The Senescence (Stay-Green)—An Important Trait to Exploit Crop Residuals for Bioenergy


Abstract
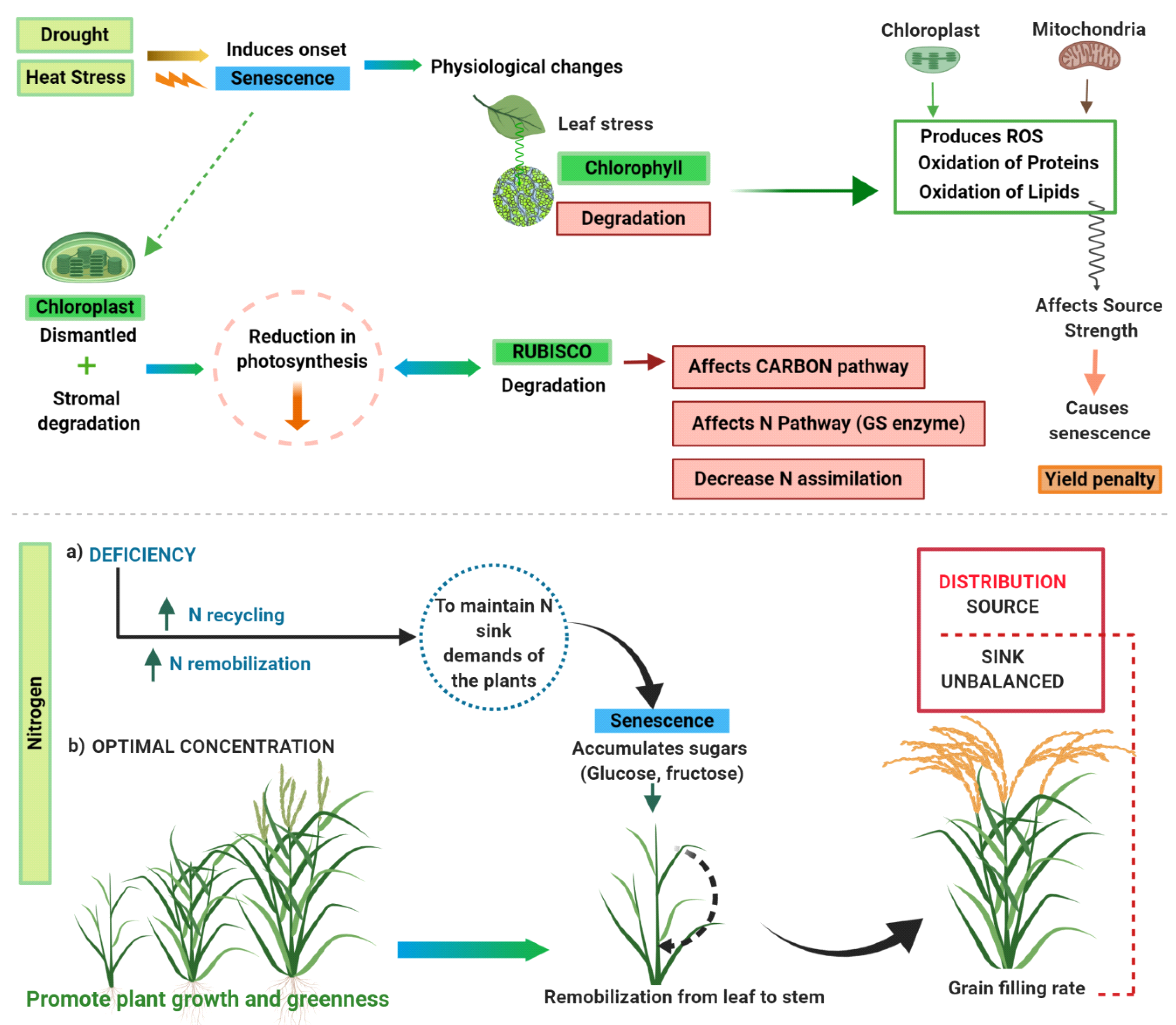
1. Introduction
2. Biological Processes of SG and Relationship with Chlorophyll
3. Genetics of SG and Associated Traits
4. The Senescence or SG in the Breeding of Bioenergy Crops
- The specific characteristics of the crop, including if the crop is annual or perennial
- The part of the plant used for obtaining energy: the whole plant or only the residuals. For example, in cereals the grain can be used for animal feeding and the remaining parts (leaves, stalks and cobs), that are traditional seen as residuals, can be used for energy.
5. Field High-Throughput Phenotyping for SG
5.1. Current Platforms and Sensors in Field-High-Throughput Phenotyping
5.2. Current Models in Field-High-Throughput Phenotyping
6. Conclusions and Further Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hörtensteiner, S.; Feller, U. Nitrogen metabolism and remobilization during senescence. J. Exp. Bot. 2002, 53, 927–937. [Google Scholar] [CrossRef]
- Aasen, H.; Honkavaara, E.; Lucieer, A.; Zarco-Tejada, J.P. Quantitative remote sensing at ultra-high resolution with UAV spectroscopy: A review of sensor technology, measurement procedures and data correction workflows. Remote Sens. 2018, 10, 1091. [Google Scholar] [CrossRef]
- Balazadeh, S. Stay-green not always stays green. Mol. Plant. 2014, 7, 1264–1266. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Assotiates: Sunderland, MA, USA, 2015. [Google Scholar]
- Lim, P.O.; Nam, H.G. The molecular and genetic control of leaf senescence and longevity in Arabidopsis. Curr. Top. Dev. Biol. 2005, 67, 50–85. [Google Scholar]
- Schippers, J.H.; Schmidt, R.; Wagstaff, C.; Jing, H.-C. Living to die and dying to live: The survival strategy behind leaf senescence. Plant Physiol. 2015, 169, 914–930. [Google Scholar] [CrossRef]
- Schippers, J.H.M. Transcriptional networks in leaf senescence. Curr. Opin. Plant Biol. 2015, 27, 77–83. [Google Scholar] [CrossRef]
- Thomas, H. Senescence, ageing and death of the whole plant. New Phytol. 2013, 197, 696–711. [Google Scholar] [CrossRef]
- Jibran, R.; Hunter, D.A.; Dijkwel, P.P. Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef]
- Jan, S.; Abbas, N.; Ashraf, M.; Ahmad, P. Roles of potential plant hormones and transcription factors in controlling leaf senescence and drought tolerance. Protoplasma 2019, 256, 313–329. [Google Scholar] [CrossRef]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51 (Suppl. 1), 329–337. [Google Scholar] [CrossRef]
- Peleg, Z.; Reguera, M.; Tumimbang, E.; Walia, H.; Blumwald, E. Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol. J. 2011, 9, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Reguera, M.; Peleg, Z.; Abdel-Tawab, Y.M.; Tumimbang, E.B.; Delatorre, C.A.; Blumwald, E. Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice. Plant Physiol. 2013, 163, 1609–1622. [Google Scholar] [CrossRef] [PubMed]
- Guiboileau, A.; Sormani, R.; Meyer, C.; Masclaux-Daubresse, C. Senescence and death of plant organs: Nutrient recycling and developmental regulation. Comptes Rendus Biol. 2010, 333, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, J.-H.; Ma, X.; Luo, D.-X.; Gong, Z.-H.; Lu, M.-H. The plant heat stress transcription factors (HSFs): Structure, regulation and function in response to abiotic stresses. Front. Plant. Sci. 2016, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Ougham, H. The stay-green trait. J. Exp. Bot. 2014, 65, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.R.; Aljadi, M.; Brewer, L. The importance of cosmetic Stay-Green in specialty crops. Plant Breed. Rev. 2018, 42, 219–256. [Google Scholar]
- Duvick, D.N. The contribution of breeding to yield advances in maize (Zea mays L.). Adv. Agron. 2005, 86, 83–145. [Google Scholar]
- Lee, E.; Tollenaar, M. Physiological basis of successful breeding strategies for maize grain yield. Crop Sci. 2007, 47 (Suppl. 3), S202–S215. [Google Scholar] [CrossRef]
- Robson, P.R.; Donnison, I.S.; Wang, K.; Frame, B.; Pegg, S.E.; Thomas, A.; Thomas, H. Leaf senescence is delayed in maize expressing the Agrobacterium IPT gene under the control of a novel maize senescence-enhanced promoter. Plant Biotechnol. J. 2004, 2, 101–112. [Google Scholar] [CrossRef]
- Joshi, S.; Choukimath, A.; Isenegger, D.; Panozzo, J.; Spangenberg, G.; Kant, S. Improved wheat growth and yield by delayed leaf senescence using developmentally regulated expression of a cytokinin biosynthesis gene. Front. Plant. Sci. 2019, 10, 1285. [Google Scholar] [CrossRef]
- Smidansky, E.D.; Meyer, F.D.; Blakeslee, B.; Weglarz, T.E.; Greene, T.W.; Giroux, M.J. Expression of a modified ADP-glucose pyrophosphorylase large subunit in wheat seeds stimulates photosynthesis and carbon metabolism. Planta 2007, 225, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Piao, W.; Kim, S.-H.; Lee, B.-D.; An, G.; Sakuraba, Y.; Paek, N.-C. Rice transcription factor OsMYB102 delays leaf senescence by down-regulating abscisic acid accumulation and signaling. J. Exp. Bot. 2019, 70, 2699–2715. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Lu, S.; Lv, B.; Zhang, B.; Shen, J.; He, J.; Luo, L.; Xi, D.; Chen, X.; Ming, F. A rice NAC transcription factor promotes leaf senescence via ABA biosynthesis. Plant Physiol. 2017, 174, 1747–1763. [Google Scholar] [CrossRef] [PubMed]
- Kusaba, M.; Ito, H.; Morita, R.; Iida, S.; Sato, Y.; Fujimoto, M.; Kawasaki, S.; Tanaka, R.; Hirochika, H.; Nishimura, M. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell 2007, 19, 1362–1375. [Google Scholar] [CrossRef]
- Sato, Y.; Morita, R.; Katsuma, S.; Nishimura, M.; Tanaka, A.; Kusaba, M. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J. 2009, 57, 120–131. [Google Scholar] [CrossRef]
- Armstead, I.; Donnison, I.; Aubry, S.; Harper, J.; Hörtensteiner, S.; James, C.; Mani, J.; Moffet, M.; Ougham, H.; Roberts, L. From crop to model to crop: Identifying the genetic basis of the staygreen mutation in the Lolium/Festuca forage and amenity grasses. New Phytol. 2006, 172, 592–597. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Park, S.-Y.; Kim, Y.-S.; Wang, S.-H.; Yoo, S.-C.; Hörtensteiner, S.; Paek, N.-C. Arabidopsis STAY-GREEN2 is a negative regulator of chlorophyll degradation during leaf senescence. Mol. Plant. 2014, 7, 1288–1302. [Google Scholar] [CrossRef]
- Jiang, H.; Li, M.; Liang, N.; Yan, H.; Wei, Y.; Xu, X.; Liu, J.; Xu, Z.; Chen, F.; Wu, G. Molecular cloning and function analysis of the stay green gene in rice. Plant J. 2007, 52, 197–209. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Deng, L.; Yan, B.; Pan, Y.; Luo, M.; Chen, X.-Q.; Hu, T.-Z.; Chen, G.-P. Silencing of the LeSGR1 gene in tomato inhibits chlorophyll degradation and exhibits a stay-green phenotype. Biol. Plant. 2011, 55, 27–34. [Google Scholar] [CrossRef]
- Zhang, J.; Fengler, K.A.; Van Hemert, J.L.; Gupta, R.; Mongar, N.; Sun, J.; Allen, W.B.; Wang, Y.; Weers, B.; Mo, H. Identification and characterization of a novel stay-green QTL that increases yield in maize. Plant Biotechnol. J. 2019, 17, 2272–2285. [Google Scholar] [CrossRef]
- Jagadish, K.S.; Kishor, K.; Polavarapu, B.; Bahuguna, R.N.; von Wirén, N.; Sreenivasulu, N. Staying alive or going to die during terminal senescence-an enigma surrounding yield stability. Front. Plant. Sci. 2015, 6, 1070. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Burch, D.; Badenhorst, P.; Palanisamy, R.; Mason, J.; Spangenberg, G. Regulated expression of a cytokinin biosynthesis gene IPT delays leaf senescence and improves yield under rainfed and irrigated conditions in canola (Brassica napus L.). PLoS ONE 2015, 10, e0116349. [Google Scholar] [CrossRef]
- Gentinetta, E.; Ceppl, D.; Lepori, C.; Perico, G.; Motto, M.; Salamini, F. A major gene for delayed senescence in maize. Pattern of photosynthates accumulation and inheritance. Plant Breed. 1986, 97, 193–203. [Google Scholar] [CrossRef]
- Beavis, W.; Smith, O.; Grant, D.; Fincher, R. Identification of quantitative trait loci using a small sample of topcrossed and F4 progeny from maize. Crop Sci. 1994, 34, 882–896. [Google Scholar] [CrossRef]
- Yang, Z.; Li, X.; Zhang, N.; Wang, X.; Zhang, Y.; Ding, Y.; Kuai, B.; Huang, X. Mapping and validation of the quantitative trait loci for leaf stay-green-associated parameters in maize. Plant Breed. 2017, 136, 188–196. [Google Scholar] [CrossRef]
- Guei, R.; Wassom, C. Genetic analysis of tassel size and leaf senescence and their relationships with yield in two tropical lowland maize populations. Afr. Crop Sci. J. 1996, 4, 275–281. [Google Scholar]
- Câmara, T.M.M.; Bento, D.A.V.; Alves, G.F.; Santos, M.F.; Moreira, J.U.V.; Souza Júnior, C.L.D. Genetic parameters of drought tolerance related traits in tropical maize. Bragantia 2007, 66, 595–603. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, A.; Zheng, C.; Wang, Y.; Cai, R.; Shen, X.; Xu, R.; Liu, P.; Kong, L.; Dong, S. QTL mapping of maize (Zea mays) stay-green traits and their relationship to yield. Plant Breed. 2009, 128, 54–62. [Google Scholar] [CrossRef]
- Wang, A.-Y.; Li, Y.; Zhang, C.-Q. QTL mapping for stay-green in maize (Zea mays). Can. J. Plant Sci. 2012, 92, 249–256. [Google Scholar] [CrossRef]
- Belícuas, P.R.; Aguiar, A.M.; Bento, D.A.V.; Camara, T.M.M.; de Souza Junior, C.L. Inheritance of the stay-green trait in tropical maize. Euphytica 2014, 198, 163–173. [Google Scholar] [CrossRef]
- Almeida, G.D.; Nair, S.; Borém, A.; Cairns, J.; Trachsel, S.; Ribaut, J.-M.; Bänziger, M.; Prasanna, B.M.; Crossa, J.; Babu, R. Molecular mapping across three populations reveals a QTL hotspot region on chromosome 3 for secondary traits associated with drought tolerance in tropical maize. Mol. Breed. 2014, 34, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Kante, M.; Revilla, P.; De La Fuente, M.; Caicedo, M.; Ordás, B. Stay-green QTLs in temperate elite maize. Euphytica 2016, 207, 463–473. [Google Scholar] [CrossRef]
- Thomas, H. Sid: A Mendelian locus controlling thylakoid membrane disassembly in senescing leaves of Festuca pratensis. Theor. Appl. Genet. 1987, 73, 551–555. [Google Scholar] [CrossRef]
- Cha, K.-W.; Lee, Y.-J.; Koh, H.-J.; Lee, B.-M.; Nam, Y.-W.; Paek, N.-C. Isolation, characterization and mapping of the stay green mutant in rice. Theor. Appl. Genet. 2002, 104, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Guiamét, J.J.; Teeri, J.A.; Noodén, L.D. Effects of nuclear and cytoplasmic genes altering chlorophyll loss on gas exchange during monocarpic senescence in soybean. Plant Cell Physiol. 1990, 31, 1123–1130. [Google Scholar]
- Kumar, U.; Joshi, A.K.; Kumari, M.; Paliwal, R.; Kumar, S.; Röder, M.S. Identification of QTLs for stay green trait in wheat (Triticum aestivum L.) in the ‘Chirya 3’בSonalika’population. Euphytica 2010, 174, 437–445. [Google Scholar] [CrossRef]
- Tao, Y.; Henzell, R.; Jordan, D.; Butler, D.; Kelly, A.; McIntyre, C. Identification of genomic regions associated with stay green in sorghum by testing RILs in multiple environments. Theor. Appl. Genet. 2000, 100, 1225–1232. [Google Scholar] [CrossRef]
- Crasta, O.; Xu, W.; Rosenow, D.; Mullet, J.; Nguyen, H. Mapping of post-flowering drought resistance traits in grain sorghum: Association between QTLs influencing premature senescence and maturity. Mol. Gen. Genet. 1999, 262, 579–588. [Google Scholar] [CrossRef]
- Jiang, G.-H.; He, Y.-Q.; Xu, C.-G.; Li, X.-H.; Zhang, Q. The genetic basis of stay-green in rice analyzed in a population of doubled haploid lines derived from an indica by japonica cross. Theor. Appl. Genet. 2004, 108, 688–698. [Google Scholar] [CrossRef]
- Yoo, S.-C.; Cho, S.-H.; Zhang, H.; Paik, H.-C.; Lee, C.-H.; Li, J.; Yoo, J.-H.; Koh, H.-J.; Seo, H.S.; Paek, N.-C. Quantitative trait loci associated with functional stay-green SNU-SG1 in rice. Mol. Cells 2007, 24, 83–94. [Google Scholar]
- Yamamoto, T.; Suzuki, T.; Suzuki, K.; Adachi, S.; Sun, J.; Yano, M.; Ookawa, T.; Hirasawa, T. Characterization of a genomic region that maintains chlorophyll and nitrogen contents during ripening in a high-yielding stay-green rice cultivar. Field Crops Res. 2017, 206, 54–64. [Google Scholar] [CrossRef]
- Ramkumar, M.; Senthil Kumar, S.; Gaikwad, K.; Pandey, R.; Chinnusamy, V.; Singh, N.K.; Singh, A.K.; Mohapatra, T.; Sevanthi, A.M. A novel stay-green mutant of rice with delayed leaf senescence and better harvest index confers drought tolerance. Plants 2019, 8, 375. [Google Scholar] [CrossRef] [PubMed]
- Borrell, A.K.; Mullet, J.E.; George-Jaeggli, B.; van Oosterom, E.J.; Hammer, G.L.; Klein, P.E.; Jordan, D.R. Drought adaptation of stay-green sorghum is associated with canopy development, leaf anatomy, root growth and water uptake. J. Exp. Bot. 2014, 65, 6251–6263. [Google Scholar] [CrossRef] [PubMed]
- Moreau, L.; Charcosset, A.; Gallais, A. Use of trial clustering to study QTL × environment effects for grain yield and related traits in maize. Theor. Appl. Genet. 2004, 110, 92–105. [Google Scholar] [CrossRef]
- Bogard, M.; Jourdan, M.; Allard, V.; Martre, P.; Perretant, M.R.; Ravel, C.; Heumez, E.; Orford, S.; Snape, J.; Griffiths, S. Anthesis date mainly explained correlations between post-anthesis leaf senescence, grain yield and grain protein concentration in a winter wheat population segregating for flowering time QTLs. J. Exp. Bot. 2011, 62, 3621–3636. [Google Scholar] [CrossRef]
- Kim, J.; Woo, H.R.; Nam, H.G. Towards systems understanding of leaf Senescence: An integrated multi-omics perspective on leaf senescence research. Mol. Plant. 2016, 9, 813–825. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Syaifullah, S.J.; Roitsch, T. Integration of multi-omics techniques and physiological phenotyping within a holistic phenomics approach to study senescence in model and crop plants. J. Exp. Bot. 2017, 69, 825–844. [Google Scholar] [CrossRef]
- Sade, N.; del Mar Rubio-Wilhelmi, M.; Umnajkitikorn, K.; Blumwald, E. Stress-induced senescence and plant tolerance to abiotic stress. J. Exp. Bot. 2017, 69, 845–853. [Google Scholar] [CrossRef]
- Penfold, C.A.; Buchanan-Wollaston, V. Modelling transcriptional networks in leaf senescence. J. Exp. Bot. 2014, 65, 3859–3873. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering quantitative trait variation for crop improvement by genome editing. Cell 2017, 171, 470–480.e8. [Google Scholar] [CrossRef]
- Gregersen, P.L. Senescence and Nutrient Remobilization in Crop Plants. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; Wiley Online Library: Hoboken, NJ, USA, 2011; pp. 83–102. [Google Scholar]
- Bernardo, R. Bandwagons I, too, have known. Theor. Appl. Genet. 2016, 129, 2323. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic selection in plant Breeding: Methods, models and perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Rajcan, I.; Tollenaar, M. Source:sink ratio and leaf senescence in maize: II. Nitrogen metabolism during grain filling. Field Crops Res. 1999, 60, 255–265. [Google Scholar] [CrossRef]
- Gregersen, P.; Holm, P.; Krupinska, K. Leaf senescence and nutrient remobilisation in barley and wheat. Plant Biol. 2008, 10, 37–49. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant senescence and crop productivity. Plant Mol. Biol. 2013, 82, 603–622. [Google Scholar] [CrossRef]
- Chen, K.; Kumudini, S.V.; Tollenaar, M.; Vyn, T.J. Plant biomass and nitrogen partitioning changes between silking and maturity in newer versus older maize hybrids. Field Crops Res. 2015, 183, 315–328. [Google Scholar] [CrossRef]
- Chen, K.; Camberato, J.J.; Tuinstra, M.R.; Kumudini, S.V.; Tollenaar, M.; Vyn, T.J. Genetic improvement in density and nitrogen stress tolerance traits over 38 years of commercial maize hybrid release. Field Crops Res. 2016, 196, 438–451. [Google Scholar] [CrossRef]
- Arriola, K.G.; Kim, S.C.; Huisden, C.M.; Adesogan, A.T. Stay-green ranking and maturity of corn hybrids: 1. Effects on dry matter yield, nutritional value, fermentation characteristics and aerobic stability of silage hybrids in Florida. J. Dairy Sci. 2012, 95, 964–974. [Google Scholar] [CrossRef]
- Mangold, A.; Lewandowski, I.; Hartung, J.; Kiesel, A. Miscanthus for biogas production: Influence of harvest date and ensiling on digestibility and methane hectare yield. GCB Bioenergy 2019, 11, 50–62. [Google Scholar] [CrossRef]
- Hoskinson, R.L.; Karlen, D.L.; Birrell, S.J.; Radtke, C.W.; Wilhelm, W.W. Engineering, nutrient removal and feedstock conversion evaluations of four corn stover harvest scenarios. Biomass Bioenergy 2007, 31, 126–136. [Google Scholar] [CrossRef]
- Fagernäs, L.; Brammer, J.; Wilén, C.; Lauer, M.; Verhoeff, F. Drying of biomass for second generation synfuel production. Biomass Bioenergy 2010, 34, 1267–1277. [Google Scholar] [CrossRef]
- Hartmann, A.; Presterl, T.; Geiger, H.H. Determination of the optimal harvest date of silage maize with low and fast stover ripening. Landbauforsch. Völkenrode Sonderh. 2000, 217, 86–93. [Google Scholar]
- Schlagheck, A.; Entrup, N.L.; Freitag, M. Effect of the ripening character (“Stay Green”/“Dry Down”) on the in vitro digestibility of maize genotypes with regard to different parts of the maize plant. Landbauforschung Völkenrode, Sonderheft 2000, 217, 94–101. [Google Scholar]
- Serrano, C.; Monedero, E.; Portero, H.; Jiménez, E.; Ordás, B. Efficient biofuel production from traditional maize under low input. Agron. Sustain. Dev. 2014, 34, 561–567. [Google Scholar] [CrossRef]
- Hudiburg, T.W.; Davis, S.C.; Parton, W.; Delucia, E.H. Bioenergy crop greenhouse gas mitigation potential under a range of management practices. GCB Bioenergy 2015, 7, 366–374. [Google Scholar] [CrossRef]
- Yang, J.; Udvardi, M. Senescence and nitrogen use efficiency in perennial grasses for forage and biofuel production. J. Exp. Bot. 2017, 69, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Robson, P.; Mos, M.; Clifton-Brown, J.; Donnison, I. Phenotypic variation in senescence in Miscanthus: Towards optimising biomass quality and quantity. Bioenergy Res. 2012, 5, 95–105. [Google Scholar] [CrossRef]
- Mangold, A.; Lewandowski, I.; Möhring, J.; Clifton-Brown, J.; Krzyżak, J.; Mos, M.; Pogrzeba, M.; Kiesel, A. Harvest date and leaf:stem ratio determine methane hectare yield of miscanthus biomass. GCB Bioenergy 2019, 11, 21–33. [Google Scholar] [CrossRef]
- Jensen, E.; Robson, P.; Farrar, K.; Thomas Jones, S.; Clifton-Brown, J.; Payne, R.; Donnison, I. Towards Miscanthus combustion quality improvement: The role of flowering and senescence. GCB Bioenergy 2017, 9, 891–908. [Google Scholar] [CrossRef]
- Salehi Jouzani, G.; Sharafi, R.; Soheilivand, S. Fueling the future; plant genetic engineering for sustainable biodiesel production. Biofuel Res. J. 2018, 5, 829–845. [Google Scholar] [CrossRef]
- Bos, H.L.; Meesters, K.P.; Conijn, S.G.; Corré, W.J.; Patel, M.K. Comparing biobased products from oil crops versus sugar crops with regard to non-renewable energy use, GHG emissions and land use. Ind. Crops Prod. 2016, 84, 366–374. [Google Scholar] [CrossRef]
- Kinney, A.; Clemente, T. Modifying soybean oil for enhanced performance in biodiesel blends. Fuel Process. Technol. 2005, 86, 1137–1147. [Google Scholar] [CrossRef]
- Woyann, L.G.; Meira, D.; Zdziarski, A.D.; Matei, G.; Milioli, A.S.; Rosa, A.C.; Madella, L.A.; Benin, G. Multiple-trait selection of soybean for biodiesel production in Brazil. Ind. Crops Prod. 2019, 140, 111721. [Google Scholar] [CrossRef]
- Nath, U.K.; Kim, H.-T.; Khatun, K.; Park, J.-I.; Kang, K.-K.; Nou, I.-S. Modification of fatty acid profiles of rapeseed (Brassica napus L.) oil for using as food, industrial feed-stock and biodiesel. Plant Breed. Biotech. 2016, 4, 123–134. [Google Scholar] [CrossRef]
- Kumudini, S.; Hume, D.; Chu, G. Genetic improvement in short-season soybeans. Crop Sci. 2002, 42, 141–145. [Google Scholar] [CrossRef]
- Kumudini, S.; Hume, D.J.; Chu, G. Genetic improvement in short season soybeans. Crop Sci. 2001, 41, 391–398. [Google Scholar] [CrossRef]
- Koeslin-Findeklee, F.; Horst, W. Contribution of nitrogen uptake and retranslocation during reproductive growth to the nitrogen efficiency of winter oilseed-rape cultivars (Brassica napus L.) differing in leaf senescence. Agronomy 2016, 6, 1. [Google Scholar] [CrossRef]
- Wang, K.; Froehlich, J.E.; Zienkiewicz, A.; Hersh, H.L.; Benning, C. A plastid phosphatidylglycerol lipase contributes to the export of acyl groups from plastids for seed oil biosynthesis. Plant Cell 2017, 29, 1678–1696. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Cao, X.; Yang, Z.; Ohlrogge, J.B. Lipid turnover during senescence. Plant Sci. 2013, 205, 13–19. [Google Scholar] [CrossRef]
- Harbach, C.; Allen, T.; Bowen, C.; Davis, J.; Hill, C.; Leitman, M.; Leonard, B.; Mueller, D.; Padgett, G.; Phillips, X. Delayed senescence in soybean: Terminology, research update and survey results from growers. Plant Health Prog. 2016, 17, 76–83. [Google Scholar] [CrossRef]
- Singh, A.; Ganapathysubramanian, B.; Singh, A.K.; Sarkar, S. Machine Learning for High-Throughput Stress Phenotyping in Plants. Trends Plant Sci. 2016, 21, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, J.; Zhao, C.; Li, Z.; Huang, Y.; Yu, H.; Xu, B.; Yang, X.; Zhu, D.; Zhang, X.; et al. Unmanned Aerial Vehicle Remote Sensing for Field-Based Crop Phenotyping: Current Status and Perspectives. Front. Plant Sci. 2017, 8, 1111. [Google Scholar] [CrossRef]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Kefauver, S.C. Breeding to adapt agriculture to climate change: Affordable phenotyping solutions. Curr. Opin. Plant Biol. 2018, 45, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating high-throughput phenotyping into genetic gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, F.; Kirchgessner, N.; Schneider, D.; Walter, A.; Hund, A. Remote, aerial phenotyping of maize traits with a mobile multi-sensor approach. Plant Methods 2015, 11, 9. [Google Scholar] [CrossRef]
- Fahlgren, N.; Gehan, M.A.; Baxter, I. Lights, camera, action: High-throughput plant phenotyping is ready for a close-up. Curr. Opin. Plant Biol. 2015, 24, 93–99. [Google Scholar] [CrossRef]
- Câmara, T. Mapeamento de QTLs de caracteres relacionados à tolerância ao estresse hídrico em milho tropical. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2006. [Google Scholar]
- Caicedo Villafuerte, M. Mejora genética de maíz para senescencia retrasada “Stay Green”. Ph.D. Thesis, Universidad de Santigo de Compostela, Santiago de Compostela, Spain, 2018. [Google Scholar]
- Gracia-Romero, A.; Kefauver, C.S.; Fernandez-Gallego, A.J.; Vergara-Díaz, O.; Nieto-Taladriz, T.M.; Araus, L.J. UAV and Ground Image-Based Phenotyping: A Proof of Concept with Durum Wheat. Remote Sens. 2019, 11, 1244. [Google Scholar] [CrossRef]
- Makanza, R.; Zaman-Allah, M.; Cairns, E.J.; Magorokosho, C.; Tarekegne, A.; Olsen, M.; Prasanna, M.B. High-Throughput Phenotyping of Canopy Cover and Senescence in Maize Field Trials Using Aerial Digital Canopy Imaging. Remote Sens. 2018, 10, 330. [Google Scholar] [CrossRef]
- Deery, D.M.; Rebetzke, G.J.; Jimenez-Berni, J.A.; James, R.A.; Condon, A.G.; Bovill, W.D.; Hutchinson, P.; Scarrow, J.; Davy, R.; Furbank, R.T. Methodology for High-Throughput Field Phenotyping of Canopy Temperature Using Airborne Thermography. Front. Plant. Sci. 2016, 7, 1808. [Google Scholar] [CrossRef] [PubMed]
- Cerrudo, D.; González Pérez, L.; Mendoza Lugo, A.J.; Trachsel, S. Stay-Green and Associated Vegetative Indices to Breed Maize Adapted to Heat and Combined Heat-Drought Stresses. Remote Sens. 2017, 9, 235. [Google Scholar] [CrossRef]
- Sagan, V.; Maimaitijiang, M.; Sidike, P.; Eblimit, K.; Peterson, K.T.; Hartling, S.; Esposito, F.; Khanal, K.; Newcomb, M.; Pauli, D. Uav-based high resolution thermal imaging for vegetation monitoring and plant phenotyping using ici 8640 p, flir vue pro r 640 and thermomap cameras. Remote Sens. 2019, 11, 330. [Google Scholar] [CrossRef]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative Aerial and Ground Based High Throughput Phenotyping for the Genetic Dissection of NDVI as a Proxy for Drought Adaptive Traits in Durum Wheat. Front. Plant. Sci. 2018, 9, 893. [Google Scholar] [CrossRef]
- Dodig, D.; Božinović, S.; Nikolić, A.; Zorić, M.; Vančetović, J.; Ignjatović-Micić, D.; Delić, N.; Weigelt-Fischer, K.; Junker, A.; Altmann, T. Image-Derived Traits Related to Mid-Season Growth Performance of Maize Under Nitrogen and Water Stress. Front. Plant. Sci. 2019, 10, 814. [Google Scholar] [CrossRef]
- Makanza, R.; Zaman-Allah, M.; Cairns, J.E.; Eyre, J.; Burgueño, J.; Pacheco, Á.; Diepenbrock, C.; Magorokosho, C.; Tarekegne, A.; Olsen, M.; et al. High-throughput method for ear phenotyping and kernel weight estimation in maize using ear digital imaging. Plant Methods 2018, 14, 49. [Google Scholar] [CrossRef]
- Miller, N.D.; Haase, N.J.; Lee, J.; Kaeppler, S.M.; de Leon, N.; Spalding, E.P. A robust, high-throughput method for computing maize ear, cob and kernel attributes automatically from images. Plant J. 2017, 89, 169–178. [Google Scholar] [CrossRef]
- Christopher, J.T.; Veyradier, M.; Borrell, A.K.; Harvey, G.; Fletcher, S.; Chenu, K. Phenotyping novel stay-green traits to capture genetic variation in senescence dynamics. Funct. Plant Biol. 2014, 41, 1035–1048. [Google Scholar] [CrossRef]
- Salas Fernandez, M.G.; Bao, Y.; Tang, L.; Schnable, P.S. A high-throughput, field-based phenotyping technology for tall biomass crops. Plant Physiol. 2017, 174, 2008. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Jimenez-Berni, J.A.; Bovill, W.D.; Deery, D.M.; James, R.A. High-throughput phenotyping technologies allow accurate selection of stay-green. J. Exp. Bot. 2016, 67, 4919–4924. [Google Scholar] [CrossRef]
- Berger, K.; Atzberger, C.; Danner, M.; D’Urso, G.; Mauser, W.; Vuolo, F.; Hank, T. Evaluation of the PROSAIL model capabilities for future hyperspectral model environments: A Review Study. Remote Sens. 2018, 10, 85. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Verhoef, W.; Baret, F.; Bacour, C.; Zarco-Tejada, P.J.; Asner, G.P.; François, C.; Ustin, S.L. PROSPECT + SAIL models: A review of use for vegetation characterization. Remote Sens. Environ. 2009, 113, S56–S66. [Google Scholar] [CrossRef]
- Parent, B.; Leclere, M.; Lacube, S.; Semenov, M.A.; Welcker, C.; Martre, P.; Tardieu, F. Maize yields over Europe may increase in spite of climate change, with an appropriate use of the genetic variability of flowering time. Proc. Natl. Acad. Sci. USA 2018, 115, 10642–10647. [Google Scholar] [CrossRef] [PubMed]
- Maes, W.H.; Steppe, K. Perspectives for remote sensing with unmanned aerial vehicles in precision agriculture. Trends Plant Sci. 2019, 24, 152–164. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Gitelson, A.A.; Schepers, J.S.; Walthall, C.L. Application of spectral remote sensing for agronomic decisions. Agron. J. 2008, 100, S117–S131. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Crop | Trait * | Locus/QTLs ** | Chromosomez | Material | Reference | Comments |
|---|---|---|---|---|---|---|
| Maize (Zea mays) | Visual scores | One locus | Lo876o2 and B73 | Gentinetta et al. (1986) [34] | Visual rating, “unexpected symptoms of senescence” in the F1 and BC1 crosses | |
| Visual scores: scale 1-9 | 3 QTLs | LG 2, 6, 9 | Mo17 and B73 | Beavis et al. (1994) [35] | ||
| Visual green appearance: scale 1 (green) and 5 (senescent) | La Posta Sequia and Pool 26 Sequia | Guei and Wasson (1996) [37] | La Posta Sequia is a late (120 days) dent, Pool 26 Sequia (flint/dent) | |||
| Visual score | 33 QTLs | Chr 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 | Two tropical maize populations, L14-04B, L08-05F, L20-01F, L02-08D | Camara (2006) [101] | 23 QTLsonchr 1, 2, 5 [38] | |
| SG value as relative green leaf area | 6 QTLs | Chr 1, 2, 3, 5 and 8 | SG inbred line (Q319) and a normal inbred line (Mo17) | Zheng et al. (2009) [39] | Measured SG at 20, 40, 50 and 60 days after flowering. Results showed at 40. Relative green leaf area = green leaf area at an interval time after flowering/maximum leaf area at flowering 100%. | |
| Green leaf area: GL1, GL2 and GL3 | 15 QTLs | Chr 1, 4 and 5 and 9 | A150-3-2 (a stay-green inbred line) and Mo17 (a normal inbred line) | Wang et al. (2012) [40] | GL1, green leaf area per plant in the whole growing period. GL2, green leaf area per plant at 30 d after flowering. GL3, green leaf area per plant at the grain-ripening stage | |
| Visual score: scale 1 (highest) to 5 (lowest) SG | 17 QTL | Chr 1, 2, 3, 4, 6 and 9 | L-14-04B (low phenotype) and L-08-05F (high phenotype) and similar flowering dates | Belícuas et al. 2014 [41] | The SG trait recorded on the 120th day after sowing, i.e., 55 days after the average silking date of this population. | |
| Visual score: scale 0 (0% senescence) to 10 (100% dead leaf) and Chlorophyll content | 8 QTLs | Chr 1, 3, 4, 5, 8 and 10 | CYMMIT parental lines—CML444, CML441, CML440 and CML504 adapted to tropical and subtropical environments | Almeida et al. 2014 [42] | QTLs detected under water stressed conditions only and QTLs of Chlorophyll content in the ear leaves using a SPAD meter. Functional stay-green FS854 QTL. | |
| Visual score: scale 1 (dead leaves) to 5 (completely healthy leaves) | 3 QTLs | Chr 1, 5 and 10 | Crosses (PHG39 × EA1070) and PHG39 × B73 [flint × dent] | Kante et al. (2016) [43] | In all regions the favorable allele derived from the SG line PHG39. Chlorophyll content used a meter (CCM200 Opti-Sciences, USA) | |
| Leaf stay-green area = stay-green part area/maximum leaf area × 100%. | 8 QTLs | Chr 1, 3, 4, 5, 6, 8 and 10 | Cross between inbred line SG (Zheng58) and inbred derived from B73 | Yang et al. (2017) [78] | Relative chlorophyll contents measured on the ear leaf using a Konica Minolta SPAD 502 (Konica Minolta Holdings, Chiyodaku, Tokyo, Japan) dual-wave-length chlorophyll meter. | |
| Chlorophyllindex | 19 QTLs | Chr 1, 3, 4, 5 and 6 | Caicedo (2018) [102] | Divided as 3QTLs, 8, 4, 3, 1 in chromosomes 1, 3, 4, 5 and 6, respectively. | ||
| Visual score: scale 1 = IHP1-like, (fully senescenced) and yellow, 3 = ILP1-like, (not senescenced and green) | 1 major QTL | Chr 3 | Illinois High Protein 1 (IHP1) and Illinois Low Protein 1 (ILP1) lines | Jun Zhang et al. (2019) [31] | Identification of a novel stay-green QTL that increases yield in maize | |
| Rice (Oryza sativa L.) | Chlorophyll con-centration and photosynthetic activity | sgr(t) locus | Chr 9 | Hwacheongbyeo crossed with a tongil rice cultivar Milyang23 | Cha et al. 2002 [45] | The stay green phenotype was controlled by a single recessive locus, sgr(t). Concentration was measured with UV/VIS spectrophotometer (Sinco, Korea) and the photosynthetic rate with LI-6400 (Li-Cor, USA) |
| Visual score: scale 1 (nearly complete leaf death) and 5 (complete green leaf) | Zhenshan97 (Indica parent) and Wuyujing2 (stay-green japonica parent) | Jiang et al. (2004) [50] | Visual scores and greenness degree measured at heading and 30 days after heading using a Minolta Chlorophyll Meter SPAD-502 (Minolta Camera Co., Japan) | |||
| Chlorophyll contents | 3 QTLs | Chr 7 and 9 | SNU-SG1 (high chlorophyll content and delayed leaf senescence) crossed by japonica cultivar, Ilpumbyeo. And SNG-SG1 ×Milyang23 (M23). | Yoo et al. (2007) [51] | Quantitative trait loci associated with functional stay-green SNU-SG1 in rice. Epistatic interactions were found. Chorophyll measures with Minolta Chlo-rophyll Meter SPAD-502 (Minolta, Japan) | |
| Photosynthetic pigments | NYC1 | Chr 1 | nyc1-1 was isolated from a rice (Oryza sativa cv Koshihikari) | Kusaba et al. (2007) [25] | Suggested nyc1, chlorophyll b reductase. nyc1 mutant cosmetic. | |
| Photosynthetic pigments | NOL gene | Chr 3 | nol-1, nol-2 and nol-3 | Sato et al. (2009) [26] | sgr mutant described as cosmetic. Lines nol 1,2,3 were isolated from SG mutant lines derived from the rice cultivar Nipponbare. | |
| Leaf chlorophyll content | 6 QTLs for higher leaf chlorophyll content 4 QTL associated to Nitrogen 5 QTL rate of transport of nitrogen to the shoots | Chr 1, 3, 4, 6, 8, 9, 11 and 12 | RILs from the cross Akenohoshi and Koshihikari | Yamamoto et al. (2017) [52] | 4 QTL associated to Nitrogen. QTL (qCHR1) stable at chr 3 associated with nitrogen 5 QTL rate of transport of nitrogen to the shoots. | |
| Chlorophyll content | Chr 9 | Three mutants in M7 generation (SGM-1, SGM-2 and SGM-3). EMS mutant resources of Nagina 22 | Ramkumar et al. (2019) [53] | The rate of net photosynthetic was measured with a portable photosynthesis system (Li-6400XT, LI-COR Biosciences, Lincoln, NE, USA). A novel SG mutant of rice with delayed leaf senescence |
| Sensors | Advantages | Limitations |
|---|---|---|
| RGB (red, green, blue) |
|
|
| Multispectraland Hyperspectral |
|
|
| Infrared-thermal |
|
|
| LIDAR (light detection and ranging) |
|
|
| Synthetic Aperture Radar (SAR) Systems |
| |
| Spectroradiometers |
|
|
| RS Method | Trait | H 2 [%] * | Crop | Sensor Deployed | Reference | Comments |
|---|---|---|---|---|---|---|
| Aerial Platforms | ||||||
| Leaf biomass | h2 > 85 | Maize | RGB and thermal sensors | Liebisch et al., 2015 [99] | Canopy Cover and NDVI indices (Zeppelin NT aircraft) | |
| Crop growth, leaf, canopy senescence | h2 > 53–60 | Maize | RGB sensor | Makanza et al., 2018 [104] | UAS | |
| Grain yield | h2 > 62–86 | Wheat | RGB, multispectral and thermal sensors | Gracia-Romero et al., 2019 [103] | Ground and aerial platforms;UAS | |
| Canopy temperature | h2 > 50–79 | Wheat | Thermal sensor | Deery et al., 2016 [105] | Airborne platform (manned helicopter) | |
| Stay-Green (SG) andGrain yield (GY) | h2 > 38–98(GY) h2 > 48–9(SG) | Maize | Multispectral sensor | Cerrudo et al., 2017 [106] | NDVI and multispectral indices Airborne platform | |
| Plant temperature | h2 > 72 | Sorghum | Thermal sensors | Sagan et al., 2019 [107] | UAS | |
| Crop biomass-Grain yield | h2 = 77–89 | Wheat | Multispectral indices; Hand-held Greenseaker | Condorelli et al., 2018 [108] | NDVI indices; UAS andPhenomobile | |
| Crop biomass-Grain yield | h2 = 77–89 | Wheat | Multispectral indices; Hand-held Greenseaker | Condorelli et al., 2018 [108] | NDVI indices; UAS andPhenomobile | |
| Ground-based platforms | ||||||
| Biomass | h2 > 56 | Maize | Fluorescence, VIS and NIR imaging | Dodig et al., 2019 [109] | CCD cameras on ground-carriers | |
| Kernel weight | h2 = 29–70 | Maize | RGB sensor | Makanza et al., 2018 [110] | Sony camera on a ground tripod | |
| Kernel attributes | h2 > 90 | Maize | RGB sensor | Miller et al., 2017 [111] | Image analysis; Web service | |
| Senescence(Stay-Green) | h2 = 50–77 | Wheat | Hand-held Greenseaker | Christopher et al., 2014 [112] | NDVI | |
| Plant height | h2>97 | Sorghum | RGB sensors | Salas Fernández et al., 2017 [113] | Phenomobile | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munaiz, E.D.; Martínez, S.; Kumar, A.; Caicedo, M.; Ordás, B. The Senescence (Stay-Green)—An Important Trait to Exploit Crop Residuals for Bioenergy. Energies 2020, 13, 790. https://doi.org/10.3390/en13040790
Munaiz ED, Martínez S, Kumar A, Caicedo M, Ordás B. The Senescence (Stay-Green)—An Important Trait to Exploit Crop Residuals for Bioenergy. Energies. 2020; 13(4):790. https://doi.org/10.3390/en13040790
Chicago/Turabian StyleMunaiz, Eduardo D., Susana Martínez, Arun Kumar, Marlon Caicedo, and Bernardo Ordás. 2020. "The Senescence (Stay-Green)—An Important Trait to Exploit Crop Residuals for Bioenergy" Energies 13, no. 4: 790. https://doi.org/10.3390/en13040790
APA StyleMunaiz, E. D., Martínez, S., Kumar, A., Caicedo, M., & Ordás, B. (2020). The Senescence (Stay-Green)—An Important Trait to Exploit Crop Residuals for Bioenergy. Energies, 13(4), 790. https://doi.org/10.3390/en13040790