Polarization Potential Has No Effect on Maximum Current Density Produced by Halotolerant Bioanodes


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Inoculum and Medium
2.2. Electrochemical Set-Up
2.3. Bioanode Growth and Electrochemical Characterization
2.4. Statistical Analysis
3. Results
3.1. Inoculum and Medium Characterization
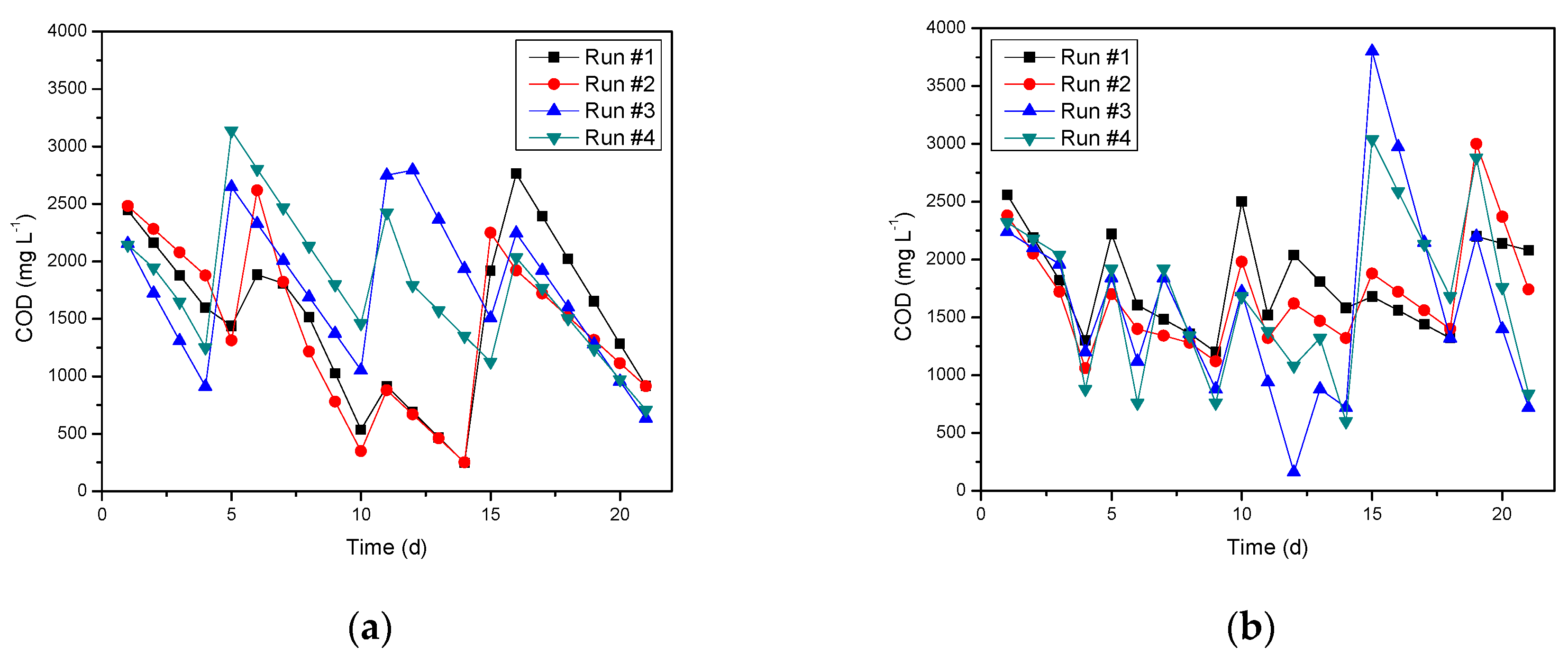
3.2. Bioanode Growth and Current Production
3.3. Maximum Current Density by Linear Voltammetry
4. Discussion
4.1. Overall Energetic Performance
4.2. Implications of Polarization Potential for Current Density
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Logan, B.E.; Rabaey, K. Conversion of Wastes into Bioelectricity and Chemicals by Using Microbial Electrochemical Technologies. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Irini, A. Microbial Electrolysis Cells Turning to Be Versatile Technology: Recent Advances and Future Challenges. Water Res. 2014, 56, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Liu, G.; Zhang, R.; Bai, Y.; Fu, S.; Hou, Y. Heavy Metal Recovery Combined with H(2) Production from Artificial Acid Mine Drainage Using the Microbial Electrolysis Cell. J. Hazard. Mater. 2014, 270, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, Z.J. Bioelectrochemical Metal Recovery from Wastewater: A Review. Water Res. 2014, 66, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Benetton, X.; Chandrakant-Varia, J.; Pozo, G.; Modin, O.; Ter Heijne, A.; Fransaer, J.; Rabbaey, K. Metal Recovery by Microbual Electro-Metallurgy. Prog. Mater. Sci. 2018, 94, 435–461. [Google Scholar] [CrossRef]
- Dominguez-Benetton, X.; Sevda, S.; Vanbroekhoven, K.; Pant, D. The Accurate Use of Impedance Analysis for the Study of Microbial Electrochemical Systems. Chem. Soc. Rev. 2012, 41, 7228–7246. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, R.; Dominguez-Benetton, X.; Délia, M.L.; Bergel, A. Microbial Bioanodes with High Salinity Tolerance for Microbial Fuel Cells and Microbial Electrolysis Cells. Electrochem. Commun. 2013, 33, 1–4. [Google Scholar] [CrossRef]
- Rousseau, R.; Santaella, C.; Bonnafous, A.; Achouak, W.; Godon, J.J.; Delia, M.L.; Bergel, A. Halotolerant Bioanodes: The Applied Potential Modulates the Electrochemical Characteristics, the Biofilm Structure and the Ratio of the Two Dominant Genera. Bioelectrochemistry 2016, 112, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, R.; Da Silva, S.; Gaig, M.V.; Rousseau, R.; Delia, M.L.; Bergel, A. Modelling Potential/Current Distribution in Microbial Electrochemical Systems Shows How the Optimal Bioanode Architecture Depends on Electrolyte Conductivity. Phys. Chem. Chem. Phys. 2014, 16, 22892–22902. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Benetton, X.; Godon, J.J.; Rousseau, R.; Erable, B.; Bergel, A.; Délia, M.L. Exploring Natural vs. Synthetic Minimal Media to Boost Current Generation with Electrochemically-Active Marine Bioanodes. J. Environ. Chem. Eng. 2016, 4, 2362–2369. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, X.; Logan, B.E.; Lee, H. Brewery Wastewater Treatment Using Air-Cathode Microbial Fuel Cells. Appl. Microbiol. Biotechnol. 2008, 78, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, O.; Quentin, S.; Torrijos, M.; Godon, J.J.; Delgenes, J.P.; Moletta, R. Impact of Increasing NaCl Concentrations on the Performance and Community Composition of Two Anaerobic Reactors. Appl. Microbiol. Biotechnol. 2007, 75, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, O.; Tan, Z.; Kharkwal, S.; Ng, H.Y. Effect of Increasing Anodic NaCl Concentration on Microbial Fuel Cell Performance. Bioresour. Technol. 2012, 112, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cheng, S.; Logan, B.E. Power Generation in Fed-Batch Microbial Fuel Cells as a Function of Ionic Strength, Temperature, and Reactor Configuration. Environ. Sci. Technol. 2005, 39, 5488–5493. [Google Scholar] [CrossRef] [PubMed]
- De Schamphelaire, L.; Rabaey, K.; Boeckx, P.; Boon, N.; Verstraete, W. Outlook for Benefits of Sediment Microbial Fuel Cells with Two Bio-Electrodes. Microbial. Biotechnol. 2008, 1, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhao, Q.; Wu, M.; Ding, J.; Zhang, W. Biodegradation of Organic Matter and Anodic Microbial Communities Analysis in Sediment Microbial Fuel Cells with/without Fe(III) Oxide Addition. Bioresour. Technol. 2017, 225, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Li, R.; Ji, M.; Ren, Z.J. Organic Content Influences Sediment Microbial Fuel Cell Performance and Community Structure. Bioresour. Technol. 2016, 220, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Erable, B.; Bergel, A. First Air-Tolerant Effective Stainless Steel Microbial Anode Obtained from a Natural Marine Biofilm. Bioresour. Technol. 2009, 100, 3302–3307. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Monzon, O.; Yang, Y.; Yu, C.; Li, Q.; Alvarez, P.J.J. Microbial Fuel Cells under Extreme Salinity: Performance and Microbial Analysis. Environ. Chem. 2014, 12, 293–299. [Google Scholar] [CrossRef]
- Oren, A. Microbial Life at High Salt Concentrations: Phylogenetic and Metabolic Diversity. Saline Syst. 2008, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Life at High Salt Concentrations. In The Prokaryotes: Prokaryotic Communities and Ecophysiology; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 421–440. [Google Scholar]
- Weyl, P.K. On the Change in Electrical Conductance of Seawater with Temperature 1. Limnol. Oceanogr. 1964, 9, 75–78. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P. Halophiles. In eLS; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar]
- Pol, L.W.; Lens, P.N.; Weijma, J.; Stams, A.J. New Developments in Reactor and Process Technology for Sulfate Reduction. Water Sci. Technol. 2001, 44, 67–76. [Google Scholar] [PubMed]
- Santoro, C.; Arbizzani, C.; Erable, B.; Ieropoulos, I. Microbial Fuel Cells: From Fundamentals to Applications. A Review. J. Power Sources 2017, 356, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Schroder, U. Anodic Electron Transfer Mechanisms in Microbial Fuel Cells and Their Energy Efficiency. Phys. Chem. Chem. Phys. 2007, 9, 2619–2629. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, R.; Santaella, C.; Achouak, W.; Godon, J.J.; Bonnafous, A.; Bergel, A.; Délia, M.L. Correlation of the Electrochemical Kinetics of High-Salinity-Tolerant Bioanodes with the Structure and Microbial Composition of the Biofilm. ChemElectroChem 2014, 1, 1966–1975. [Google Scholar] [CrossRef]
- Abrevaya, X.C.; Sacco, N.; Mauas, P.J.D.; Cortón, E. Archaea-Based Microbial Fuel Cell Operating at High Ionic Strength Conditions. Extremophiles 2011, 15, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.E.; Yung, P.Y.; Mitra, S.D.; Wuertz, S.; Williams, R.B.H.; Lauro, F.M.; Marsili, E. Electrochemical and Genomic Analysis of Novel Electroactive Isolates Obtained Via Potentiostatic Enrichment from Tropical Sediment. J. Power Sources 2017, 356, 539–548. [Google Scholar] [CrossRef]
- Grattieri, M.; Suvira, M.; Hasan, K.; Minteer, S.D. Halotolerant Extremophile Bacteria from the Great Salt Lake for Recycling Pollutants in Microbial Fuel Cells. J. Power Sources 2017, 356, 310–318. [Google Scholar] [CrossRef]
- Holmes, D.E.; Bond, D.R.; O’Neil, R.A.; Reimers, C.E.; Tender, L.R.; Lovley, D.R. Microbial Communities Associated with Electrodes Harvesting Electricity from a Variety of Aquatic Sediments. Microb. Ecol. 2004, 48, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Monzon, O.; Yang, Y.; Kim, J.; Heldenbrand, A.; Li, Q.; Alvarez, P.J.J. Microbial Fuel Cell Fed by Barnett Shale Produced Water: Power Production by Hypersaline Autochthonous Bacteria and Coupling to a Desalination Unit. Biochem. Eng. J. 2017, 117, 87–91. [Google Scholar] [CrossRef]
- Finkelstein, D.A.; Tender, L.M.; Zeikus, J.G. Effect of Electrode Potential on Electrode-Reducing Microbiota. Environ. Sci. Technol. 2006, 40, 6990–6995. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Cheng, S.; Zhang, F.; Logan, B.E. Current Density Reversibly Alters Metabolic Spatial Structure of Exoelectrogenic Anode Biofilms. J. Power Sources 2017, 356, 566–571. [Google Scholar] [CrossRef]
- Lewis, A.J.; Borole, A.P. Adapting Microbial Communities to Low Anode Potentials Improves Performance of Mecs at Negative Potentials. Electrochim. Acta 2017, 254, 79–88. [Google Scholar] [CrossRef]
- Torres, C.I.; Krajmalnik-Brown, R.; Parameswaran, P.; Marcus, A.K.; Wanger, G.; Gorby, Y.A.; Rittmann, B.E. Selecting Anode-Respiring Bacteria Based on Anode Potential: Phylogenetic, Electrochemical, and Microscopic Characterization. Environ. Sci. Technol. 2009, 43, 9519–9524. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yates, M.D.; Hatzell, M.C.; Rao, H.A.; Saikaly, P.E.; Logan, B.E. Microbial Community Composition Is Unaffected by Anode Potential. Environ. Sci. Technol. 2014, 48, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Commault, A.S.; Lear, G.; Packer, M.A.; Weld, R.J. Influence of Anode Potentials on Selection of Geobacter Strains in Microbial Electrolysis Cells. Bioresour. Technol. 2013, 139, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.C.; Call, D.F.; Logan, B.E. Optimal Set Anode Potentials Vary in Bioelectrochemical Systems. Environ. Sci. Technol. 2010, 44, 6036–6041. [Google Scholar] [CrossRef] [PubMed]
- Babauta, J.; Renslow, R.; Lewandowski, Z.; Beyenal, H. Electrochemically active biofilms: Facts and fiction. A review. Biofouling 2012, 28, 789–812. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association (APHA). Standard Methods of Water and Wastewater; American Public Health Association; American Water Works Association; Water Environment Federation Publication: Washington, DC, USA, 1996. [Google Scholar]
- Rousseau, R.; Rimboud, M.; Délia, M.L.; Bergel, A.; Basséguy, R. Electrochemical Characterization of Microbial Bioanodes Formed on a Collector/Electrode System in a Highly Saline Electrolyte. Bioelectrochemistry 2015, 106, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.S. Techniques for Cultivating Methanogens. Methods Enzymol. 2011, 494, 1–22. [Google Scholar] [PubMed]
- Erable, B.; Roncato, M.A.; Achouak, W.; Bergel, A. Sampling Natural Biofilms: A New Route to Build Efficient Microbial Anodes. Environ. Sci. Technol. 2009, 43, 3194–3199. [Google Scholar] [CrossRef] [PubMed]
- White, H.K.; Reimers, C.E.; Cordes, E.E.; Dilly, G.F.; Girguis, P.R. Quantitative population dynamics of microbial communities in plankton-fed microbial fuel cells. ISME J. 2009, 3, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Microbial Fuel Cells: Methodology and Technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E. Microbial Fuel Cells; Wiley-Interscience: Hoboken, NJ, USA, 2008. [Google Scholar]
- Wang, X.; Feng, Y.; Ren, N.; Wang, H.; Lee, H.; Li, N.; Zhao, Q. Accelerated Start-up of Two-Chambered Microbial Fuel Cells: Effect of Anodic Positive Poised Potential. Electrochim. Acta 2009, 54, 1109–1114. [Google Scholar] [CrossRef]
- Heijne, A.T.; Liu, F.; Weijden, R.; Weijma, J.; Buisman, C.J.; Hamelers, H.V. Copper Recovery Combined with Electricity Production in a Microbial Fuel Cell. Environ. Sci. Technol. 2010, 44, 4376–4381. [Google Scholar] [CrossRef] [PubMed]
- Libes, S. Introduction to Marine Biogeochemistry, 2nd ed.; John Willey and Sons: Hoboken, NJ, USA, 2009; 1715p, ISBN 9780120885305. [Google Scholar]
- Valdes, D.; Real, E. Nitrogen and Phosphorus in Water and Sediments at Ria Lagartos Coastal Lagoon, Yucatan, Gulf of Mexico. Indian J. Mar. Sci. 2004, 33, 338–345. [Google Scholar]
- Rabaey, K.; Van de Sompel, K.; Maignien, L.; Boon, N.; Aelterman, P.; Clauwaert, P.; De Schamphelaire, L.; Pham, H.T.; Vermeulen, J.; Verhaege, M.; et al. Microbial Fuel Cells for Sulfide Removal. Environ. Sci. Technol. 2006, 40, 5218–5224. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Microbial Fuel Cells: Novel Microbial Physiologies and Engineering Approaches. Curr. Opin. Biotechnol. 2006, 17, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; Stams, A.J. The Ecology and Biotechnology of Sulphate-Reducing Bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Bug Juice: Harvesting Electricity with Microorganisms. Nat. Rev. Microbiol. 2006, 4, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yates, M.D.; Hatzell, M.C.; Rao, H.A.; Saikaly, P.E.; Logan, B.E. Response to Comment on Microbial Community Composition Is Unaffected by Anode Potential. Environ. Sci. Technol. 2014, 48, 14853–14854. [Google Scholar] [CrossRef] [PubMed]
- Ichihashi, O.; Vishnivetskaya, T.A.; Borole, A.P. High-Performance Bioanode Development for Fermentable Substrates via Controlled Electroactive Biofilm Growth. ChemElectroChem 2014, 1, 1940–1947. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Polarization Potential | Maximum Current Density (A m−2) | ||||
|---|---|---|---|---|---|
| Run #1 | Run #2 | Run #3 | Run #4 | Mean | |
| +0.34 V/SHE | 2.64 (day 19) | 1.17 (day 16) | 1.58 (day 16) | 1.29 (day 16) | 1.67 ± 0.67a |
| −0.16 V/SHE | 1.82 (day 21) | 1.53 (day 20) | 1.07 (day 20) | 2.45 (day 16) | 1.72 ± 0.58a |
| Polarization Potential | Maximum Power Density (W m−2) | ||||
|---|---|---|---|---|---|
| Run #1 | Run #2 | Run #3 | Run #4 | Mean | |
| +0.34 V/SHE | 1.39 (day 19) | 0.62 (day 16) | 0.77 (day 16) | 0.64 (day 16) | 0.85 ± 0.36a |
| −0.16 V/SHE | 0.88 (day 21) | 0.77 (day 20) | 0.55 (day 20) | 1.27 (day 16) | 0.88 ± 0.30a |
| Polarization Potential | Maximum Current Density (A m−2) | ||||
|---|---|---|---|---|---|
| Run #1 | Run #2 | Run #3 | Run #4 | Mean | |
| +0.34 V/SHE | 10.12 | 4.80 | 3.00 | 3.17 | 5.27 ± 3.38a |
| −0.16 V/SHE | 5.54 | 4.32 | 3.18 | 6.91 | 4.99 ± 1.60a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Muñoz, M.; Dominguez-Benetton, X.; Domínguez-Maldonado, J.; Valdés-Lozano, D.; Pacheco-Catalán, D.; Ortega-Morales, O.; Alzate-Gaviria, L. Polarization Potential Has No Effect on Maximum Current Density Produced by Halotolerant Bioanodes. Energies 2018, 11, 529. https://doi.org/10.3390/en11030529
González-Muñoz M, Dominguez-Benetton X, Domínguez-Maldonado J, Valdés-Lozano D, Pacheco-Catalán D, Ortega-Morales O, Alzate-Gaviria L. Polarization Potential Has No Effect on Maximum Current Density Produced by Halotolerant Bioanodes. Energies. 2018; 11(3):529. https://doi.org/10.3390/en11030529
Chicago/Turabian StyleGonzález-Muñoz, Muriel, Xochitl Dominguez-Benetton, Jorge Domínguez-Maldonado, David Valdés-Lozano, Daniella Pacheco-Catalán, Otto Ortega-Morales, and Liliana Alzate-Gaviria. 2018. "Polarization Potential Has No Effect on Maximum Current Density Produced by Halotolerant Bioanodes" Energies 11, no. 3: 529. https://doi.org/10.3390/en11030529
APA StyleGonzález-Muñoz, M., Dominguez-Benetton, X., Domínguez-Maldonado, J., Valdés-Lozano, D., Pacheco-Catalán, D., Ortega-Morales, O., & Alzate-Gaviria, L. (2018). Polarization Potential Has No Effect on Maximum Current Density Produced by Halotolerant Bioanodes. Energies, 11(3), 529. https://doi.org/10.3390/en11030529