesized in planta via intein-based multimerization. Transgenic Res. 2013, 22, 369–377. [Google Scholar]
- Andersson, M.; Jia, Q.; Abella, A.; Lee, X.-Y.; Landreh, M.; Purhonen, P.; Hebert, H.; Tenje, M.; Robinson, C.V.; Meng, Q.; et al. Biomimetic spinning of artificial spider silk from a chimeric minispidroin. Nat. Chem. Biol. 2017, 13, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Tokareva, O.; Javobsen, M.; Buehler, M.; Wong, J.; Kaplan, D.L. Structure-function-property-design interplay in biopolymers: Spider silk. Acta Biomater. 2014, 10, 1612–1626. [Google Scholar] [CrossRef] [PubMed]
- Scheibel, T. Spider silks: Recombinant synthesis, assembly, spinning, and engineering of synthetic proteins. Microb. Cell Fact. 2004, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Rammensee, S.; Slotta, U.; Scheibel, T.; Bauch, A.R. Assembly mechanism of recombinant spider silk proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 6590–6595. [Google Scholar]
- Heim, M.; Keerl, D.; Scheibel, T. Spider silk: From soluble protein to extraordinary fiber. Angew. Chem. Int. Ed. 2009, 48, 3584–3596. [Google Scholar] [CrossRef] [PubMed]
- Koeppel, A.; Holland, C. Progress and trends in artificial silk spinning: A systematic review. ACS Biomater. Sci. Eng. 2017, 3, 226–237. [Google Scholar] [CrossRef]
- Knight, D.P.; Vollrath, F. Changes in element composition along the spinning duct in a Nephila spider. Naturwissenschaften 2001, 88, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Johansson, J.; Rising, A. Silk spinning in silkworms and spiders. Int. J. Mol. Sci. 2016, 17, 1290. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.L.; Peakall, D.B. Changes in fine structure during silk protein production in the ampullate gland of the spider Araneus sericatus. J. Cell Biol. 1969, 42, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Kovoor, J.; Zylberberg, L. Morphology and ultrastructure of the ampullate gland duct in Araenus diadematus Clerck (Arachnida, Araneidae). Z. Zellforsch. Mikrosk. Anat. 1972, 128, 188–211. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S. The structure of the dragline control valves in the garden spider. J. Cell Sci. 1962, 103, 549–555. [Google Scholar]
- Witt, P.N.; Reed, C.F.; Peakall, D.B. A Spider’s Web. Problems in Regulatory Biology; Springer: New York, NY, USA, 1968. [Google Scholar]
- Tillinghast, E.; Chase, S.; Townley, M. Water extraction by the major ampullate duct during silk formation in the spider. J. Insect Physiol. 1984, 30, 591–596. [Google Scholar] [CrossRef]
- Davies, G.J.; Knight, D.P.; Vollrath, F. Structure and function of the major ampullate spinning duct of the golden orb weaver, Nephila edulis. Tissue Cell 2013, 45, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.P.; Knight, M.M.; Vollrath, F. Beta transition and stress-induced phase separation in the spinning of spider dragline silk. Int. J. Biol. Macromol. 2000, 27, 205–210. [Google Scholar] [CrossRef]
- Chen, X.; Knight, D.P.; Vollrath, F. Rheological characterization of Nephila spidroin solution. Biomacromolecules 2002, 3, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Viney, C.; Huber, A.E.; Dunaway, D.L.; Kerkam, K.; Case, S.T. Optical characterization of silk secretions and fibers. In Silk Polymers: Materials Sciences and Biotechnology; Adams, W.W., Kaplan, D., Farmer, B., Viney, C., Eds.; ACS: Washington, DC, USA, 1994; pp. 120–136. [Google Scholar]
- Bird, R.B.; Stewart, W.E.; Lightfoot, E.N. Transport Phenomena, 2nd Wiley international ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2002; 895p. [Google Scholar]
- Kojic, N.; Bico, J.; Clasen, C.; McKinley, G.H. Ex vivo rheology of spider silk. J. Exp. Biol. 2006, 209, 4355–4362. [Google Scholar] [CrossRef] [PubMed]
- Blamires, S.J.; Wu, C.-C.; Wu, C.-L.; Sheu, H.-S.; Tso, I.-M. Uncovering spider silk nanocrystalline variations that facilitate wind-induced mechanical property changes. Biomacromolecules 2013, 14, 3484–3490. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, T.H.; Machado Fde, A.; Kaneto, G.E.; Japyassú, H.F. Resting metabolic rates of two orbweb spiders: A first approach to evolutionary success of ecribellate spiders. J. Insect Physiol. 2011, 57, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F. Metabolic rates of spiders. Comp. Biochem. Physiol. 1970, 33, 51–72. [Google Scholar] [CrossRef]
- World_Spider_Catalog. World Spider Catalog; Natural History Museum Bern: Bern, Switzerland, 2017. [Google Scholar]
- Blamires, S.J.; Wu, C.L.; Blackledge, T.A.; Tso, I.-M. Post-secretion processing influences spider silk performance. J. R. Soc. Interface 2012, 9, 2479–2487. [Google Scholar] [CrossRef] [PubMed]
- Tso, I.M.; Wu, H.C.; Hwang, I.R. Giant wood spider Nephila pilipes alters silk protein in response to prey variation. J. Exp. Biol. 2005, 208, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Hijirida, D.H.; Do, K.G.; Michal, C.; Wong, S.; Zax, D.; Jelinski, L.W. 13C NMR of Nephila clavipes major ampullate silk gland. Biophys. J. 1996, 71, 3442–3447. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Spider | No. 1 | No. 2 | No. 3 | No. 4 | No. 5 | |
|---|---|---|---|---|---|---|
| Item | ||||||
| Weight of spider (mg) | 3041.0 | 2850.6 | 3013.6 | 2698.1 | 2873.1 | |
| Weight loss during silking (mg) | 12.0 | 15.5 | 44.8 | 33.1 | 12.6 | |
| Metabolic rate (mg/min) | 0.3 | 0.3 | 0.4 | 0.5 | 0.3 | |
| Silking time (min) | 29.0 | 25.0 | 31.0 | 23.0 | 30.0 | |
| Weight of dried silk (mg) | 0.5 | 0.7 | 1.3 | 1.2 | 0.6 | |
| Silk conc. exiting spinneret (wt%) | 19.1 | 7.4 | 3.9 | 5.4 | 12.0 | |
| Water conc. exiting spinneret (wt%) | 80.9 | 92.6 | 96.1 | 94.6 | 88.0 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.-C.; Wu, S.-R.; Yang, T.C.-K.; Yang, J.-C. A Facile Measurement for Monitoring Dragline Silk Dope Concentration in Nephila pilipes upon Spinning. Materials 2018, 11, 1951. https://doi.org/10.3390/ma11101951
Wu H-C, Wu S-R, Yang TC-K, Yang J-C. A Facile Measurement for Monitoring Dragline Silk Dope Concentration in Nephila pilipes upon Spinning. Materials. 2018; 11(10):1951. https://doi.org/10.3390/ma11101951
Chicago/Turabian StyleWu, Hsuan-Chen, Shang-Ru Wu, Thomas Chung-Kuang Yang, and Jen-Chang Yang. 2018. "A Facile Measurement for Monitoring Dragline Silk Dope Concentration in Nephila pilipes upon Spinning" Materials 11, no. 10: 1951. https://doi.org/10.3390/ma11101951
APA StyleWu, H.-C., Wu, S.-R., Yang, T. C.-K., & Yang, J.-C. (2018). A Facile Measurement for Monitoring Dragline Silk Dope Concentration in Nephila pilipes upon Spinning. Materials, 11(10), 1951. https://doi.org/10.3390/ma11101951