
Gut Microbiome Interactions with Oxidative Stress: Mechanisms and Consequences for Health

 ,
,  ,
,  and
and
Abstract
1. Introduction
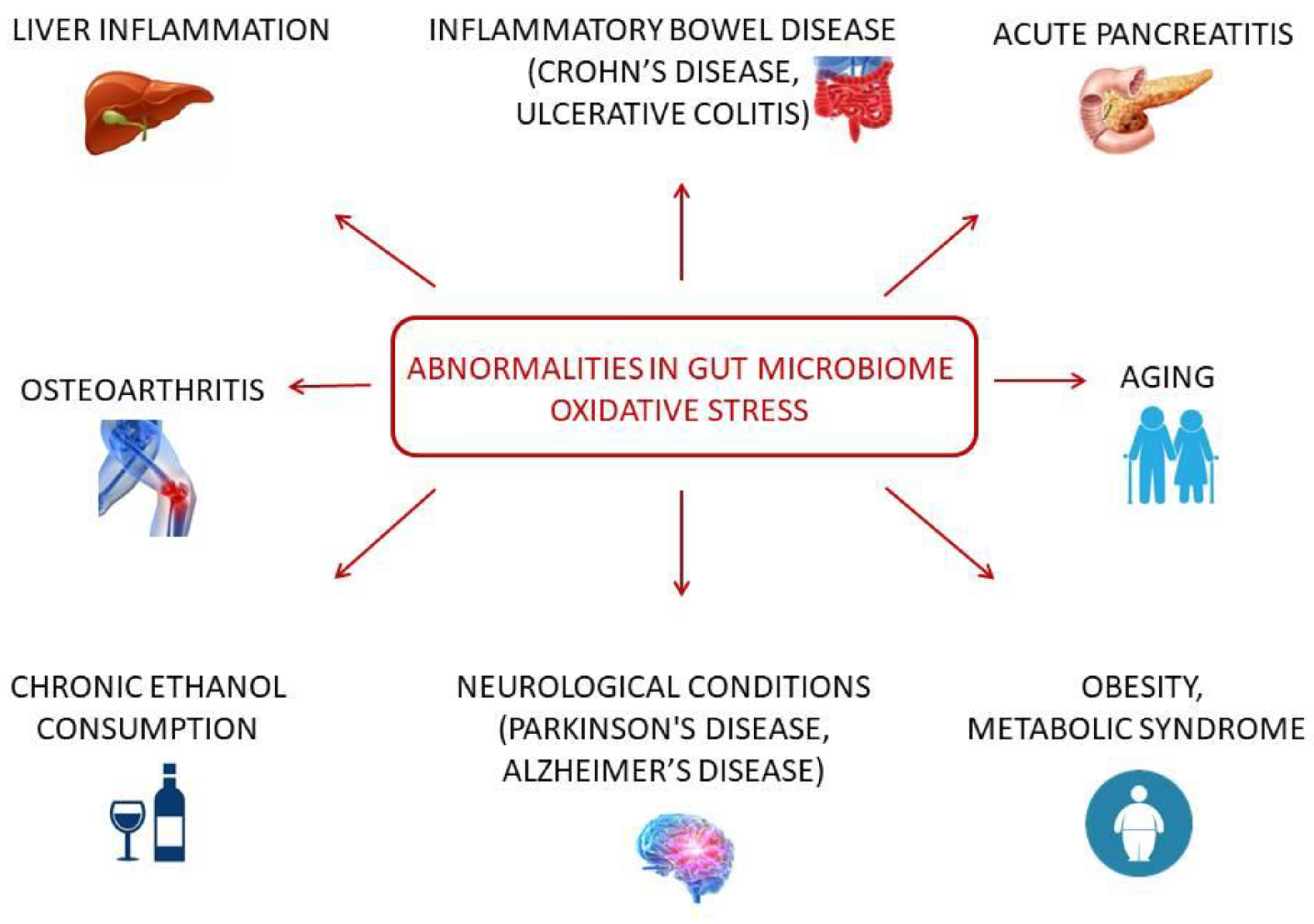
2. The Cooperation of the Microbiome and OS in Disease Development and Other Conditions
3. Possible Interconnections between OS and the Microbiome
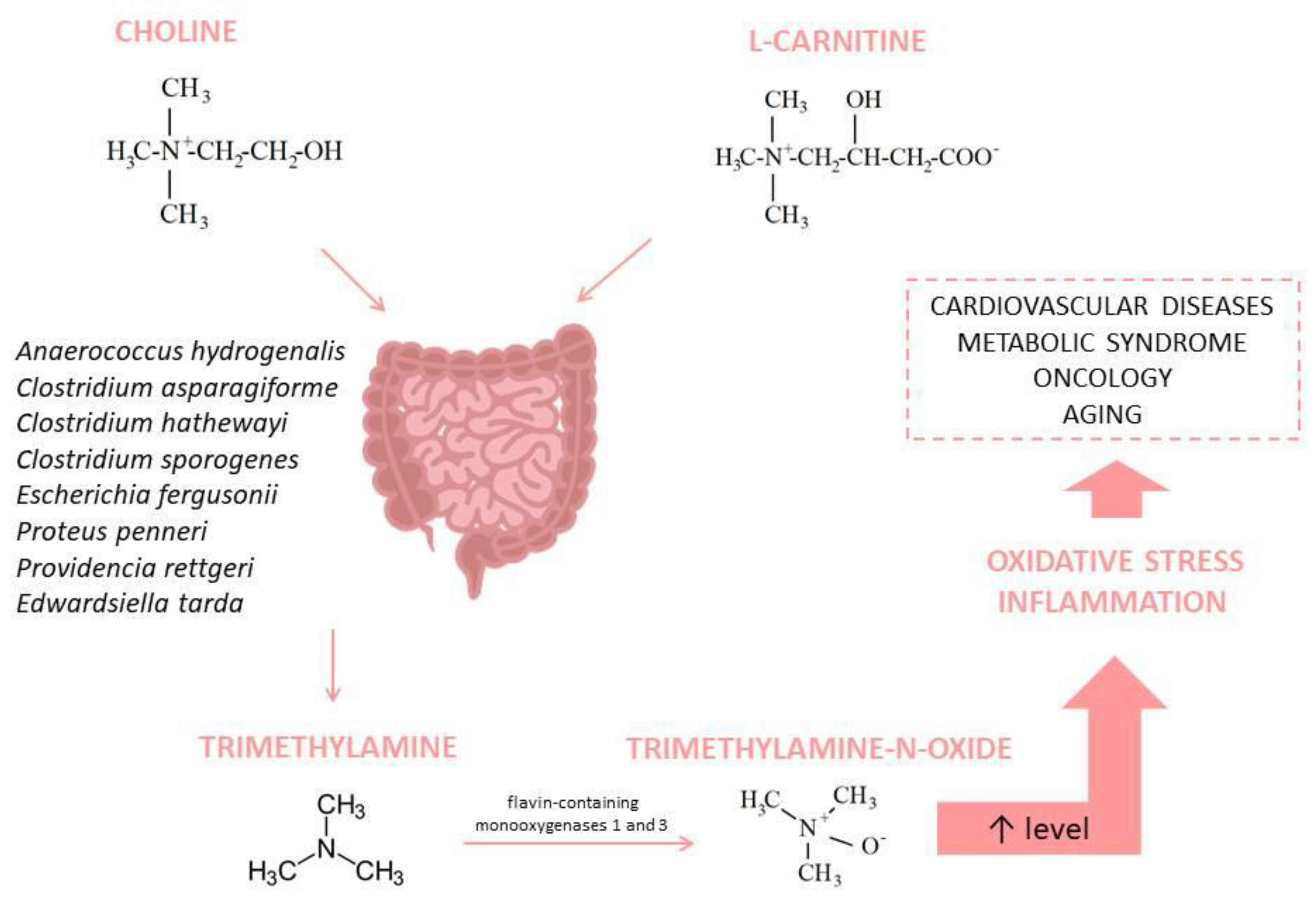
3.1. Trimethylamine-N-Oxide (TMAO)
3.2. Mitochondria and Short-Chain Fatty Acids
3.3. Melatonin
4. Impact of Food Supplements on the Gut Microbiome and OS
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rose, S.; Bennuri, S.C.; Davis, J.E.; Wynne, R.; Slattery, J.C.; Tippett, M.; Delhey, L.; Melnyk, S.; Kahler, S.G.; MacFabe, D.F.; et al. Butyrate Enhances Mitochondrial Function during Oxidative Stress in Cell Lines from Boys with Autism. Transl. Psychiatry 2018, 8, 42. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of Mammals and Their Gut Microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The Gut Microbiome as a Modulator of Healthy Ageing. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 565–584. [Google Scholar] [CrossRef]
- Xu, F.; Fu, Y.; Sun, T.; Jiang, Z.; Miao, Z.; Shuai, M.; Gou, W.; Ling, C.; Yang, J.; Wang, J.; et al. The Interplay between Host Genetics and the Gut Microbiome Reveals Common and Distinct Microbiome Features for Complex Human Diseases. Microbiome 2020, 8, 145. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Das, M.; Jeffery, I.B.; O’Toole, P.W. Adjusting for Age Improves Identification of Gut Microbiome Alterations in Multiple Diseases. eLife 2020, 9, e50240. [Google Scholar] [CrossRef]
- Almeida, A.; Mitchell, A.L.; Boland, M.; Forster, S.C.; Gloor, G.B.; Tarkowska, A.; Lawley, T.D.; Finn, R.D. A New Genomic Blueprint of the Human Gut Microbiota. Nature 2019, 568, 499–504. [Google Scholar] [CrossRef]
- Peters, B.; Santoro, N.; Kaplan, R.; Qi, Q. Spotlight on the Gut Microbiome in Menopause: Current Insights. Int. J. Women’s Health 2022, 14, 1059–1072. [Google Scholar] [CrossRef]
- Kedia, S.; Ahuja, V. Human Gut Microbiome: A Primer for the Clinician. JGH Open 2023, 7, 337–350. [Google Scholar] [CrossRef]
- Konopelski, P.; Mogilnicka, I. Biological Effects of Indole-3-Propionic Acid, a Gut Microbiota-Derived Metabolite, and Its Precursor Tryptophan in Mammals’ Health and Disease. Int. J. Mol. Sci. 2022, 23, 1222. [Google Scholar] [CrossRef]
- Wang, J.; Dong, P.; Zheng, S.; Mai, Y.; Ding, J.; Pan, P.; Tang, L.; Wan, Y.; Liang, H. Advances in Gut Microbiome in Metabonomics Perspective: Based on Bibliometrics Methods and Visualization Analysis. Front. Cell. Infect. Microbiol. 2023, 13, 1196967. [Google Scholar] [CrossRef]
- Barko, P.C.; McMichael, M.A.; Swanson, K.S.; Williams, D.A. The Gastrointestinal Microbiome: A Review. Vet. Intern. Med. 2018, 32, 9–25. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Li, L.; Zhong, C.; Zhang, Y.; Yang, X.; Li, M.; Yang, C. The Role of Gut Microbiota in Intestinal Disease: From an Oxidative Stress Perspective. Front. Microbiol. 2024, 15, 1328324. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: A Concept in Redox Biology and Medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative Stress and Autophagy: The Clash Between Damage and Metabolic Needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef]
- Vatner, S.F.; Zhang, J.; Oydanich, M.; Berkman, T.; Naftalovich, R.; Vatner, D.E. Healthful Aging Mediated by Inhibition of Oxidative Stress. Ageing Res. Rev. 2020, 64, 101194. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Sallam, R.M. Reactive Oxygen Species in Health and Disease. J. Biomed. Biotechnol. 2012, 2012, 936486. [Google Scholar] [CrossRef]
- Wei, Y.-H.; Lee, H.-C. Oxidative Stress, Mitochondrial DNA Mutation, and Impairment of Antioxidant Enzymes in Aging. Exp. Biol. Med. (Maywood) 2002, 227, 671–682. [Google Scholar] [CrossRef]
- Zhao, J.; Tian, F.; Yan, S.; Zhai, Q.; Zhang, H.; Chen, W. Lactobacillus plantarum CCFM10 Alleviating Oxidative Stress and Restoring the Gut Microbiota in d-Galactose-Induced Aging Mice. Food Funct. 2018, 9, 917–924. [Google Scholar] [CrossRef]
- Darenskaya, M.; Kolesnikov, S.; Semenova, N.; Kolesnikova, L. Diabetic Nephropathy: Significance of Determining Oxidative Stress and Opportunities for Antioxidant Therapies. Int. J. Mol. Sci. 2023, 24, 12378. [Google Scholar] [CrossRef]
- Semenova, N.V.; Madaeva, I.M.; Brichagina, A.S.; Kolesnikov, S.I.; Kolesnikova, L.I. 8-Hydroxy-2′-Deoxyguanosine as an Oxidative Stress Marker in Insomnia. Bull. Exp. Biol. Med. 2021, 171, 384–387. [Google Scholar] [CrossRef]
- Darenskaya, M.A.; Belenkaya, L.V.; Atalyan, A.V.; Danusevich, I.N.; Lazareva, L.M.; Nadelyaeva, Y.G.; Kolesnikova, L.I. Oxidative Stress Reactions in Women of Reproductive Age with Metabolic Syndrome. Bull. Exp. Biol. Med. 2023, 174, 601–604. [Google Scholar] [CrossRef]
- Zhang, K.; Ping, L.; Du, T.; Wang, Y.; Sun, Y.; Liang, G.; Wang, X.; Xie, X.; Wei, W.; Xiao, X.; et al. A Novel Systematic Oxidative Stress Score Predicts the Prognosis of Patients with Operable Breast Cancer. Oxidative Med. Cell. Longev. 2021, 2021, 9441896. [Google Scholar] [CrossRef]
- Ünal, E.D.; Vural, G.; Eren, F.; Neşelioğlu, S.; Erel, Ö. Clinicoradiological Evaluation of Oxidative Stress Activity in Acute Cerebral Infarction in the First 24 h and the Qualitative Importance of Dysfunctional HDL in Stroke. Turk. J. Med. Sci. 2022, 52, 1917–1925. [Google Scholar] [CrossRef]
- Shanahan, F.; Ghosh, T.S.; O’Toole, P.W. The Healthy Microbiome—What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. Toward an Improved Definition of a Healthy Microbiome for Healthy Aging. Nat Aging 2022, 2, 1054–1069. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Targeting Oxidative Stress in Disease: Promise and Limitations of Antioxidant Therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Aljeradat, B.; Kumar, D.; Abdulmuizz, S.; Kundu, M.; Almealawy, Y.F.; Batarseh, D.R.; Atallah, O.; Ennabe, M.; Alsarafandi, M.; Alan, A.; et al. Neuromodulation and the Gut–Brain Axis: Therapeutic Mechanisms and Implications for Gastrointestinal and Neurological Disorders. Pathophysiology 2024, 31, 244–268. [Google Scholar] [CrossRef]
- Zhao, W.; Kodancha, P.; Das, S. Gut Microbiome Changes in Anorexia Nervosa: A Comprehensive Review. Pathophysiology 2024, 31, 68–88. [Google Scholar] [CrossRef]
- Tang, J.; Song, X.; Zhao, M.; Chen, H.; Wang, Y.; Zhao, B.; Yu, S.; Ma, T.; Gao, L. Oral Administration of Live Combined Bacillus Subtilis and Enterococcus Faecium Alleviates Colonic Oxidative Stress and Inflammation in Osteoarthritic Rats by Improving Fecal Microbiome Metabolism and Enhancing the Colonic Barrier. Front. Microbiol. 2022, 13, 1005842. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.; Elsabagh, M.; Zhang, Y.; Ma, Y.; Jin, Y.; Wang, M.; Wang, H.; Jiang, H. Effects of the Gut Microbiota and Barrier Function on Melatonin Efficacy in Alleviating Liver Injury. Antioxidants 2022, 11, 1727. [Google Scholar] [CrossRef]
- Mei, Q.; Hu, J.; Huang, Z.; Fan, J.; Huang, C.; Lu, Y.; Wang, X.; Zeng, Y. Pretreatment with Chitosan Oligosaccharides Attenuate Experimental Severe Acute Pancreatitis via Inhibiting Oxidative Stress and Modulating Intestinal Homeostasis. Acta Pharmacol. Sin. 2021, 42, 942–953. [Google Scholar] [CrossRef]
- Wan, F.; Wang, M.; Zhong, R.; Chen, L.; Han, H.; Liu, L.; Zhao, Y.; Lv, H.; Hou, F.; Yi, B.; et al. Supplementation With Chinese Medicinal Plant Extracts From Lonicera Hypoglauca and Scutellaria Baicalensis Mitigates Colonic Inflammation by Regulating Oxidative Stress and Gut Microbiota in a Colitis Mouse Model. Front. Cell. Infect. Microbiol. 2022, 11, 798052. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yan, A.; Liu, X.; Ma, Y.; Zhao, F.; Wang, M.; Loor, J.J.; Wang, H. Melatonin Ameliorates Ochratoxin A Induced Liver Inflammation, Oxidative Stress and Mitophagy in Mice Involving in Intestinal Microbiota and Restoring the Intestinal Barrier Function. J. Hazard. Mater. 2021, 407, 124489. [Google Scholar] [CrossRef]
- Gao, T.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin Ameliorates Corticosterone-Mediated Oxidative Stress-Induced Colitis in Sleep-Deprived Mice Involving Gut Microbiota. Oxidative Med. Cell. Longev. 2021, 2021, 9981480. [Google Scholar] [CrossRef]
- Li, H.; Wei, Y.; Li, X.; Zhang, S.; Zhang, R.; Li, J.; Ma, B.; Shao, S.; Lv, Z.; Ruan, H.; et al. Diosmetin Has Therapeutic Efficacy in Colitis Regulating Gut Microbiota, Inflammation, and Oxidative Stress via the Circ-Sirt1/Sirt1 Axis. Acta Pharmacol. Sin. 2022, 43, 919–932. [Google Scholar] [CrossRef]
- Bruner, L.P.; White, A.M.; Proksell, S. Inflammatory Bowel Disease. Prim. Care 2023, 50, 411–427. [Google Scholar] [CrossRef]
- Alemany-Cosme, E.; Sáez-González, E.; Moret, I.; Mateos, B.; Iborra, M.; Nos, P.; Sandoval, J.; Beltrán, B. Oxidative Stress in the Pathogenesis of Crohn’s Disease and the Interconnection with Immunological Response, Microbiota, External Environmental Factors, and Epigenetics. Antioxidants 2021, 10, 64. [Google Scholar] [CrossRef]
- Annese, V. Genetics and Epigenetics of IBD. Pharmacol. Res. 2020, 159, 104892. [Google Scholar] [CrossRef]
- Hendrix, J.; Nijs, J.; Ickmans, K.; Godderis, L.; Ghosh, M.; Polli, A. The Interplay between Oxidative Stress, Exercise, and Pain in Health and Disease: Potential Role of Autonomic Regulation and Epigenetic Mechanisms. Antioxidants 2020, 9, 1166. [Google Scholar] [CrossRef]
- Nikitina, O.A.; Darenskaya, M.A.; Semenova, N.V.; Kolesnikova, L.I. Antioxidant Defense System: Regulation of Metabolic Processes, Genetic Determinants, and Methods of Determination. Cell Tiss. Biol. 2023, 17, 16–24. [Google Scholar] [CrossRef]
- Jurcau, A. Insights into the Pathogenesis of Neurodegenerative Diseases: Focus on Mitochondrial Dysfunction and Oxidative Stress. Int. J. Mol. Sci. 2021, 22, 11847. [Google Scholar] [CrossRef] [PubMed]
- Balmus, I.; Ciobica, A.; Trifan, A.; Stanciu, C. The Implications of Oxidative Stress and Antioxidant Therapies in Inflammatory Bowel Disease: Clinical Aspects and Animal Models. Saudi J. Gastroenterol. 2016, 22, 3. [Google Scholar] [CrossRef]
- Kruidenier, L.; Kuiper, I.; Lamers, C.B.; Verspaget, H.W. Intestinal Oxidative Damage in Inflammatory Bowel Disease: Semi-quantification, Localization, and Association with Mucosal Antioxidants. J. Pathol. 2003, 201, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, X.; Zhang, S.; Qi, C.; Zhang, Z.; Ma, R.; Xiang, L.; Chen, L.; Zhu, Y.; Tang, C.; et al. Oxidative Stress Gene Expression, DNA Methylation, and Gut Microbiota Interaction Trigger Crohn’s Disease: A Multi-Omics Mendelian Randomization Study. BMC Med. 2023, 21, 179. [Google Scholar] [CrossRef] [PubMed]
- Dunnill, C.J.; Ibraheem, K.; Mohamed, A.; Southgate, J.; Georgopoulos, N.T. A Redox State-dictated Signalling Pathway Deciphers the Malignant Cell Specificity of CD40-mediated Apoptosis. Oncogene 2017, 36, 2515–2528. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.; Hausenloy, D.J.; Andreadou, I.; Horman, S.; Bertrand, L.; Beauloye, C. AMP-Activated Protein Kinase: A Remarkable Contributor to Preserve a Healthy Heart against ROS Injury. Free. Radic. Biol. Med. 2021, 166, 238–254. [Google Scholar] [CrossRef] [PubMed]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin. Microbiol. Rev. 2022, 35, e00338-20. [Google Scholar] [CrossRef]
- Shandilya, S.; Kumar, S.; Kumar Jha, N.; Kumar Kesari, K.; Ruokolainen, J. Interplay of Gut Microbiota and Oxidative Stress: Perspective on Neurodegeneration and Neuroprotection. J. Adv. Res. 2022, 38, 223–244. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Li, B.; He, B.; Li, L.; Nice, E.C.; Zhang, W.; Xu, J. New Insights into the Gut Microbiota in Neurodegenerative Diseases from the Perspective of Redox Homeostasis. Antioxidants 2022, 11, 2287. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, Z.; Zhao, L.; Zhao, Y.; Yang, G.; Wang, C.; Gao, L.; Niu, C.; Li, S. Lactobacillus plantarum DP189 Reduces α-SYN Aggravation in MPTP-Induced Parkinson’s Disease Mice via Regulating Oxidative Damage, Inflammation, and Gut Microbiota Disorder. J. Agric. Food Chem. 2022, 70, 1163–1173. [Google Scholar] [CrossRef]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef]
- Cobb, C.A.; Cole, M.P. Oxidative and Nitrative Stress in Neurodegeneration. Neurobiol. Dis. 2015, 84, 4–21. [Google Scholar] [CrossRef]
- Pan, S.; Wei, H.; Yuan, S.; Kong, Y.; Yang, H.; Zhang, Y.; Cui, X.; Chen, W.; Liu, J.; Zhang, Y. Probiotic Pediococcus Pentosaceus Ameliorates MPTP-Induced Oxidative Stress via Regulating the Gut Microbiota–Gut–Brain Axis. Front. Cell. Infect. Microbiol. 2022, 12, 1022879. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Kaul, R.; Abdellatif, B.; Arabi, M.; Upadhyay, R.; Saliba, R.; Sebah, M.; Chaari, A. The Promising Role of Microbiome Therapy on Biomarkers of Inflammation and Oxidative Stress in Type 2 Diabetes: A Systematic and Narrative Review. Front. Nutr. 2022, 9, 906243. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Yan, Y.; Ji, F.; Zhou, H. Maternal Obesity Increases Oxidative Stress in Placenta and It Is Associated with Intestinal Microbiota. Front. Cell. Infect. Microbiol. 2021, 11, 671347. [Google Scholar] [CrossRef] [PubMed]
- Klimenko, E.S.; Belkova, N.L.; Romanitsa, A.I.; Pogodina, A.V.; Rychkova, L.V.; Darenskaya, M.A. Differences in Gut Microbiota Composition and Predicted Metabolic Functions: A Pilot Study of Adolescents with Normal Weight and Obesity. Bull. Exp. Biol. Med 2022, 173, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Darenskaya, M.; Rychkova, L.; Semenova, N.; Belkova, N.; Kolesnikova, L. The Role of Oxidative Stress and Changes in the Composition of the Gut Microbiota in the Genesis of Adolescent Obesity. Int. J. Biol. Macromol. 2022, 12, 344–348. [Google Scholar] [CrossRef]
- Darenskaya, M.; Gavrilova, O.; Rychkova, L.; Kravtsova, O.; Grebenkina, L.; Osipova, E.; Kolesnikov, S.; Kolesnikova, L. The Assessment of Oxidative Stress Intensity in Adolescents with Obesity by the Integral Index. Int. J. Biol. Macromol. 2018, 8, 37–41. [Google Scholar] [CrossRef]
- Shantir’, I.I.; Rodionov, G.G.; Fominykh, Y.A.; Batskov, S.S.; Ushal, I.E.; Kolobova, E.A.; Svetkina, E.V.; Sannikov, M.V. Intestinal Microbiota and Oxidative Stress in Patients with Metabolic Syndrome. Ekol. Cheloveka (Hum. Ecol.) 2019, 26, 23–29. [Google Scholar] [CrossRef]
- Ohira, H.; Tsuruya, A.; Oikawa, D.; Nakagawa, W.; Mamoto, R.; Hattori, M.; Waki, T.; Takahashi, S.; Fujioka, Y.; Nakayama, T. Alteration of Oxidative-Stress and Related Marker Levels in Mouse Colonic Tissues and Fecal Microbiota Structures with Chronic Ethanol Administration: Implications for the Pathogenesis of Ethanol-Related Colorectal Cancer. PLoS ONE 2021, 16, e0246580. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhou, H.; Deng, L.; Wang, L.; Chen, J.; Zhou, X. Serine Deficiency Exacerbates Inflammation and Oxidative Stress via Microbiota-Gut-Brain Axis in D-Galactose-Induced Aging Mice. Mediat. Inflamm. 2020, 2020, 5821428. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Cheng, Y.; Su, W.; Wang, C.; Lu, Z.; Jin, M.; Wang, F.; Wang, Y. Pediococcus Pentosaceus ZJUAF-4 Relieves Oxidative Stress and Restores the Gut Microbiota in Diquat-Induced Intestinal Injury. Appl. Microbiol. Biotechnol. 2021, 105, 1657–1668. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-L.; Zhao, Y.-C.; Zhu, H.-Y.; Wu, M.; Zheng, Y.-N.; Yang, M.; Cheng, Z.-Q.; Ding, C.-B.; Liu, W.-C. Taxifolin Retards the d-Galactose-Induced Aging Process through Inhibiting Nrf2-Mediated Oxidative Stress and Regulating the Gut Microbiota in Mice. Food Funct. 2021, 12, 12142–12158. [Google Scholar] [CrossRef] [PubMed]
- Tsuruya, A.; Kuwahara, A.; Saito, Y.; Yamaguchi, H.; Tsubo, T.; Suga, S.; Inai, M.; Aoki, Y.; Takahashi, S.; Tsutsumi, E.; et al. Ecophysiological Consequences of Alcoholism on Human Gut Microbiota: Implications for Ethanol-Related Pathogenesis of Colon Cancer. Sci. Rep. 2016, 6, 27923. [Google Scholar] [CrossRef] [PubMed]
- Kansakar, U.; Trimarco, V.; Mone, P.; Varzideh, F.; Lombardi, A.; Santulli, G. Choline Supplements: An Update. Front. Endocrinol. 2023, 14, 1148166. [Google Scholar] [CrossRef] [PubMed]
- Romano, K.A.; Vivas, E.I.; Amador-Noguez, D.; Rey, F.E. Intestinal Microbiota Composition Modulates Choline Bioavailability from Diet and Accumulation of the Proatherogenic Metabolite Trimethylamine-N-Oxide. mBio 2015, 6, e02481-14. [Google Scholar] [CrossRef] [PubMed]
- Day-Walsh, P.; Shehata, E.; Saha, S.; Savva, G.M.; Nemeckova, B.; Speranza, J.; Kellingray, L.; Narbad, A.; Kroon, P.A. The Use of An In-vitro Batch Fermentation (Human Colon) Model for Investigating Mechanisms of TMA Production From Choline, L-carnitine and Related Precursors by The Human Gut Microbiota. Eur. J. Nutr. 2021, 60, 3987–3999. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Fletcher, C. Trimethylamine N-oxide: Breathe New Life. Br. J. Pharmacol. 2018, 175, 1344–1353. [Google Scholar] [CrossRef]
- Shi, C.; Pei, M.; Wang, Y.; Chen, Q.; Cao, P.; Zhang, L.; Guo, J.; Deng, W.; Wang, L.; Li, X.; et al. Changes of Flavin-containing Monooxygenases and Trimethylamine-N-oxide May be Involved in The Promotion of Non-alcoholic Fatty Liver Disease by Intestinal Microbiota Metabolite Trimethylamine. Biochem. Biophys. Res. Commun. 2022, 594, 1–7. [Google Scholar] [CrossRef]
- Luo, Z.; Yu, X.; Wang, C.; Zhao, H.; Wang, X.; Guan, X. Trimethylamine N-oxide Promotes Oxidative Stress and Lipid Accumulation in Macrophage Foam Cells Via the Nrf2/ABCA1 Pathway. J. Physiol. Biochem. 2024, 80, 67–79. [Google Scholar] [CrossRef]
- Li, T.; Chen, Y.; Gua, C.; Li, X. Elevated Circulating Trimethylamine N-Oxide Levels Contribute to Endothelial Dysfunction in Aged Rats through Vascular Inflammation and Oxidative Stress. Front. Physiol. 2017, 8, 350. [Google Scholar] [CrossRef]
- Brunt, V.E.; Gioscia-Ryan, R.A.; Richey, J.J.; Zigler, M.C.; Cuevas, L.M.; Gonzalez, A.; Vázquez-Baeza, Y.; Battson, M.L.; Smithson, A.T.; Gilley, A.D.; et al. Suppression of the Gut Microbiome Ameliorates Age-related Arterial Dysfunction and Oxidative Stress in Mice. J. Physiol. 2019, 597, 2361–2378. [Google Scholar] [CrossRef]
- Li, L.; Chen, B.; Zhu, R.; Li, R.; Tian, Y.; Liu, C.; Jia, Q.; Wang, L.; Tang, J.; Zhao, D.; et al. Fructus Ligustri Lucidi Preserves Bone Quality through the Regulation of Gut Microbiota Diversity, Oxidative Stress, TMAO and Sirt6 Levels in Aging Mice. Aging 2019, 11, 9348–9368. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, S.; Zhang, H.; Gu, Y.; Chen, H.; Huang, Z.; Yang, F.; Li, W.; Chen, C.; Men, L.; et al. The Triangular Relationship Between Traditional Chinese Medicines, Intestinal Flora, and Colorectal Cancer. Med. Res. Rev. 2024, 44, 539–567. [Google Scholar] [CrossRef]
- Zhu, T.; Wang, L.; Feng, Y.; Sun, G.; Sun, X. Classical Active Ingredients and Extracts of Chinese Herbal Medicines: Pharmacokinetics, Pharmacodynamics, and Molecular Mechanisms for Ischemic Stroke. Oxid. Med. Cell Longev. 2021, 2021, 8868941. [Google Scholar] [CrossRef] [PubMed]
- Omar, N.N.; Mosbah, R.A.; Sarawi, W.S.; Rashed, M.M.; Badr, A.M. Rifaximin Protects against Malathion-Induced Rat Testicular Toxicity: A Possible Clue on Modulating Gut Microbiome and Inhibition of Oxidative Stress by Mitophagy. Molecules 2022, 27, 4069. [Google Scholar] [CrossRef] [PubMed]
- Magliocca, G.; Mone, P.; Di Iorio, B.R.; Heidland, A.; Marzocco, S. Short-Chain Fatty Acids in Chronic Kidney Disease: Focus on Inflammation and Oxidative Stress Regulation. Int. J. Mol. Sci. 2022, 23, 5354. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Barcenilla, A.; Stewart, C.S.; Pryde, S.E.; Flint, H.J. Acetate Utilization and Butyryl Coenzyme A (CoA):Acetate-CoA Transferase in Butyrate-Producing Bacteria from the Human Large Intestine. Appl. Environ. Microbiol. 2002, 68, 5186–5190. [Google Scholar] [CrossRef]
- Miyamoto, J.; Hasegawa, S.; Kasubuchi, M.; Ichimura, A.; Nakajima, A.; Kimura, I. Nutritional Signaling via Free Fatty Acid Receptors. Int. J. Mol. Sci. 2016, 17, 450. [Google Scholar] [CrossRef]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, Y.; Jiang, H.; Nie, D. Short-Chain Fatty Acids Induced Autophagy Serves as an Adaptive Strategy for Retarding Mitochondria-Mediated Apoptotic Cell Death. Cell Death Differ. 2011, 18, 602–618. [Google Scholar] [CrossRef]
- Semenova, N.V.; Madaeva, I.M.; Kolesnikova, L.I. The Role of Melatonin as a Component of the Antioxidant Defense System in Perimenopausal Women with Insomnia. Z. Nevrol. Psikhiatr. Im. SS Korsakova 2019, 119, 7. [Google Scholar] [CrossRef]
- Verma, A.K.; Khan, M.I.; Ashfaq, F.; Rizvi, S.I. Crosstalk Between Aging, Circadian Rhythm, and Melatonin. Rejuvenation Res. 2023, 26, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, F.; Sutradhar, S.; Das, P.; Mukherjee, S. Gut Melatonin: A Potent Candidate In the Diversified Journey of Melatonin Research. Gen. Comp. Endocrinol. 2021, 303, 113693. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef]
- Bubenik, G.A. Localization, Physiological Significance and Possible Clinical Implication of Gastrointestinal Melatonin. Neurosignals 2001, 10, 350–366. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Q. Distribution, Function and Physiological Role of Melatonin in the Lower Gut. World J. Gastroenterol. 2011, 17, 3888. [Google Scholar] [CrossRef]
- Iesanu, M.I.; Zahiu, C.D.M.; Dogaru, I.-A.; Chitimus, D.M.; Pircalabioru, G.G.; Voiculescu, S.E.; Isac, S.; Galos, F.; Pavel, B.; O’Mahony, S.M.; et al. Melatonin–Microbiome Two-Sided Interaction in Dysbiosis-Associated Conditions. Antioxidants 2022, 11, 2244. [Google Scholar] [CrossRef]
- Rode, J.; Yang, L.; König, J.; Hutchinson, A.N.; Wall, R.; Venizelos, N.; Brummer, R.-J.; Rangel, I.; Vumma, R. Butyrate Rescues Oxidative Stress-Induced Transport Deficits of Tryptophan: Potential Implication in Affective or Gut-Brain Axis Disorders. Neuropsychobiology 2021, 80, 253–263. [Google Scholar] [CrossRef]
- Wang, T.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin Prevents the Dysbiosis of Intestinal Microbiota in Sleep-Restricted Mice by Improving Oxidative Stress and Inhibiting Inflammation. Saudi J. Gastroenterol. 2022, 28, 209. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Yu, S.; Fang, A.; Shen, X.; Li, K. Association Between Protein Intake and the Risk of Hypertension among Chinese Men and Women: A Longitudinal Study. Nutrients 2022, 14, 1276. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Pérez, D.; Bressa, C.; Bailén, M.; Hamed-Bousdar, S.; Naclerio, F.; Carmona, M.; Pérez, M.; González-Soltero, R.; Montalvo-Lominchar, M.; Carabaña, C.; et al. Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study. Nutrients 2018, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Liu, Y.; Zhen, J.; Mou, P.; Li, J.; Xu, Z.; Song, B. Protective Effect of Methionine on the Intestinal Oxidative Stress and Microbiota Change Induced by Nickel. Ecotoxicol. Environ. Saf. 2022, 244, 114037. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.-H.; Zheng, W.-X.; Zhao, Z.-T.; Meng, N.; Zhang, Q.-R.; Zhu, W.-J.; Li, R.-D.; Liang, X.-L.; Li, Q.-Y. A Pilot Study of the Effect of Lactobacillus Casei Obtained from Long-Lived Elderly on Blood Biochemical, Oxidative, and Inflammatory Markers, and on Gut Microbiota in Young Volunteers. Nutrients 2021, 13, 3891. [Google Scholar] [CrossRef]
- Chen, J.; Ding, X.; Wu, R.; Tong, B.; Zhao, L.; Lv, H.; Meng, X.; Liu, Y.; Ren, B.; Li, J.; et al. Novel Sesquiterpene Glycoside from Loquat Leaf Alleviates Type 2 Diabetes Mellitus Combined with Nonalcoholic Fatty Liver Disease by Improving Insulin Resistance, Oxidative Stress, Inflammation, and Gut Microbiota Composition. J. Agric. Food Chem. 2021, 69, 14176–14191. [Google Scholar] [CrossRef]
- Zou, Y.-F.; JiZe, X.-P.; Li, C.-Y.; Zhang, C.-W.; Fu, Y.-P.; Yin, Z.-Q.; Li, Y.-P.; Song, X.; Li, L.-X.; Zhao, X.-H.; et al. Polysaccharide from Aerial Part of Chuanminshen Violaceum Alleviates Oxidative Stress and Inflammatory Response in Aging Mice through Modulating Intestinal Microbiota. Front. Immunol. 2023, 14, 1159291. [Google Scholar] [CrossRef]
- Gao, Y.; Yu, T.; Wu, Y.; Huang, X.; Teng, J.; Zhao, N.; Zheng, X.; Yan, F. Bacillus Coagulans (Weizmannia Coagulans) XY2 Attenuates Cu-Induced Oxidative Stress via DAF-16/FoxO and SKN-1/Nrf2 Pathways and Gut Microbiota Regulation. J. Hazard. Mater. 2023, 457, 131741. [Google Scholar] [CrossRef]
- Zhu, Y.-Y.; Thakur, K.; Feng, J.-Y.; Zhang, J.-G.; Hu, F.; Cespedes-Acuña, C.L.; Liao, C.; Wei, Z.-J. Riboflavin Bioenriched Soymilk Alleviates Oxidative Stress Mediated Liver Injury, Intestinal Inflammation, and Gut Microbiota Modification in B 2 Depletion–Repletion Mice. J. Agric. Food Chem. 2022, 70, 3818–3831. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Yang, C.; Deng, Q.; Xu, J.; Wang, X.; Hu, C.; Tang, H.; Huang, F. Sinapic Acid and Resveratrol Alleviate Oxidative Stress with Modulation of Gut Microbiota in High-Fat Diet-Fed Rats. Food Res. Int. 2019, 116, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Merecz-Sadowska, A.; Sitarek, P.; Kowalczyk, T.; Zajdel, K.; Jęcek, M.; Nowak, P.; Zajdel, R. Food Anthocyanins: Malvidin and Its Glycosides as Promising Antioxidant and Anti-Inflammatory Agents with Potential Health Benefits. Nutrients 2023, 15, 3016. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Wang, Y.; Wu, Y.; Zhang, M.; Li, F.; He, Y.; Wen, L.; Yue, H. Effect of Stabilization Malvids Anthocyanins on the Gut Microbiota in Mice with Oxidative Stress. J. Food Biochem. 2021, 45, 4892–4902. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| SCFAs | OS | Microbiome | Experiment | References |
|---|---|---|---|---|
| The levels of acetic and butyric acid were significantly lower, while propionic acid was undetected in the malathion group/control group. The levels of serum acetic and propionic acid significantly increased in the rifaximin group/malathion group. There were no significant differences in the butyric acid levels in the malathion group/malathion + rifaximin group. | The activities of SOD and CAT were lower, and MDA levels were higher in the malathion-treated group/control group. The activities of SOD and CAT significantly increased after rifaximin administration for two and four weeks in comparison with the malathion group, whereas the levels of MDA decreased after four weeks of rifaximin administration. | The main phyla in the gut microbiota were Bacteroidetes, Firmicutes, and Proteobacteria. The relative abundance of Firmicutes increased, and Bacteroidetes and Proteobacteria decreased in the malathion group/control group. Rifaximin treatment attenuated malathion-induced changes. The ratio of Firmicutes to Bacteroidetes increased in the malathion group/control group. It was mitigated by rifaximin treatment. | Disease: Testicular dysfunction Subjects: Rats Agent: Malathionan (an organophosphorus pesticide) Treatment: Rifaximin is a poorly absorbed antibiotic | [77] |
| The level was higher in the group without disease but not in the chitosan oligosaccharide group/control group. The level was higher in the group with disease and treated/group with disease and not treated. | Treatment decreased OS, showing a reduction in MDA levels and an increase in SOD levels. | The disease changes the Firmicutes/Bacteroidetes ratio. The treatment reversed it. The abundance of the probiotic family Muribaculaceae and genus Akkermansia rose after treatment. In the disease group, proinflammatory Desulfovibrio and Dubosiella increased. In the cohort with treated disease, it was lower. The same tendency was shown in Escherichia–Shigella and Enterococcus. | Disease: Acute pancreatitis Subjects: Mice Agent: Caerulein Treatment: Chitosan oligosaccharide administration | [32] |
| There was a significant decrease in butyric acid in the disease group/control group. The levels of acetic, propionic, butyric, isobutyric, valeric, and isovaleric acids were significantly improved in the treated group/disease group. | AOC and antioxidative enzyme activities, including GSH-Px, SOD, and CAT, significantly decreased, and the level of MDA and ROS were significantly higher in the disease group/control group. The SOD and CAT activities increased, and MDA and ROS levels decreased in the treated group/disease group. | The relative abundance of Bacteroidetes decreased in the treated group/disease group. The Firmicutes/Bacteroidetes ratio was significantly higher in the treated group/control and disease groups. Bacteroides, Turicibacter, Actinobacteria, Acholeplasmatales, Eubacterium, Staphylococcaceae, Enterobacterales, and Rhodespirillales predominated in the disease group. Dubosiella, Erysipelotrichales, Desulfovibrionia, Peptostreptococcaceae, and Patescibacteria predominated in the treated group. | Disease: IBD Subjects: Mice Agent: Dextran sulfate sodium Treatment: Lonicera hypoglauca and Scutellaria baicalensis as the beneficial bioactivities of extracts from L. hypoglauca and S. baicalensis | [33] |
| Melatonin | OS | Microbiome | Experiment | References |
|---|---|---|---|---|
| Per oral administration for three weeks. | The activity of SOD, CAT, and GSH-Px was higher in the control group/others and was lower in the condition group/treated group. | The ratio of Firmicutes to Bacteroidetes increased in the disease group/control group. Melatonin administration mitigated the changes. | Condition: Liver inflammation Subjects: Mice Agent: Ochratoxin A (the mycotoxin generated by Aspergillus spp.) Treatment: Melatonin | [34] |
| Melatonin supplementation. | There was a decrease in the CAT, GSH-Px, SOD, and AOC levels and an increase in the MDA level in the corticosterone group/control group. Sleep deprivation increased ROS production. ROS production in other groups showed no significant differences. | The ratio of Firmicutes/Bacteroidetes increased in the corticosterone group/control group. The relative abundances of Firmicutes and Proteobacteria increased, and the abundance of Bacteroidetes decreased in the corticosterone group/control group. After melatonin administration, it was mitigated. There was no statistically significant difference between the control group and the melatonin-supplemented group. The relative abundances of Firmicutes and Proteobacterium increased, and Bacteroidetes and Prevotellaceae decreased in the sleep deprivation group/control group. Melatonin pretreatment attenuated the effects of sleep deprivation on the intestinal microbiota disorder. | Condition: IBD Subjects: Mice Agent: Sleep deprivation or corticosterone supplementation Treatment: Melatonin | [35] |
| Supplementation with 10−5 mol/L of melatonin. The plasma melatonin level was decreased in the sleep restriction group/control group. No significant difference in melatonin was observed between the treated group and the control group mice. | The activities of antioxidant enzymes (GSH-Px, CAT, and SOD), AOC, and MDA content in the jejunum: The activity of enzymes and AOC were lower in the sleep restriction group/control group. The level of MDA was higher in the sleep restriction group/control group. No significant differences in AOC, MDA, GSH-Px, and CAT were observed between the treated group and the control group mice. SOD level was increased in the treated group/control group. | The relative abundances of Bacteroidetes and Lactobacillus were decreased, whereas that of Firmicutes, Helicobacter, and Clostridium and the difference in abundance between Firmicutes and Bacteroidetes were increased in the sleep restriction group/control group. Melatonin supplementation mitigated these patterns. | Condition: Sleep restriction Subjects: Mice Treatment: Melatonin | [91] |
| Compound | OS | Microbiome | Subjects | References |
|---|---|---|---|---|
| Proteins and amino acids | Proteins: No significant difference in MDA level between the two groups was detected. Methionine: Heavy metal exposure increased the intestinal MDA and decreased the antioxidant enzyme activity of GSH-Px, glutathione reductase, SOD, and CAT. MDA decreased in methionine-treated groups. Methionine significantly increased the content of antioxidative enzymes. | Proteins: The Bacteroidetes increased, and the Firmicutes decreased in the main group/control group. The presence of health-related taxa, including Roseburia, Blautia, and Bifidobacterium longum, was lower. Methionine: The abundances of Proteobacteria, Actinobacteria, Patescibacteria, and Cyanobacteria were higher in the methionine-treated group/control group. | Human Mice | [93] [94] |
| Probiotics (lactic acid bacteria) | Pediococcus pentosaceus increased the levels of SOD and GSH-Px. Lactobacillus casei LTL1879 increased the AOC level and decreased the MDA level. | The relative abundances of Firmicutes and Proteobacteria were increased in the disease group, which was reversed in the treated group. The decline in Bacteroidetes was restored by treatment. Supplement reversed the decrease in Muribaculaceae, Lachnospiraceae, and Defluviitaleaceae as well as the increase in Erysipelotrichaceae, Enterococcaceae, Dubosiella, and Enterococcus in the disease group. Lactobacillus casei LTL1879 reduced Escherichia coli, Enterococcus, and Bacteroides expression and increased Clostridium leptum, Bifidobacterium, and Lactobacillus expression. | Mice Human | [63] [95] |
| Other supplements | Sesquiterpene glycoside 3 was able to prevent oxidative stress. Taxifolin significantly mitigated oxidative stress injury by alleviating the levels of ROS and MDA as well as increasing antioxidant enzyme activity. Bacillus coagulans XY2 caused SOD, CAT, GSH-Px, and glutathione reductase to increase after Cu exposure. | Sesquiterpene glycoside 3 administration could decrease the ratio of Firmicutes/Bacteroidetes and increase the relative abundances of Lachnospiraceae, Muribaculaceae, and Lactobacillacea. Taxifolin treatment decelerated the D-galactose-induced aging process by regulating the composition of the intestinal flora and abundance of beneficial bacteria, including Enterorhabdus, Clostridium, Bifidobacterium, and Parvibacter. Bacillus coagulans XY2 reversed Cu and caused an increased level of Enterorhabdus abundance and decreased abundances of Intestinimonas, Faecalibaculu, Ruminococcaceae, and Coriobacteriaceae_UCG-002. | Mice | [64,96,97,98] |
| Riboflavin | Supplement elevated CAT, SOD, AOC, and decreased MDA level in the serum. | The relative abundances of Prevotella and Absiella were higher, and the relative abundances of Proteobacteria, Fusobacteria, Synergistetes, and Cyanobacteria were lower in the supplemented group. | Mice | [99] |
| Polyphenols | MDA level in colon tissues was decreased, and the levels of SOD, GSH, and GSH-Px were increased by diosmetin. Sinapic acid consumption decreased ROS and MDA levels in the colon and increased AOC in the liver. Malvidin anthocyanins increased the activities of SOD, GSH-Px, CAT, and AOC but decreased the levels of MDA in the serum and liver. | The diosmetin-treated group had increased relative abundances of Bacteroidetes and Cyanobacteria and a decreased relative abundance of Firmicutes compared with the disease group. The abundances of Eggerthella, Flavobacterium, and Clostridium were lower, and the abundances of Odoribacteraceae, Prevotella, Rikenellaceae, Ruminococcus, Coprococcus, Roseburia, Oscillospira, Anaeroplasma, and Synergistales were higher in the diosmetin-treated group/disease group. Polyphenol supplementation (resveratrol and sinapic acid) increased the proportion of butyrate produced in the Lachaospiraceae family and inhibited the growth of bacterial species associated with diseases and inflammation such as Bacteroides and Desulfovibrio spp. Malvidin anthocyanins increased the relative abundance of Firmicutes and decreased the relative abundance of Bacteroidetes. | Mice Rats | [36,103] [101] |
| Agent | Possible Mechanism | References |
|---|---|---|
| TMAO | An imbalance in the microbiome composition; precisely, Bifidobacterium abundance declines and Firmicutes/Bacteroidetes ratio changes, which causes a rise in TMAO level, which, in turn, leads to OS. | [74] |
| SCFAs | Several SCFAs might modulate mitochondrial function, including oxidative processes. The gut microbiome may influence autophagy processes by producing SCFAs. The damaged mitochondria generate excess ROS, and their autophagy leads to decreased OS in cells. | [1,77] |
| Melatonin | Melatonin alters colonic microbial composition and diversity, resulting in decreased Gram-negative bacterial-derived lipopolysaccharides that could enter into the blood and liver via defects in the intestinal barrier. These effects lead to the alleviation of ochratoxin A-induced liver inflammation, oxidative stress, and mitophagy. | [34] |
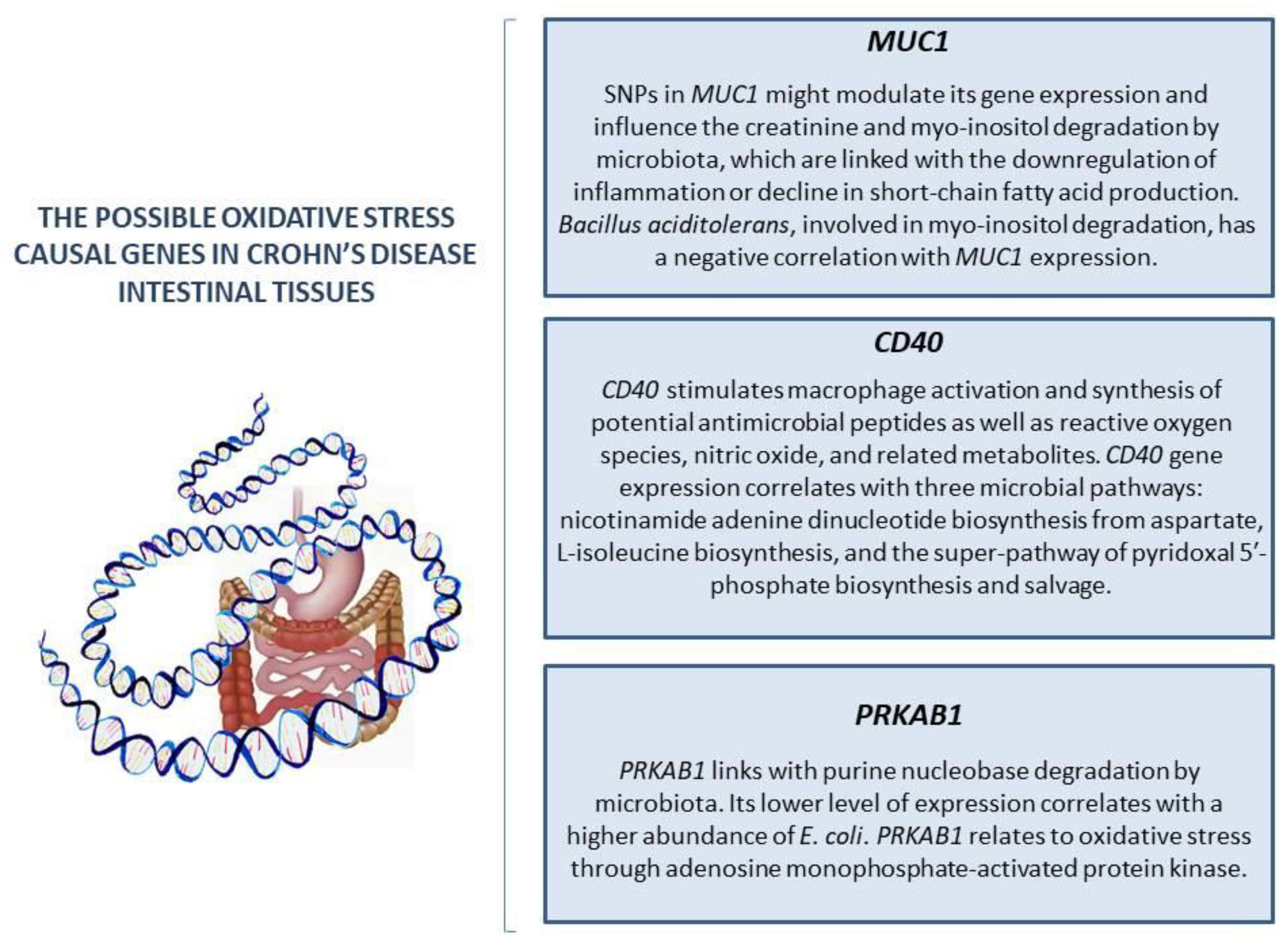
| Genes | It was suggested that high MUC1 (OS causal gene) expression accompanied by decreased production of microbiota-derived metabolites could confer an increased risk of CD. CD40 and its ligand (CD40L) are associated with ROS production. It was suggested that genetic variants regulate CD40 expression and interact with inflammation-related microbial activities and, therefore, contribute to CD pathogenesis through three microbial pathways associated with CD40 gene expression: nicotinamide adenine dinucleotide biosynthesis from aspartate, L-isoleucine biosynthesis, and the superpathway of pyridoxal 5-phosphate biosynthesis and salvage. PRKAB1 expression may be negatively associated with the risk of developing CD because of the interaction between gene expression and intestinal microbial nucleotide metabolism. | [45] |
| Supplements | No mechanisms were presented in this study. The influence of supplements could be parallel or co-dependent. | [19,36,63,64,74,97,98,101,103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenova, N.; Garashchenko, N.; Kolesnikov, S.; Darenskaya, M.; Kolesnikova, L. Gut Microbiome Interactions with Oxidative Stress: Mechanisms and Consequences for Health. Pathophysiology 2024, 31, 309-330. https://doi.org/10.3390/pathophysiology31030023
Semenova N, Garashchenko N, Kolesnikov S, Darenskaya M, Kolesnikova L. Gut Microbiome Interactions with Oxidative Stress: Mechanisms and Consequences for Health. Pathophysiology. 2024; 31(3):309-330. https://doi.org/10.3390/pathophysiology31030023
Chicago/Turabian StyleSemenova, Natalya, Nadezhda Garashchenko, Sergey Kolesnikov, Marina Darenskaya, and Liubov Kolesnikova. 2024. "Gut Microbiome Interactions with Oxidative Stress: Mechanisms and Consequences for Health" Pathophysiology 31, no. 3: 309-330. https://doi.org/10.3390/pathophysiology31030023
APA StyleSemenova, N., Garashchenko, N., Kolesnikov, S., Darenskaya, M., & Kolesnikova, L. (2024). Gut Microbiome Interactions with Oxidative Stress: Mechanisms and Consequences for Health. Pathophysiology, 31(3), 309-330. https://doi.org/10.3390/pathophysiology31030023