
Optimization of Pretreatment Conditions and Enzymatic Hydrolysis of Corn Cobs for Production of Microbial Lipids by Trichosporon oleaginosus

 ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pretreatment of Corn Cobs
2.2. Enzymatic Digestibility of Corn Cobs
2.3. Analysis of Sugar Concentration
2.4. Effect of Substrate and Enzyme Loading on Sugar Yield
2.5. Cultivation of T. oleaginosus
2.6. Lignocellulose Biomass Analysis
2.7. Lipid Analysis
2.8. Fatty Acid Methyl Esters (FAMEs) Analysis
3. Results
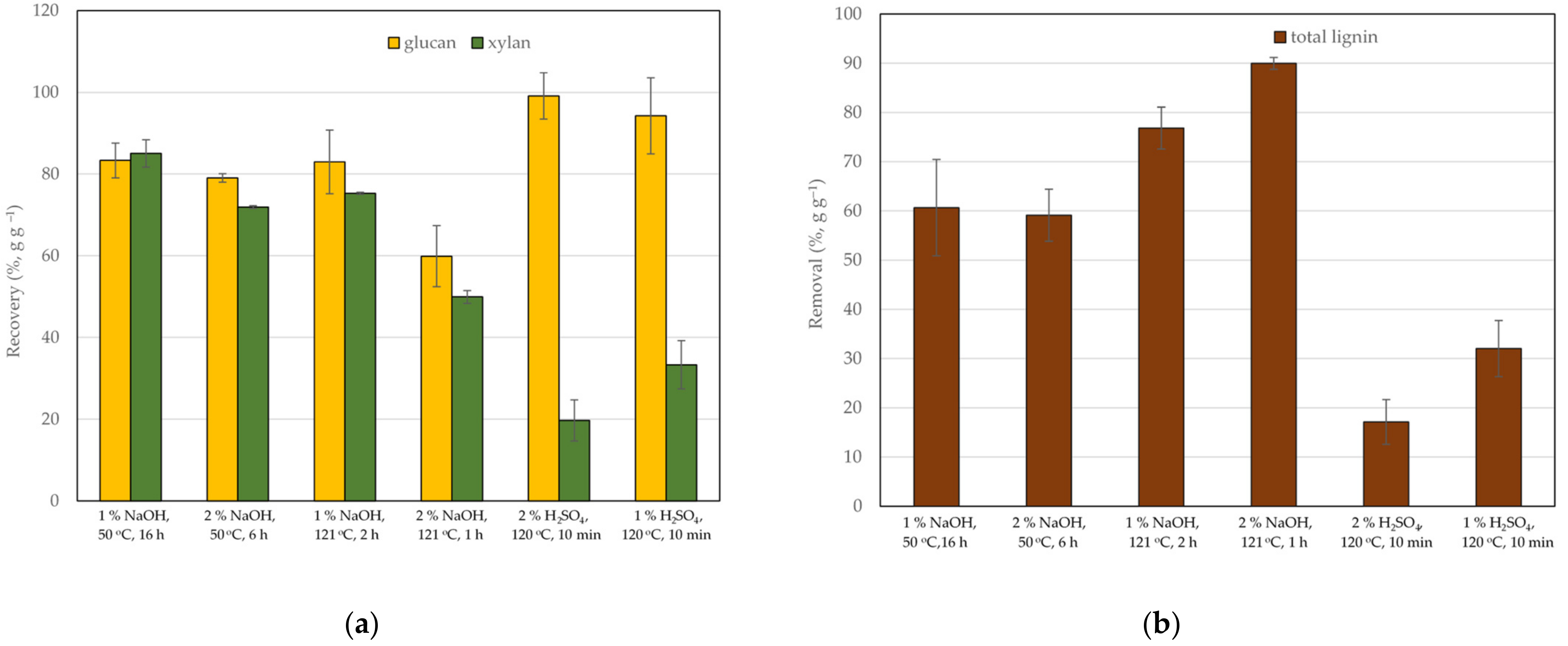
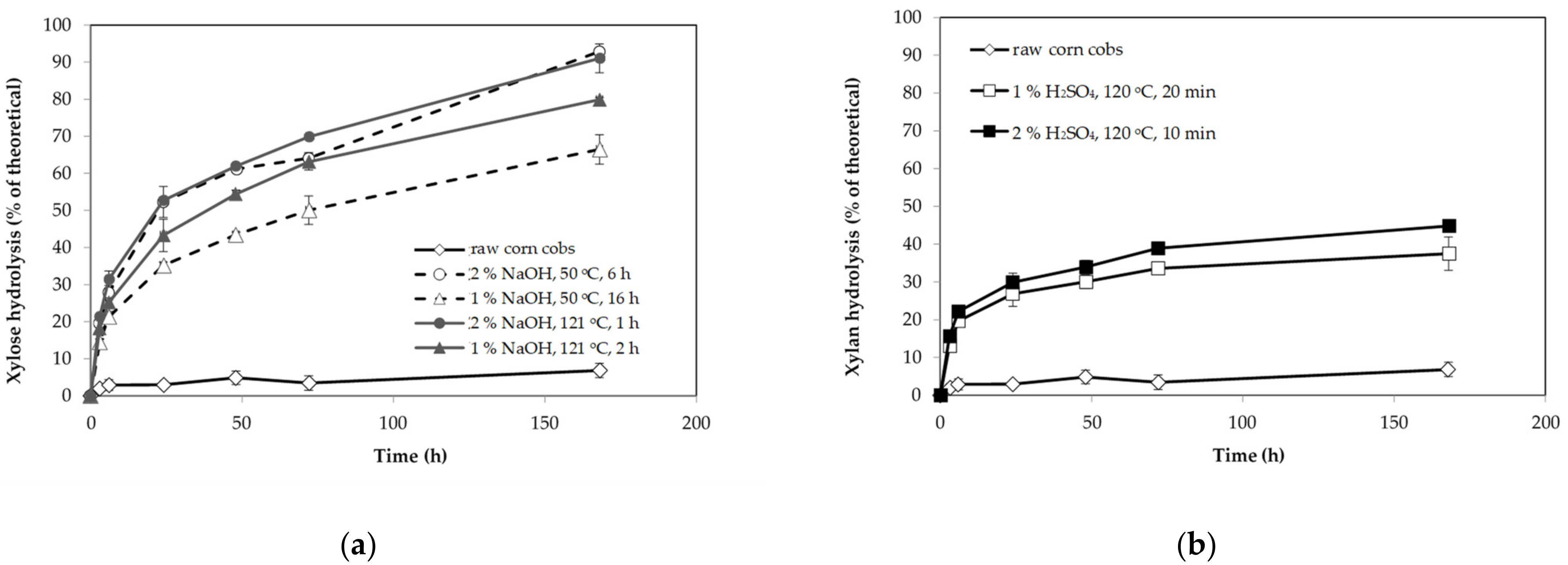
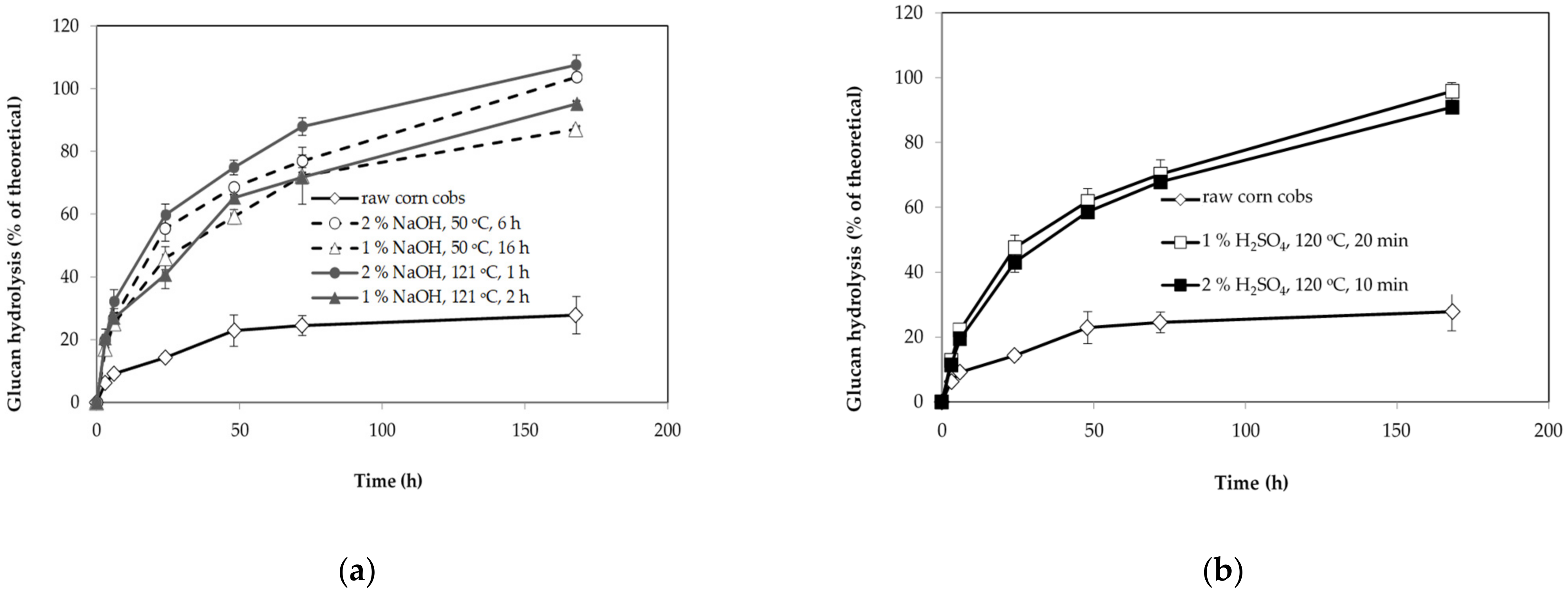
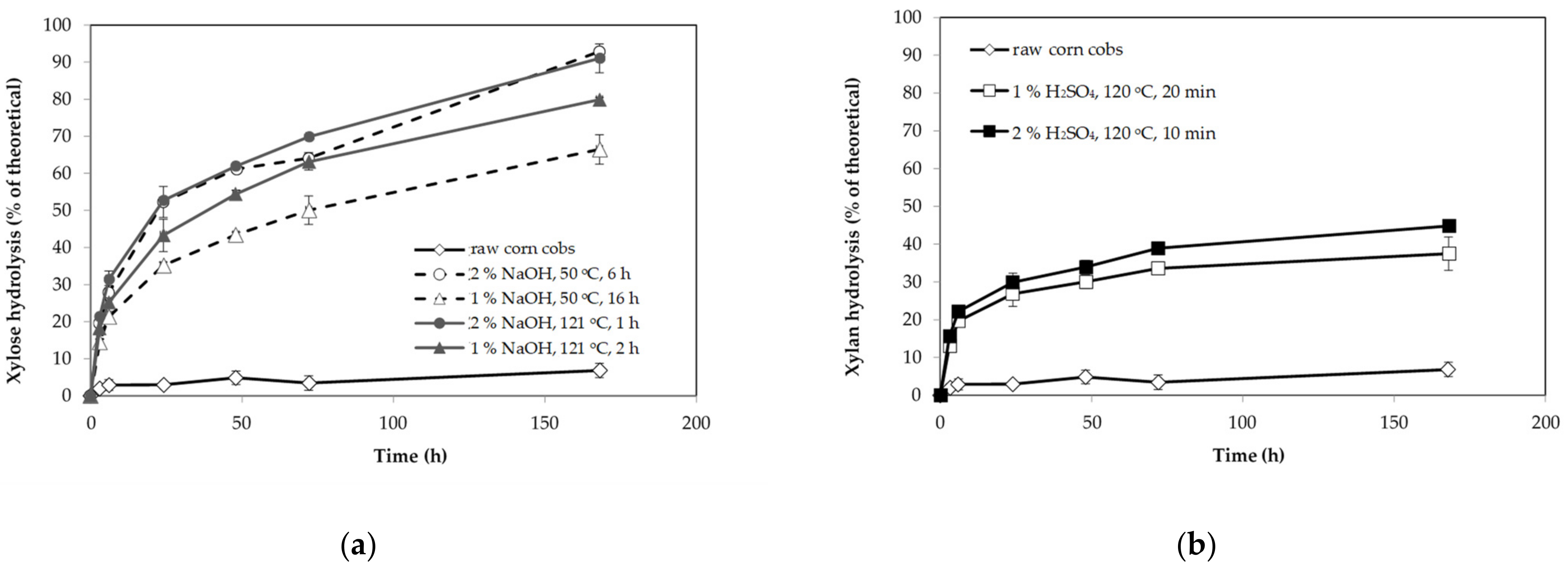
3.1. Composition and Enzyme Digestibility of Untreated and Pretreated Corn Cobs
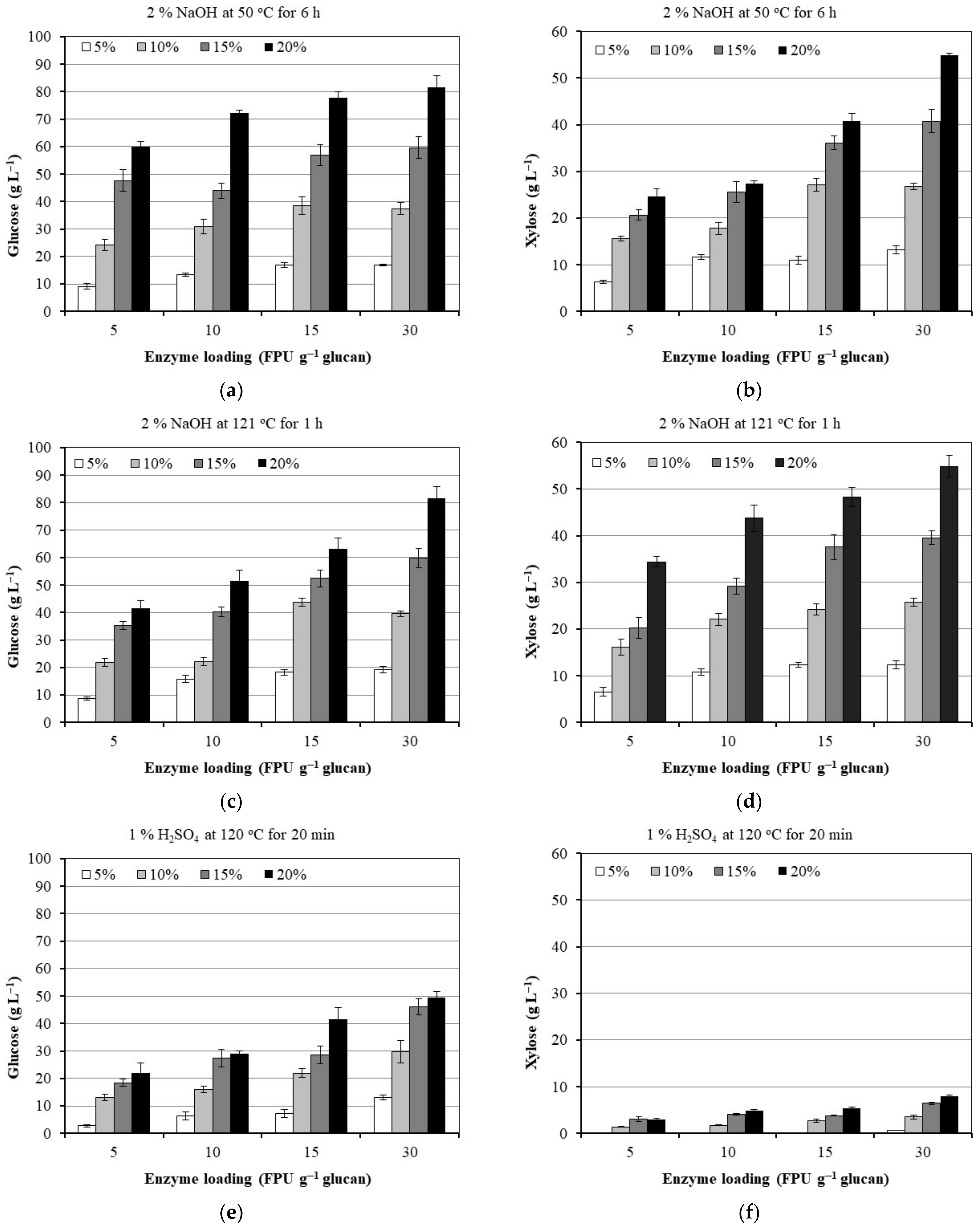
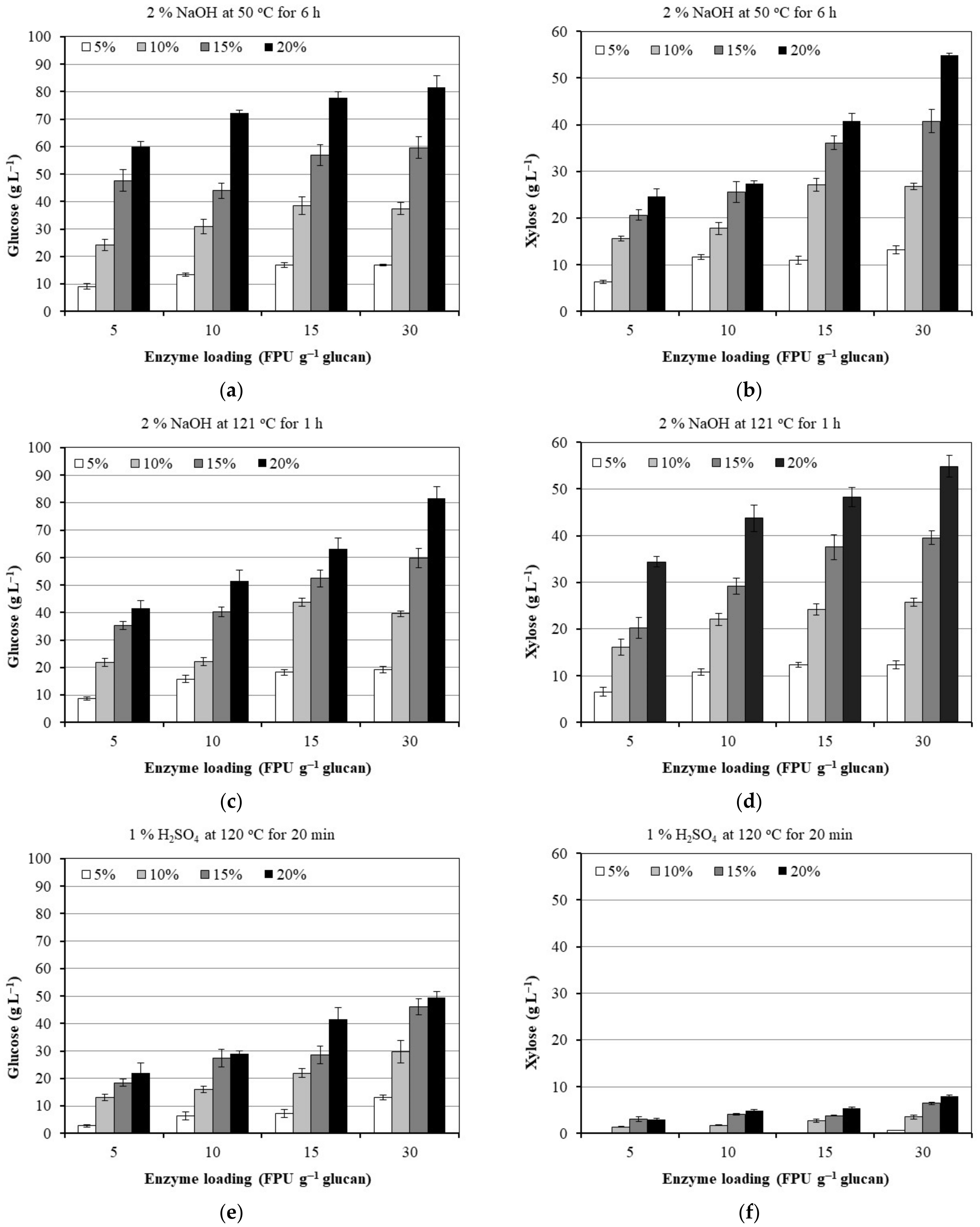
3.2. Effect of Enzyme and Substrate Loading
3.3. Cultivation of T. oleaginosus on Pretreated Lignocellulosic Biomasses and Filtrates of Lignocellulosic Slurries
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO Vegetable Oil Price Index Hits 13-Year High. Available online: https://www.ofimagazine.com/news/fao-vegetable-oil-price-index-hits-13-year-high (accessed on 25 March 2022).
- Rapeseed Oil-Monthly Price-Commodity Prices-Price Charts, Data, and News-IndexMundi. Available online: https://www.indexmundi.com/commodities/?commodity=rapeseed-oil&months=120 (accessed on 29 March 2022).
- Palm oil-Monthly Price-Commodity Prices-Price Charts, Data, and News-IndexMundi. Available online: https://www.indexmundi.com/commodities/?commodity=palm-oil&months=240 (accessed on 29 March 2022).
- Ravindranath, N.H.; Manuvie, R.; Fargione, J.; Canadell, J.G.; Berndes, G. Greenhouse Gas Implications of Land Use and Land Conversion to Biofuel Crops; Cornell University: Ithaca, NY, USA, 2009; pp. 111–125. [Google Scholar]
- Ivančić Šantek, M.; Miškulin, E.; Petrović, M.; Beluhan, S.; Šantek, B. Effect of carbon and nitrogen source concentrations on the growth and lipid accumulation of yeast Trichosporon oleaginosus in continuous and batch culture. J. Chem. Technol. Biotechnol. 2017, 92, 1620–1629. [Google Scholar] [CrossRef]
- Masri, M.A.; Garbe, D.; Mehlmer, N.; Brück, T.B. A sustainable, high-performance process for the economic production of waste-free microbial oils that can replace plant-based equivalents. Energy Environ. Sci. 2019, 12, 2717–2732. [Google Scholar] [CrossRef]
- Chen, M.; Tang, H.; Ma, H.; Holland, T.C.; Ng, K.Y.S.; Salley, S.O. Bioresource Technology Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresour. Technol. 2011, 102, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Zhu, Z.; Shen, H.; Lin, X.; Jin, X.; Jiao, X.; Zhao, Z.K. Biotechnology for Biofuels Systems analysis of phosphate-limitation-induced lipid accumulation by the oleaginous yeast Rhodosporidium toruloides. Biotechnol. Biofuels 2018, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafari, M.; Rashidi, B.; Haznedaroglu, B.Z. Effects of macro and micronutrients on neutral lipid accumulation in oleaginous microalgae. Biofuels 2018, 9, 147–156. [Google Scholar] [CrossRef]
- Sabu, S.; Sarojini, I.; Singh, B.; Joseph, V. Improved lipid production in oleaginous brackish diatom Navicula phyllepta MACC8 using two-stage cultivation approach. 3 Biotech 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Bellou, S.; Moustogianni, A.; Makri, A.; Aggelis, G. Lipids Containing Polyunsaturated Fatty Acids Synthesized by Zygomycetes Grown on Glycerol. Appl. Biochem. Biotechnol. 2012, 166, 146–158. [Google Scholar] [CrossRef]
- Valdés, G.; Mendonça, R.T.; Aggelis, G. Lignocellulosic biomass as a substrate for oleaginous microorganisms: A review. Appl. Sci. 2020, 10, 7698. [Google Scholar] [CrossRef]
- Vasconcelos, B.; Teixeira, J.C.; Dragon, G.; Teixeira, J.A. Oleaginous yeasts for sustainable lipid production—from biodiesel to surf boards, a wide range of “green” applications. Appl. Microbiol. Biotechnol. 2019, 103, 3651–3667. [Google Scholar] [CrossRef] [Green Version]
- Grubišić, M.; Mihajlovski, K.; Gruičić, A.M.; Beluhan, S.; Šantek, B.; Ivančić Šantek, M. Strategies for Improvement of Lipid Production by Yeast Trichosporon oleaginosus from Lignocellulosic Biomass. J. Fungi 2021, 7, 934. [Google Scholar] [CrossRef]
- Bao, R.; Wu, X.; Liu, S. Efficient Conversion of Fructose-Based Biomass into Lipids with Trichosporon fermentans Under Phosphate-Limited Conditions. Appl. Biochem. Biotechnol. 2018, 184, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Awad, D.; Bohnen, F.; Mehlmer, N.; Brueck, T. Multi-Factorial-Guided Media Optimization for Enhanced Biomass and Lipid Formation by the Oleaginous Yeast Cutaneotrichosporon oleaginosus. Front. Bioeng. Biotechnol. 2019, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, S.; Conte, C.; De Andrade, P.; Ghiselli, G.; Maugeri, F. Bioresource Technology Exploration of Brazilian biodiversity and selection of a new oleaginous yeast strain cultivated in raw glycerol. Bioresour. Technol. 2013, 138, 377–381. [Google Scholar]
- Fontanille, P.; Kumar, V.; Christophe, G.; Nouaille, R.; Larroche, C. Bioresource Technology Bioconversion of volatile fatty acids into lipids by the oleaginous yeast Yarrowia lipolytica. Bioresour. Technol. 2012, 114, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Muniraj, I.; Zappi, M.; El-Ghonemy, D.H. Microbial lipid production from potato processing wastewater using oleaginous filamentous fungi Aspergillus oryzae. Water Res. 2013, 47, 3477–3483. [Google Scholar] [CrossRef]
- Spagnuolo, M.; Yaguchi, A.; Blenner, M. Oleaginous yeast for biofuel and oleochemical production. Curr. Opin. Biotechnol. 2019, 57, 73–81. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I.K. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Park, G.W.; Chang, H.N.; Jung, K.; Seo, C.; Kim, Y.; Choi, J.H.; Woo, H.C.; Hwang, I. Production of microbial lipid by Cryptococcus curvatus on rice straw hydrolysates. Process Biochem. 2017, 56, 147–153. [Google Scholar] [CrossRef]
- Ricardo, C.; José, C.; Neto, D.; Thomaz, V.; Bittencourt, E.; Scopel, E.; Bianchi, A.; Medeiros, P.; Porto, L.; Vandenberghe, D.S. Pilot scale biodiesel production from microbial oil of Rhodosporidium toruloides DEBB 5533 using sugarcane juice: Performance in diesel engine and preliminary economic study. Bioresour. Technol. 2017, 223, 259–268. [Google Scholar]
- Ivančić Šantek, M.; Grubišić, M.; Galić Perečinec, M.; Beluhan, S.; Šantek, B. Lipid production by Mortierella isabellina from pretreated corn cobs and effect of lignocellulose derived inhibitors on growth and lipid synthesis. Process Biochem. 2021, 109, 46–58. [Google Scholar] [CrossRef]
- Tang, M.; Wang, Y.; Zhou, W.; Yang, M.; Liu, Y.; Gong, Z. Efficient conversion of chitin-derived carbon sources into microbial lipid by the oleaginous yeast Cutaneotrichosporon oleaginosum. Bioresour. Technol. 2020, 315, 123897. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zong, M.; Wu, H.; Liu, Q. Bioresource Technology Microbial oil production from rice straw hydrolysate by Trichosporon fermentans. Bioresour. Technol. 2009, 100, 4535–4538. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, Y.; Yu, Z.; Bao, J. Simultaneous saccharification and microbial lipid fermentation of corn stover by oleaginous yeast Trichosporon cutaneum. Bioresour. Technol. 2012, 118, 13–18. [Google Scholar] [CrossRef]
- Brar, K.K.; Sarma, A.K.; Aslam, M.; Polikarpov, I.; Chadha, B.S. Technology Potential of oleaginous yeast Trichosporon sp., for conversion of sugarcane bagasse hydrolysate into biodiesel. Bioresour. Technol. 2017, 242, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of structural carbohydrates and lignin in Biomass—NREL/TP-510-42618. Lab. Anal. Proced. 2012, 1617, 1–16. [Google Scholar]
- Schneiter, R.; Daum, G. Extraction of yeast lipids. Methods Mol. Biol. 2006, 313, 41–45. [Google Scholar] [PubMed]
- Šantek, M.I.; Lisičar, J.; Mušak, L.; Špoljarić, I.V.; Beluhan, S.; Šantek, B. Lipid Production by Yeast Trichosporon oleaginosus on the Enzymatic Hydrolysate of Alkaline Pretreated Corn Cobs for Biodiesel Production. Energy Fuels 2018, 32, 12501–12513. [Google Scholar] [CrossRef]
- Gao, K.; Rehmann, L. ABE fermentation from enzymatic hydrolysate of NaOH-pretreated corncobs. Biomass Bioenergy 2014, 66, 110–115. [Google Scholar] [CrossRef]
- Baadhe, R.R.; Potumarthi, R.; Mekala, N.K. Influence of dilute acid and alkali pretreatment on reducing sugar production from corncobs by crude enzymatic method: A comparative study. Bioresour. Technol. 2014, 162, 213–217. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Taherzadeh, M.J.; Karimi, K. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production: A review. Int. J. Mol. Sci. 2008, 9, 1621–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Eylen, D.; Van Dongen, F.; Kabel, M.; De Bont, J. Corn fiber, cobs and stover: Enzyme-aided saccharification and co-fermentation after dilute acid pretreatment. Bioresour. Technol. 2011, 102, 5995–6004. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Chuan, J. Microbial cellulases: Engineering, production and applications. Renew. Sustain. Energy Rev. 2014, 33, 188–203. [Google Scholar] [CrossRef]
- Expasy. EC 3.2.1.21 ß—Glucosidase. Available online: https://enzyme.expasy.org/EC/3.2.1.21 (accessed on 21 March 2022).
- Expasy. EC 3.2.1.4 Endoglucanase or ß-1,4-Endoglucan Hydrolase. Available online: https://enzyme.expasy.org/EC/3.2.1.4 (accessed on 21 March 2022).
- Expasy. EC 3.2.1.91 Exoglucanase. Available online: https://enzyme.expasy.org/EC/3.2.1.91 (accessed on 21 March 2022).
- Expasy. EC 3.2.1.176 Exoglucanase. Available online: https://enzyme.expasy.org/EC/3.2.1.176 (accessed on 21 March 2022).
- Kaar, W.E.; Holtzapple, M.T. Using lime pretreatment to facilitate the enzymic hydrolysis of corn stover. Biomass Bioenergy. 2000, 18, 189–199. [Google Scholar] [CrossRef]
- Sathitsuksanoh, N.; Zhu, Z.; Zhang, Y.P. Bioresource Technology Cellulose solvent- and organic solvent-based lignocellulose fractionation enabled efficient sugar release from a variety of lignocellulosic feedstocks. Bioresour. Technol. 2012, 117, 228–233. [Google Scholar] [CrossRef]
- Hu, F.; Ragauskas, A. Pretreatment and Lignocellulosic Chemistry. Bioenergy Resour. 2012, 5, 1043–1066. [Google Scholar] [CrossRef]
- Dai, X.; Shen, H.; Li, Q.; Rasool, K.; Wang, Q.; Yu, X.; Wang, L.; Bao, J.; Yu, D.; Zhao, Z.K. Microbial lipid production from corn stover by the oleaginous yeast Rhodosporidium toruloides using the presslp process. Energies 2019, 12, 1053. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Peng, F.; Du, W.; Liu, C.; Liu, D. Effects of some inhibitors on the growth and lipid accumulation of oleaginous yeast Rhodosporidium toruloides and preparation of biodiesel by enzymatic transesterification of the lipid. Bioprocess Biosyst. Eng. 2012, 35, 993–1004. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Mehtani, J.; Pruthi, V.; Pruthi, P.A. Assessment of fuel properties on the basis of fatty acid pro fi les of oleaginous yeast for potential biodiesel production. Renew. Sustain. Energy Rev. 2017, 77, 604–616. [Google Scholar] [CrossRef] [Green Version]
- Koppram, R.; Tomás-Pejó, E.; Xiros, C.; Olsson, L. Lignocellulosic ethanol production at high-gravity: Challenges and perspectives. Trends Biotechnol. 2014, 32, 46–53. [Google Scholar] [CrossRef]
- Sahare, P.; Singh, R.; Laxman, R.S.; Rao, M. Effect of Alkali Pretreatment on the Structural Properties and Enzymatic Hydrolysis of Corn Cob. Appl. Biochem. Biotechnol. 2012, 168, 1806–1819. [Google Scholar] [CrossRef] [PubMed]
- Arantes, V.; Saddler, J.N. Cellulose accessibility limits the effectiveness of minimum cellulase loading on the efficient hydrolysis of pretreated lignocellulosic substrates. Biotechnol. Biofuels 2011, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öhgren, K.; Bura, R.; Saddler, J.; Zacchi, G. Effect of hemicellulose and lignin removal on enzymatic hydrolysis of steam pretreated corn stover. Bioresour. Technol. 2007, 98, 2503–2510. [Google Scholar] [CrossRef] [PubMed]
- Zanchetta, A.; Carlos, A.; Ximenes, E.; Carreira, C.; Boscolo, M.; Gomes, E.; Ladisch, R. Temperature Dependent Cellulase Adsorption on Lignin from Sugarcane Bagasse. Bioresour. Technol. 2017, 252, 143–149. [Google Scholar] [CrossRef]
- Jing, X.; Zhang, X.; Bao, J. Inhibition Performance of Lignocellulose Degradation Products on Industrial Cellulase Enzymes During Cellulose Hydrolysis. Appl. Biochem. Biotechnol. 2009, 159, 696–707. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Alriksson, B.; Nilvebrant, N.O. Bioconversion of lignocellulose: Inhibitors and detoxification. Biotechnol. Biofuels 2013, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Renewable Energy Statistics. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Renewable_energy_statistics&oldid=515129#of_renewable_energy_used_in_transport_activities_in_2019 (accessed on 24 September 2021).
- Faria, N.T.; Santos, M.; Ferreira, C.; Marques, S.; Ferreira, F.C. Conversion of cellulosic materials into glycolipid biosurfactants, mannosylerythritol lipids, by Pseudozyma spp. under SHF and SSF processes. Microb. Cell Fact. 2014, 13, 155. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Pejó, E.; Oliva, J.M.; González, A.; Ballesteros, I.; Ballesteros, M. Bioethanol production from wheat straw by the thermotolerant yeast Kluyveromyces marxianus CECT 10875 in a simultaneous saccharification and fermentation fed-batch process. Fuel 2009, 88, 2142–2147. [Google Scholar] [CrossRef]
- Brandenburg, J.; Blomqvist, J.; Shapaval, V.; Kohler, A.; Sampels, S. Biotechnology for Biofuels Oleaginous yeasts respond differently to carbon sources present in lignocellulose hydrolysate. Biotechnol. Biofuels 2021, 14, 124. [Google Scholar] [CrossRef]
- Gong, Z.; Shen, H.; Yang, X.; Wang, Q.; Xie, H.; Zhao, Z.K. Lipid production from corn stover by the oleaginous yeast Cryptococcus curvatus. Biotechnol. Biofuels 2014, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Rane, D.V.; Pawar, P.P.; Odaneth, A.A.; Lali, A.M. Microbial oil production by the oleaginous red yeast, Rhodotorula glutinis NCIM 3168, using corncob hydrolysate. Biomass Convers. Biorefinery 2021, 1–11. [Google Scholar] [CrossRef]
- Yu, X.; Zeng, J.; Zheng, Y.; Chen, S. Effect of lignocellulose degradation products on microbial biomass and lipid production by the oleaginous yeast Cryptococcus curvatus. Process Biochem. 2014, 49, 457–465. [Google Scholar] [CrossRef]
- Poontawee, R.; Yongmanitchai, W.; Limtong, S. Efficient oleaginous yeasts for lipid production from lignocellulosic sugars and effects of lignocellulose degradation compounds on growth and lipid production. Process Biochem. 2017, 53, 44–60. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Robbins, C.; Ceniceros, E.; Natarajan, M. Review of biodiesel composition, properties, and specifications. Renew. Sustain. Energy Rev. 2012, 16, 143–169. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (%, g g−1) | Time | Temperature (°C) | Biomass Recovery (%, g g−1) 2 | Water Usage (mL/100 g) 1 | |
|---|---|---|---|---|---|
| NaOH | 1 | 16 h | 50 | 73.4 | 545 |
| 2 | 6 h | 50 | 67.2 | 407 | |
| 1 | 2 h | 121 | 63.6 | 437 | |
| 2 | 1 h | 121 | 44.9 | 398 | |
| H2SO4 | 2 | 10 min | 120 | 45.2 | 2896 |
| 1 | 20 min | 120 | 44.8 | 2664 |
| Composition (%, g g−1) | |||
|---|---|---|---|
| Pretreatment Conditions | Glucan | Xylan | Lignin |
| raw corn cobs | 33.96 ± 4.57 | 30.45 ± 30.73 | 13.34 ± 0.15 |
| 1% NaOH, 50 °C, 16 h | 38.68 ± 7.15 | 35.60 ± 0.96 | 7.14 ± 1.70 |
| 2% NaOH, 50 °C, 6 h | 39.92 ± 4.86 | 32.89 ± 0.25 | 8.10 ± 0.96 |
| 1% NaOH, 121 °C, 2 h | 44.02 ± 1.79 | 36.37 ± 0.34 | 4.85 ± 0.84 |
| 2% NaOH, 121 °C,1 h | 44.92 ± 0.46 | 34.16 ± 0.64 | 2.98 ± 0.39 |
| 2% H2SO4, 120 °C, 10 min | 74.18 ± 5.76 | 13.37 ± 3.24 | 24.45 ± 1.07 |
| 1% H2SO4, 120 °C, 20 min | 70.98 ± 2.55 | 22.88 ± 4.34 | 20.23 ± 1.48 |
| Pretreatment Conditions | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NaOH (%) | - | 2 | 1 | 1 | 2 | - | - | - | 2 | 1 | 1 | 2 | - | - |
| H2SO4 (%) | - | - | - | - | - | 2 | 1 | - | - | - | - | - | 2 | 1 |
| T (°C) | - | 50 | 50 | 121 | 120 | 120 | 120 | - | 50 | 50 | 121 | 120 | 120 | 120 |
| Time | - | 6 h | 16 h | 2 h | 1 h | 10 min | 20 min | - | 6 h | 16 h | 2 h | 1 h | 10 min | 20 min |
| FPU g−1 | 15 | 15 | 15 | 15 | 15 | 15 | 15 | 30 | 30 | 30 | 30 | 30 | 30 | 30 |
| Glucose (g L−1) | 9.97 * 0.12 | 46.61 * 7.94 | 51.28 * 0.44 | 56.89 * 4.64 | 59.99 * 9.67 | 8.72 * 6.21 | 52.28 * 0.21 | 19.64 * 0.12 | 63.38 * 12.38 | 51.23 * 1.13 | 62.04 * 19.30 | 66.05 * 17.87 | 15.70 * 9.74 | 50.28 * 3.17 |
| Xylose (g L−1) | 3.99 * - | 28.35 * 23.90 | 32.45 * 20.40 | 35.81 * 22.19 | 37.30 * 26.38 | 1.20 * 2.09 | 7.11 * 0.10 | 7.40 * - | 42.78 * 24.44 | 37.20 * 21.90 | 42.64 * 31.98 | 44.87 * 32.48 | 5.70 * 2.52 | 7.58 * 3.21 |
| Lipid concentration (g L−1) | 5.13 | 14.37 | 18.97 | 14.60 | 14.89 | 0.85 | 14.37 | 3.41 | 10.64 | 16.44 | 6.39 | 8.01 | 0.33 | 12.77 |
| Lipid content (%, g g−1) | 2.67 | 30.69 | 39.15 | 25.14 | 32.85 | 0.28 | 13.96 | 4.18 | 37.1 | 41.07 | 40.23 | 41.7 | 0.73 | 14.47 |
| Lipid yield (g g−1) | 0.023 | 0.096 | 0.126 | 0.097 | 0.099 | 0.006 | 0.096 | 0.034 | 0.071 | 0.110 | 0.043 | 0.053 | 0.002 | 0.085 |
| Fatty acid (%, g g−1) | ||||||||||||||
| C14:0 | 0.19 | 1.05 | 0.84 | 0.78 | 0.88 | 0.29 | 0.58 | 0.15 | 0.58 | 0.47 | 0.59 | 0.56 | - | 0.16 |
| C16:0 | 24.23 | 30.34 | 27.68 | 27.25 | 30.28 | 37.03 | 26.71 | 23.99 | 26.55 | 24.36 | 26.31 | 25.89 | 16.08 | 26.00 |
| C16:1 cis 9 | - | 1.10 | 0.87 | 0.60 | 0.73 | - | 0.42 | - | 0.45 | 0.45 | 0.49 | 0.58 | - | 0.20 |
| C17:0 | - | 0.27 | 0.03 | 0.53 | 0.21 | - | 0.18 | - | 0.17 | 0.03 | 0.16 | 0.24 | - | - |
| C18:0 | 6.94 | 16.19 | 16.98 | 17.32 | 14.51 | 3.47 | 17.46 | 12.18 | 16.96 | 17.37 | 13.68 | 12.98 | - | 15.75 |
| C18:1 cis 9 | 53.69 | 42.28 | 46.44 | 46.46 | 46.99 | 59.83 | 48.85 | 54.77 | 46.93 | 49.36 | 50.50 | 49.72 | 82.42 | 53.55 |
| C18:2 cis 9, 12 | 14.48 | 5.14 | 4.45 | 3.49 | 0.01 | 1.88 | 4.72 | 8.57 | 5.38 | 5.31 | 5.77 | 7.07 | - | 3.98 |
| C20:2 cis 11, 14 | - | 0.33 | - | - | - | 0.64 | 0.09 | 0.80 | 0.29 | - | 0.13 | 0.28 | 3.01 | - |
| C24:0 | - | 1.85 | 1.36 | 1.57 | 1.30 | - | 1.13 | 0.26 | 1.72 | 1.31 | 1.59 | 1.60 | - | 0.67 |
| SFA a | 31.36 | 49.70 | 46.89 | 47.45 | 47.18 | 40.79 | 46.06 | 36.58 | 45.98 | 43.54 | 42.33 | 41.27 | 16.08 | 42.58 |
| MUFA b | 53.69 | 43.38 | 47.31 | 47.06 | 47.72 | 59.83 | 49.27 | 54.77 | 47.38 | 49.81 | 50.99 | 50.30 | 82.42 | 53.75 |
| PUFA c | 14.48 | 5.47 | 4.45 | 3.49 | 0.01 | 2.52 | 4.81 | 9.37 | 5.67 | 5.31 | 5.90 | 7.35 | 3.01 | 3.98 |
| Pretreatment Conditions | ||||||
|---|---|---|---|---|---|---|
| NaOH (%) | 2 | 1 | 1 | 2 | - | - |
| H2SO4 (%) | - | - | - | - | 1 | 2 |
| T (°C) | 50 | 121 | 50 | 121 | 121 | 121 |
| t | 6 h | 2 h | 16 h | 1 h | 20 min | 10 min |
| Lipid concentration (g L−1) | 1.05 | 1.08 | 0.48 | 1.46 | 2.48 | 2.38 |
| Lipid content (%, g g−1) | 14.21 | 8.41 | 23.1 | 14.85 | 28.19 | 24.52 |
| Fatty acid (%, g g−1) | ||||||
| C14:0 | 1.65 | 1.25 | 1.49 | 1.53 | 0.79 | 0.80 |
| C16:0 | 31.13 | 29.94 | 32.23 | 32.89 | 26.67 | 27.70 |
| C16:1 (cis 9) | 1.55 | 1.29 | 1.32 | 1.52 | 0.91 | 0.91 |
| C17:0 | 2.75 | 2.01 | 2.61 | 2.81 | 1.35 | 0.80 |
| C18:0 | 11.69 | 10.03 | 11.90 | 11.33 | 10.51 | 12.35 |
| C18:1 (cis 9) | 42.46 | 46.63 | 42.95 | 42.36 | 44.22 | 44.21 |
| C18:2 (cis 9, 12) | 6.07 | 6.66 | 5.44 | 5.59 | 13.31 | 11.98 |
| C20:2 (cis 11, 14) | 0.53 | - | 0.39 | 0.44 | 0.21 | - |
| C24:0 | 0.71 | 0.67 | 0.68 | 0.38 | 0.71 | 0.65 |
| SFA a | 47.93 | 43.90 | 48.91 | 48.94 | 40.03 | 42.30 |
| MUFA b | 44.01 | 47.92 | 44.27 | 43.88 | 45.13 | 45.12 |
| PUFA c | 6.60 | 6.66 | 5.83 | 6.03 | 13.52 | 11.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grubišić, M.; Galić Perečinec, M.; Peremin, I.; Mihajlovski, K.; Beluhan, S.; Šantek, B.; Ivančić Šantek, M. Optimization of Pretreatment Conditions and Enzymatic Hydrolysis of Corn Cobs for Production of Microbial Lipids by Trichosporon oleaginosus. Energies 2022, 15, 3208. https://doi.org/10.3390/en15093208
Grubišić M, Galić Perečinec M, Peremin I, Mihajlovski K, Beluhan S, Šantek B, Ivančić Šantek M. Optimization of Pretreatment Conditions and Enzymatic Hydrolysis of Corn Cobs for Production of Microbial Lipids by Trichosporon oleaginosus. Energies. 2022; 15(9):3208. https://doi.org/10.3390/en15093208
Chicago/Turabian StyleGrubišić, Marina, Maja Galić Perečinec, Ines Peremin, Katarina Mihajlovski, Sunčica Beluhan, Božidar Šantek, and Mirela Ivančić Šantek. 2022. "Optimization of Pretreatment Conditions and Enzymatic Hydrolysis of Corn Cobs for Production of Microbial Lipids by Trichosporon oleaginosus" Energies 15, no. 9: 3208. https://doi.org/10.3390/en15093208
APA StyleGrubišić, M., Galić Perečinec, M., Peremin, I., Mihajlovski, K., Beluhan, S., Šantek, B., & Ivančić Šantek, M. (2022). Optimization of Pretreatment Conditions and Enzymatic Hydrolysis of Corn Cobs for Production of Microbial Lipids by Trichosporon oleaginosus. Energies, 15(9), 3208. https://doi.org/10.3390/en15093208