

Ameliorative Effects of Oyster Protein Hydrolysates on Cadmium-Induced Hepatic Injury in Mice

Abstract
1. Introduction
2. Results
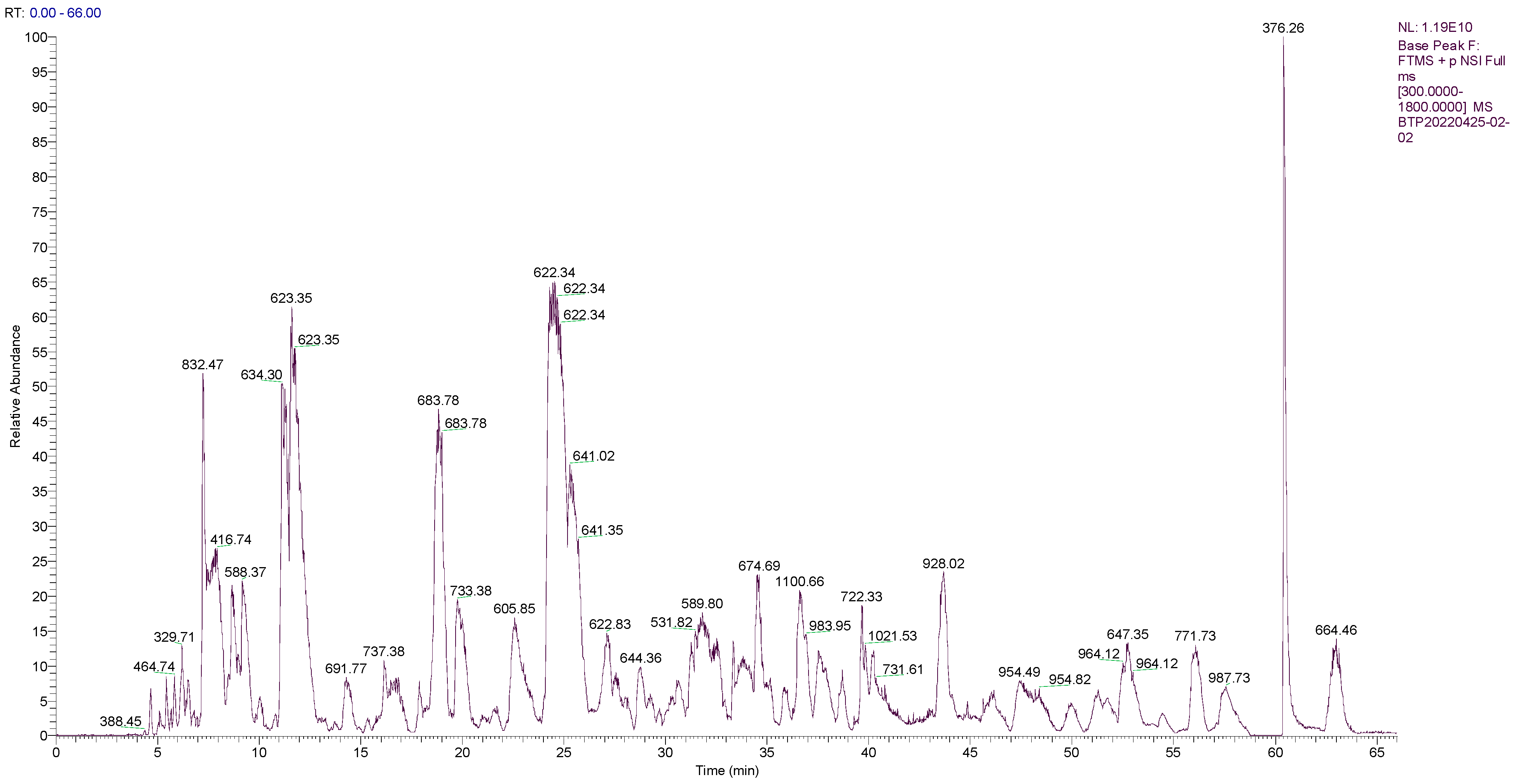
2.1. Sequence Analysis of the Main Peptides of OPs
2.2. Composition of Amino Acid in OPs
2.3. Contents of Free Amino Acids in OPs
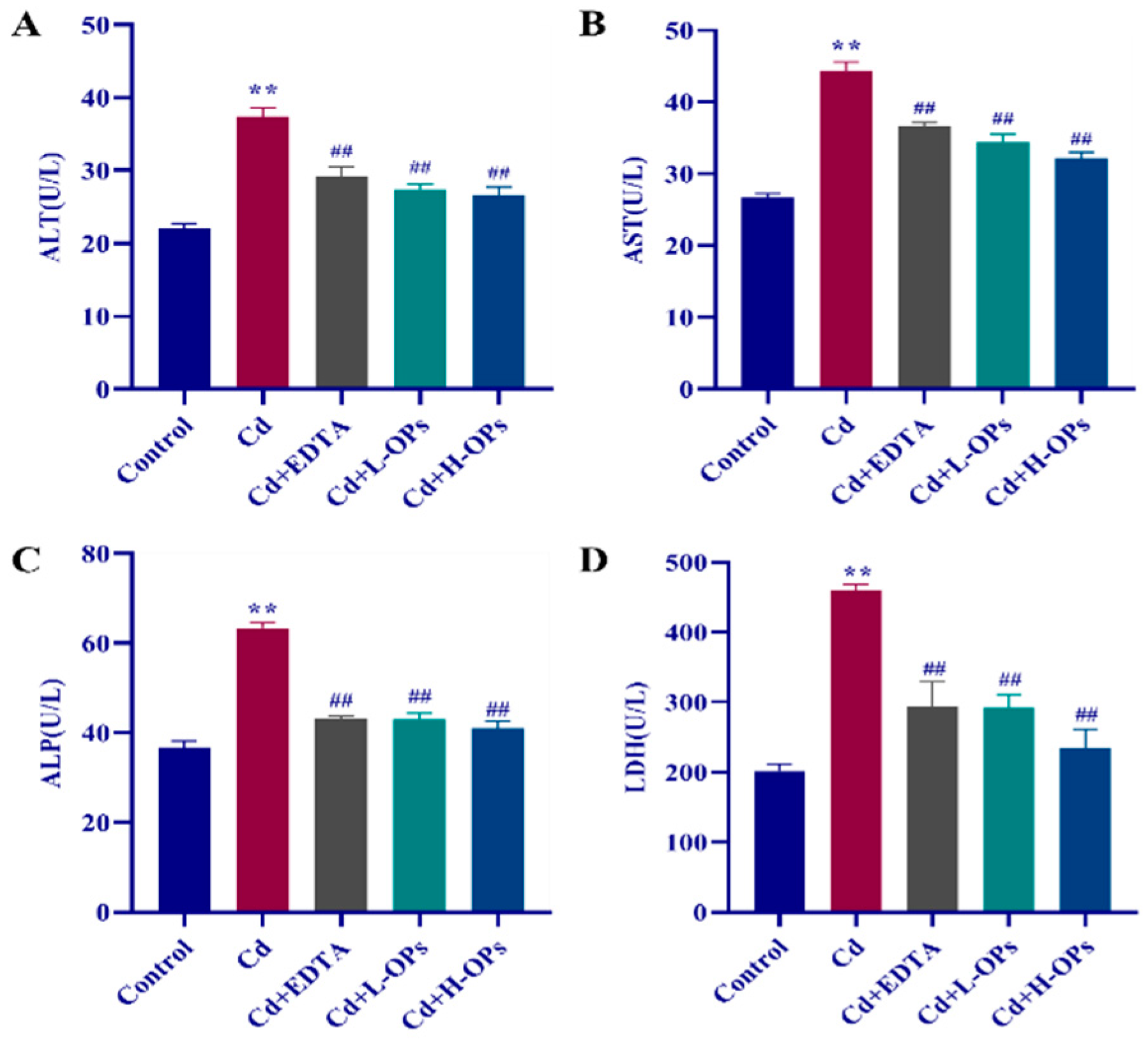
2.4. Effects of OPs on Hepatic Dysfunction in Cd-Exposed Mice
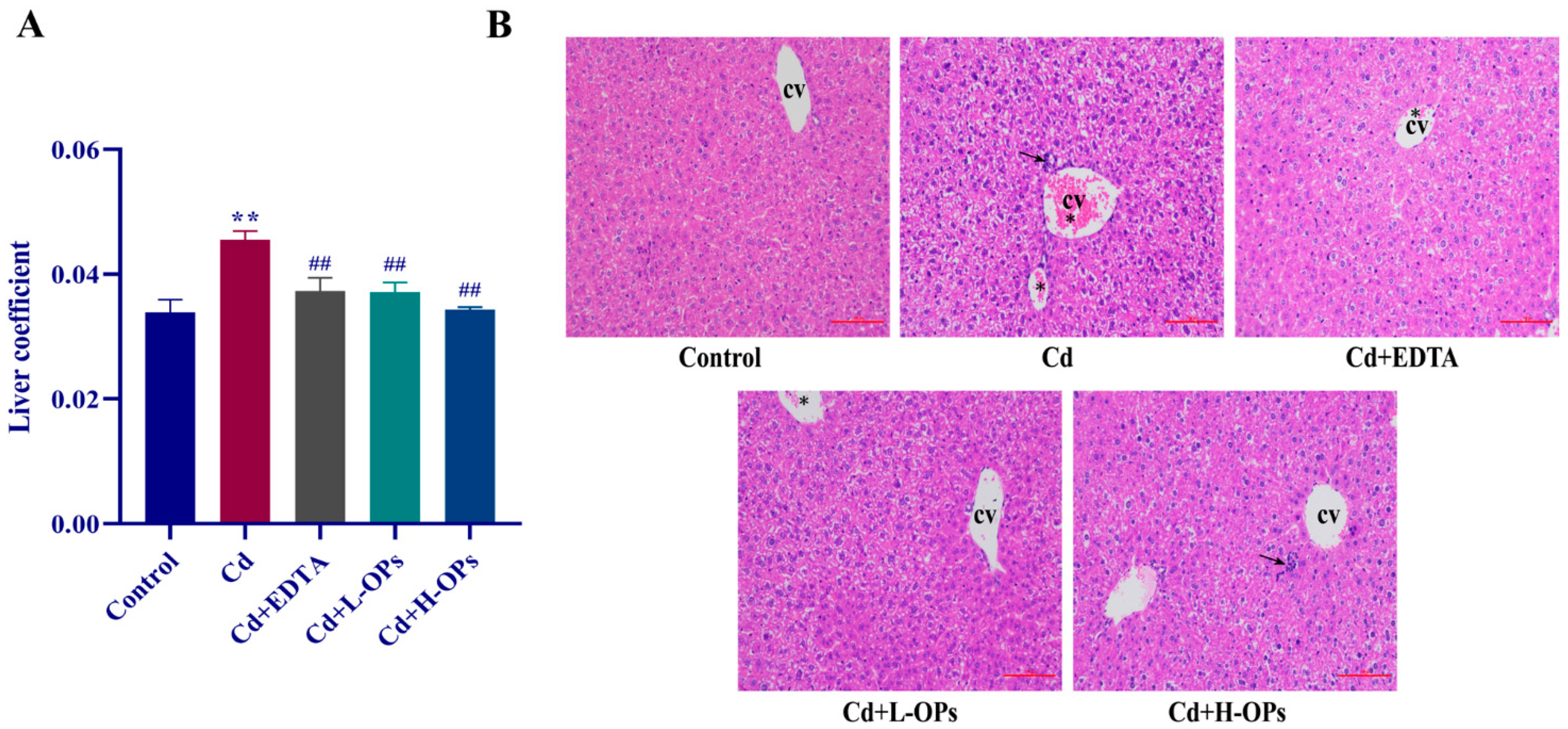
2.5. Effect of OPs on Hepatic Injury in Cd-Exposed Mice
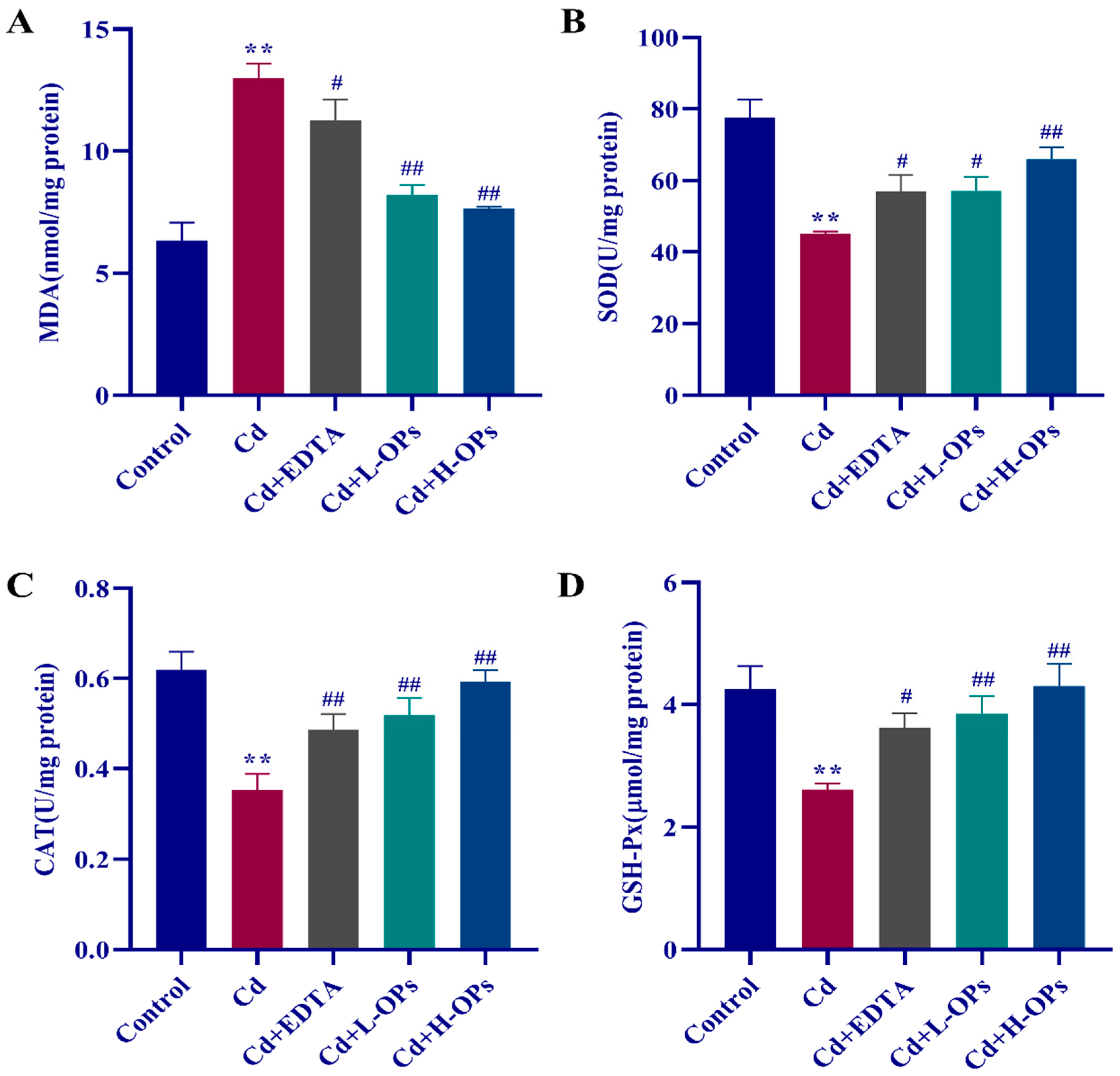
2.6. Effect of OPs on Hepatic Oxidative Indexes in Cd-Exposed Mice
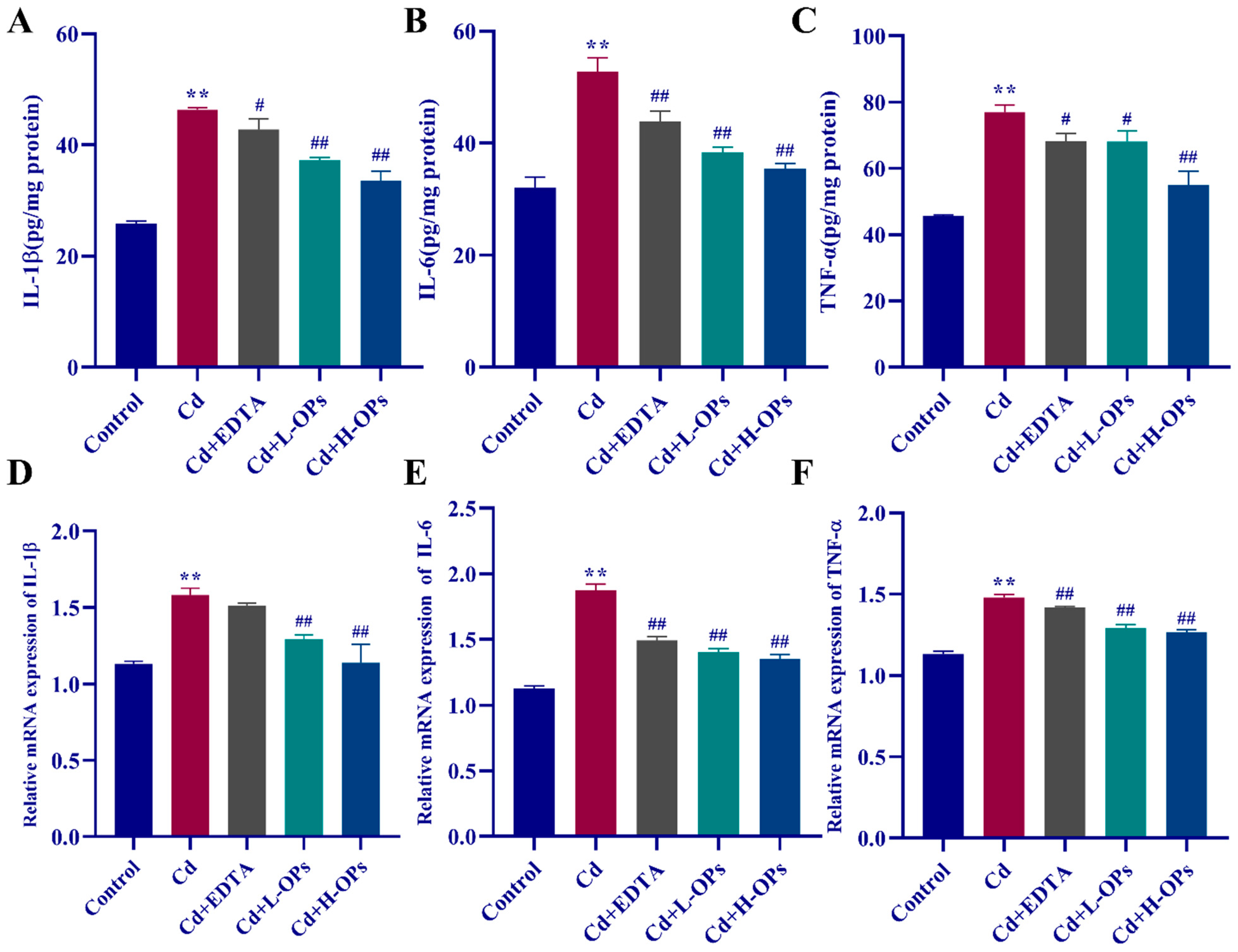
2.7. Effect of OPs on The Hepatic Inflammatory Response (IL-1β, IL-6, TNF-α) in Cd-Exposed Mice
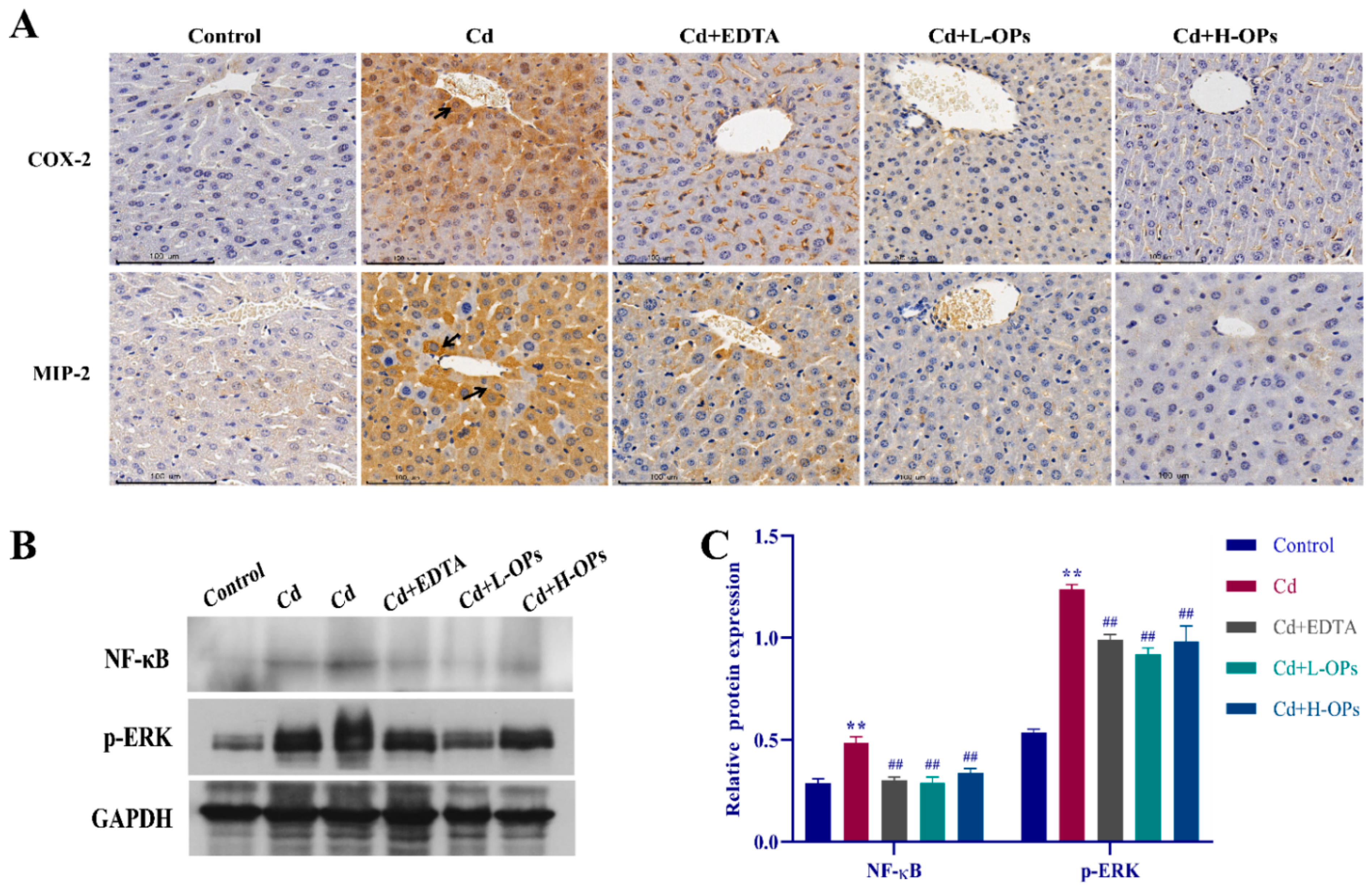
2.8. Effect of OPs on The Expression of Hepatic COX-2, MIP-2, NF-κB, and p-ERK in Cd-Exposed Mice
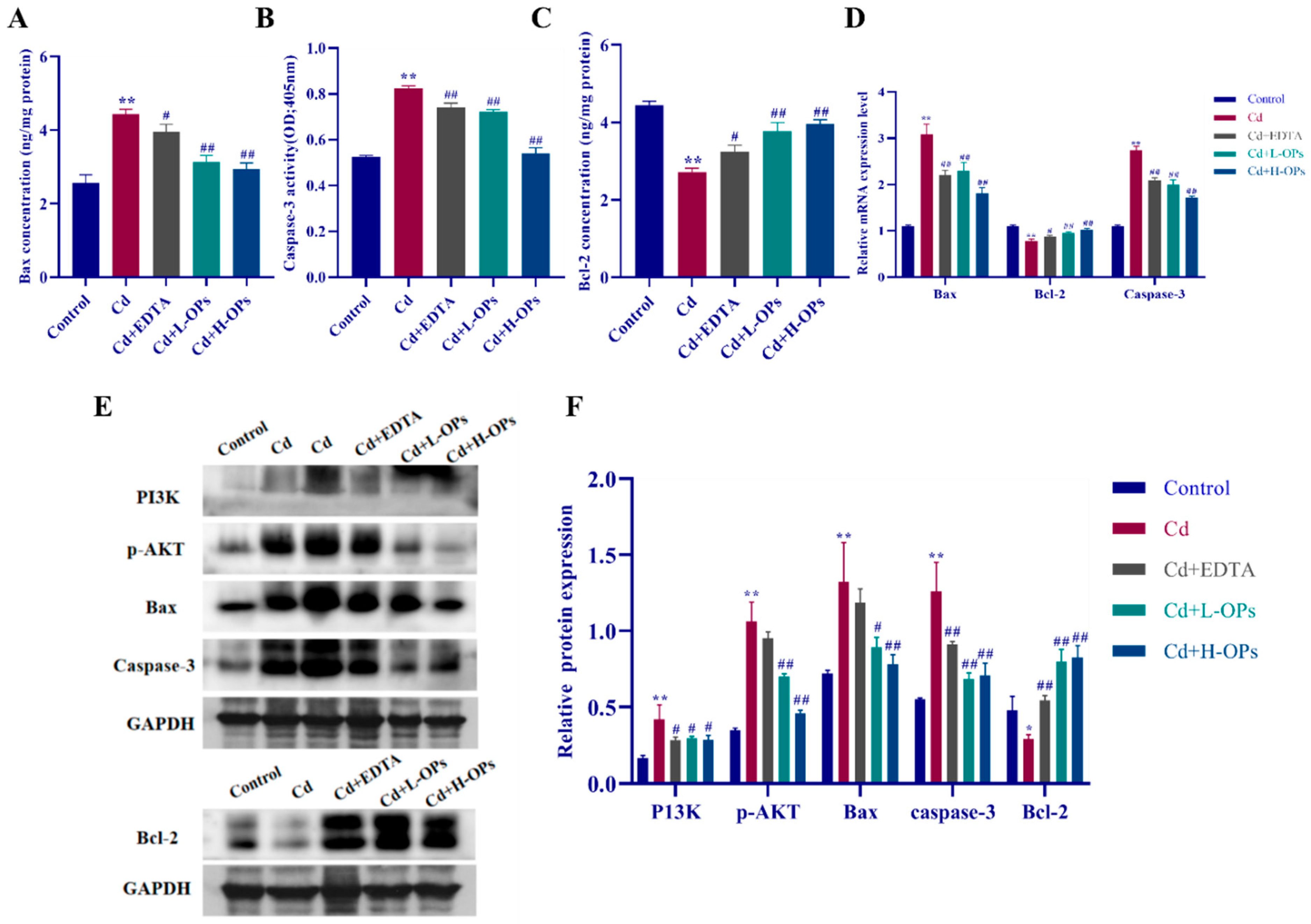
2.9. Effect of OPs on Hepatic Apoptosis in Cd-Exposed Mice
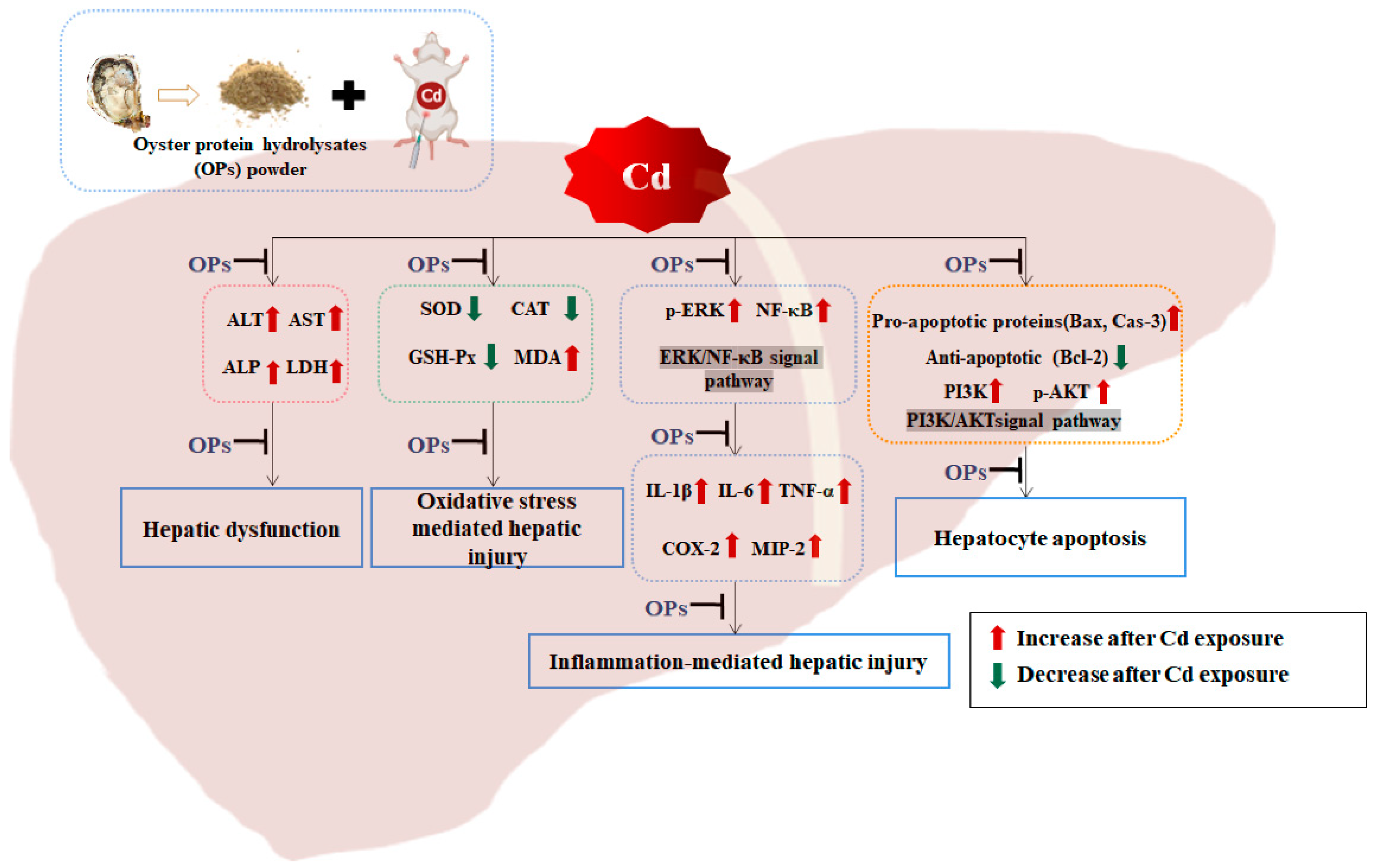
3. Discussion
4. Materials and Methods
4.1. Chemical and Materials
4.2. Animals and Experimental Design
4.3. Preparation of OPs
4.4. Peptide Sequence Analysis Based on LC-MS/MS
4.5. LC-MS/MS Analysis of Free Amino Acids
4.6. Analysis of Amino Acid Composition
4.7. Histopathology Examination
4.8. Analysis of Liver Function
4.9. Measurement of MDA, SOD, CAT and GSH-Px Activities
4.10. Measurement of IL-1β, IL-6 and TNF-α in Hepatic Tissue by ELISA Kits
4.11. Immunohistochemistry Analysis
4.12. Measurement of Bax, Caspase-3 and Bcl-2 in Hepatic Tissue
4.13. Quantitative Reverse-Transcription PCR (qRT-PCR) Analyses
4.14. Western Blotting Analyses
4.15. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, A.; Abou-Zaid, O.A.; Abdel-Maksoud, H.A.; Aboubakr, M.; Abdelkader, A.; Abdelnaby, A.; Abo-Ahmed, A.I.; El-Mleeh, A.; Mostafa, O.; Abdel-Daim, M.; et al. Cadmium overload modulates piroxicam-regulated oxidative damage and apoptotic pathways. Env. Sci. Pollut. Res. Int. 2019, 26, 25167–25177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Fu, L.; Wang, B.; Ji, Y.L.; Wang, H.; Xu, D.X. Chronic cadmium exposure induced hepatic cellular stress and inflammation in aged female mice. J. Appl. Toxicol. 2019, 39, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, T.S.; Staessen, J.A.; Roels, H.A.; Munters, E.; Cuypers, A.; Richart, T.; Ruttens, A.; Smeets, K.; Clijsters, H.; Vangronsveld, J. Cadmium exposure in the population: From health risks to strategies of prevention. BioMetals 2010, 23, 769–782. [Google Scholar] [CrossRef]
- Arroyo, V.; Flores, K.; Ortiz, L.; Gómez-Quiroz, L.; Gutiérrez-Ruiz, M. Liver and Cadmium Toxicity. J. Drug Metab. Toxicol. 2012, 5, 1–7. [Google Scholar]
- Andjelkovic, M.; Buha Djordjevic, A.; Antonijevic, E.; Antonijevic, B.; Stanic, M.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; Jovanovic, M.; Boricic, N.; Wallace, D.; et al. Toxic Effect of Acute Cadmium and Lead Exposure in Rat Blood, Liver, and Kidney. Int. J. Env. Res. Public Health 2019, 16, 274. [Google Scholar] [CrossRef]
- Almeer, R.S.; Alarifi, S.; Alkahtani, S.; Ibrahim, S.R.; Ali, D.; Moneim, A. The potential hepatoprotective effect of royal jelly against cadmium chloride-induced hepatotoxicity in mice is mediated by suppression of oxidative stress and upregulation of Nrf2 expression. Biomed. Pharm. 2018, 106, 1490–1498. [Google Scholar] [CrossRef]
- Miao, J.; Liao, W.; Kang, M.; Jia, Y.; Wang, Q.; Duan, S.; Xiao, S.; Cao, Y.; Ji, H. Anti-fatigue and anti-oxidant activities of oyster (Ostrea rivularis) hydrolysate prepared by compound protease. Food Funct. 2018, 9, 6577–6585. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef]
- Fuda, H.; Watanabe, M.; Hui, S.P.; Joko, S.; Okabe, H.; Jin, S.; Takeda, S.; Miki, E.; Watanabe, T.; Chiba, H. Anti-apoptotic effects of novel phenolic antioxidant isolated from the Pacific oyster (Crassostrea gigas) on cultured human hepatocytes under oxidative stress. Food Chem. 2015, 176, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Umayaparvathi, S.; Arumugam, M.; Meenakshi, S.; Dräger, G.; Kirschning, A.; Balasubramanian, T. Purification and Characterization of Antioxidant Peptides from Oyster (Saccostrea cucullata) Hydrolysate and the Anticancer Activity of Hydrolysate on Human Colon Cancer Cell Lines. Int. J. Pept. Res. Ther. 2013, 20, 231–243. [Google Scholar] [CrossRef]
- Siregar, A.S.; Nyiramana, M.M.; Kim, E.J.; Cho, S.B.; Woo, M.S.; Lee, D.K.; Hong, S.G.; Han, J.; Kang, S.S.; Kim, D.R.; et al. Oyster-Derived Tyr-Ala (YA) Peptide Prevents Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Suppressing Inflammatory, Apoptotic, Ferroptotic, and Pyroptotic Signals. Mar. Drugs 2021, 19, 614. [Google Scholar] [CrossRef] [PubMed]
- Umayaparvathi, S.; Meenakshi, S.; Vimalraj, V.; Arumugam, M.; Sivagami, G.; Balasubramanian, T. Antioxidant activity and anticancer effect of bioactive peptide from enzymatic hydrolysate of oyster (Saccostrea cucullata). Biomed. Prev. Nutr. 2014, 4, 343–353. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Jing, X.; Zhang, B.; Zhang, Q.; Niu, Q.; Wang, J.; Tian, Z. Protective effects of oyster extract against hepatic tissue injury in alcoholic liver diseases. J. Ocean Univ. China 2014, 13, 262–270. [Google Scholar] [CrossRef]
- Li, H.; Xia, X.; Zang, J.; Tan, X.; Wang, Z.; Xu, X.; Du, M. Oyster (Crassostrea gigas) ferritin can efficiently reduce the damage of Pb2+ in vivo by electrostatic attraction. Int. J. Biol. Macromol. 2022, 210, 365–376. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive peptides from marine processing waste and shellfish: A review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Rakesh, K.P.; Suhas, R.; Gowda, D.C. Anti-inflammatory and Antioxidant Peptide-Conjugates: Modulation of Activity by Charged and Hydrophobic Residues. In. J. Pept. Res. Ther. 2019, 25, 227–234. [Google Scholar] [CrossRef]
- Mahmood, S.; Wahid, A.; Azeem, M.; Zafar, S.; Bashir, R.; Sharif, O.; Ali, S. Tyrosine or lysine priming modulated phenolic metabolism and improved cadmium stress tolerance in mung bean (Vigna radiata L.). S. Afr. J. Bot. 2022, 149, 397–406. [Google Scholar] [CrossRef]
- Zemanová, V.; Pavlík, M.; Pavlíková, D.; Tlustoš, P. The significance of methionine, histidine and tryptophan in plant responses and adaptation to cadmium stress. Plant Soil Environ. 2014, 60, 426–432. [Google Scholar] [CrossRef]
- Domínguez-Solís, J.R.; López-Martín, M.C.; Ager, F.J.; Ynsa, M.D.; Romero, L.C.; Gotor, C. Increased cysteine availability is essential for cadmium tolerance and accumulation in Arabidopsis thaliana. Plant Biotechnol. J. 2004, 2, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Hoque, M.A.; Okuma, E.; Banu, M.N.A.; Shimoishi, Y.; Nakamura, Y.; Murata, Y. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Born, T.; Kontoghiorghe, C.N.; Spyrou, A.; Kolnagou, A.; Kontoghiorghes, G.J. EDTA chelation reappraisal following new clinical trials and regular use in millions of patients: Review of preliminary findings and risk/benefit assessment. Toxicol. Mech. Methods 2013, 23, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Michael, B.; Yano, B.; Sellers, R.S.; Perry, R.; Morton, D.; Roome, N.; Johnson, J.K.; Schafer, K. Evaluation of Organ Weights for Rodent and Non-Rodent Toxicity Studies: A Review of Regulatory Guidelines and a Survey of Current Practices. Toxicol. Pathol. 2007, 35, 742–750. [Google Scholar] [CrossRef]
- Arranz, J.; Soriano, A.; Garcia, I.; García, I.; Concepción, M.T.; Navarro, J.; Arteaga, A.; Filella, X.; Bravo, P.; Barrera, M.; et al. Effect of proinflammatory cytokines (IL-6, TNF-α, IL-1β) on hemodynamic performance during orthotopic liver transplantation. Transpl. P. 2003, 35, 1884–1887. [Google Scholar] [CrossRef]
- Arras, D.S.; John, S.R. IL-6 pathway in the liver: From physiopathology to therapy. J. Hepatol. 2016, 64, 1403–1415. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Brenner, D.A. Mechanisms of Liver Injury. I. TNF-α-induced liver injury: Role of IKK, JNK, and ROS pathways. Am. J. Physiol. Gastr. L. 2006, 290, G583–G589. [Google Scholar] [CrossRef]
- Yu, J.; Ip, E.; dela Peña, A.; Hou, J.Y.; Sesha, J.; Pera, N.; Hall, P.; Kirsch, R.; Leclercq, I.; Farrell, G.C. COX-2 induction in mice with experimental nutritional steatohepatitis: Role as pro-inflammatory mediator. Hepatology 2006, 43, 826–836. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, S.; Liu, J.; Wu, X.; Zhou, S.; Dai, K.; Kou, Y. Mitophagy Contributes to the Pathogenesis of Inflammatory Diseases. Inflammation 2018, 41, 1590–1600. [Google Scholar] [CrossRef]
- Huang, Y.-Y.; Xia, M.-Z.; Wang, H.; Liu, X.-J.; Hu, Y.-F.; Chen, Y.-H.; Zhang, C.; Xu, D.-X. Cadmium Selectively Induces MIP-2 and COX-2 Through PTEN-Mediated Akt Activation in RAW264.7 Cells. Toxicol. Sci. 2014, 138, 310–321. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Zhang, J.; Qi, X.; Cui, Y.; Yin, K.; Lin, H. Cadmium induced inflammation and apoptosis of porcine epididymis via activating RAF1/MEK/ERK and NF-κB pathways. Toxicol. Appl. Pharmacol. 2021, 415, 115449. [Google Scholar] [CrossRef] [PubMed]
- Habeebu, S.S.M.; Liu, J.; Klaassen, C.D. Cadmium-Induced Apoptosis in Mouse Liver. Toxicol. Appl. Pharm. 1998, 149, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Thomadaki, H.; Scorilas, A. BCL2 Family of Apoptosis-Related Genes: Functions and Clinical Implications in Cancer. Crit. Rev. Clin. Lab. Sci. 2006, 43, 1–67. [Google Scholar] [CrossRef] [PubMed]
- Yiming, L.; Yanfei, H.; Hang, Y.; Yimei, C.; Guangliang, S.; Shu, L. Cadmium induces apoptosis of pig lymph nodes by regulating the PI3K/AKT/HIF-1α pathway. Toxicology 2021, 451, 152694. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Zhou, S.; Wang, Y.; Shi, X.; Liu, H. Cadmium exposure activates the PI3K/AKT signaling pathway through miRNA-21, induces an increase in M1 polarization of macrophages, and leads to fibrosis of pig liver tissue. Ecotoxicol. Environ. Saf. 2021, 228, 113015. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Barbero, A.; Dorado, F.; Velasco, S.; Pandiella, A.; Banas, B.; López Novoa, J.M. TGF-β1 induces COX-2 expression and PGE2 synthesis through MAPK and PI3K pathways in human mesangial cells. Kidney Int. 2006, 70, 901–909. [Google Scholar] [CrossRef]
- Matović, V.; Buha, A.; Ðukić-Ćosić, D.; Bulat, Z. Insight into the oxidative stress induced by lead and/or cadmium in blood, liver and kidneys. Food Chem. Toxicol. 2015, 78, 130–140. [Google Scholar] [CrossRef]
- Wu, H.-C.; Chen, H.-M.; Shiau, C.-Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Nasri, M. Chapter Four—Protein Hydrolysates and Biopeptides: Production, Biological Activities, and Applications in Foods and Health Benefits. A Review. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 109–159. [Google Scholar]
- Ghaly, A.E.; Vv, R.; Brooks, M.S.-L.; Budge, S.M.; Dave, D. Fish Processing Wastes as a Potential Source of Proteins, Amino Acidsand Oils: A Critical Review. J. Microb. Biochem. Technol. 2013, 2, 107–129. [Google Scholar]
- Yang, B.; Yang, H.; Li, J.; Li, Z.; Jiang, Y. Amino acid composition, molecular weight distribution and antioxidant activity of protein hydrolysates of soy sauce lees. Food Chem. 2011, 124, 551–555. [Google Scholar] [CrossRef]
- Rania, J.H.; Hassan, H.M.M.; Afify, A.S. Evaluation of Antioxidant and Metal Chelating Activities of Protein Hydrolysates Produced from Leather Waste by Alkaline and Enzymatic Hydrolysis. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 910–919. [Google Scholar]
- Gu, H.; Song, I.-B.; Han, H.-J.; Lee, N.-Y.; Cha, J.-Y.; Son, Y.-K.; Kwon, J. Antioxidant Activity of Royal Jelly Hydrolysates Obtained by Enzymatic Treatment. Korean J. Food Sci. Anim. Resour. 2018, 38, 135–142. [Google Scholar] [PubMed]
- Ovissipour, M.; Rasco, B.; Shiroodi, S.G.; Modanlow, M.; Gholami, S.; Nemati, M. Antioxidant activity of protein hydrolysates from whole anchovy sprat (Clupeonella engrauliformis) prepared using endogenous enzymes and commercial proteases. J. Sci. Food Agric. 2013, 93, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- DeRosa, G.; Swick, R.W. Metabolic implications of the distribution of the alanine aminotransferase isoenzymes. J. Biol. Chem. 1975, 250, 7961–7967. [Google Scholar] [CrossRef]
- Fernando, S.; Wijewickrama, A.; Gomes, L.; Punchihewa, C.T.; Madusanka, S.D.; Dissanayake, H.; Jeewandara, C.; Peiris, H.; Ogg, G.S.; Malavige, G.N. Patterns and causes of liver involvement in acute dengue infection. BMC Infect. Dis. 2016, 16, 319. [Google Scholar] [CrossRef]
- Noor, K.K.; Ijaz, M.U.; Ehsan, N.; Tahir, A.; Yeni, D.K.; Neamul Kabir Zihad, S.M.; Uddin, S.J.; Ashraf, A.; Simal-Gandara, J. Hepatoprotective role of vitexin against cadmium-induced liver damage in male rats: A biochemical, inflammatory, apoptotic and histopathological investigation. Biomed. Pharmacother. 2022, 150, 112934. [Google Scholar] [CrossRef]
- Wang, K.; Shi, J.; Gao, S.; Hong, H.; Tan, Y.; Luo, Y. Oyster protein hydrolysates alleviated chronic alcohol-induced liver injury in mice by regulating hepatic lipid metabolism and inflammation response. Food Res. Int. 2022, 160, 111647. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, J.; He, H.; Guo, H.; Zhang, S. Hepatoprotective effects of Ganoderma lucidum peptides against d-galactosamine-induced liver injury in mice. J. Ethnopharmacol. 2008, 117, 415–419. [Google Scholar] [CrossRef]
- Mumtaz, S.; Ali, S.; Khan, R.; Andleeb, S.; Ulhaq, M.; Khan, M.A.; Shakir, H.A. The protective role of ascorbic acid in the hepatotoxicity of cadmium and mercury in rabbits. Environ. Sci. Pollut. R. 2019, 26, 14087–14096. [Google Scholar] [CrossRef]
- Holeček, M.; Mráz, J.; Tilšer, I. Plasma amino acids in four models of experimental liver injury in rats. Amino Acids 1996, 10, 229–241. [Google Scholar] [CrossRef]
- Ryu, B.; Shin, K.-H.; Kim, S.-K. Muscle Protein Hydrolysates and Amino Acid Composition in Fish. Mar. Drugs 2021, 19, 377. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, Y.; Arisaka, H.; Yoshida, S.; Mori, M.; Takahashi, M. Proline protects liver from d-galactosamine hepatitis by activating the IL-6/STAT3 survival signaling pathway. Amino Acids 2012, 43, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.-W.; Tian, L.-X.; Li, Y.-M.; Zhou, W.; Zeng, S.-L.; Yang, H.-J.; Liu, Y.-J. Effect of proline supplementation on anti-oxidative capacity, immune response and stress tolerance of juvenile Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2015, 448, 105–111. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, L.; Chen, Y.; Ni, Q.I.; Zhou, M.; Qu, C.; Zhang, Y. Antifibrotic effect of N-acetyl-seryl-aspartyl-lysyl-proline on bile duct ligation induced liver fibrosis in rats. World J. Gastroenterol. 2012, 18, 5283–5288. [Google Scholar]
- Gujjar, R.; Pathak, A.D.; Karkute, S.; Supaibulwattana, K. Multifunctional proline rich proteins and their role in regulating cellular proline content in plants under stress. Biol. Plant. 2019, 63, 448–454. [Google Scholar] [CrossRef]
- Yao Chai, T.; Didierjean, L.; Burkard, G.; Genot, G. Expression of a green tissue-specific 11 kDa proline-rich protein gene in bean in response to heavy metals. Plant Sci. 1998, 133, 47–56. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef]
- Komatsu, T.; Kobayashi, K.; Morimoto, Y.; Helmerhorst, E.; Oppenheim, F.; Chang-Il Lee, M. Direct evaluation of the antioxidant properties of salivary proline-rich proteins. J. Clin. Biochem. Nutr. 2020, 67, 131–136. [Google Scholar] [CrossRef]
- Galoyan, A.A.; Shakhlamov, V.A.; Aghajanov, M.I.; Vahradyan, H.G. Hypothalamic Proline-Rich Polypeptide Protects Brain Neurons in Aluminum Neurotoxicosis. Neurochem. Res. 2004, 29, 1349–1357. [Google Scholar] [CrossRef]
- Das, S.C.; Al-Naemi, H.A. Cadmium Toxicity: Oxidative Stress, Inflammation and Tissue Injury. Occup. Dis. Environ. Med. 2019, 7, 144–163. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Albano, E.; Dianzani, M.U. The role of lipid peroxidation in liver damage. Chem. Phys. Lipids 1987, 45, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Fang, Z.; Sun, L.; Wang, Y.; Liu, Y.; Xu, D.; Nie, F.; Gooneratne, R. Oleic Acid Alleviates Cadmium-Induced Oxidative Damage in Rat by Its Radicals Scavenging Activity. Biol. Trace Elem. Res. 2019, 190, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Elkomy, A.; Elkammar, R.; Youssef, G.; Abdelhiee, E.Y.; Abdo, W.; Fadl, S.E.; Soliman, A.; Aboubakr, M. Synergistic protective effects of lycopene and N-acetylcysteine against cisplatin-induced hepatorenal toxicity in rats. Sci. Rep. 2021, 11, 13979. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Elkomy, A.; Alkafafy, M.; Elkammar, R.; Fadl, S.E.; Abdelhiee, E.Y.; Abdeen, A.; Youssef, G.; Shaheen, H.; Soliman, A.; et al. Ameliorating Effect of Lycopene and N-Acetylcysteine against Cisplatin-Induced Cardiac Injury in Rats. Pak. Vet. J. 2022, 42, 107–111. [Google Scholar]
- Elsayed, A.; Elkomy, A.; Alkafafy, M.; Elkammar, R.; El-Shafey, A.; Soliman, A.; Aboubakr, M. Testicular toxicity of cisplatin in rats: Ameliorative effect of lycopene and N-acetylcysteine. Environ. Sci. Pollut. R. 2022, 29, 24077–24084. [Google Scholar] [CrossRef]
- Fan, R.; Hu, P.-C.; Wang, Y.; Lin, H.-Y.; Su, K.; Feng, X.-S.; Wei, L.; Yang, F. Betulinic acid protects mice from cadmium chloride-induced toxicity by inhibiting cadmium-induced apoptosis in kidney and liver. Toxicol. Lett. 2018, 299, 56–66. [Google Scholar] [CrossRef]
- Shaikh, Z.A.; Vu, T.T.; Zaman, K. Oxidative Stress as a Mechanism of Chronic Cadmium-Induced Hepatotoxicity and Renal Toxicity and Protection by Antioxidants. Toxicol. Appl. Pharmacol. 1999, 154, 256–263. [Google Scholar] [CrossRef]
- Jemai, H.; Messaoudi, I.; Chaouch, A.; Kerkeni, A. Protective effect of zinc supplementation on blood antioxidant defense system in rats exposed to cadmium. J. Trace Elem. Med. Biol. 2007, 21, 269–273. [Google Scholar] [CrossRef]
- Jemai, H.; Mahmoudi, A.; Feryeni, A.; Fki, I.; Bouallagui, Z.; Choura, S.; Chamkha, M.; Sayadi, S. Hepatoprotective Effect of Oleuropein-Rich Extract from Olive Leaves against Cadmium-Induced Toxicity in Mice. Biomed. Res. Int. 2020, 2020, 4398924. [Google Scholar] [CrossRef]
- Li, Y.; Yu, J. Research Progress in Structure-Activity Relationship of Bioactive Peptides. J. Med. Food 2014, 18, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, J.; Tang, X.; Xiong, Y.L. Reducing, Radical Scavenging, and Chelation Properties of in Vitro Digests of Alcalase-Treated Zein Hydrolysate. J. Agric. Food Chem. 2008, 56, 2714–2721. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Wree, A.; Holtmann, T.M.; Inzaugarat, M.E.; Feldstein, A.E. Novel Drivers of the Inflammatory Response in Liver Injury and Fibrosis. Semin. Liver Dis. 2019, 39, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.; Li, N.; Brayman, T.G.; Hotz, K.J.; Lai, Y. Regulation of MRP2/ABCC2 and BSEP/ABCB11 Expression in Sandwich Cultured Human and Rat Hepatocytes Exposed to Inflammatory Cytokines TNF-α, IL-6, and IL-1β. J. Biol. Chem. 2010, 285, 31185–31192. [Google Scholar] [CrossRef]
- Zhao, Z.; Hyun, J.S.; Satsu, H.; Kakuta, S.; Shimizu, M. Oral exposure to cadmium chloride triggers an acute inflammatory response in the intestines of mice, initiated by the over-expression of tissue macrophage inflammatory protein-2 mRNA. Toxicol. Lett. 2006, 164, 144–154. [Google Scholar] [CrossRef]
- Qian, B.; Zhao, X.; Yang, Y.; Tian, C. Antioxidant and anti-inflammatory peptide fraction from oyster soft tissue by enzymatic hydrolysis. Food Sci. Nutr. 2020, 8, 3947–3956. [Google Scholar] [CrossRef]
- Li, S.; Sun, W.; Zhang, K.; Zhu, J.; Jia, X.; Guo, X.; Zhao, Q.; Tang, C.; Yin, J.; Zhang, J. Selenium deficiency induces spleen pathological changes in pigs by decreasing selenoprotein expression, evoking oxidative stress, and activating inflammation and apoptosis. J. Anim. Sci. Biotechno. 2021, 12, 65. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, B.; Zheng, Q.; Zhu, G.; Cao, W.; Qin, X.; Zhang, C. Ameliorative Effects of Peptides from the Oyster (Crassostrea hongkongensis) Protein Hydrolysates against UVB-Induced Skin Photodamage in Mice. Mar. Drugs 2020, 18, 288. [Google Scholar] [CrossRef]
- Qian, Z.-J.; Chen, M.-F.; Chen, J.; Zhang, Y.; Zhou, C.; Hong, P.; Yang, P. Intracellular ethanol-mediated oxidation and apoptosis in HepG2/CYP2E1 cells impaired by two active peptides from seahorse (Hippocampus kuda bleeler) protein hydrolysates via the Nrf2/HO-1 and akt pathways. Food Sci. Nutr. 2021, 9, 1584–1602. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, L.; Luo, N.; Wang, Y.Q.; Gao, H.M. Inhibition of PI3K/AKt/mTOR signaling pathway protects against d-galactosamine/lipopolysaccharide-induced acute liver failure by chaperone-mediated autophagy in rats. Biomed Pharm. 2017, 92, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Qian, K.; Xiong, J.; Ma, K.; Wang, A.; Zou, Y. Curcumin alleviates lipopolysaccharide induced sepsis and liver failure by suppression of oxidative stress-related inflammation via PI3K/AKT and NF-kappaB related signaling. Biomed. Pharm. 2016, 83, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gu, J.; Hu, L.; Kong, L.; Wang, T.; Di, M.; Li, C.; Gui, S. miR-130a alleviates neuronal apoptosis and changes in expression of Bcl-2/Bax and caspase-3 in cerebral infarction rats through PTEN/PI3K/Akt signaling pathway. Exp. Med. 2020, 19, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Zhang, Y.; Xing, H.; Xu, S. Ameliorative Effect of Selenomethionine on Cadmium-Induced Hepatocyte Apoptosis via Regulating PI3K/AKT Pathway in Chickens. Biol. Trace Elem. Res. 2020, 195, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Augustine, N.; Ani, C.; Eze, W.; Ugwudike, P.; Anyaeji, P.; Ude, V.C.; Agu, F.U.; Nworgu, C.; Ikwuka, D.; Ugwuishi, E.; et al. The effect of aqueous extract of zest of citrus sinensis (AEZCs) on cadmium chloride induced liver toxicity in wistar rats. Afr. J. Biochem. R. 2020, 14, 5–17. [Google Scholar] [CrossRef][Green Version]
- Najeeb, U.; Jilani, G.; Ali, S.; Sarwar, M.; Xu, L.; Zhou, W. Insights into cadmium induced physiological and ultra-structural disorders in Juncus effusus L. and its remediation through exogenous citric acid. J. Hazard. Mater. 2011, 186, 565–574. [Google Scholar] [CrossRef]
- El-Naggar, S.A.; El-Said, K.S.; Elwan, M.; Mobasher, M.; Mansour, F.; Elbakry, M.; Kabil, D.I. Toxicity of bean cooking media containing EDTA in mice. Toxicol. Ind. Health 2020, 36, 436–445. [Google Scholar] [CrossRef]
- Zhang, X.; Peng, Z.; Zheng, H.; Zhang, C.; Lin, H.; Qin, X. The Potential Protective Effect and Possible Mechanism of Peptides from Oyster (Crassostrea hongkongensis) Hydrolysate on Triptolide-Induced Testis Injury in Male Mice. Mar. Drugs 2021, 19, 566. [Google Scholar] [CrossRef]
- Jansen, R.; Lachatre, G.; Marquet, P. LC-MS/MS systematic toxicological analysis: Comparison of MS/MS spectra obtained with different instruments and settings. Clin. Biochem. 2005, 38, 362–372. [Google Scholar] [CrossRef]
- Łozowicka, B.; Kaczyński, P.; Iwaniuk, P. Analysis of 22 free amino acids in honey from Eastern Europe and Central Asia using LC-MS/MS technique without derivatization step. J. Food Compos. Anal. 2021, 98, 103837. [Google Scholar] [CrossRef]
- He, S.; Zhuo, L.; Cao, Y.; Liu, G.; Zhao, H.; Song, R.; Liu, Z. Effect of cadmium on osteoclast differentiation during bone injury in female mice. Environ. Toxicol. 2020, 35, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, X.; Rao, X.; Zheng, C.; Peng, X. Saikosaponin a ameliorates lipopolysaccharide and d-galactosamine-induced liver injury via activating LXRα. Int. Immunopharmacol. 2019, 72, 131–137. [Google Scholar] [CrossRef]
- Yang, P.; Xu, F.; Li, H.-F.; Wang, Y.; Li, F.-C.; Shang, M.-Y.; Liu, G.-X.; Wang, X.; Cai, S.-Q. Detection of 191 Taxifolin Metabolites and Their Distribution in Rats Using HPLC-ESI-IT-TOF-MSn. Molecules 2016, 21, 1209. [Google Scholar] [CrossRef] [PubMed]
- Fornetti, J.; Jindal, S.; Middleton, K.; Borges, V.; Schedin, P. Physiological COX-2 Expression in Breast Epithelium Associates with COX-2 Levels in Ductal Carcinoma in Situ and Invasive Breast Cancer in Young Women. Am. J. Pathol. 2014, 184, 1220–1229. [Google Scholar] [CrossRef][Green Version]
- He, Q.; Luo, Y.; Xie, Z. Sulforaphane ameliorates cadmium induced hepatotoxicity through the up-regulation of /Nrf2/ARE pathway and the inactivation of NF-κB. J. Funct. Foods 2021, 77, 104297. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Peptide Sequence | Length | Molecular Mass (Da) | Observed Mass (m/z) | Peak Area | Relative Peak Area (%) | Scores |
|---|---|---|---|---|---|---|---|
| 1 | GEPGPEGPAGPIGPR | 15 | 1387.697 | 694.350 | 18,567,000 | 0.15 | 614.6 |
| 2 | YEETRGVLK | 9 | 1094.584 | 547.795 | 9,895,900 | 0.08 | 494.2 |
| 3 | GPTGPVGPL | 9 | 794.441 | 794.437 | 3,857,400 | 0.03 | 477.2 |
| 4 | GPSGEPGPE | 9 | 826.358 | 826.354 | 12,054,000 | 0.10 | 468.7 |
| 5 | DIERPTYT | 8 | 994.484 | 497.745 | 18,359,000 | 0.15 | 461.4 |
| 6 | ENPVPVPS | 8 | 838.431 | 838.427 | 6,999,200 | 0.06 | 453.3 |
| 7 | TEAPLNPK | 8 | 869.473 | 869.471 | 9,380,400 | 0.08 | 451.1 |
| 8 | TPEEFIPR | 8 | 988.510 | 494.758 | 12,323,000 | 0.10 | 437.8 |
| 9 | AGFAGDDAPR | 10 | 976.448 | 488.727 | 171,200,000 | 1.39 | 430.9 |
| 10 | TPTYGDL | 7 | 766.362 | 766.359 | 46,716,000 | 0.38 | 428.1 |
| 11 | PDVPAGDVDKGK | 12 | 1197.611 | 399.874 | 14,901,000 | 0.12 | 425.1 |
| 12 | GPIGGPL | 7 | 610.356 | 610.354 | 5,358,100 | 0.04 | 423.7 |
| 13 | SPVGVGA | 7 | 586.320 | 586.318 | 13,332,000 | 0.11 | 411.4 |
| 14 | YTPVAYPV | 8 | 586.320 | 586.318 | 4,007,500 | 0.03 | 388.5 |
| 15 | LTPSGLPY | 8 | 647.342 | 647.342 | 14,591,000 | 0.12 | 386.7 |
| 16 | STPFEGF | 7 | 571.308 | 571.308 | 13,332,000 | 0.11 | 384.4 |
| 17 | VSDTVVEPYN | 10 | 550.250 | 550.250 | 27,505,000 | 0.22 | 383.3 |
| 18 | DIERPTYTN | 9 | 909.420 | 909.420 | 8,461,100 | 0.07 | 383.2 |
| 19 | QGETGDRGPFG | 11 | 879.456 | 440.231 | 34,716,000 | 0.28 | 383.0 |
| 20 | PRPPTQVGGS | 10 | 995.525 | 498.266 | 38,429,000 | 0.31 | 382.8 |
| Amino Acids | Contents (g/100 g) | Amino Acids | Contents (g/100 g) |
|---|---|---|---|
| Alanine (Ala) # | 2.63 | Leucine (Leu) *# | 2.02 |
| Cystine (Cys) | 0.40 | Methionine (Met) *# | 0.65 |
| Aspartic acid (Asp) | 3.36 | Proline (Pro) # | 2.61 |
| Glutamic acid (Glu) | 4.33 | Arginine (Arg) | 1.71 |
| Phenylalanine (Phe) *# | 0.72 | Serine (Ser) | 1.83 |
| Glycine (Gly) | 2.92 | Threonine (Thr) * | 1.69 |
| Histidine (His) | 0.50 | Valine (Val) *# | 1.81 |
| Isoleucine (Ile) *# | 1.40 | Tyrosine (Tyr) | 1.62 |
| Lysine (Lys) * | 2.61 | Tryptophan (Trp) *# | 0.92 |
| Total amino acids | 33.73 | ||
| Essential amino acid | 11.82 | ||
| Hydrophobic amino acids | 12.76 |
| Free Amino Acids | Contents (g/100 g) | Free Amino Acids | Contents (g/100 g) |
|---|---|---|---|
| Ala # | 0.59 | Met *# | 0.36 |
| Cys | ND | Asn | 0.26 |
| Asp | 0.37 | Pro # | 2.49 |
| Glu | 2.12 | Gln | 0.85 |
| Phe *# | 0.18 | Arg | 0.53 |
| Gly | 1.38 | Ser | 0.29 |
| His | 0.08 | Thr * | 0.84 |
| Ile *# | 0.59 | Val *# | 0.91 |
| Lys * | 1.43 | Trp *# | 0.52 |
| Leu *# | 0.50 | Tyr | 1.51 |
| Total free amino acids | 15.80 | ||
| Essential free amino acid | 5.33 | ||
| Hydrophobic free amino acids | 6.14 |
| Gene Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| IL-1β | GACTTCACCATGGAACCCGT | GGAGACTGCCCATTCTCGAC |
| IL-6 | GGCCCTTGCTTTCTCTTCG | ATAATAAAGTTTTGATTATGT |
| TNF-α | AGCCCTGGTATGAGCCCATGTA | CCGGACTCCGTGATGTCTAAGT |
| Bax | CTGAGCTGACCTTGGAGC | GACTCCAGCCACAAGAGATG |
| Caspase-3 | GAGCTTGGAACGGTACGCTA | CCGTACCAGAGCGAGATGAC |
| Bcl-2 | GACAGAAGATCATGCCGTCC | GGTACCAATGGCACTTCAAG |
| GAPDH | TCACCACCATGGAGAAGGC | GCTAAGCAGTTGGTGGTGCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Fang, Z.; Li, Y.; Sun, L.; Liu, Y.; Deng, Q.; Zhong, S. Ameliorative Effects of Oyster Protein Hydrolysates on Cadmium-Induced Hepatic Injury in Mice. Mar. Drugs 2022, 20, 758. https://doi.org/10.3390/md20120758
Wang J, Fang Z, Li Y, Sun L, Liu Y, Deng Q, Zhong S. Ameliorative Effects of Oyster Protein Hydrolysates on Cadmium-Induced Hepatic Injury in Mice. Marine Drugs. 2022; 20(12):758. https://doi.org/10.3390/md20120758
Chicago/Turabian StyleWang, Jingwen, Zhijia Fang, Yongbin Li, Lijun Sun, Ying Liu, Qi Deng, and Saiyi Zhong. 2022. "Ameliorative Effects of Oyster Protein Hydrolysates on Cadmium-Induced Hepatic Injury in Mice" Marine Drugs 20, no. 12: 758. https://doi.org/10.3390/md20120758
APA StyleWang, J., Fang, Z., Li, Y., Sun, L., Liu, Y., Deng, Q., & Zhong, S. (2022). Ameliorative Effects of Oyster Protein Hydrolysates on Cadmium-Induced Hepatic Injury in Mice. Marine Drugs, 20(12), 758. https://doi.org/10.3390/md20120758