Pigments Production, Growth Kinetics, and Bioenergetic Patterns in Dunaliella tertiolecta (Chlorophyta) in Response to Different Culture Media

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgal Cultivations
2.2. Analytical Methods
2.3. Growth Parameters
2.4. Photosynthetic Efficiency
2.5. Bioenergetic Parameters
2.6. Statistical Analysis
3. Results and Discussion
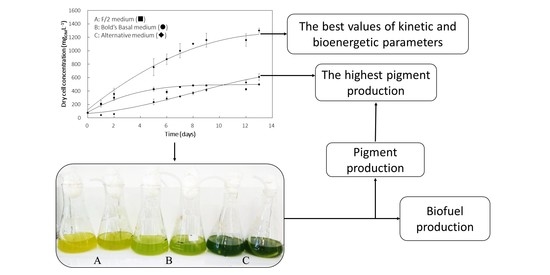
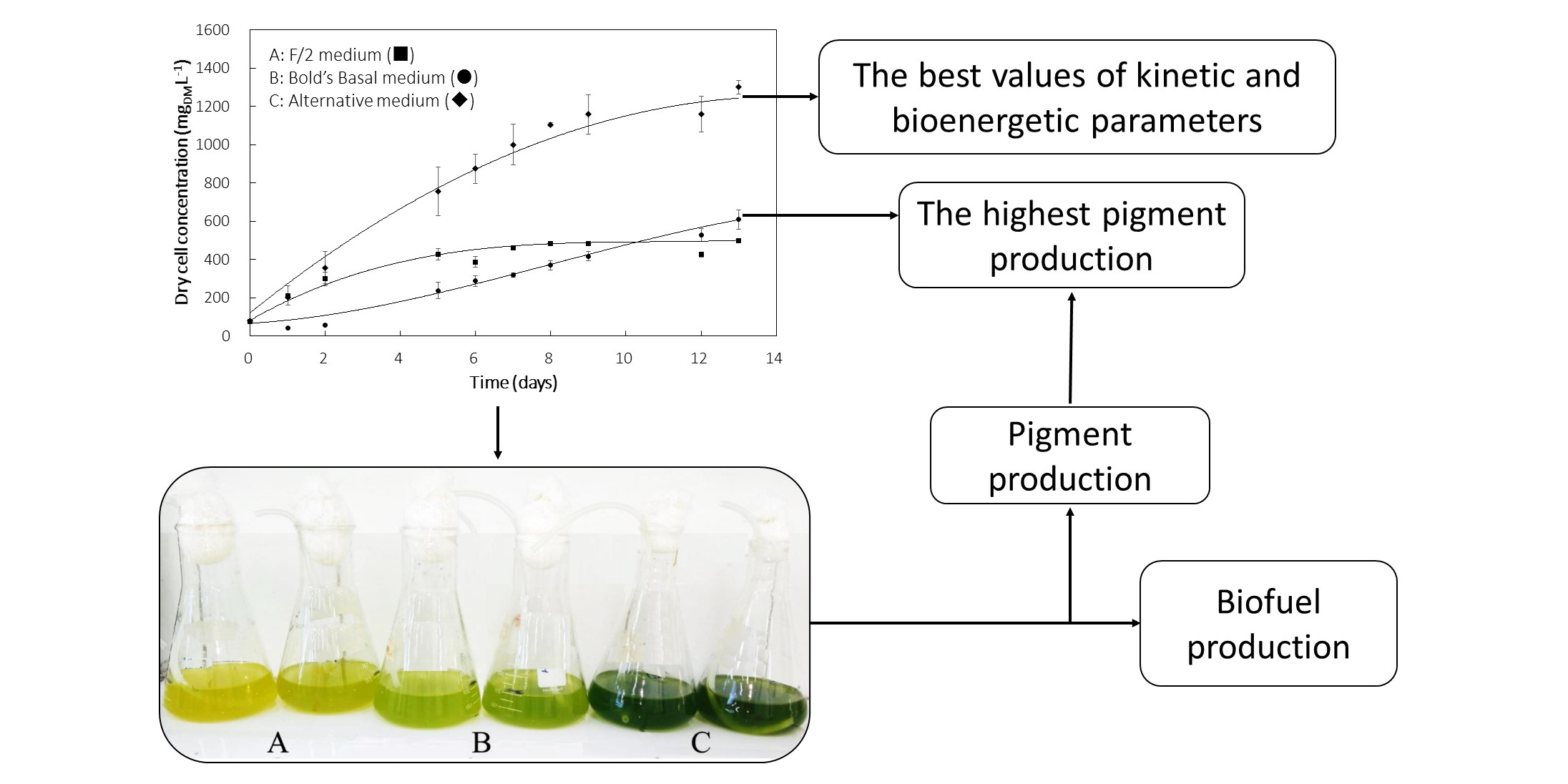
3.1. Cell Growth Profile
3.2. Kinetic Parameters
3.3. Pigments Production
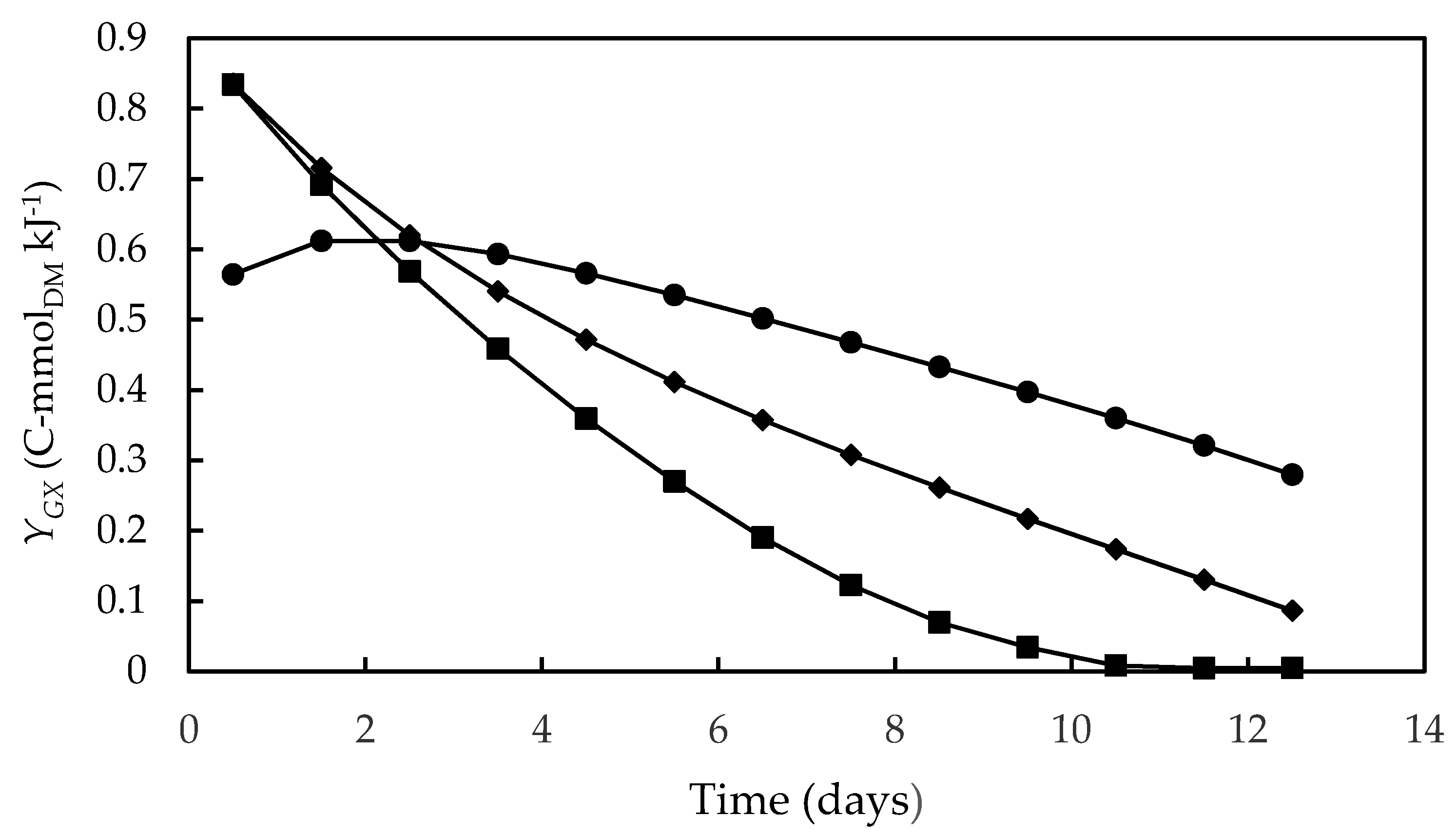
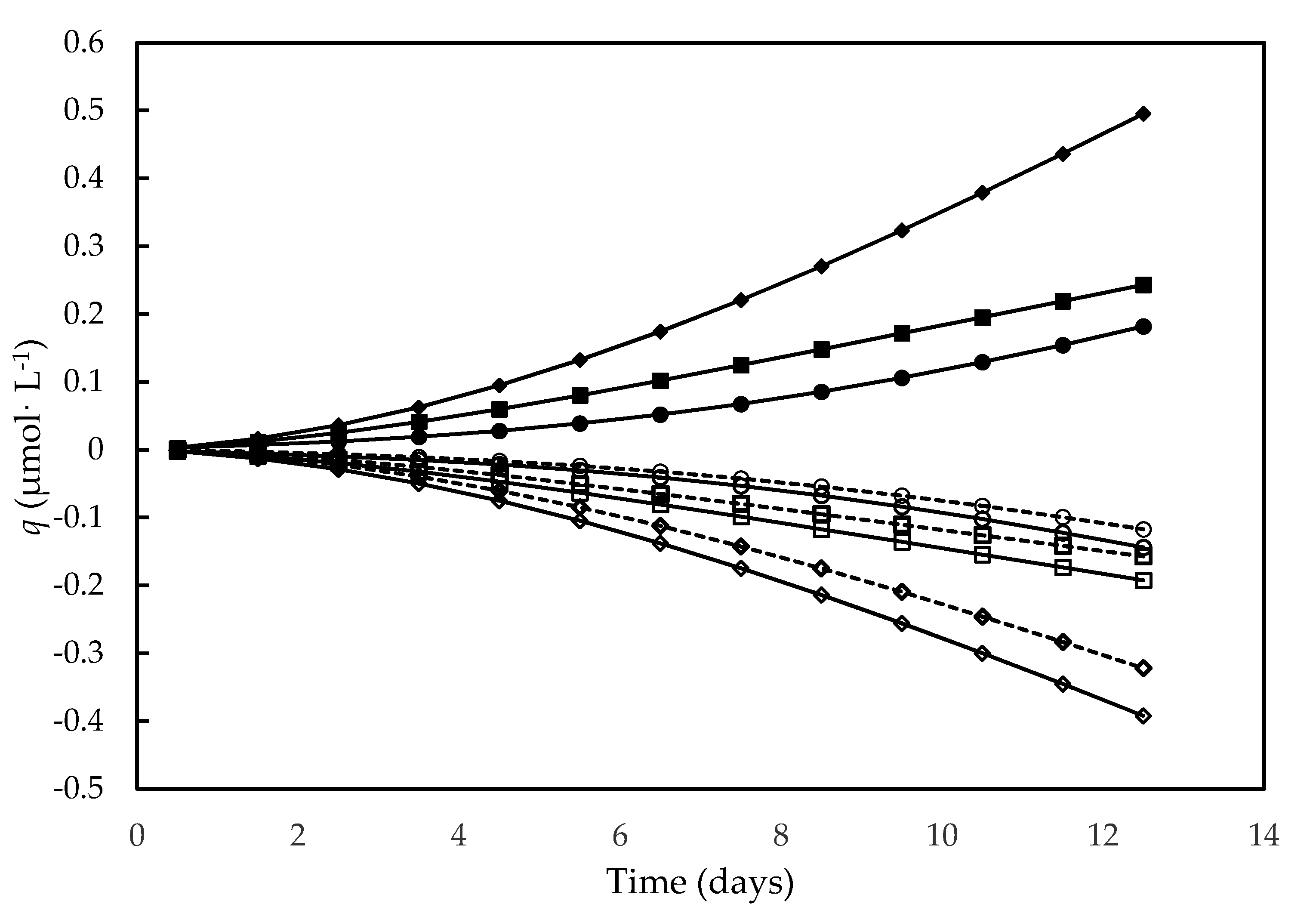
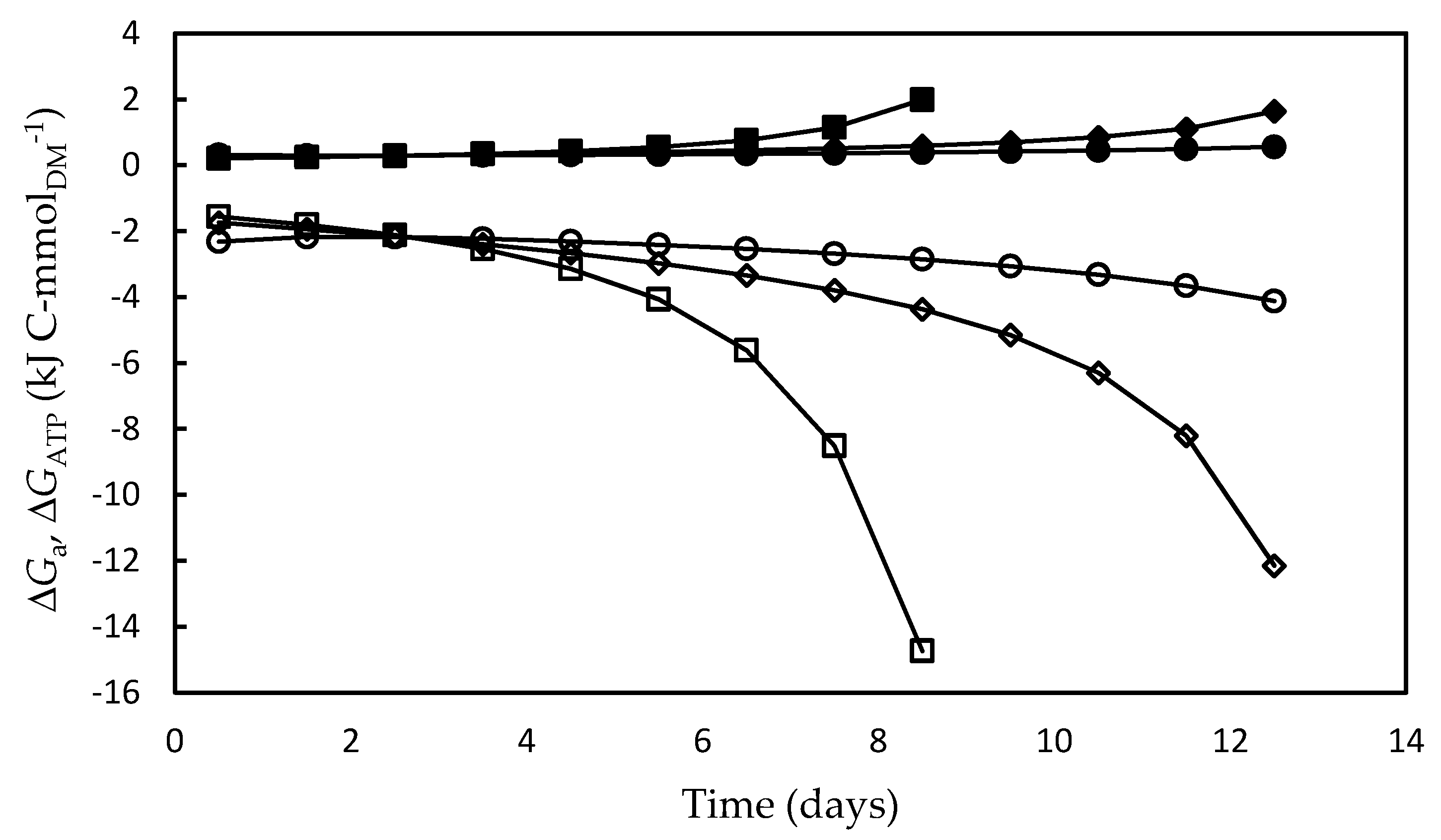
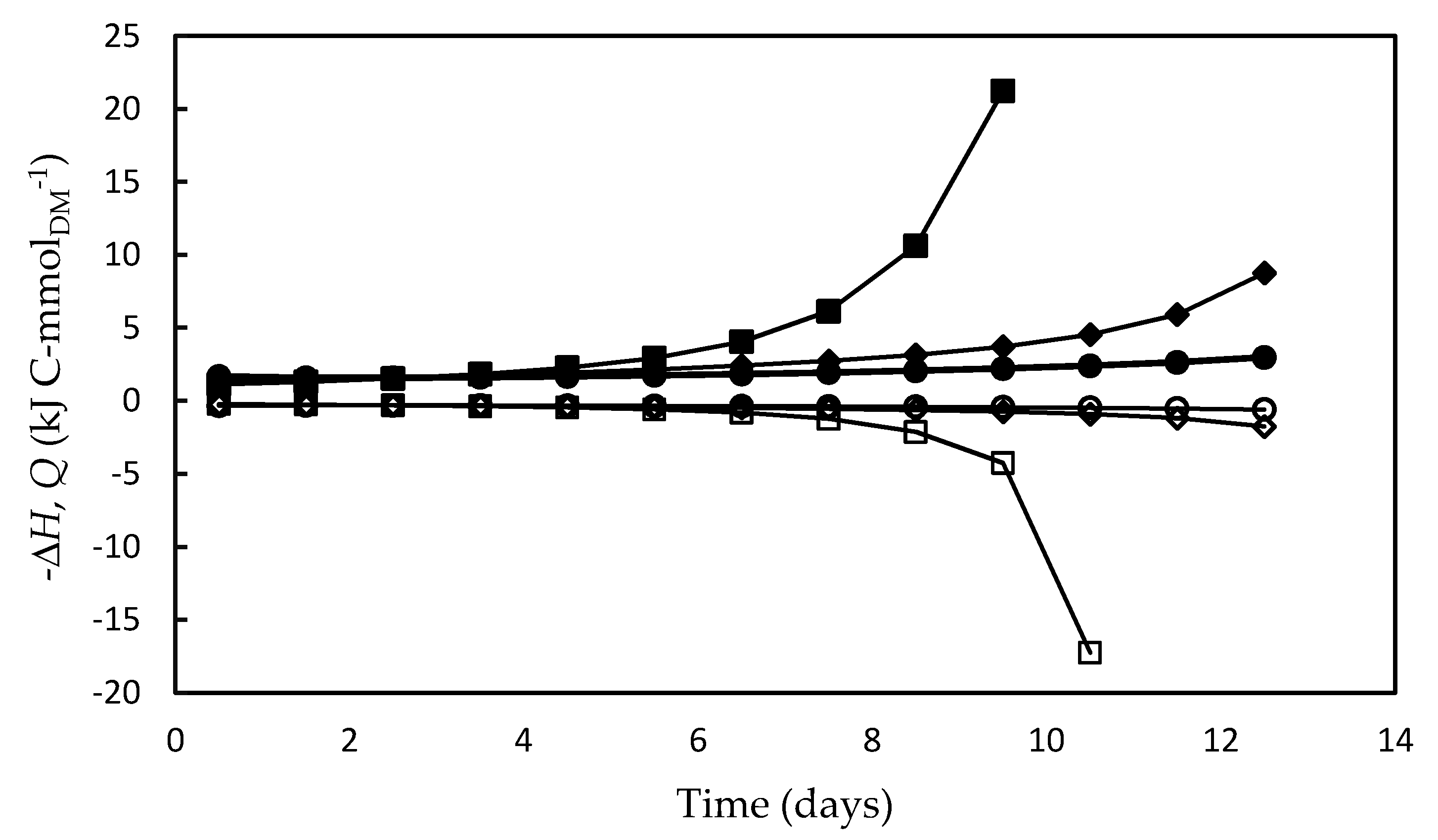
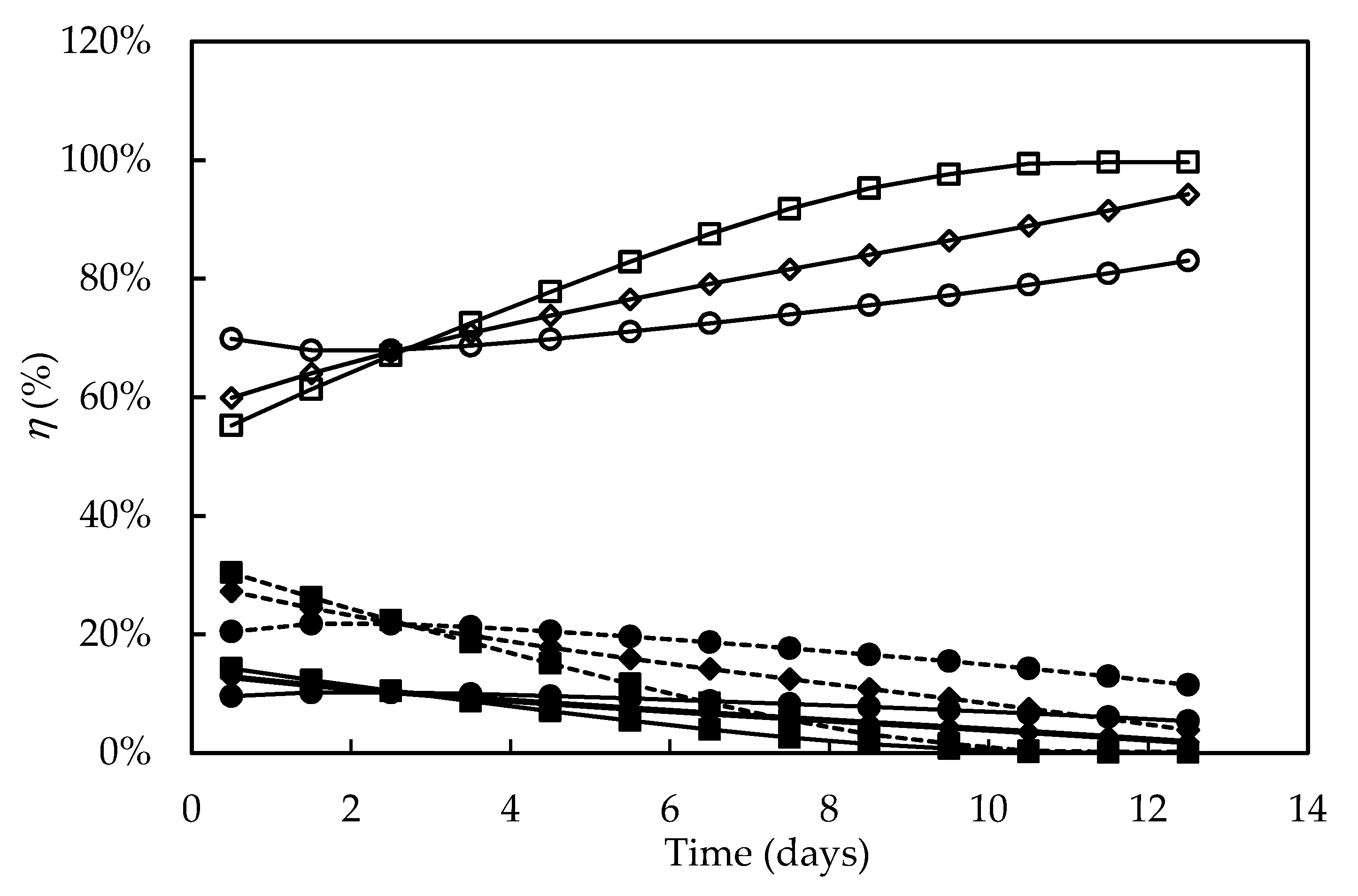
3.4. Bioenergetic Parameters
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, X.J.; Wu, M.J.; Jiang, Y.; Yang, Y.; Yan, Y.B. Dunaliella salina Hsp90 Is Halotolerant. Int. J. Biol. Macromol. 2015, 75, 418–425. [Google Scholar] [CrossRef]
- Madkour, F.F.; Abdel-Daim, M.M. Hepatoprotective and Antioxidant Activity of Dunaliella salina in Paracetamol-Induced Acute Toxicity in Rats. Indian J. Pharm. Sci. 2013, 75, 642–648. [Google Scholar]
- Srinivasan, R.; Chaitanyakumar, A.; Mageswari, A.; Gomathi, A.; Pavan Kumar, J.G.S.; Jayasindu, M.; Bharath, G.; Shravan, J.S.; Gothandam, K.M. Oral Administration of Lyophilized Dunaliella salina, a Carotenoid-Rich Marine Alga, Reduces Tumor Progression in Mammary Cancer Induced Rats. Food Funct. 2017, 8, 4517–4527. [Google Scholar] [CrossRef]
- Caroprese, M.; Albenzio, M.; Ciliberti, M.G.; Francavilla, M.; Sevi, A. A Mixture of Phytosterols from Dunaliella tertiolecta Affects Proliferation of Peripheral Blood Mononuclear Cells and Cytokine Production in Sheep. Vet. Immunol. Immunopathol. 2012, 150, 27–35. [Google Scholar] [CrossRef]
- Talebi, A.F.; Tohidfar, M.; Mousavi Derazmahalleh, S.M.; Sulaiman, A.; Baharuddin, A.S.; Tabatabaei, M. Biochemical Modulation of Lipid Pathway in Microalgae Dunaliella sp. for Biodiesel Production. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Innovative Natural Functional Ingredients from Microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Khozingoldberg, I.; Cohen, Z. The Effect of Phosphate Starvation on the Lipid and Fatty Acid Composition of the Fresh Water Eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal Triacylglycerols as Feedstocks for Biofuel Production: Perspectives and Advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Prathima Devi, M.; Venkata Mohan, S. CO2 Supplementation to Domestic Wastewater Enhances Microalgae Lipid Accumulation under Mixotrophic Microenvironment: Effect of Sparging Period and Interval. Bioresour. Technol. 2012, 112, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Tanaka, M.; Shinkawa, H.; Nakada, T.; Ano, Y.; Kurano, N.; Soga, T.; Tomita, M. Metabolic and Morphological Changes of an Oil Accumulating Trebouxiophycean Alga in Nitrogen-Deficient Conditions. Metabolomics 2013, 9, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, K.; Rohit, M.V.; Venkata Mohan, S. Microalgae-Based Carotenoids Production. Algal Green Chem. 2017, 139–147. [Google Scholar] [CrossRef]
- Khoo, K.S.; Chew, K.W.; Yew, G.Y.; Leong, W.H.; Chai, Y.H.; Show, P.L.; Chen, W.-H. Recent Advances in Downstream Processing of Microalgae Lipid Recovery for Biofuel Production. Bioresour. Technol. 2020, 304, 122996. [Google Scholar] [CrossRef] [PubMed]
- Zabed, H.M.; Akter, S.; Yun, J.; Zhang, G.; Zhang, Y.; Qi, X. Biogas from Microalgae: Technologies, Challenges and Opportunities. Renew. Sust. Energy Rev. 2020, 117, 109503. [Google Scholar] [CrossRef]
- Sun, J.; Xiong, X.; Wang, M.; Du, H.; Li, J.; Zhou, D.; Zuo, J. Microalgae Biodiesel Production in China: A Preliminary Economic Analysis. Renew. Sust. Energy Rev. 2019, 104, 296–306. [Google Scholar] [CrossRef]
- Liang, M.-H.; Qv, X.-Y.; Chen, H.; Wang, Q.; Jiang, J.-G. Effects of Salt Concentrations and Nitrogen and Phosphorus Starvations on Neutral Lipid Contents in the Green Microalga Dunaliella tertiolecta. J. Agric. Food Chem. 2017, 65, 3190–3197. [Google Scholar] [CrossRef]
- Rodríguez, M.B.R. Simulation of an Assisted Culture Medium for Production of Dunaliella tertiolecta. Algal Res. 2020, 47, 101838. [Google Scholar] [CrossRef]
- García Morales, J.; López Elías, J.A.; Medina Félix, D.; García Lagunas, N.; Fimbres Olivarría, D. Efecto Del Estrés Por Nitrógeno y Salinidad En El Contenido de β-Caroteno de La Microalga Dunaliella tertiolecta//Effect of Nitrogen and Salinity Stress on the β-Carotene Content of the Microalgae Dunaliella tertiolecta. Biotecnia 2020, 22, 13–19. [Google Scholar] [CrossRef]
- Lü, J.-M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and Molecular Mechanisms of Antioxidants: Experimental Approaches and Model Systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef]
- McWilliams, A. The Global Market for Carotenoids; BCC Research: Wellesley, MA, USA, 2018. [Google Scholar]
- Guedes, A.C.; Amaro, H.M.; Sousa-Pinto, I.; Malcata, F.X. Algal Spent Biomass—A Pool of Applications. Biofuels Algae 2019, 397–433. [Google Scholar] [CrossRef]
- Sedjati, S.; Santosa, G.; Yudiati, E.; Supriyantini, E.; Ridlo, A.; Kimberly, F. Chlorophyll and Carotenoid Content of Dunaliella salina at Various Salinity Stress and Harvesting Time. IOP Conf. Ser. Earth Environ. Sci. 2019, 246, 012025. [Google Scholar] [CrossRef]
- Tredici, M.R. Photobiology of Microalgae Mass Cultures: Understanding the Tools for the next Green Revolution. Biofuels 2010, 1, 143–162. [Google Scholar] [CrossRef]
- Küçük, K.; Tevatia, R.; Sorgüven, E.; Demirel, Y.; Özilgen, M. Bioenergetics of Growth and Lipid Production in Chlamydomonas reinhardtii. Energy 2015, 83, 503–510. [Google Scholar] [CrossRef]
- Demirel, Y.; Sieniutycz, S. Nonequilibrium Thermodynamics: Transport and Rate Processes in Physical and Biological Systems, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies on Marine Planktonic Diatoms I. Cyclotella Nana Hustedt and Detonula Confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Bischoff, H.W.; Bold, H.C. Some Soil Algae from Enchanted Rock and Related Algal Species. Phycol. Stud. IV 1963, 6318, 1–95. [Google Scholar]
- Jesus, S.S.; Maciel Filho, R. Modeling Growth of Microalgae Dunaliella salina under Different Nutritional Conditions. Am. J. Biochem. Biotechnol. 2010, 6, 279–283. [Google Scholar] [CrossRef]
- Lichtenthaler, H. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzym. 1987, 148, 350–382. [Google Scholar]
- Soletto, D.; Binaghi, L.; Ferrari, L.; Lodi, A.; Carvalho, J.C.M.; Zilli, M.; Converti, A. Effects of Carbon Dioxide Feeding Rate and Light Intensity on the Fed-Batch Pulse-Feeding Cultivation of Spirulina platensis in Helical Photobioreactor. Biochem. Eng. J. 2008, 39, 369–375. [Google Scholar] [CrossRef]
- Heijnen, J.J. Stoichiometry and Kinetics of Microbial Growth from a Thermodynamic Perspective. In Basic Biotechnology; Cambridge University Press: Cambridge, UK, 2001; pp. 45–58. [Google Scholar] [CrossRef]
- Torre, P.; Sassano, C.E.; Sato, S.; Converti, A.; Gioielli, L.A.; Carvalho, J.C. Fed-Batch Addition of Urea for Spirulina platensis Cultivation. Enzym. Microb. Technol. 2003, 33, 698–707. [Google Scholar] [CrossRef]
- Kim, S.-S.; Ly, H.V.; Kim, J.; Lee, E.Y.; Woo, H.C. Pyrolysis of Microalgae Residual Biomass Derived from Dunaliella tertiolecta after Lipid Extraction and Carbohydrate Saccharification. Chem. Eng. J. 2015, 263, 194–199. [Google Scholar] [CrossRef]
- Geider, R.; Macintyre; Graziano, L.; McKay, R.M. Responses of the Photosynthetic Apparatus of Dunaliella tertiolecta (Chlorophyceae) to Nitrogen and Phosphorus Limitation. Eur. J. Phycol. 1998, 33, 315–332. [Google Scholar] [CrossRef]
- Wongsnansilp, T.; Juntawong, N.; Wu, Z. Effects of Phosphorus on the Growth and Chlorophyll Fluorescence of a Dunaliella salina Strain Isolated from Saline Soil under Nitrate Limitation. J. Biol. Res. Boll. Soc. Ital. Biol. Sper. 2016, 89. [Google Scholar] [CrossRef]
- Kumar, A.; Guria, C.; Pathak, A.K. Optimal Cultivation towards Enhanced Algae-Biomass and Lipid Production Using Dunaliella tertiolecta for Biofuel Application and Potential CO2 Bio-Fixation: Effect of Nitrogen Deficient Fertilizer, Light Intensity, Salinity and Carbon Supply Strategy. Energy 2018. [Google Scholar] [CrossRef]
- Polle, J.E.W.; Tran, D.; Ben-Amotz, A. History, Distribution, and Habitats of Algae of the Genus Dunaliella Teodoresco (Chlorophyceae). In The Alga Dunaliella: Biodiversity, Physiology, Genomics and Biotechnology; Ben-Amotz, A., Polle, J.E.W., Rao, D.V.S., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2009; pp. 1–13. [Google Scholar]
- Kulshreshtha, J.; Singh, G.P. Evaluation of Various Inorganic Media for Growth and Biopigment of Dunaliella salina. Int. J. Pharm. Bio. Sci. 2013, 4, 1083–1089. [Google Scholar]
- Ohse, S.; Derner, R.B.; Ozório, R.Á.; Cunha, P.C.R.; Lamarca, C.P.; Santos, M.E.; Mender, L.B.B. Revisão: Sequestro de Carbono Realizado Por Microalgas e Florestas e a Capacidade de Produção de Lipídios Pelas Microalgas. Insula 2007, 36, 39–74. [Google Scholar]
- Millero, F.J.; Feistel, R.; Wright, D.G.; McDougall, T.J. The Composition of Standard Seawater and the Definition of the Reference-Composition Salinity Scale. Deep Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 50–72. [Google Scholar] [CrossRef]
- Jin, H.; Zhang, H.; Zhou, Z.; Li, K.; Hou, G.; Xu, Q.; Chuai, W.; Zhang, C.; Han, D.; Hu, Q. Ultrahigh-cell-density Heterotrophic Cultivation of the Unicellular Green Microalga Scenedesmus acuminatus and Application of the Cells to Photoautotrophic Culture Enhance Biomass and Lipid Production. Biotechnol. Bioeng. 2020, 117, 96–108. [Google Scholar] [CrossRef]
- Xinyi, E.; Crofcheck, C.; Crocker, M. Application of Recycled Media and Algae-Based Anaerobic Digestate in Scenedesmus Cultivation. J. Renew. Sust. Energy 2016, 8, 1–14. [Google Scholar] [CrossRef]
- Yeh, K.-L.; Chang, J.-S.; Chen, W. Effect of Light Supply and Carbon Source on Cell Growth and Cellular Composition of a Newly Isolated Microalga Chlorella vulgaris ESP-31. Eng. Life Sci. 2010, 10, 201–208. [Google Scholar] [CrossRef]
- Chen, M.; Tang, H.; Ma, H.; Holland, T.C.; Ng, K.Y.S.; Salley, S.O. Effect of Nutrients on Growth and Lipid Accumulation in the Green Algae Dunaliella tertiolecta. Bioresour. Technol. 2011, 102, 1649–1655. [Google Scholar] [CrossRef]
- Hexin, L.; Xianggan, C.; Zhilei, T.; Shiru, J. Analysis of Metabolic Responses of Dunaliella salina to Phosphorus Deprivation. J. Appl. Phycol. 2017, 29, 1251–1260. [Google Scholar] [CrossRef]
- Tammam, A.A.; Fakhry, E.M.; El-Sheekh, M. Effect of Salt Stress on Antioxidant System and the Metabolism of the Reactive Oxygen Species in Dunaliella salina and Dunaliella tertiolecta. Afr. J. Biotechnol. 2011, 10, 3795–3808. [Google Scholar]
- Katz, A.; Pick, U. Plasma Membrane Electron Transport Coupled to Na+ Extrusion in the Halotolerant Alga Dunaliella. Biochim. Biophys. Acta Bioenerg. 2001, 1504, 423–431. [Google Scholar] [CrossRef]
- Oren, A. A Hundred Years of Dunaliella Research: 1905–2005. Saline Syst. 2005, 1, 2. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J.-G.; Wu, G.-H. Effects of Salinity Changes on the Growth of Dunaliella salina and Its Isozyme Activities of Glycerol-3-Phosphate Dehydrogenase. J. Agric. Food Chem. 2009, 57, 6178–6182. [Google Scholar] [CrossRef]
- Jiang, Y.; Yoshida, T.; Quigg, A. Photosynthetic Performance, Lipid Production and Biomass Composition in Response to Nitrogen Limitation in Marine Microalgae. Plant Physiol. Biochem. 2012, 54, 70–77. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of Nitrogen Sources on Cell Growth and Lipid Accumulation of Green Alga Neochloris oleoabundans. Appl. Microbiol. Biotechnol. 2008, 81, 629–636. [Google Scholar] [CrossRef]
- Wykoff, D.D.; Davies, J.P.; Melis, A.; Grossman, A.R. The Regulation of Photosynthetic Electron Transport during Nutrient Deprivation in Chlamydomonas reinhardtii. Plant Physiol. 1998, 117, 129–139. [Google Scholar] [CrossRef]
- Zhang, L.; Happe, T.; Melis, A. Biochemical and Morphological Characterization of Sulfur-Deprived and H2-Producing Chlamydomonas reinhardtii (Green Alga). Planta 2002, 214, 552–561. [Google Scholar] [CrossRef]
- Volgusheva, A.A.; Zagidullin, V.E.; Antal, T.K.; Korvatovsky, B.N.; Krendeleva, T.E.; Paschenko, V.Z.; Rubin, A.B. Examination of Chlorophyll Fluorescence Decay Kinetics in Sulfur Deprived Algae Chlamydomonas reinhardtii. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 559–564. [Google Scholar] [CrossRef][Green Version]
- Srinivasan, R.; Mageswari, A.; Subramanian, P.; Suganthi, C.; Chaitanyakumar, A.; Aswini, V.; Gothandam, K.M. Bicarbonate Supplementation Enhances Growth and Biochemical Composition of Dunaliella salina V-101 by Reducing Oxidative Stress Induced during Macronutrient Deficit Conditions. Sci. Rep. 2018, 8, 6972. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, M.; Wang, C. Nutrient Deprivation Enhances Lipid Content in Marine Microalgae. Bioresour. Technol. 2013, 147, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Watanabe, Y.; Saiki, H. Photosynthetic Productivity of Conical Helical Tubular Photobioreactor Incorporatin Chlorella sorokiniana under Field Conditions. Biotechnol. Bioeng. 2002, 77, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-C.; Karam, A.L.; Sederoff, H.W.; Ducoste, J.J.; de los Reyes, F.L. Relating Nitrogen Concentration and Light Intensity to the Growth and Lipid Accumulation of Dunaliella viridis in a Photobioreactor. J. Appl. Phycol. 2019, 31, 3397–3409. [Google Scholar] [CrossRef]
- Sheehan, J.; Dunahay, T.; Benemann, J.; Roessler, P.A. Look Back at the U.S. Department of Energy’s Aquatic Species Program Biodiesel from Algae. Nat. Renew. Energy Lab. 1998, 328, 1–294. [Google Scholar]
- Antal, T.K.; Volgusheva, A.A.; Kukarskikh, G.P.; Krendeleva, T.E.; Tusov, V.B.; Rubin, A.B. Examination of Chlorophyll Fluorescence in Sulfur-Deprived Cells of Chlamydomonas reinhardtii. Biophysics 2006, 51, 251–257. [Google Scholar] [CrossRef]
- Shaker, S.; Morowvat, M.H.; Ghasemi, Y. Effects of Sulfur, Iron and Manganese Starvation on Growth, β-Carotene Production and Lipid Profile of Dunaliella salina. J. Young Pharm. 2017, 9, 43–46. [Google Scholar] [CrossRef]
- Johnson, M.K.; Johnson, E.J.; Macelroy, R.D.; Speer, H.L.; Bruff, B.S. Effects of Salts on the Halophilic Alga Dunaliella viridis. J. Bacteriol. Res. 1968, 95, 1461–1468. [Google Scholar] [CrossRef]
- Fazeli, M.R.; Tofighi, H.; Samadi, N.; Jamalifar, H. Carotenoids Accumulation by Dunaliella tertiolecta (Lake Urmia Isolate) and Dunaliella salina (CCAP 19/18 & Wt) under Stress Conditions. DARU J. Pharm. Sci. 2012, 14, 146–150. [Google Scholar]
- Ye, Z.-W.; Jiang, J.-G.; Wu, G.-H. Biosynthesis and Regulation of Carotenoids in Dunaliella: Progresses and Prospects. Biotechnol. Adv. 2008, 26, 352–360. [Google Scholar] [CrossRef]
- Ahmed, R.A.; He, M.; Aftab, R.A.; Zheng, S.; Nagi, M.; Bakri, R.; Wang, C. Bioenergy Application of Dunaliella salina SA 134 Grown at Various Salinity Levels for Lipid Production. Sci. Rep. 2017, 7, 8118. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.F.; Casazza, A.A.; Ferrari, P.F.; Perego, P.; Bezerra, R.P.; Converti, A.; Porto, A.L.F. A New Bioenergetic and Thermodynamic Approach to Batch Photoautotrophic Growth of Arthrospira (Spirulina) Platensis in Different Photobioreactors and under Different Light Conditions. Bioresour. Technol. 2016, 207, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Sassano, C.E.N.; Carvalho, J.C.M.; Gioielli, L.A.; Sato, S.; Torre, P.; Converti, A. Kinetics and Bioenergetics of Spirulina platensis Cultivation by Fed-Batch Addition of Urea as Nitrogen Source. Appl. Biochem. Biotechnol. 2004, 112, 143–150. [Google Scholar] [CrossRef]
- Stephanopoulos, G.; Aristidou, A.A.; Nielsen, J. Metabolic Engineering: Principles and Methodologies, 1st ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Bezerra, R.P.; Matsudo, M.C.; Sato, S.; Perego, P.; Converti, A.; de Carvalho, J.C.M. Effects of Photobioreactor Configuration, Nitrogen Source and Light Intensity on the Fed-Batch Cultivation of Arthrospira (Spirulina) Platensis. Bioenergetic Aspects. Biomass Bioenerg 2012, 37, 309–317. [Google Scholar] [CrossRef]
- Richmond, A. Phototrophic Microalgae. In Biotechnology; Rehm, H.J., Reed, G., Dellweg, H., Eds.; Verlag Chemie: Weinheim, Germany, 1983; Volume 3, pp. 109–143. [Google Scholar]
- Yang, C.; Hua, Q.; Shimizu, K. Energetics and Carbon Metabolism during Growth of Microalgal Cells under Photoautotrophic, Mixotrophic and Cyclic Light-Autotrophic/Dark-Heterotrophic Conditions. Biochem. Eng. J. 2000, 6, 87–102. [Google Scholar] [CrossRef]
- Escoubas, J.M.; Lomas, M.; LaRoche, J.; Falkowski, P.G. Light Intensity Regulation of Cab Gene Transcription Is Signaled by the Redox State of the Plastoquinone Pool. Proc. Natl. Acad. Sci. USA 1995, 92, 10237–10241. [Google Scholar] [CrossRef]
- Mukhanov, V.S.; Kemp, R.B. Simultaneous Photocalorimetric and Oxygen Polarographic Measurements on Dunaliella Maritima Cells Reveal a Thermal Discrepancy That Could Be Due to Nonphotochemical Quenching. Thermochim. Acta 2006, 446, 11–19. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. The Wavelength Dependence Of Massive Carotene Synthesis In Dunaliella Bardawil (Chlorophyceae)1. J. Phycol. 1989, 25, 175–178. [Google Scholar] [CrossRef]
- Binaghi, L.; Del Borghi, A.; Lodi, A.; Converti, A.; Del Borghi, M. Batch and Fed-Batch Uptake of Carbon Dioxide by Spirulina platensis. Process Biochem. 2003, 38, 1341–1346. [Google Scholar] [CrossRef]
- Sakurai, H.; Masukawa, H.; Kitashima, M.; Inoue, K. Photobiological Hydrogen Production: Bioenergetics and Challenges for Its Practical Application. J. Photochem. Photobiol. C Photochem. Rev. 2013, 17, 1–25. [Google Scholar] [CrossRef]
- Kayser, A.; Weber, J.; Hecht, V.; Rinas, U. Metabolic Flux Analysis of Escherichia coli in Glucose-Limited Continuous Culture. I. Growth-Rate-Dependent Metabolic Efficiency at Steady State. Microbiology 2005, 151, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Taymaz-Nikerel, H.; Borujeni, A.E.; Verheijen, P.J.T.; Heijnen, J.J.; van Gulik, W.M. Genome-Derived Minimal Metabolic Models for Escherichia coli MG1655 with Estimated in Vivo Respiratory ATP Stoichiometry. Biotechnol. Bioeng. 2010, 107, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Kliphuis, A.M.J.; Klok, A.J.; Martens, D.E.; Lamers, P.P.; Janssen, M.; Wijffels, R.H. Metabolic Modeling of Chlamydomonas reinhardtii: Energy Requirements for Photoautotrophic Growth and Maintenance. J. Appl. Phycol. 2012, 24, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, L.; Liu, J.; Qin, R. Light Absorption and Growth Response of Dunaliella under Different Light Qualities. J. Appl. Phycol. 2020, 32, 1041–1052. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salts | Concentration (mM) |
|---|---|
| NaCl | 470 |
| MgCl2 | 420 |
| MgSO4 | 640 |
| CaCl2 | 280 |
| KCl | 8 |
| NaHCO3 | 580 |
| Constituent | Medium | ||
|---|---|---|---|
| A (mM) a | B (mM) b | C (mM) a | |
| NaNO3 | 0.8823 | 2.9410 | 2.9410 |
| MgSO4·7H2O | - | 0.6231 | 0.6231 |
| NaCl | - | 0.4277 | 0.4277 |
| K2HPO4 | - | 0.4305 | 0.4305 |
| KH2PO4 | - | 1.2860 | 1.286 |
| CaCl2·2H2O | - | 0.1702 | 0.1702 |
| ZnSO4·7H2O | 0.0765 | 0.0306 | 0.0306 |
| MnCl2·4H2O | 0.9098 | 0.0072 | 0.0072 |
| MoO3 | - | 0.0047 | 0.0047 |
| CuSO4·5H2O | 0.0392 | 0.0062 | 0.0062 |
| Co(NO3)2·6H2O | - | 0.0016 | 0.0016 |
| H3BO3 | - | 0.1846 | 0.1846 |
| EDTA | - | 0.1710 | 0.1710 |
| KOH | - | 0.5525 | 0.5525 |
| FeSO4·7H2O | - | 0.0125 | 0.0125 |
| H2SO4·(conc) | - | 0.0071 | 0.0071 |
| NaH2PO4·H2O | 0.0362 | - | - |
| Na2SiO3·9H2O | 0.1055 | - | - |
| FeCl3·6H2O | 0.0116 | - | - |
| Na2EDTA·2H2O | 0.0116 | - | - |
| Na2MoO4·2H2O | 0.0240 | - | - |
| CoCl2·6H2O | 0.0770 | - | - |
| Thiamine HCl (vit. B1) | 0.0003 | - | - |
| Biotin (vit. H) | 0.0020 | - | - |
| Cyanocobalamin (vit. B12) | 0.0003 | - | - |
| Medium | Xmax (mgDM·L−1) | PX (mgDM·L−1·d−1) | µmax (d−1) | PEmax (%) | Carotenoids (mg·g −1) | Chlorophyll (mg·g−1) |
|---|---|---|---|---|---|---|
| F/2 | 452 ± 28 a | 75 | 0.500 | 9.14 | 18.2 ± 2.1 a | 65.7 ± 4.8 a |
| Bold’s Basal | 567± 33 b | 47 | 0.269 | 5.13 | 52.0 ± 7.2 b | 162.6 ± 15.7 b |
| Alternative | 1223 ± 91 c | 102 | 0.535 | 14.54 | 33.4 ± 5.7 c | 108.0 ± 11.3 c |
| Compound | HCO3− | NO3− | H2O | O2 | H+ | Biomass |
|---|---|---|---|---|---|---|
| Δgf′i (kJ·mol−1) | −587.2 | −111.4 | −273.3 | 0 | −39.9 | −67.0 |
| Stoichiometric Coefficients | ||||||
| Symbol | a | b | c | d | e | γX |
| Value (mol·C-molDM−1) | −1 | −0.24 | 1.0 | 1.52 | −1.20 | 6.06 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, Y.A.S.; Viana-Marques, D.d.A.; Porto, A.L.F.; Bezerra, R.P.; Converti, A. Pigments Production, Growth Kinetics, and Bioenergetic Patterns in Dunaliella tertiolecta (Chlorophyta) in Response to Different Culture Media. Energies 2020, 13, 5347. https://doi.org/10.3390/en13205347
Moura YAS, Viana-Marques DdA, Porto ALF, Bezerra RP, Converti A. Pigments Production, Growth Kinetics, and Bioenergetic Patterns in Dunaliella tertiolecta (Chlorophyta) in Response to Different Culture Media. Energies. 2020; 13(20):5347. https://doi.org/10.3390/en13205347
Chicago/Turabian StyleMoura, Yanara Alessandra Santana, Daniela de Araújo Viana-Marques, Ana Lúcia Figueiredo Porto, Raquel Pedrosa Bezerra, and Attilio Converti. 2020. "Pigments Production, Growth Kinetics, and Bioenergetic Patterns in Dunaliella tertiolecta (Chlorophyta) in Response to Different Culture Media" Energies 13, no. 20: 5347. https://doi.org/10.3390/en13205347
APA StyleMoura, Y. A. S., Viana-Marques, D. d. A., Porto, A. L. F., Bezerra, R. P., & Converti, A. (2020). Pigments Production, Growth Kinetics, and Bioenergetic Patterns in Dunaliella tertiolecta (Chlorophyta) in Response to Different Culture Media. Energies, 13(20), 5347. https://doi.org/10.3390/en13205347