
Altered Production and Cellular Levels of Hydrogen Sulfide (H2S) in Placental Trophoblasts from Pregnancies Affected by Pre-Eclampsia

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Placenta Collection
2.3. Human Placenta Trophoblast Cells Isolation and Culture
2.4. Free H2S Measurement
2.5. Immunohistochemistry
2.6. Protein Expression by Western Blot
2.7. Statistical Analysis
3. Results
3.1. Demographic and Clinical Data of Study Subjects
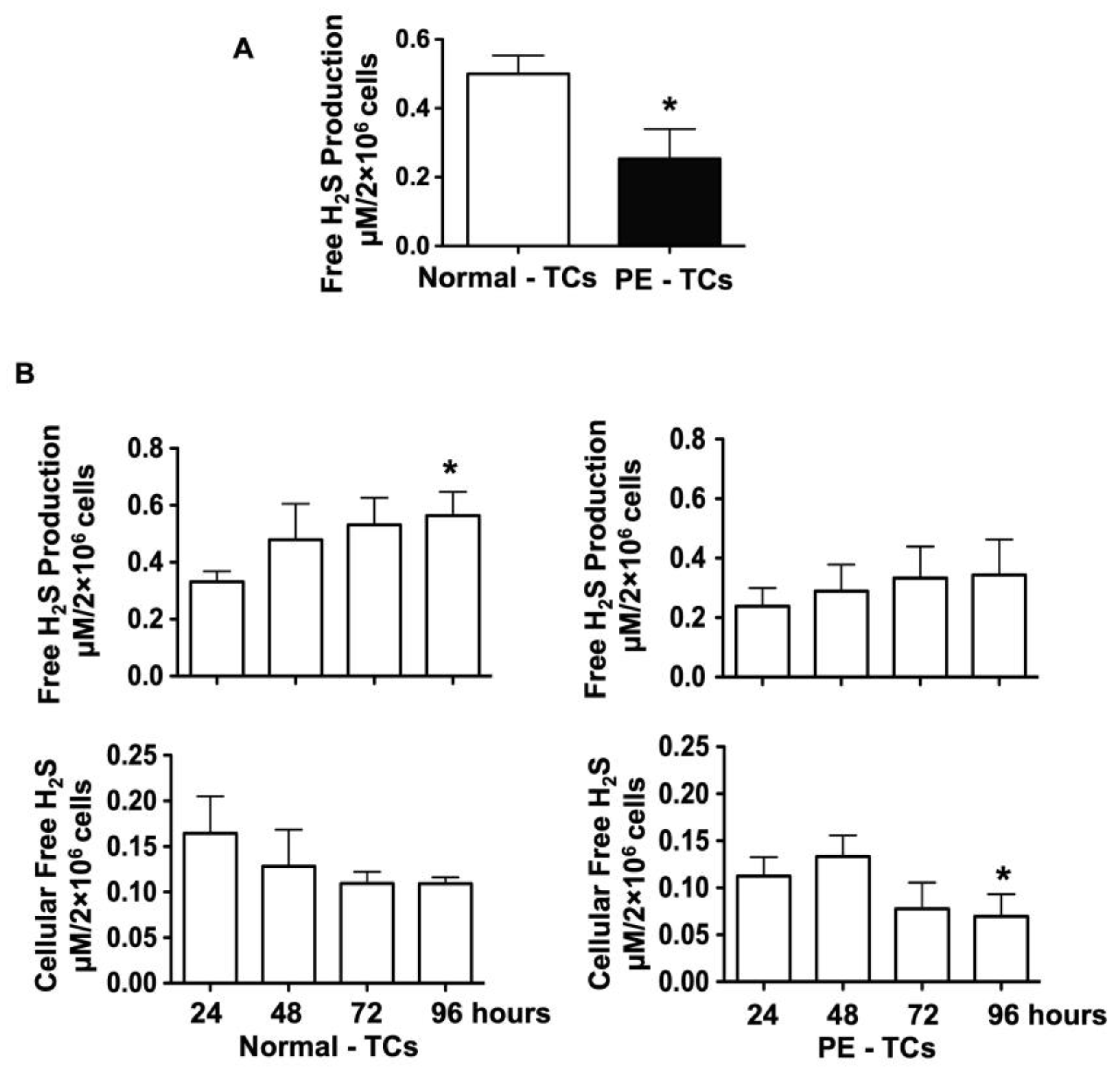
3.2. H2S Production and Cellular Levels in Trophoblasts from Normal and Pre-Eclamptic Pregnancies
3.3. Expression of CBS, CSE, and 3-MST in Placental Villous Tissue from Normal and Pre-Eclamptic Pregnancies
3.4. Downregulation of CBS and CSE Expression in Placental Trophoblasts from Pre-Eclamptic Pregnancies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roberts, J.M.; Hubel, C.A. The two stage model of preeclampsia: Variations on the theme. Placenta 2009, 30, S32–S37. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Walsh, S.W.; Kay, H.H. Placental lipid peroxides and thromboxane are increased and prostacyclin is decreased in women with preeclampsia. Am. J. Obstet. Gynecol. 1992, 167, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Walsh, S.W. Antioxidant activities and mRNA expression of superoxide dismutase, catalase, and glutathione peroxidase in normal and preeclamptic placentas. J. Soc. Gynecol. Investig. 1996, 3, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Walsh, S.W. Increased superoxide generation is associated with decreased superoxide dismutase activity and mRNA expression in placental trophoblast cells in preeclampsia. Placenta 2001, 22, 206–212. [Google Scholar] [CrossRef]
- Bowen, R.S.; Gu, Y.; Zhang, Y.; Lewis, D.F.; Wang, Y. Hypoxia promotes interleukin-6 and -8 but reduces interleukin-10 production by placental trophoblast cells from preeclamptic pregnancies. J. Soc. Gynecol. Investig. 2005, 12, 428–432. [Google Scholar] [CrossRef]
- Gude, N.M.; King, R.G.; Brennecke, S.P. Endothelin: Release by and potent constrictor effect on the fetal vessels of human perfused placental lobules. Reprod. Fertil. Dev. 1991, 3, 495–500. [Google Scholar] [CrossRef]
- Wang, Y.; Walsh, S.W.; Parnell, R.; Han, J. Placental production of nitric oxide and endothelin in normal and preeclamptic pregnancies. Hypertens. Pregnancy 1994, 13, 171–178. [Google Scholar] [CrossRef]
- Singh, S.; Padovani, D.; Leslie, R.A.; Chiku, T.; Banerjee, R. Relative contributions of cystathionine beta-synthase and gamma-cystathionase to H2S biogenesis via alternative trans-sulfuration reaction. J. Biol. Chem. 2009, 284, 22457–22466. [Google Scholar] [CrossRef]
- Rao, S.P.; Dobariya, P.; Bellamkonda, H.; More, S.S. Role of 3-mercaptopyruvate sulfurtransferase (3-MST) in physiology and disease. Antioxidants 2023, 12, 603. [Google Scholar] [CrossRef]
- Pattillo, R.A.; Gey, G.O. The establishment of a cell line of human hormone-synthesizing trophoblastic cells in vitro. Cancer Res. 1968, 28, 1231–1236. [Google Scholar]
- Chen, D.B.; Feng, L.; Hodges, J.K.; Lechuga, T.J.; Zhang, H. Human trophoblast-derived hydrogen sulfide stimulates placental artery endothelial cell angiogenesis. Biol. Reprod. 2017, 97, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Holwerda, K.M.; Bos, E.M.; Rajakumar, A.; Ris-Stalpers, C.; Van Pampus, M.G.; Timmer, A.; Erwich, J.J.H.M.; Faas, M.M.; Goor, H.; Lely, A.T. Hydrogen sulfide producing enzymes in pregnancy and preeclampsia. Placenta 2012, 33, 518–521. [Google Scholar] [CrossRef]
- Wang, K.; Ahmad, S.; Cai, M.; Rennie, J.; Fujisawa, T.; Crispi, F.; Baily, J.; Miller, M.R.; Cudmore, M.; Hadoke, P.W.F.; et al. Dysregulation of hydrogen sulfide producing enzyme cystathionine γ-lyase contributes to maternal hypertension and placental abnormalities in preeclampsia. Circulation 2013, 127, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Gu, Y.; Zhao, S.; Sun, J.; Groome, L.J.; Wang, Y. Expressions of vitamin D metabolic components VDBP, CYP2R1, CYP27B1, CYP24A1, and VDR in placentas from normal and preeclamptic pregnancies. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E928–E935. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Kolluru, G.K.; Yuan, S.; Kevil, C.G. Measurement of H2S in vivo and in vitro by the monobromobimane method. Methods Enzymol. 2015, 554, 31–45. [Google Scholar]
- Gu, Y.; Chu, X.; Morgan, J.A.; Lewis, D.F.; Wang, Y. Upregulation of METTL3 expression and m6A RNA methylation in placental trophoblasts in preeclampsia. Placenta 2021, 103, 43–49. [Google Scholar] [CrossRef]
- Li, Z.; Polhemus, D.J.; Lefer, D.J. Evolution of Hydrogen Sulfide Therapeutics to Treat Cardiovascular Disease. Circ. Res. 2018, 123, 590–600. [Google Scholar] [CrossRef]
- Kanagy, N.L.; Szabo, C.; Papapetropoulos, A. Vascular biology of hydrogen sulfide. Am. J. Physiol. Cell Physiol. 2017, 312, C537–C549. [Google Scholar] [CrossRef]
- Bibli, S.I.; Hu, J.; Sigala, F.; Wittig, I.; Heidler, J.; Zukunft, S.; Tsilimigras, D.I.; Randriamboavonjy, V.; Wittig, J.; Kojonazarov, B.; et al. Cystathionine γ Lyase Sulfhydrates the RNA Binding Protein Human Antigen R to Preserve Endothelial Cell Function and Delay Atherogenesis. Circulation 2019, 139, 101–114. [Google Scholar] [CrossRef]
- Kuang, Q.; Xue, N.; Chen, J.; Shen, Z.; Cui, X.; Fang, Y.; Ding, X. Low plasma hydrogen sulfide is associated with impaired renal function and cardiac dysfunction. Am. J. Nephrol. 2018, 47, 361–371. [Google Scholar] [CrossRef]
- Greaney, J.L.; Kutz, J.L.; Shank, S.W.; Jandu, S.; Santhanam, L.; Alexander, L.M. Impaired hydrogen sulfide-mediated vasodilation contributes to microvascular endothelial dysfunction in hypertensive adults. Hypertension 2017, 69, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, P.; Gou, Q.; Jin, S.; Xue, H.; Li, D.; Tian, D.; Sun, J.; Zhang, X.; Teng, X.; et al. Hydrogen sulfide ameliorated preeclampsia via suppression of toll-like receptor 4-activated inflammation in the rostral ventrolateral medulla of rats. Biomed. Pharmacother. 2022, 150, 113018. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, B.; Gordon, L.; Wong, N.C.; Moffett, A.; Manuelpillai, U.; Craig, J.M.; Sharkey, A.; Saffery, R. Wide-ranging DNA Methylation Differences of Primary Trophoblast Cell Populations and Derived Cell Lines: Implications and Opportunities for Understanding Trophoblast Function. Mol. Hum. Reprod. 2011, 17, 344–353. [Google Scholar] [CrossRef]
- Morales-Prieto, D.M.; Chaiwangyen, W.; Ospina-Prieto, S.; Schneider, U.; Herrmann, J.; Gruhn, B.; Markert, U.R. MicroRNA expression profiles of trophoblastic cells. Placenta 2012, 33, 725–734. [Google Scholar] [CrossRef]
- Bazhanov, N.; Ansar, M.; Ivanciuc, T.; Garofalo, R.P.; Casola, A. Hydrogen sulfide: A novel player in airway development, pathophysiology of respiratory diseases, and antiviral defenses. Am. J. Respir. Cell Mol. Biol. 2017, 57, 403–410. [Google Scholar] [CrossRef]
- Tomita, M.; Nagahara, N.; Ito, T. Expression of 3-mercaptopyruvate sulfurtransferase in the mouse. Molecules 2016, 21, 1707. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, N.; Ito, T.; Kitamura, H.; Nishino, T. Tissue and subcellular distribution of mercaptopyruvate sulfurtransferase in the rat: Confocal laser fluorescence and immunoelectron microscopic studies combined with biochemical analysis. Histochem. Cell Biol. 1998, 110, 243–250. [Google Scholar] [CrossRef]
- Huerta de la Cruz, S.; Rodríguez-Palma, E.J.; Santiago-Castañeda, C.L.; Beltrán-Ornelas, J.H.; Sánchez-López, A.; Rocha, L.; Centurión, D. Exogenous hydrogen sulfide restores CSE and CBS but no 3-MST protein expression in the hypothalamus and brainstem after severe traumatic brain injury. Metab. Brain Dis. 2022, 37, 1863–1874. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Variables | Normal (n = 18) | Pre-Eclampsia (n = 12) | p Value |
|---|---|---|---|
| Maternal Age | 29 ± 6 | 29 ± 7 | 0.938 |
| Racial Status ¶ White Black Other | |||
| 5 12 | 2 9 | 0.6693 | |
| 1 | 1 | ||
| BMI | 36.4 ± 6.2 | 37.9 ± 9.5 | 0.593 |
| Blood Pressure (mmHg) Systolic Diastolic | 120 ± 9 67 ± 9 | 171 ± 12 99 ± 15 | <0.001 <0.001 |
| Primigravida ¶ | 2 | 3 | 0.3644 |
| Gestational Age (weeks+days) at Delivery | 38+3 ± 1+2 | 33+0 ± 3+4 | <0.001 |
| Delivery Mode ¶ Vaginal Delivery C-Section | |||
| 8 | 3 | 0.2789 | |
| 10 | 9 | ||
| Fetal Gender ¶ Male Female | 7 11 | 6 6 | 0.7106 |
| Baby Weight (gram) | 3341 ± 565 | 1832 ± 778 | <0.001 |
| Placental Weight (gram) | 619 ± 131 | 402 ± 137 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, X.; Xu, J.; Shen, X.; Sheng, W.; Sun, J.; Gu, Y.; Lewis, D.F.; Cooper, D.; Zoorob, D.; Wang, Y. Altered Production and Cellular Levels of Hydrogen Sulfide (H2S) in Placental Trophoblasts from Pregnancies Affected by Pre-Eclampsia. Pathophysiology 2025, 32, 10. https://doi.org/10.3390/pathophysiology32010010
Chu X, Xu J, Shen X, Sheng W, Sun J, Gu Y, Lewis DF, Cooper D, Zoorob D, Wang Y. Altered Production and Cellular Levels of Hydrogen Sulfide (H2S) in Placental Trophoblasts from Pregnancies Affected by Pre-Eclampsia. Pathophysiology. 2025; 32(1):10. https://doi.org/10.3390/pathophysiology32010010
Chicago/Turabian StyleChu, Xiaodan, Jie Xu, Xinggui Shen, Wenji Sheng, Jingxia Sun, Yang Gu, David F. Lewis, Danielle Cooper, Dani Zoorob, and Yuping Wang. 2025. "Altered Production and Cellular Levels of Hydrogen Sulfide (H2S) in Placental Trophoblasts from Pregnancies Affected by Pre-Eclampsia" Pathophysiology 32, no. 1: 10. https://doi.org/10.3390/pathophysiology32010010
APA StyleChu, X., Xu, J., Shen, X., Sheng, W., Sun, J., Gu, Y., Lewis, D. F., Cooper, D., Zoorob, D., & Wang, Y. (2025). Altered Production and Cellular Levels of Hydrogen Sulfide (H2S) in Placental Trophoblasts from Pregnancies Affected by Pre-Eclampsia. Pathophysiology, 32(1), 10. https://doi.org/10.3390/pathophysiology32010010