
Evaluation of the Maturity and Gene Expression of Sheep Oocytes and Embryos Cultured in Media Supplemented with Marjoram (Origanum vulgare) Extract

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. O. vulgare Plants and Extract Preparation
2.2. Ovary and Oocyte Collection
2.3. Experimental Design
2.4. Evaluating the DNA in the Cultured Oocytes
2.5. Sperm Capacitation Medium Preparation
2.6. In vitro Fertilization of Oocytes: Fertilization Medium Preparation
2.7. Assessment of Gene Expression
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Máximo, D.M.; da Silva, I.G.M.; Mondadori, R.G.; Neves, J.P.; Lucci, C.M. Ultrastructural characteristics of sheep oocytes during in vitro maturation (IVM). Small Rumin. Res. 2012, 105, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y. Melatonin as a free radical scavenger in the ovarian follicle. Endocr. J. 2013, 60, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.; Reddy, I.J.; Gupta, P.S.P.; Mondal, S. l-carnitine mediated reduction in oxidative stress and alteration in transcript level of antioxidant enzymes in sheep embryos produced in vitro. Reprod. Domest. Anim. 2016, 51, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Cánepa, M.J.; Ortega, N.M.; Monteleone, M.C.; Mucci, N.; Kaiser, G.G.; Brocco, M.; Mutto, A. Expression profile of genes as indicators of developmental competence and quality of in vitro fertilization and somatic cell nuclear transfer bovine embryos. PLoS ONE 2014, 9, e108139. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sharma, R. Oxidative stress and its implications in female infertility–a clinician’s perspective. Reprod. Biomed. Online 2005, 11, 641–650. [Google Scholar] [CrossRef]
- Jamnik, P.; Goranovič, D.; Raspor, P. Antioxidative action of royal jelly in the yeast cell. Exp. Gerontol. 2007, 42, 594–600. [Google Scholar] [CrossRef]
- Nagai, T.; Inoue, R. Preparation and the functional properties of water extract and alkaline extract of royal jelly. Food Chem. 2004, 84, 181–186. [Google Scholar] [CrossRef]
- Nagai, T.; Inoue, R.; Suzuki, N.; Nagashima, T. Antioxidant properties of enzymatic hydrolysates from royal jelly. J. Med. Food 2006, 9, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Olson, S.E.; Seidel, G.E., Jr. Culture of in vitro-produced bovine embryos with vitamin E improves development in vitro and after transfer to recipients. Biol. Reprod. 2000, 62, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Urdaneta, A.; Jiménez-Macedo, A.-R.; Izquierdo, D.; Paramio, M.-T. Supplementation with cysteamine during maturation and embryo culture on embryo development of prepubertal goat oocytes selected by the brilliant cresyl blue test. Zygote 2003, 11, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Robker, R.L. The role of L-carnitine during oocyte in vitro maturation: Essential co-factor? Anim. Reprod. 2018, 14, 469–475. [Google Scholar] [CrossRef]
- Do, L.T.K.; Shibata, Y.; Taniguchi, M.; Nii, M.; Nguyen, T.V.; Tanihara, F.; Takagi, M.; Otoi, T. Melatonin supplementation during in vitro maturation and development supports the development of porcine embryos. Reprod. Domest. Anim. 2015, 50, 1054–1058. [Google Scholar] [CrossRef] [PubMed]
- Barakat, I.A.H.; RAl-Himaidi, A. Effects of fenugreek seed extract on in vitro maturation and subsequent development of sheep oocytes. Pak. J. Zool. 2013, 45. [Google Scholar]
- Barakat, I. a H.; Al-himaidi, A.R.; Rady, A.M. Antioxidant Effect of Green Tea Leaves Extract on in vitro Production of Sheep Embryos. Pak. Zool. J. 2014, 46, 167–175. [Google Scholar]
- Chakravarthi, V.P.; Kona, S.S.R.; Kumar, A.V.N.S.; Bhaskar, M.; Rao, V.H. Quantitative expression of antiapoptotic and proapoptotic genes in sheep ovarian follicles grown in vivo or cultured in vitro. Theriogenology 2015, 83, 590–595. [Google Scholar] [CrossRef]
- Ozawa, M.; Hirabayashi, M.; Kanai, Y. Developmental competence and oxidative state of mouse zygotes heat-stressed maternally or in vitro. Reprod. Camb. 2002, 124, 683–689. [Google Scholar] [CrossRef]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Dube, J.L.; Wang, P.; Elvin, J.; Lyons, K.M.; Celeste, A.J.; Matzuk, M.M. The bone morphogenetic protein 15 gene is X-linked and expressed in oocytes. Mol. Endocrinol. 1998, 12, 1809–1817. [Google Scholar] [CrossRef]
- Li, H.-K.; Kuo, T.-Y.; Yang, H.-S.; Chen, L.-R.; Li, S.S.-L.; Huang, H.-W. Differential gene expression of bone morphogenetic protein 15 and growth differentiation factor 9 during in vitro maturation of porcine oocytes and early embryos. Anim. Reprod. Sci. 2008, 103, 312–322. [Google Scholar] [CrossRef]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef] [PubMed]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Orisaka, M.; Orisaka, S.; Jiang, J.-Y.; Craig, J.; Wang, Y.; Kotsuji, F.; Tsang, B.K. Growth differentiation factor 9 is antiapoptotic during follicular development from preantral to early antral stage. Mol. Endocrinol. 2006, 20, 2456–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, T.S.; Froiland, D.A.; Amato, F.; Thompson, J.G.; Gilchrist, R.B. Oocytes prevent cumulus cell apoptosis by maintaining a morphogenic paracrine gradient of bone morphogenetic proteins. J. Cell Sci. 2005, 118, 5257–5268. [Google Scholar] [CrossRef] [Green Version]
- Bussmann, R.W.; Glenn, A. Medicinal plants used in Northern Peru for reproductive problems and female health. J. Ethnobiol. Ethnomed. 2010, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Alenezy, E.S.; Barakat, I.A.H.; Al Musayeib, N.M. Effect of wild marjoram (Origanum vulgare) plant extracts on capacitation of sheep spermatozoa in vitro. Adv. Biosci. Biotechnol. 2019, 10, 82–97. [Google Scholar] [CrossRef] [Green Version]
- Bendifallah, L.; Tchoulak, Y.; Djouabi, M.; Oukili, M.; Ghezraoui, R. Phytochemical study and antimicrobial activity of Origanum vulgare L. (Lamiaceae) in boumerdes mountainous region (Algeria). J. Med. Bioeng. Vol 2015, 4. [Google Scholar] [CrossRef]
- Saxena, M.; Saxena, J.; Pradhan, A. Flavonoids and phenolic acids as antioxidants in plants and human health. Int. J. Pharm. Sci. Rev. Res 2012, 16, 130–134. [Google Scholar]
- Abshenas, J.; Sakhaee, E.; Mostafavi, A. Effects of Origanum vulgare on epididymal sperm in mice after chronic copper sulfate administration. Online J. Vet. Res. 2014, 18, 859–868. [Google Scholar]
- Paderes, N.M.; Lagangilang, A.P. Antiangiogenic Activity of Oregano (Origanum Vulgare) Aqueous Extract In-Vivo through Chick Chorioallantoic Membrane (Cam) Assay. Syst. Rev. Pharm. 2020, 11, 1222–1227. [Google Scholar]
- Mbaye, M.M.; El Khalfi, B.; Ouzamode, S.; Saadani, B.; Louanjli, N.; Soukri, A. Effect of Origanum vulgare essential oil supplementation on the advanced parameters of mobility and on the integrity of human sperm DNA. Int. J. Reprod. Med. 2020, 2020, 1230274. [Google Scholar] [CrossRef] [PubMed]
- Dorman, H.J.D.; Peltoketo, A.; Hiltunen, R.; Tikkanen, M.J. Characterisation of the antioxidant properties of de-odourised aqueous extracts from selected Lamiaceae herbs. Food Chem. 2003, 83, 255–262. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Valiollahpoor Amiri, M.; Deldar, H.; Ansari Pirsaraei, Z. Impact of supplementary royal jelly on in vitro maturation of sheep oocytes: Genes involved in apoptosis and embryonic development. Syst. Biol. Reprod. Med. 2016, 62, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, S.S.; Celi, P.; Fahri, F.T.; Leury, B.J.; Dunshea, F.R. Dietary antioxidants at supranutritional doses modulate skeletal muscle heat shock protein and inflammatory gene expression in sheep exposed to heat stress. J. Anim. Sci. 2014, 92, 4897–4908. [Google Scholar] [CrossRef] [PubMed]
- Kyasari, O.R.; Valojerdi, M.R.; Farrokhi, A.; Ebrahimi, B. Expression of maturation genes and their receptors during in vitro maturation of sheep COCs in the presence and absence of somatic cells of cumulus origin. Theriogenology 2012, 77, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Rao, B.S.; Mahesh, Y.U.; Charan, K.V.; Suman, K.; Sekhar, N.; Shivaji, S. Effect of vitrification on meiotic maturation and expression of genes in immature goat cumulus oocyte complexes. Cryobiology 2012, 64, 176–184. [Google Scholar] [CrossRef]
- Wei, W.; Howard, P.S.; Kogan, B.; Macarak, E.J. Altered extracellular matrix expression in the diverted fetal sheep bladder. J. Urol. 2007, 178, 1104–1107. [Google Scholar] [CrossRef]
- Mishra, A.; Reddy, I.J.; Gupta, P.S.P.; Mondal, S. Expression of apoptotic and antioxidant enzyme genes in sheep oocytes and in vitro produced embryos. Anim. Biotechnol. 2017, 28, 18–25. [Google Scholar] [CrossRef]
- Germoush, M.O.; Al-Mutary, M.G.; Al-himaidi, A.R.; Al-Ghadi, M.G.; Iwamoto, D.; Al-anazi, Y.; Ammari, A.; Ahmad, J.; Al-Khedhairy, A. Effects of follicular fluid on developmental competence and gene expression of in vitro fertilized sheep embryos. Pak. J. Zool. 2018, 50. [Google Scholar] [CrossRef]
- Mishra, A.; Reddy, I.J.; Gupta, P.S.P.; Mondal, S. Developmental regulation and modulation of apoptotic genes expression in sheep oocytes and embryos cultured in vitro with L-carnitine. Reprod. Domest. Anim. 2016, 51, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Lee, E. Modification of maturation condition improves oocyte maturation and in vitro development of somatic cell nuclear transfer pig embryos. J. Vet. Sci. 2007, 8, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deleuze, S.; Goudet, G. Cysteamine supplementation of in vitro maturation media: A review. Reprod. Domest. Anim. 2010, 45, e476–e482. [Google Scholar] [CrossRef]
- Singhal, S.; Prasad, S.; Singh, B.; Prasad, J.K.; Gupta, H.P. Effect of including growth factors and antioxidants in maturation medium used for in vitro culture of buffalo oocytes recovered in vivo. Anim. Reprod. Sci. 2009, 113, 44–50. [Google Scholar] [CrossRef]
- Ali, A.A.; Bilodeau, J.F.; Sirard, M.A. Antioxidant requirements for bovine oocytes varies during in vitro maturation, fertilization and development. Theriogenology 2003, 59, 939–949. [Google Scholar] [CrossRef]
- Mukherjee, A.; Malik, H.; Saha, A.P.; Dubey, A.; Singhal, D.K.; Boateng, S.; Saugandhika, S.; Kumar, S.; De, S.; Guha, S.K. Resveratrol treatment during goat oocytes maturation enhances developmental competence of parthenogenetic and hand-made cloned blastocysts by modulating intracellular glutathione level and embryonic gene expression. J. Assist. Reprod. Genet. 2014, 31, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.T.; Nath, A.; Prasad, S.; Singhal, S.; Singh, N.; Gade, N.E.; Dubey, P.K.; Saikumar, G. Expression and characterization of constitutive heat shock protein 70.1 (HSPA-1A) gene in in vitro produced and in vivo-derived buffalo (Bubalus bubalis) embryos. Reprod. Domest. Anim. 2012, 47, 975–983. [Google Scholar] [CrossRef]
- Hendrey, J.; Kola, I. Thermolability of mouse oocytes is due to the lack of expression and/or inducibility of Hsp70. Mol. Reprod. Dev. 1991, 28, 1–8. [Google Scholar] [CrossRef]
- Duffy, D.M. Growth differentiation factor-9 is expressed by the primate follicle throughout the periovulatory interval. Biol. Reprod. 2003, 69, 725–732. [Google Scholar] [CrossRef]
- Robert, C.; Gagné, D.; Bousquet, D.; Barnes, F.L.; Sirard, M.-A. Differential display and suppressive subtractive hybridization used to identify granulosa cell messenger RNA associated with bovine oocyte developmental competence. Biol. Reprod. 2001, 64, 1812–1820. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Ulloa, S.M.; Heinzmann, J.; Herrmann, D.; Hadeler, K.-G.; Aldag, P.; Winkler, S.; Pache, D.; Baulain, U.; Lucas-Hahn, A.; Niemann, H. Cyclic AMP affects oocyte maturation and embryo development in prepubertal and adult cattle. PLoS ONE 2016, 11, e0150264. [Google Scholar] [CrossRef] [PubMed]
- Gode, F.; Gulekli, B.; Dogan, E.; Korhan, P.; Dogan, S.; Bige, O.; Cimrin, D.; Atabey, N. Influence of follicular fluid GDF9 and BMP15 on embryo quality. Fertil. Steril. 2011, 95, 2274–2278. [Google Scholar] [CrossRef] [PubMed]
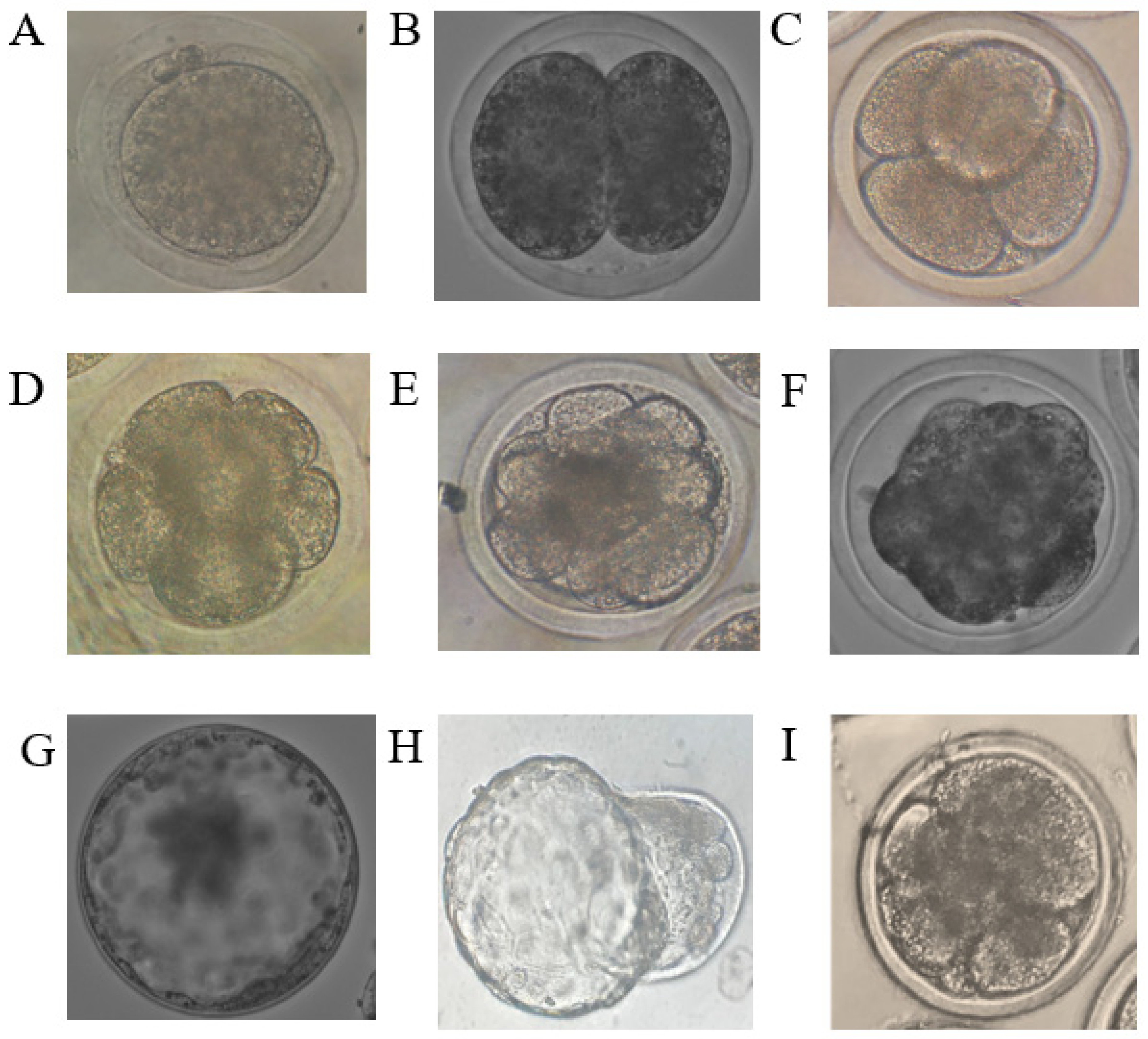

{kind=link}
| Function | Gene | Primer Sequence (5-3) Forward and Reverse | Fragment Size (bp) | GenBank Accession No. |
|---|---|---|---|---|
| Apoptosis | BCL-2 | F:ATGTGTTGGAGAGCGTCA R:AGAGACAGCCAGGAGAAATC | 182 | NM 001166486.1 [34] |
| Stress | HSP70 | F:AACATGAAGAGCGCCGTGGAGG R:GTTAACACCTGCTCCAGCTCC | 171 | NM 001267874.1 [35] |
| Nuclear maturation and embryonic development | GDF-9 | F:TAGTCAGCTGAAGTGGGACA R:AGCCATCAGGCTCGATGGCC | 224 | AF078545 [36] |
| TGFßR1 | F:GGTTCCGTGAAGCAGAGATT R:GACACCAACCAG AGCTGAGTC | 115 | AY656799 [37] | |
| EGR-1 | F:ACCAGTCCCAGCTCATCAA R:GAGCTCATCTGAGCGAGAGA | 134 | NM 001142506 [38] | |
| Reference gene | GAPDH | F:ATGGGCGTGAACCACGAGAA R: ATGGCGTGGACAGTGGTCAT | 146 | NM 001190390 [39] |
| Conc. Of O.V.E | Total of Oocyte | Nuclear Maturation (Mean ± SEM) | Deg. | ||||
|---|---|---|---|---|---|---|---|
| GV | GVBDV | M I | Anaphase I | M II | |||
| Control (0 µg/mL) | 151 | 9.27 ± 0.02 a (14) | 24.50 ± 0.03 a (37) | 5.96 ± 0.01 a (9) | 10.60 ± 0.02 a (16) | 43.71 ± 0.04 a (66) | 5.96 ± 0.01 a (9) |
| Treatment 1 (1 µg/mL) | 152 | 13.16 ± 0.02 a (20) | 11.84 ± 0.02 b (18) | 5.26 ± 0.01 a (8) | 13.82 ± 0.02 a (21) | 47.37 ± 0.04 a (72) | 8.55 ± 0.02 a (13) |
| Treatment 2 (10 µg/mL) | 152 | 7.89 ± 0.02 a (12) | 17.11 ± 0.03 ab (26) | 4.61 ± 0.0 a (7) | 9.87 ± 0.02 a (15) | 56.61 ± 0.04 a (83) | 5.92 ± 0.0 a (9) |
| Conc. Of O.V.E | Total of Oocyte | Nuclear Maturation )mean ± SEM( | Deg. | ||||
|---|---|---|---|---|---|---|---|
| GV | GVBDV | M I | Anaphase I | M II | |||
| Control (0 µg/mL) | 151 | 9.27 ± 0.02 a (14) | 24.50 ± 0.03 a (37) | 5.96 ± 0.01 a (9) | 10.60 ± 0.02 a (16) | 43.71 ± 0.04 a (66) | 5.96 ± 0.01a (9) |
| Treatment 1 (1 µg/mL) | 152 | 13.16 ± 0.02 a (20) | 11.84 ± 0.02 b (18) | 5.26 ± 0.01 a (8) | 13.82 ± 0.02 a (21) | 47.37 ± 0.04 a (72) | 8.55 ± 0.02 a (13) |
| Treatment 2 (10 µg/mL) | 152 | 7.89 ± 0.02 a (12) | 17.11 ± 0.03 ab (26) | 4.61 ± 0.0 a (7) | 9.87 ± 0.02 a (15) | 56.61 ± 0.04 a (83) | 5.92 ± 0.0 a (9) |
| Conc. of O.V.E | No, of Cutured Oocytes | One-Cell Stage No, Rate | 2-Cell stage No, Rate | 4-Cell Stage No, Rate | 8-Cell Stage No, Rate | 16-Cell Stage No, Rate | Morula Stage No, Rate | Cleavage No, Rate | Blastocyst No, Rate | Fragmented Embryos | Degenerated Oocytes |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control (0 µg/mL) | 353 | 66 18.70± 0.02 a | 1 0.28 ± 0.05 a | 1 0.28 ± 0.05 a | 3 0.85 ± 0.04 a | 6 1.70 ± 0.06 a | 78 22.10 ± 0.02 a | 108 30.59 ± 0.02 ab | 19 17.59 ± 0.04 a | 105 29.75 ± 0.02 a | 74 20.96 ± 0.02 b |
| Treatment 1 (1 µg/mL) | 341 | 64 19.35± 0.02 a | 2 0.59 ± 0.07 a | 1 0.29 ± 0.05 a | 1 0.29 ± 0.02 a | 4 1.17 ± 0.05 a | 84 25.22 ± 0.02 a | 106 31.08 ± 0.03 a | 16 14.95 ± 0.03 a | 104 30.50 ± 0.02 a | 65 19.65 ± 0.02 b |
| Treatment 2 (10 µg/mL) | 257 | 50 19.46 ± 0.02 a | 0 0 ± 0.00 a | 1 0.39 ± 0.06 a | 1 0.39 ± 0.03 a | 6 2.33 ± 0.09 a | 38 14.79± 0.02 b | 60 23.34 ± 0.03 b | 14 23.33 ± 0.05 a | 72 28.02 ± 0.02 a | 75 29.18 ± 0.02 a |
| The Fold Change in Gene Expression | |||||
|---|---|---|---|---|---|
| Conc. Of O.V.E | BCL2 | HSP70 | GDF9 | TGFRβ1 | EGR1 |
| Control (0 µg/mL) | 1.05 ± 0.20 c | 1.32 ± 0.70 a | 1.01 ± 0.12 a | 1.06 ± 0.26 b | 1.15 ± 0.36 b |
| Treatment 1 (1 µg/mL) | 9.57 ± 0.76 a | 0.61 ± 0.57 a | 0.41 ± 0.02 a | 1.36 ± 0.06 b | 1.82 ± 0.79 b |
| Treatment 2 (10 µg/mL) | 4.21 ± 0.48 b | 0.48 ± 0.06 a | 0.62 ± 0.36 a | 2.48 ± 0.13 a | 8.05 ± 2.84 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alotaibi, M.M.; Alhimaidi, A.R.; Al-Ghadi, M.Q.; Ammari, A.A.; Al-Malahi, N.M. Evaluation of the Maturity and Gene Expression of Sheep Oocytes and Embryos Cultured in Media Supplemented with Marjoram (Origanum vulgare) Extract. Genes 2022, 13, 1844. https://doi.org/10.3390/genes13101844
Alotaibi MM, Alhimaidi AR, Al-Ghadi MQ, Ammari AA, Al-Malahi NM. Evaluation of the Maturity and Gene Expression of Sheep Oocytes and Embryos Cultured in Media Supplemented with Marjoram (Origanum vulgare) Extract. Genes. 2022; 13(10):1844. https://doi.org/10.3390/genes13101844
Chicago/Turabian StyleAlotaibi, Mashael M., Ahmad R. Alhimaidi, Muath Q. Al-Ghadi, Aiman A. Ammari, and Nawal M. Al-Malahi. 2022. "Evaluation of the Maturity and Gene Expression of Sheep Oocytes and Embryos Cultured in Media Supplemented with Marjoram (Origanum vulgare) Extract" Genes 13, no. 10: 1844. https://doi.org/10.3390/genes13101844
APA StyleAlotaibi, M. M., Alhimaidi, A. R., Al-Ghadi, M. Q., Ammari, A. A., & Al-Malahi, N. M. (2022). Evaluation of the Maturity and Gene Expression of Sheep Oocytes and Embryos Cultured in Media Supplemented with Marjoram (Origanum vulgare) Extract. Genes, 13(10), 1844. https://doi.org/10.3390/genes13101844