
A Review on Synchronous Microalgal Lipid Enhancement and Wastewater Treatment


Abstract
:1. Introduction
2. Functioning of Microalgae
2.1. Wastewater Treatment
2.2. Biofuel Production
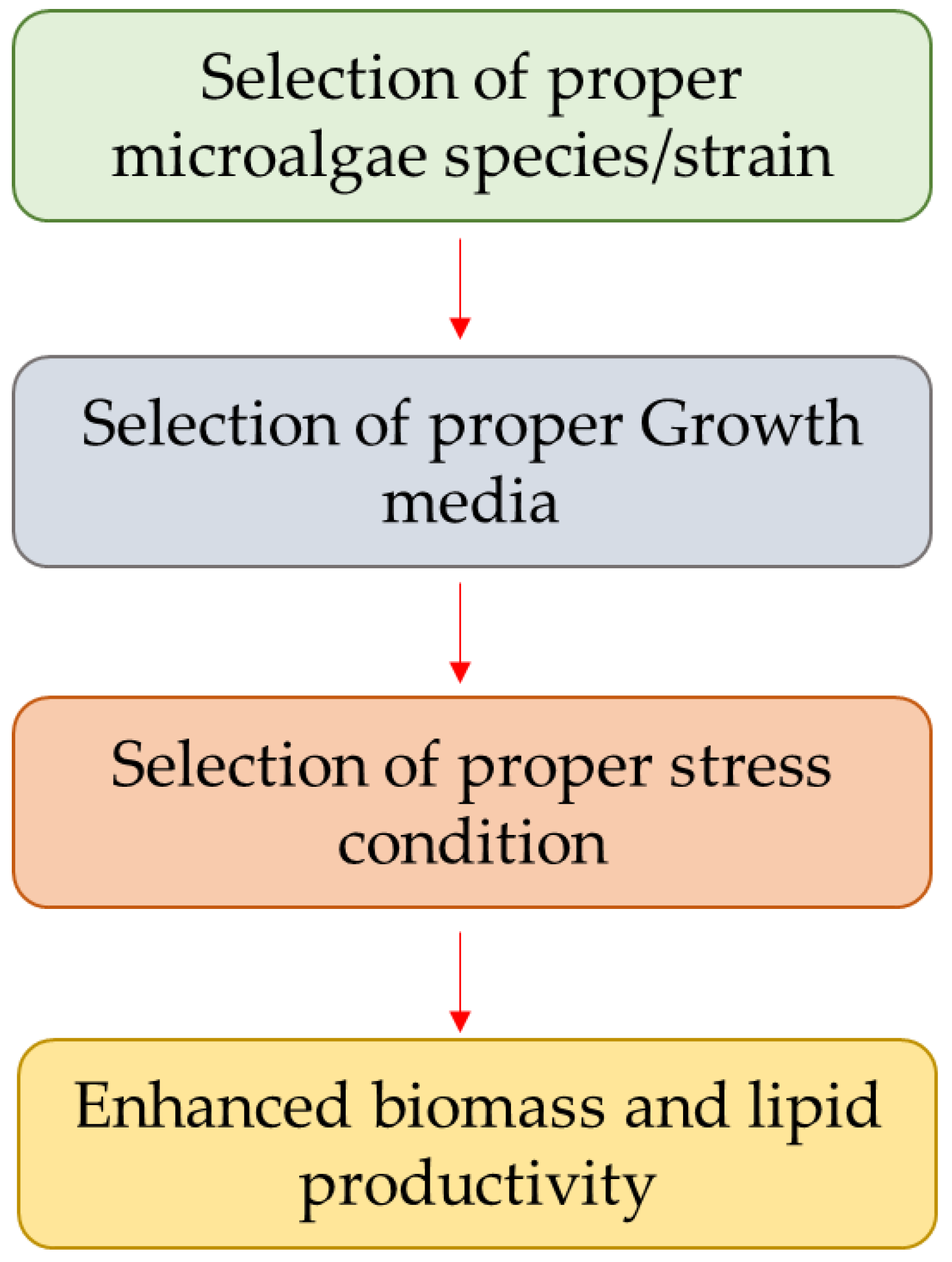
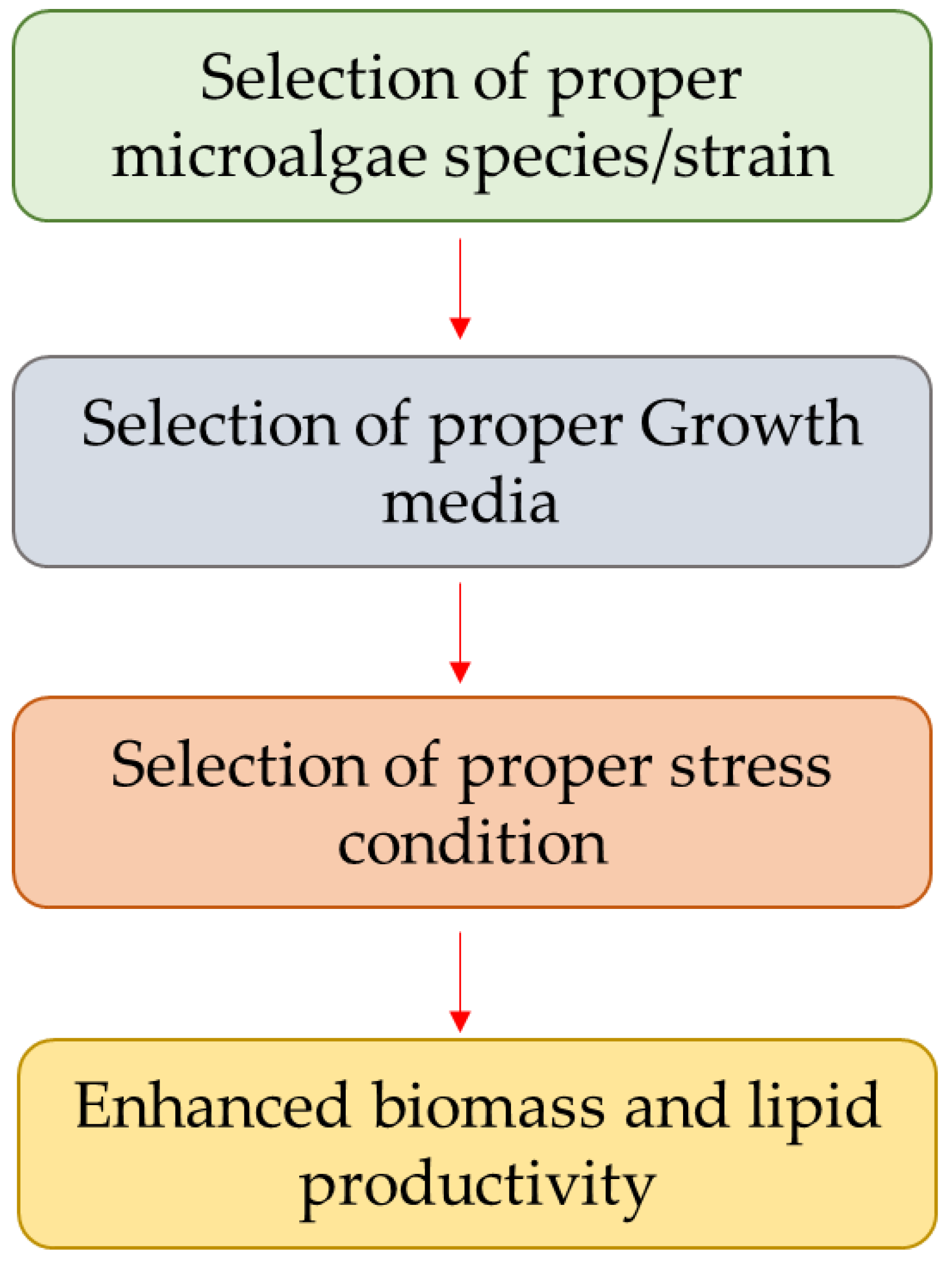
3. Strategies for Lipid Enhancement
3.1. Selection of Proper Microalgae Species or Strain
3.2. Selection of Proper Growth Media
3.3. Selection of Proper Stress Condition
3.3.1. Effect of Light
3.3.2. Effect of Temperature
3.3.3. Effect of Carbon Dioxide (CO2)
3.3.4. Effect of Nutrients

{kind=link}
{kind=link}
| Microalgae | Stress Condition | Result | Reference |
|---|---|---|---|
| Nannochloropsis oculata Chlorella vulgaris | Nitrogen deficiency | 15.31% lipid yield for N. oculata and 16.41% for C. vulgaris | [42] |
| Nannochloropsis sp. F&M-M24 | Nitrogen-deficient condition | Lipid productivity of 204 mg L−1 d−1 | [90] |
| N.oceanica DUT01 | Nitrogen-rich condition | 31 mg L−1 d−1 lipid productivity | [45] |
| Microcystis panniformis Microcystis novacekii | Phosphorus and nitrogen | Lipid accumulation had an inverse and direct correlation with nitrogen and phosphorus concentration | [100] |
| Dunaliella tertiolecta | Nitrogen | 10 folds increased nitrogen led to lipid productivity of 47.4 mg L−1 d−1 and content of 33.5% | [101] |
| Chlorella vulgaris | High iron | 3–7 folds enhanced lipid accumulation | [102] |
| Ankistrodesmus falcatus | 3–6 mg L−1 iron | 3 mg L−1 decreased the lipid content and productivity whereas 6 mg L−1 enhanced lipid content and productivity | [46] |
3.3.5. Effect of Salinity Stress
4. Integration of Wastewater Treatment, Enhanced Biomass, and Lipid Production Strategies
5. Conclusions and Future Recommendation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hemalatha, M.; Mohan, S.V. Microalgae cultivation as tertiary unit operation for treatment of pharmaceutical wastewater associated with lipid production. Bioresour. Technol. 2016, 215, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.A.H.; Yaakob, Z.; Abdullah, S.R.S.; Takriff, M.S. Assessing the feasibility of microalgae cultivation in agricultural wastewater: The nutrient characteristics. Environ. Technol. Innov. 2019, 15, 100402. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Ling, T.C.; Show, P.L.; Juan, J.C.; Chang, J.-S.; Lee, D.-J. Cultivation in wastewaters for energy: A microalgae platform. Appl. Energy 2016, 179, 609–625. [Google Scholar] [CrossRef]
- Pittman, J.K.; Dean, A.; Osundeko, O. The potential of sustainable algal biofuel production using wastewater resources. Bioresour. Technol. 2011, 102, 17–25. [Google Scholar] [CrossRef]
- Lundquist, T.J.; Woertz, I.C.; Quinn, W.M.T.; Benemann, J. A Realistic Technology and Engineering Assessment of Algae Biofuel Production. Available online: https://digitalcommons.calpoly.edu/cgi/viewcontent.cgi?article=1189&context=cenv_fac (accessed on 5 October 2021).
- Chen, G.; Zhao, L.; Qi, Y. Enhancing the productivity of microalgae cultivated in wastewater toward biofuel production: A critical review. Appl. Energy 2015, 137, 282–291. [Google Scholar] [CrossRef]
- Demirbas, A.; Demirbas, M.F. Importance of algae oil as a source of biodiesel. Energy Convers. Manag. 2011, 52, 163–170. [Google Scholar] [CrossRef]
- Zhu, L.D.; Li, Z.H.; Hiltunen, E. Strategies for Lipid Production Improvement in Microalgae as a Biodiesel Feedstock. BioMed Res. Int. 2016, 2016, 8792548. [Google Scholar] [CrossRef] [Green Version]
- Min, M.; Wang, L.; Li, Y.; Mohr, M.J.; Hu, B.; Zhou, W.; Chen, P.; Ruan, R. Cultivating Chlorella sp. in a Pilot-Scale Photobioreactor Using Centrate Wastewater for Microalgae Biomass Production and Wastewater Nutrient Removal. Appl. Biochem. Biotechnol. 2011, 165, 123–137. [Google Scholar] [CrossRef]
- Mar, C.C.; Fan, Y.; Li, F.-L.; Hu, G.-R. Bioremediation of wastewater from edible oil refinery factory using oleaginous microalga Desmodesmus sp. S1. Int. J. Phytoremediation 2016, 18, 1195–1201. [Google Scholar] [CrossRef]
- Daneshvar, E.; Antikainen, L.; Koutra, E.; Kornaros, M.; Bhatnagar, A. Investigation on the feasibility of Chlorella vulgaris cultivation in a mixture of pulp and aquaculture effluents: Treatment of wastewater and lipid extraction. Bioresour. Technol. 2018, 255, 104–110. [Google Scholar] [CrossRef]
- Chinnasamy, S.; Bhatnagar, A.; Hunt, R.W.; Das, K.C. Microalgae cultivation in a from poultry litter anaerobic digestion. Bioresour. Technol. 2010, 102, 10841–10848. [Google Scholar]
- Hongyang, S.; Yalei, Z.; Chunmin, Z.; Xuefei, Z.; Jinpeng, L. Cultivation of Chlorella pyrenoidosa in soybean processing wastewater. Bioresour. Technol. 2011, 102, 9884–9890. [Google Scholar] [CrossRef]
- Jiang, L.; Luo, S.; Fan, X.; Yang, Z.; Guo, R. Biomass and lipid production of marine microalgae using municipal wastewater and high concentration of CO2. Appl. Energy 2011, 88, 3336–3341. [Google Scholar] [CrossRef]
- Singh, M.; Reynolds, D.L.; Das, K.C. Microalgal system for treatment of effluent wastewater dominated by carpet mill effluents for biofuel applications. Bioresour. Technol. 2011, 101, 3097–3105. [Google Scholar]
- Wu, L.F.; Chen, P.C.; Huang, A.P.; Lee, C.M. The feasibility of biodiesel production by microalgae using industrial wastewater. Bioresour. Technol. 2012, 113, 14–18. [Google Scholar] [CrossRef]
- Komolafe, O.; Orta, S.B.V.; Monje-Ramirez, I.; Noguez, I.Y.; Harvey, A.; Ledesma, M.T.O. Biodiesel production from indigenous microalgae grown in wastewater. Bioresour. Technol. 2014, 154, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ramanna, L.; Guldhe, A.; Rawat, I.; Bux, F. The optimization of biomass and lipid yields of Chlorella sorokiniana when using wastewater supplemented with different nitrogen sources. Bioresour. Technol. 2014, 168, 127–135. [Google Scholar] [CrossRef]
- Ansari, F.A.; Singh, P.; Guldhe, A.; Bux, F. Microalgal cultivation using aquaculture wastewater: Integrated biomass generation and nutrient remediation. Algal Res. 2017, 21, 169–177. [Google Scholar] [CrossRef]
- Hernández-García, A.; Velásquez-Orta, S.B.; Novelo, E.; Yáñez-Noguez, I.; Monje-Ramírez, I.; Ledesma, M.T.O. Wastewater-leachate treatment by microalgae: Biomass, carbohydrate and lipid production. Ecotoxicol. Environ. Saf. 2019, 174, 435–444. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Sivaramakrishnan, R.; Incharoensakdi, A.; Cornejo, P.; Kamaraj, B.; Chi, N.T.L. Removal of nutrients from domestic wastewater by microalgae coupled to lipid augmentation for biodiesel production and influence of deoiled algal biomass as biofertilizer for Solanum lycopersicum cultivation. Chemosphere 2021, 268, 129323. [Google Scholar] [CrossRef]
- Han, W.; Jin, W.; Li, Z.; Wei, Y.; He, Z.; Chen, C.; Qin, C.; Chen, Y.; Tu, R.; Zhou, X. Cultivation of microalgae for lipid production using municipal wastewater. Process. Saf. Environ. Prot. 2021, 155, 155–165. [Google Scholar] [CrossRef]
- Vitova, M.; Bisova, K.; Kawano, S.; Zachleder, V. Accumulation of energy reserves in algae: From cell cycles to biotechnological applications. Biotechnol. Adv. 2015, 33, 1204–1218. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.H.; Raja, I.A.; Rizwan, M.; Rashid, N.; Mahmood, Q.; Shah, F.A.; Pervez, A. Potential of microalgal biodiesel production and its sustainability perspectives in Pakistan. Renew. Sustain. Energy Rev. 2018, 81, 76–92. [Google Scholar] [CrossRef]
- Taher, H.; Al-Zuhair, S.; Al-Marzouqi, A.; Haik, Y.; Farid, M. Growth of microalgae using CO2 enriched air for biodiesel production in supercritical CO2. Renew. Energy 2015, 82, 61–70. [Google Scholar] [CrossRef]
- Han, S.-F.; Jin, W.-B.; Tu, R.-J.; Wu, W. Biofuel production from microalgae as feedstock: Current status and potential. Crit. Rev. Biotechnol. 2013, 35, 255–268. [Google Scholar] [CrossRef]
- Okoro, V.; Azimov, U.; Munoz, J.; Hernandez, H.; Phan, A.N. Microalgae cultivation and harvesting: Growth performance and use of flocculants—A review. Renew. Sustain. Energy Rev. 2019, 115, 109364. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Nakano, H.; Murakami, M.; Konosu, S.; Nakayama, O.; Kanda, M.; Nakamura, A.; Iwamoto, H. Lipid Composition of a Green Alga, Botryococcus braunii. Agric. Bioi. Chem. 1987, 51, 493–498. [Google Scholar]
- Aketo, T.; Hoshikawa, Y.; Nojima, D.; Yabu, Y.; Maeda, Y.; Yoshino, T.; Takano, H.; Tanaka, T. Selection and characterization of microalgae with potential for nutrient removal from municipal wastewater and simultaneous lipid production. J. Biosci. Bioeng. 2020, 129, 565–572. [Google Scholar] [CrossRef]
- Ruangsomboon, S. Effect of light, nutrient, cultivation time and salinity on lipid production of newly isolated strain of the green microalga, Botryococcus braunii KMITL 2. Bioresour. Technol. 2012, 109, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Remmers, I.M.; Wijffels, R.H.; Barbosa, M.J.; Lamers, P.P. Can We Approach Theoretical Lipid Yields in Microalgae? Trends Biotechnol. 2018, 36, 265–276. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, W.; Li, Y.; Yang, W.; Shen, G. Mixotrophic cultivation of Botryococcus braunii. Biomass- Bioenergy 2011, 35, 1710–1715. [Google Scholar] [CrossRef]
- Toledo-Cervantes, A.; Morales, M.; Novelo, E.; Revah, S. Carbon dioxide fixation and lipid storage by Scenedesmus obtusiusculus. Bioresour. Technol. 2013, 130, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, I.A.; Cabanelas, I.T.D.; dos Santos, J.N.; Nascimento, M.A.; Sousa, L.; Sansone, G. Biodiesel yields and fuel quality as criteria for algal-feedstock selection: Effects of CO2-supplementation and nutrient levels in cultures. Algal Res. 2015, 8, 53–60. [Google Scholar] [CrossRef]
- Ge, Y.; Liu, J.; Tian, G. Growth characteristics of Botryococcus braunii 765 under high CO2 concentration in photobioreactor. Bioresour. Technol. 2011, 102, 130–134. [Google Scholar] [CrossRef]
- Mehrabadi, A.; Craggs, R.; Farid, M.M. Biodiesel production potential of wastewater treatment high rate algal pond biomass. Bioresour. Technol. 2016, 221, 222–233. [Google Scholar] [CrossRef]
- Yeh, K.L.; Chang, J.S. Nitrogen starvation strategies and photobioreactor design for enhancing lipid content and lipid production of a newly isolated microalga Chlorella vulgaris ESP-31: Implications for biofuels. Biotechnol. J. 2011, 6, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, M.; Zhang, X.; Hu, Q.; Sommerfeld, M.; Chen, Y. Life-cycle analysis on biodiesel production from microalgae: Water footprint and nutrients balance. Bioresour. Technol. 2011, 102, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Saranya, D.; Shanthakumar, S. Green microalgae for combined sewage and tannery effluent treatment: Performance and lipid accumulation potential. J. Environ. Manag. 2019, 241, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Krzemińska, I.; Oleszek, M.; Wiącek, D. Liquid Anaerobic Digestate as a Source of Nutrients for Lipid and Fatty Acid Accumulation by Auxenochlorella Protothecoides. Molecules 2019, 24, 3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, A.L.; Pires, J.C.M.; Simões, M. A review on the use of microalgal consortia for wastewater treatment. Algal Res. 2017, 24, 403–415. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process. Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Xin, L.; Hu, H.Y.; Ke, G.; Sun, Y.X. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef]
- Salama, E.-S.; Kim, H.-C.; Abou-Shanab, R.; Ji, M.-K.; Oh, Y.-K.; Kim, S.-H.; Jeon, B.-H. Biomass, lipid content, and fatty acid composition of freshwater Chlamydomonas mexicana and Scenedesmus obliquus grown under salt stress. Bioprocess Biosyst. Eng. 2013, 36, 827–833. [Google Scholar] [CrossRef]
- Wan, C.; Bai, F.-W.; Zhao, X.-Q. Effects of nitrogen concentration and media replacement on cell growth and lipid production of oleaginous marine microalga Nannochloropsis oceanica DUT01. Biochem. Eng. J. 2013, 78, 32–38. [Google Scholar] [CrossRef]
- Singh, P.; Guldhe, A.; Kumari, S.; Rawat, I.; Bux, F. Investigation of combined effect of nitrogen, phosphorus and iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surface methodology. Biochem. Eng. J. 2015, 94, 22–29. [Google Scholar] [CrossRef]
- Álvarez-Díaz, P.D.; Ruiz, J.; Arbib, Z.; Barragán, J.; Garrido-Pérez, M.C.; Perales, J.A. Freshwater microalgae selection for simultaneous wastewater nutrient removal and lipid production. Algal Res. 2017, 24, 477–485. [Google Scholar] [CrossRef]
- García, L.M.; Gariépy, Y.; Barnabé, S.; Raghavan, G. Effect of environmental factors on the biomass and lipid production of microalgae grown in wastewaters. Algal Res. 2019, 41, 101521. [Google Scholar] [CrossRef]
- Patel, A.K.; Joun, J.; Sim, S.J. A sustainable mixotrophic microalgae cultivation from dairy wastes for carbon credit, bioremediation and lucrative biofuels. Bioresour. Technol. 2020, 313, 123681. [Google Scholar] [CrossRef]
- Poh, Z.L.; Kadir, W.N.A.; Lam, M.K.; Uemura, Y.; Suparmaniam, U.; Lim, J.W.; Show, P.L.; Lee, K.T. The effect of stress environment towards lipid accumulation in microalgae after harvesting. Renew. Energy 2020, 154, 1083–1091. [Google Scholar] [CrossRef]
- Lu, M.-M.; Gao, F.; Li, C.; Yang, H.-L. Response of microalgae Chlorella vulgaris to Cr stress and continuous Cr removal in a membrane photobioreactor. Chemosphere 2021, 262, 128422. [Google Scholar] [CrossRef]
- Wahidin, S.; Idris, A.; Shaleh, S.R.M. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef]
- Hallenbeck, P.C.; Grogger, M.; Mraz, M.; Veverka, D. The use of Design of Experiments and Response Surface Methodology to optimize biomass and lipid production by the oleaginous marine green alga, Nannochloropsis gaditana in response to light intensity, inoculum size and CO2. Bioresour. Technol. 2015, 184, 161–168. [Google Scholar] [CrossRef]
- Guo, X.; Su, G.; Li, Z.; Chang, J.; Zeng, X.; Sun, Y.; Lu, Y.; Lin, L. Light intensity and N/P nutrient affect the accumulation of lipid and unsaturated fatty acids by Chlorella sp. Bioresour. Technol. 2015, 191, 385–390. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Yang, H.; Wu, L.; Hu, C. Effect of light intensity on physiological changes, carbon allocation and neutral lipid accumulation in oleaginous microalgae. Bioresour. Technol. 2015, 191, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yuan, C.; Hu, G.; Li, F. Effects of Light Intensity on the Growth and Lipid Accumulation of Microalga Scenedesmus sp. 11-1 Under Nitrogen Limitation. Appl. Biochem. Biotechnol. 2012, 166, 2127–2137. [Google Scholar] [CrossRef]
- Takeshita, T.; Ota, S.; Yamazaki, T.; Hirata, A.; Zachleder, V.; Kawano, S. Starch and lipid accumulation in eight strains of six Chlorella species under comparatively high light intensity and aeration culture conditions. Bioresour. Technol. 2014, 158, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Liao, Q.; Sun, Y.; Huang, Y.; Xia, A.; Fu, Q.; Zhu, X. Simultaneous enhancement of Chlorella vulgaris growth and lipid accumulation through the synergy effect between light and nitrate in a planar waveguide flat-plate photobioreactor. Bioresour. Technol. 2017, 243, 528–538. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers, P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. Effect of light intensity, pH, and temperature on triacylglycerol (TAG) accumulation induced by nitrogen starvation in Scenedesmus obliquus. Bioresour. Technol. 2013, 143, 1–9. [Google Scholar] [CrossRef]
- Teo, C.L.; Atta, M.; Bukhari, A.; Taisir, M.; Yusuf, A.M.; Idris, A. Enhancing growth and lipid production of marine microalgae for biodiesel production via the use of different LED wavelengths. Bioresour. Technol. 2014, 162, 38–44. [Google Scholar] [CrossRef]
- Das, P.; Lei, W.; Aziz, S.S.; Obbard, J.P. Enhanced algae growth in both phototrophic and mixotrophic culture under blue light. Bioresour. Technol. 2011, 102, 3883–3887. [Google Scholar] [CrossRef] [PubMed]
- Hultberg, M.; Jönsson, H.L.; Bergstrand, K.-J.; Carlsson, A.S. Impact of light quality on biomass production and fatty acid content in the microalga Chlorella vulgaris. Bioresour. Technol. 2014, 159, 465–467. [Google Scholar] [CrossRef]
- Rai, M.P.; Gautom, T.; Sharma, N. Effect of Salinity, pH, Light Intensity on Growth and Lipid Production of Microalgae for Bioenergy Application. Online J. Biol. Sci. 2015, 15, 260–267. [Google Scholar] [CrossRef]
- Wong, Y.-K.; Ho, Y.H.; Ho, K.C.; Leung, H.M.; Chow, K.P.; Yung, K.K.L. Effect of different light sources on algal biomass and lipid production in internal leds-illuminated photobioreactor. J. Mar. Biol. Aquacult. 2016, 2, 1–8. [Google Scholar] [CrossRef]
- Severes, A.; Hegde, S.; D’souza, L.; Hedge, S. Use of light emitting diodes (LEDs) for enhanced lipid production in microalgae-based biofuels. J. Photochem. Photobiol. B. 2017, 170, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Solovchenko, A.E.; Ismagulova, T.T.; Lukyanov, A.A.; Vasilieva, S.G.; Konyukhov, I.V.; Pogosyan, S.I.; Lobakova, E.S.; Gorelova, O.A. Luxury phosphorus uptake in microalgae. J. Appl. Phycol. 2019, 31, 2755–2770. [Google Scholar] [CrossRef]
- Xin, L.; Hong-Ying, H.; Yu-Ping, Z. Growth and lipid accumulation properties of a freshwater microalga Scenedesmus sp. under different cultivation temperature. Bioresour. Technol. 2011, 102, 3098–3102. [Google Scholar] [CrossRef]
- Dickinson, S.; Mientus, M.; Frey, D.; Amini-Hajibashi, A.; Ozturk, S.; Shaikh, F.; Sengupta, D.; El-Halwagi, M.M. A review of biodiesel production from microalgae. Clean Technol. Environ. Policy 2017, 19, 637–668. [Google Scholar] [CrossRef]
- Subhash, G.V.; Rohit, M.; Devi, M.P.; Swamy, Y.; Mohan, S.V. Temperature induced stress influence on biodiesel productivity during mixotrophic microalgae cultivation with wastewater. Bioresour. Technol. 2014, 169, 789–793. [Google Scholar] [CrossRef]
- Bohnenberger, J.; Crossetti, L.O. Influence of temperature and nutrient content on lipid production in freshwater microalgae cultures. An. Acad. Bras. Ciências 2014, 86, 1239–1248. [Google Scholar] [CrossRef] [Green Version]
- Freire, I.; Cortina-Burgueño, A.; Grille, P.; Arizcun, M.A.; Abellán, E.; Segura, M.; Sousa, F.W.; Otero, A. Nannochloropsis limnetica: A freshwater microalga for marine aquaculture. Aquaculture 2016, 459, 124–130. [Google Scholar] [CrossRef]
- Menegol, T.; Diprat, A.B.; Rodrigues, E.; Rech, R. Effect of temperature and nitrogen concentration on biomass composition of Heterochlorella luteoviridis. Food Sci. Technol. 2017, 37, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Huang, X.; Huang, Z. Temperature effects on lipid properties of microalgae Tetraselmis subcordiformis and Nannochloropsis oculata as biofuel resources. Chin. J. Oceanol. Limnol. 2015, 33, 99–106. [Google Scholar] [CrossRef]
- Renaud, S.M.; Thinh, L.-V.; Lambrinidis, G.; Parry, D.L. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- Los, D.A.; Murata, N. Membrane fluidity and its roles in the perception of environmental signals. Biochim. Biophys. Acta Biomembr. 2004, 1666, 142–157. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Singh, P. Effect of CO2 concentration on algal growth: A review. Renew. Sustain. Energy Rev. 2014, 38, 172–179. [Google Scholar] [CrossRef]
- Chiu, S.-Y.; Kao, C.-Y.; Tsai, M.-T.; Ong, S.-C.; Chen, C.-H.; Lin, C.-S. Lipid accumulation and CO2 utilization of Nannochloropsis oculata in response to CO2 aeration. Bioresour. Technol. 2009, 100, 833–838. [Google Scholar] [CrossRef]
- Yoo, C.; Choi, G.-G.; Kim, S.-C.; Oh, H.-M. Ettlia sp. YC001 showing high growth rate and lipid content under high CO2. Bioresour. Technol. 2013, 127, 482–488. [Google Scholar] [CrossRef]
- Montoya, E.Y.O.; Casazza, A.; Aliakbarian, B.; Perego, P.; de Carvalho, J.C.M.; Converti, A. Production of Chlorella vulgaris as a source of essential fatty acids in a tubular photobioreactor continuously fed with air enriched with CO2 at different concentrations. Biotechnol. Prog. 2014, 30, 916–922. [Google Scholar] [CrossRef]
- Nakanishi, A.; Aikawa, S.; Ho, S.-H.; Chen, C.-Y.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Development of lipid productivities under different CO2 conditions of marine microalgae Chlamydomonas sp. JSC4. Bioresour. Technol. 2014, 152, 247–252. [Google Scholar] [CrossRef]
- Bagchi, S.K.; Mallick, N. Carbon dioxide biofixation and lipid accumulation potential of an indigenous microalga Scenedesmus obliquus (Turpin) Kützing GA 45 for biodiesel production. RSC Adv. 2016, 6, 29889–29898. [Google Scholar] [CrossRef]
- Hui, W.; Wenjun, Z.; Wentao, C.; Lili, G.; Tianzhong, L. Strategy study on enhancing lipid productivity of filamentous oleaginous microalgae Tribonema. Bioresour. Technol. 2016, 218, 161–166. [Google Scholar] [CrossRef]
- Kao, C.-Y.; Chen, T.-Y.; Chang, Y.-B.; Chiu, T.-W.; Lin, H.-Y.; Chen, C.-D.; Chang, J.S.; Lin, C.S. Utilization of carbon dioxide in industrial flue gases for the cultivation of microalga Chlorella sp. Bioresour. Technol. 2014, 166, 485–493. [Google Scholar] [CrossRef]
- Ying, K.; Zimmerman, W.; Gilmour, D. Effects of CO2 and pH on growth of the microalga Dunaliella salina. J. Microbial. Biochem. Technol. 2014, 6, 167–173. [Google Scholar] [CrossRef]
- Peng, L.; Lan, C.Q.; Zhang, Z.; Sarch, C.; Laporte, M. Control of protozoa contamination and lipid accumulation in Neochloris oleoabundans culture: Effects of pH and dissolved inorganic carbon. Bioresour. Technol. 2015, 197, 143–151. [Google Scholar] [CrossRef]
- Ferreira, A.; Marques, P.; Ribeiro, B.; Assemany, P.; de Mendonça, H.V.; Barata, A.; Oliveira, A.C.; Reis, A.; Pinheiro, H.; Gouveia, L. Combining biotechnology with circular bioeconomy: From poultry, swine, cattle, brewery, dairy and urban wastewaters to biohydrogen. Environ. Res. 2018, 164, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.T.; Yeh, K.L.; Chang, J.S. Enzymatic transesterification of microalgal oil from Chlorella vulgaris ESP-31 for biodiesel synthesis using immobilized Burkholderia lipase. Biotechnol. Bioresour. 2012, 108, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.J.; Hill, D.R.; Olmstead, I.L.; Bergamin, A.; Shears, M.J.; Dias, D.A.; Kentish, S.E.; Scales, P.J.; Botte, C.Y.; Callahan, D.L. Lipid profile remodeling in response to nitrogen deprivation in the microalgae Chlorella sp. (Trebouxiophyceae) and Nannochloropsis sp. (Eustigmatophyceae). PLoS ONE 2014, 9, e103389. [Google Scholar] [CrossRef] [Green Version]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-H.; Wu, W.-T. Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresour. Technol. 2009, 100, 3921–3926. [Google Scholar] [CrossRef] [PubMed]
- Praveenkumar, R.; Shameera, K.; Mahalakshmi, G.; Akbarsha, M.A.; Thajuddin, N. Influence of nutrient deprivations on lipid accumulation in a dominant indigenous microalga Chlorella sp., BUM11008: Evaluation for biodiesel production. Biomass- Bioenergy 2012, 37, 60–66. [Google Scholar] [CrossRef]
- Li, Y.; Han, F.; Xu, H.; Mu, J.; Chen, D.; Feng, B.; Zeng, H. Potential lipid accumulation and growth characteristic of the green alga Chlorella with combination cultivation mode of nitrogen (N) and phosphorus (P). Bioresour. Technol. 2014, 174, 24–32. [Google Scholar] [CrossRef]
- Scragg, A.; Illman, A.; Carden, A.; Shales, S. Growth of microalgae with increased calorific values in a tubular bioreactor. Biomass- Bioenergy 2002, 23, 67–73. [Google Scholar] [CrossRef]
- Adams, C.; Godfrey, V.; Wahlen, B.; Seefeldt, L.; Bugbee, B. Understanding precision nitrogen stress to optimize the growth and lipid content tradeoff in oleaginous green microalgae. Bioresour. Technol. 2013, 131, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Lin, J. Effects of nitrogen source and concentration on biomass and oil production of a Scenedesmus rubescens like microalga. Bioresour. Technol. 2011, 102, 1615–1621. [Google Scholar] [CrossRef]
- Tao, L.; Linglin, W.; Aifen, L.; Chengwu, Z. Responses in growth, lipid accumulation, and fatty acid composition of four oleaginous microalgae to different nitrogen sources and concentrations. Chin. J. Oceanol. Limnol. 2013, 31, 1306–1314. [Google Scholar]
- Yang, L.; Chen, J.; Qin, S.; Zeng, M.; Jiang, Y.; Hu, L.; Xiao, P.; Hao, W.; Hu, Z.; Lei, A.; et al. Growth and lipid accumulation by different nutrients in the microalga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2018, 11, 40. [Google Scholar] [CrossRef]
- Kamalanathan, M.; Pierangelini, M.; Shearman, L.A.; Gleadow, R.; Beardall, J. Impacts of nitrogen and phosphorus starvation on the physiology of Chlamydomonas reinhardtii. Environ. Boil. Fishes 2016, 28, 1509–1520. [Google Scholar] [CrossRef]
- Cordeiro, R.S.; Vaz, I.C.D.; Sergia, M.S.M.; Barbosa, F.A.R. Effects of nutritional conditions on lipid production by cyanobacteria. An. Acad. Bras. Ciências 2017, 89, 2021–2031. [Google Scholar] [CrossRef]
- Mata, T.M.; Almeidab, R.; Caetanoa, N.S. Effect of the culture nutrients on the biomass and lipid productivities of microalgae Dunaliella tertiolecta. Chem. Eng. Transac. 2013, 32, 978–988. [Google Scholar]
- Liu, Z.-Y.; Wang, G.-C.; Zhou, B.-C. Effect of iron on growth and lipid accumulation in Chlorella vulgaris. Bioresour. Technol. 2008, 99, 4717–4722. [Google Scholar] [CrossRef]
- El Baky, H.A.; El-Baroty, G.S.; Bouaid, A.; Martinez, M.; Aracil, J. Enhancement of lipid accumulation in Scenedesmus obliquus by Optimizing CO2 and Fe3+ levels for biodiesel production. Bioresour. Technol. 2012, 119, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.-Y.; Liu, B.-F.; Kong, F.; Zhao, L.; Xie, G.-J.; Ren, N.-Q. Enhanced lipid accumulation of green microalga Scenedesmus sp. by metal ions and EDTA addition. Bioresour. Technol. 2014, 169, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Gorain, P.C.; Bagchi, S.K.; Mallick, N. Effects of calcium, magnesium and sodium chloride in enhancing lipid accumulation in two green microalgae. Environ. Technol. 2013, 34, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Battah, M.; El-Ayoty, Y.; Abomohra, A.E.-F.; El-Ghany, S.A.; Esmael, A. Effect of Mn2+, Co2+ and H2O2 on biomass and lipids of the green microalga Chlorella vulgaris as a potential candidate for biodiesel production. Ann. Microbiol. 2014, 65, 155–162. [Google Scholar] [CrossRef]
- Arguelles, E.D.L.R.; Laurena, A.C.; Monsalud, R.G.; Martinez-Goss, M.R. Fatty acid profile and fuel-derived physico- chemical properties of biodiesel obtained from an indigenous green microalga, Desmodesmus sp. (I-AU1), as potential source of renewable lipid and highquality biodiesel. J. Appl. Phycol. 2018, 30, 411–419. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Cohen, Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Corteggiani, C.E.C.; Telatin, A.; Vitulo, N.; Forcato, C.; D’Angelo, M.; Schiavon, R.; Vezzi, A.; Giacometti, G.M.; Morosinotto, T.; Valle, G. Chromosome scale genome assembly and transcriptome profiling of Nannochloropsis gaditana in nitrogen depletion. Mol. Plant 2014, 7, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of Environmental Factors and Nutrient Availability on the Biochemical Composition of Algae for Biofuels Production: A Review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Yuan, H.; Li, B.; Yang, J. Significance evaluation of the effects of environmental factors on the lipid accumulation of Chlorella minutissima UTEX 2341 under low-nutrition heterotrophic condition. Bioresour. Technol. 2014, 152, 177–184. [Google Scholar] [CrossRef]
- Kan, G.; Shi, C.; Wang, X.; Xie, Q.; Wang, M.; Wang, X.; Miao, J. Acclimatory responses to high-salt stress in Chlamydomonas (Chlorophyta, Chlorophyceae) from Antarctica. Acta Oceanol. Sin. 2012, 31, 116–124. [Google Scholar] [CrossRef]
- Rao, A.R.; Dayananda, C.; Sarada, R.; Shamala, T.; Ravishankar, G. Effect of salinity on growth of green alga Botryococcus braunii and its constituents. Bioresour. Technol. 2007, 98, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.K.; Schuhmann, H.; Schenk, P.M. High Lipid Induction in Microalgae for Biodiesel Production. Energies 2012, 5, 1532–1553. [Google Scholar] [CrossRef] [Green Version]
- Bartley, M.; Boeing, W.J.; Corcoran, A.A.; Holguin, F.O.; Schaub, T. Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass- Bioenergy 2013, 54, 83–88. [Google Scholar] [CrossRef]
- Figler, A.; Márton, K.; B-Béres, V.; Bácsi, I. Effects of Nutrient Content and Nitrogen to Phosphorous Ratio on the Growth, Nutrient Removal and Desalination Properties of the Green Alga Coelastrum morus on a Laboratory Scale. Energies 2021, 14, 2112. [Google Scholar] [CrossRef]
- Pandit, P.R.; Fulekar, M.H.; Karuna, M.S.L. Effect of salinity stress on growth, lipid productivity, fatty acid composition, and biodiesel properties in Acutodesmus obliquus and Chlorella vulgaris. Environ. Sci. Pollut. Res. 2017, 24, 13437–13451. [Google Scholar] [CrossRef]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Salinity induced oxidative stress alters the physiological responses and improves the biofuel potential of green microalgae Acutodesmus dimorphus. Bioresour. Technol. 2017, 244, 1376–1383. [Google Scholar] [CrossRef]
- Srivastava, G.; Nishchal; Goud, V.V. Salinity induced lipid production in microalgae and cluster analysis (ICCB 16-BR_047). Bioresour. Technol. 2017, 242, 244–252. [Google Scholar] [CrossRef]
- Sydney, E.; da Silva, T.; Tokarski, A.; Novak, A.; de Carvalho, J.; Woiciecohwski, A.; Larroche, C.; Soccol, C. Screening of microalgae with potential for biodiesel production and nutrient removal from treated domestic sewage. Appl. Energy 2011, 88, 3291–3294. [Google Scholar] [CrossRef]
- Jayakumar, S.; Yusoff, M.; Rahim, M.H.A.; Maniam, G.P.; Govindan, N. The prospect of microalgal biodiesel using agro-industrial and industrial wastes in Malaysia. Renew. Sustain. Energy Rev. 2017, 72, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Ngo, H.; Guo, W.; Chang, S.; Nguyen, D.D.; Kumar, S. Microalgae biomass from swine wastewater and its conversion to bioenergy. Bioresour. Technol. 2019, 275, 109–122. [Google Scholar] [CrossRef]
- Diniz, G.S.; Silva, A.F.; Araujo, O.; Chaloub, R.M. The potential of microalgal biomass production for biotechnological purposes using wastewater resources. J. Appl. Phycol. 2017, 29, 821–832. [Google Scholar] [CrossRef]
- Fields, M.W.; Hise, A.; Lohman, E.J.; Bell, T.; Gardner, R.D.; Corredor, L.; Moll, K.; Peyton, B.M.; Characklis, G.W.; Gerlach, R. Sources and resources: Importance of nutrients, resource allocation, and ecology in microalgal cultivation for lipid accumulation. Appl. Microbiol. Biotechnol. 2014, 98, 4805–4816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Li, Y.; Min, M.; Hu, B.; Chen, P.; Ruan, R. Local bioprospecting for high-lipid producing microalgal strains to be grown on concentrated municipal wastewater for biofuel production. Bioresour. Technol. 2011, 102, 6909–6919. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, W.; Hu, B.; Min, M.; Chen, P.; Ruan, R.R. Effect of light intensity on algal biomass accumulation and biodiesel production for mixotrophic strains Chlorella kessleri and Chlorella protothecoide cultivated in highly concentrated municipal wastewater. Biotechnol. Bioeng. 2012, 109, 2222–2229. [Google Scholar] [CrossRef]
- Cabanelas, I.T.D.; Arbib, Z.; Chinalia, F.A.; Souza, C.O.; Perales, J.A.; Almeida, P.F.; Druzian, J.I.; Nascimento, I.A. From waste to energy: Microalgae production in wastewater and glycerol. Appl. Energy 2013, 109, 283–290. [Google Scholar] [CrossRef]
- Singh, A.; Ummalyma, S.B.; Sahoo, D. Bioremediation and biomass production of microalgae cultivation in river water contaminated with pharmaceutical effluent. Bioresour. Technol. 2020, 307, 123233. [Google Scholar] [CrossRef]
- Collos, Y.; Harrison, P. Acclimation and toxicity of high ammonium concentrations to unicellular algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef]
- Samorì, G.; Samorì, C.; Guerrini, F.; Pistocchi, R. Growth and nitrogen removal capacity of Desmodesmus communis and of a natural microalgae consortium in a batch culture system in view of urban wastewater treatment: Part I. Water Res. 2013, 47, 791–801. [Google Scholar] [CrossRef]
- Salama, E.-S.; Kurade, M.; Abou-Shanab, R.A.; El-Dalatony, M.M.; Yang, I.-S.; Min, B.; Jeon, B.-H. Recent progress in microalgal biomass production coupled with wastewater treatment for biofuel generation. Renew. Sustain. Energy Rev. 2017, 79, 1189–1211. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kazimierowicz, J.; Kujawska, N.; Talbierz, S. Microalgae Cultivation Technologies as an Opportunity for Bioenergetic System Development—Advantages and Limitations. Sustainability 2020, 12, 9980. [Google Scholar] [CrossRef]
- Bošnjaković, M.; Sinaga, N. The Perspective of Large-Scale Production of Algae Biodiesel. Appl. Sci. 2020, 10, 8181. [Google Scholar] [CrossRef]
- Mehariya, S.; Goswami, R.; Verma, P.; Lavecchia, R.; Zuorro, A. Integrated Approach for Wastewater Treatment and Biofuel Production in Microalgae Biorefineries. Energies 2021, 14, 2282. [Google Scholar] [CrossRef]
- Silkina, A.; Ginnever, N.E.; Fernandes, F.; Fuentes-Grünewald, C. Large-Scale Waste Bio-Remediation Using Microalgae Cultivation as a Platform. Energies 2019, 12, 2772. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Ahn, Y.; Pandi, K.; Ji, M.-K.; Yun, H.-S.; Choi, J.-Y. Microalgae Cultivation in Pilot Scale for Biomass Production Using Exhaust Gas from Thermal Power Plants. Energies 2019, 12, 3497. [Google Scholar] [CrossRef] [Green Version]
- Sasongko, N.A.; Noguchi, R.; Ito, J.; Demura, M.; Ichikawa, S.; Nakajima, M.; Watanabe, M.M. Engineering Study of a Pilot Scale Process Plant for Microalgae-Oil Production Utilizing Municipal Wastewater and Flue Gases: Fukushima Pilot Plant. Energies 2018, 11, 1693. [Google Scholar] [CrossRef] [Green Version]

| Microalgae | Wastewater | Results | Reference |
|---|---|---|---|
| Chlorella sp. UMN271 | Swine manure-based | Biomass productivity ranged from 8.08–14.59 g/m2 × day and lipid content of 1.77–3.55% | [9] |
| Desmodesmus sp. S1 | Oil refinery | Biomass and lipid content were found to be 2.98 g/L and 21.95%, respectively | [10] |
| Chlorella vulgaris | Pulp and aquaculture | Biomass productivity of 187 mg/L/d and lipid content of 9.07% | [11] |
| Mix consortium | Carpet mill treated | Biomass productivity of 41 mg/L/d and lipid content of 12.2% | [12] |
| Chlorella pyrenoidosa | Soybean processing | Biomass productivity of 0.64 g/L/d | [13] |
| Chlorella vulgaris | Tertiary-treated domestic | Biomass productivity of 197 gL−1 and lipid productivity of 0.164 gL−1 | [14] |
| Scenedesmus bijuga | 6% effluent from poultry litter anaerobic digestion | Biomass productivity of 31–76 mg/L/d | [15] |
| Chlamydomonas sp. TAI-2 | Untreated industrial | Biomass yield of 1.5 g/L | [16] |
| Desmodesmus sp. | Municipal | Biomass productivity of 500 mg/L/d and lipid content of 3.3% | [17] |
| C. sorokiniana | Domestic wastewater with urea supplementation | Biomass productivity of 200 mg/L/d and lipid content of 61.52% | [18] |
| Scenedesmus obliquus Chlorella sorokiniana Ankistrodesmus falcatus | Aquaculture | A. falcatus showed the highest biomass productivity (160.79 mg L−1 d−1) and lipid productivity (57.72 mg L−1 d−1) | [19] |
| Desmodesmus spp. S. obliquus | Municipal wastewater with different leachate | 33% increase in lipid content | [20] |
| Chlorella sp. | Municipal wastewater | Lipid content of 34.83% | [21] |
| Scenedesmus obliquus | Municipal wastewater | 0.33–0.38 g L−1 of total lipid | [22] |
| Microalgae | Media | Stress Condition | Result | Reference |
|---|---|---|---|---|
| Desmodesmus spp. | Municipal wastewater with leachate | Intensity of 53 µmol/m2/s and light cycles of 12:12. High ammonia concentration (≥167 mg/L) | Biomass productivity of 1.95 g/L and lipid content of 20% | [20] |
| P.kessleri NKG021201 | Municipal wastewater | Illumination at 40 mmol/m2/s at 25 °C | Biomass productivity of 125 ± 8 mg/L/d and lipid content of 38 ± 1% | [29] |
| Nannochloropsis oculate Chlorella vulgaris | Bold’s Basal Medium for C. vulgari f2 for N. oculata | 20 °C for N. oculata and 30 °C for C. vulgaris Nitrogen deficiency CO2 contained in air (about 300 ppm) | Lipid yield enhanced to 15.31% for N. oculata and 16.41% for C. vulgaris | [42] |
| Scenedesmus sp. | BG11 | Light intensity of 55–60 μmol photon·m−2 s−1, light/dark ratio of 14:10 at 25 °C Phosphate deficiency | Lipid accumulation of 53% | [43] |
| C. mexicana GU73240 S. obliquus HM103382 | Bold basal medium | Illumination at 40 μmol (photon)/m2s at 27 °C for 20 days Increased NaCl dose | Highest dry weight (0.8 and 0.65 g/L) and lipid content (37 and 34%) of C. mexicana and S. obliquus repectively | [44] |
| N. oceanica DUT01 | f/2 seawater BG 11 | 14/10 h light/dark cycle under an intensity of 60 μmol m−2 s−1 at 25 °C 2% CO2 Nitrogen rich | Lipid productivity of 31 mg L−1 d−1 | [45] |
| Ankistrodesmus falcatus | Blue-Green (BG11) medium | Photon flux of 120 μmol m−2 s−1, under a 16 h:8 h light dark cycle at 25 °C Iron sufficient and deficient | At 3 mg L−1 the lipid content and productivity decreased whereas lipid content and productivity enhanced at 6 mg L−1 iron concentration | [46] |
| Chlorella vulgaris Chlorella kessleri Scenedesmus obliquus | Secondary treated urban wastewater | Illumination of 143 μmol·m−2·s−1 and 14/10 light/dark cycle at 20 ± 1 °C 4% CO2 | Scenedesmus obliquus yielded the highest biomass concentration (1.4 g/L) and lipid content (36.75%) C. vulgaris reached the highest biomass productivity (0.107 g/L·d) followed by S.obliquus (4.4 mg Total N/L·d) | [47] |
| Mixed consortium mainly consisted ofChlorella sp. | Mixed waste streams (liquid digestate obtained after the filtration of the compost, liquid coming from the septic system sludge treatment plant and an effluent coming from the wastewater treatment plant) | Photoperiod of 12:12 h at 20 °C CO2 supplemented at a rate of 0.2 volume of air per volume of medium per minute (vvm). | Biomass productivity (105.2 mg⋅L−1⋅d−1) was negatively affected by CO2 addition and positively affected by light intensity. Higher lipid contents (17.2%) were found at low light intensity | [48] |
| C. protothecoides UTEX-256 | Pretreated dairy wastewater | 5% CO2 was supplied light intensity was 150 μmol m−2 s−1 for 9 days | 40% pretreated whey was most productive for biomass and lipid fractions, respectively, 4.54 and 1.80 gl−1 with daily productivities 0.50 and 0.20 gl−1d−1 | [49] |
| Chlorella vulgaris | Chicken waste compost mixed with tap water | 1 day of nutrient starvation with 6 g/L of salinity stress at dark | Lipid content was recorded at 40.28% | [50] |
| Chlorella vulgaris | BG11 medium | Illumination at 135 μmol photon m−2 s−1 at 25 °C low concentration of Cr(VI) | Biomass productivity of 28.3–35.9 mg L−1 d−1 | [51] |
| Microalgae | Stress Condition | Result | Reference |
|---|---|---|---|
| Botryococcus braunii KMITL 2 | Light intensities of 200 and 538 μE −2 s−1 | More lipid accumulation than lower light intensity of 87.5 μE m−2 s−1 | [30] |
| Phaeodactylum tricornutum | Light intensity of 60 µmol photons m−2 s−1 | TAG yield of 112 mg molph−1 | [31] |
| Nannochloropsis sp. | 100 µmol m−2 s−1 light intensity photoperiod of 18 h light: 6 h dark cycle | Enhanced lipid accumulation of up to 31.3% | [52] |
| Chlorella sp. L1 | 400 µmol photon m−2 s−1 light intensity | 51.4 mg L−1 d−1 lipid productivity | [55] |
| Nannochloropsis sp. | Light intensity of 700 μmol photons/m2/s | Enhanced lipid content (47% of dry weight) | [56] |
| Scenedesmus sp. | 143 μmol·m−2·s−1 and 14/10 light/dark cycle at 20 ± 1 °C | Lipid accumulation enhanced | [57] |
| Chlorella vulgaris | Light intensity of 560 μE m−2 s−1 | 92.89% enhanced lipid yield | [59] |
| Scenedesmus obliquus | 200–1500 μmol photons/m2/s light intensity | Lipid content remained the same | [60] |
| Microalgae | Stress Condition | Result | Reference |
|---|---|---|---|
| N. oculata C. vulgaris | 20–25 °C | Increased temperature decreases lipid content in C. vulgaris and increases lipid production for N.oculata | [42] |
| Mixed microalgae culture | 30 °C | 5-fold rise in neutral lipid | [70] |
| Monoraphidium consortiums Desmodesmus quadricauda | 13 °C | Aids in lipid accumulation | [71] |
| Nannochloropsis limnetica | 22 °C | highest growth and lipid productivity was observed | [72] |
| Heterochlorella luteoviridis | 22–27 °C | 40.7% of PUFAs at a temperature of 22 °C, whereas 52.9% saturated fatty acids increased at 27 °C | [73] |
| Scenedesmus sp. LX1. | 10–25 °C | Enhanced lipid build-up at low temperature | [43] |
| Scenedesmus sp. LX1 | 30 °C | Most of the fatty acids saturated | [68] |
| Microalgae | Stress Condition | Result | Reference |
|---|---|---|---|
| N. oculata | 2% and 15% CO2 | Lipid productivity of 142 and 82 mg L−1 d−1 | [78] |
| Nannochloropsis sp. | 15% CO2 | Increased biomass productivity (0.39–1.43 gL−1) and growth rate (0.33–0.52 d−1) | [14] |
| Ettlia sp. YC001 | 10% CO2 | 3.1 g L−1 cell density and 80.0 mg L−1 d−1 lipid productivity | [79] |
| C. vulgaris | 8% CO2 | 29.5 mg L−1 d−1 lipid productivity | [80] |
| Chlamydomonas sp. JSC4 | 4% CO2 | 169.1 mg L−1 d−1 lipid productivity | [81] |
| Scenedesmus Obliquus | 15% CO2 | 850 mg/L of lipid in 16 days | [82] |
| Tribonema minus | (0.03–20%) CO2 | Reduced lipid productivity with increased CO2 | [83] |
| Microalgae | Stress Condition | Response | Reference |
|---|---|---|---|
| Botrycococcus braunii N. oculata | Salinity concentration of 34 mM and 85 mM | 1.7–2.25-fold increase in palmitic acid and 2-fold increase in oleic acid at 34 mM and 85 mM for B. braunii. For N. exican, rise in temperature led to an enhanced lipid production by 2-fold | [113] |
| Nannochloropsis salina | Salt concentration of 34, 46, and 58 PSU | Highest total fatty acids content of 36% dry tissue mass at 34 PSU | [115] |
| Chlamydomonas mexicana Scenedesmus obliquus | 25 mM NaCl | Maximum lipid content of 37% and 34% for C. mexicana and S. obliquus | [44] |
| C. vulgaris Acutodesmus obliquus | 0.4 M NaCl | Highest growth and lipid productivity was observed | [117] |
| Acutodesmus dimorphus | 200 mM NaCl | 33.40 ± 2.29% lipid accumulation | [118] |
| Chlorella sorokiniana CG12(KR905186) Desmodesmus GS12(KR905187) | NaCl, KCl, MgCl2 and CaCl2 | CaCl2 improved up to 40.02–44.97% in Chlorella sorokiniana CG12(KR905186) and Desmodesmus GS12(KR905187) | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barua, V.B.; Munir, M. A Review on Synchronous Microalgal Lipid Enhancement and Wastewater Treatment. Energies 2021, 14, 7687. https://doi.org/10.3390/en14227687
Barua VB, Munir M. A Review on Synchronous Microalgal Lipid Enhancement and Wastewater Treatment. Energies. 2021; 14(22):7687. https://doi.org/10.3390/en14227687
Chicago/Turabian StyleBarua, Visva Bharati, and Mariya Munir. 2021. "A Review on Synchronous Microalgal Lipid Enhancement and Wastewater Treatment" Energies 14, no. 22: 7687. https://doi.org/10.3390/en14227687
APA StyleBarua, V. B., & Munir, M. (2021). A Review on Synchronous Microalgal Lipid Enhancement and Wastewater Treatment. Energies, 14(22), 7687. https://doi.org/10.3390/en14227687