
Rab11a Controls Cell Shape via C9orf72 Protein: Possible Relationships to Frontotemporal Dementia/Amyotrophic Lateral Sclerosis (FTDALS) Type 1

 , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Antibodies and Chemicals
2.2. Synthetic siRNAs and DNA Primers
2.3. Reverse Transcription-Polymerase Chain (RT-PCR) Reaction
2.4. Cell Culture and Differentiation
2.5. Isolation and Culture of Primary Cortical Neuronal Cells
2.6. siRNA Transfection
2.7. Polyacrylamide Gel Electrophoresis and Immunoblotting
2.8. Statistical Analyses
2.9. Ethics Statement
3. Results
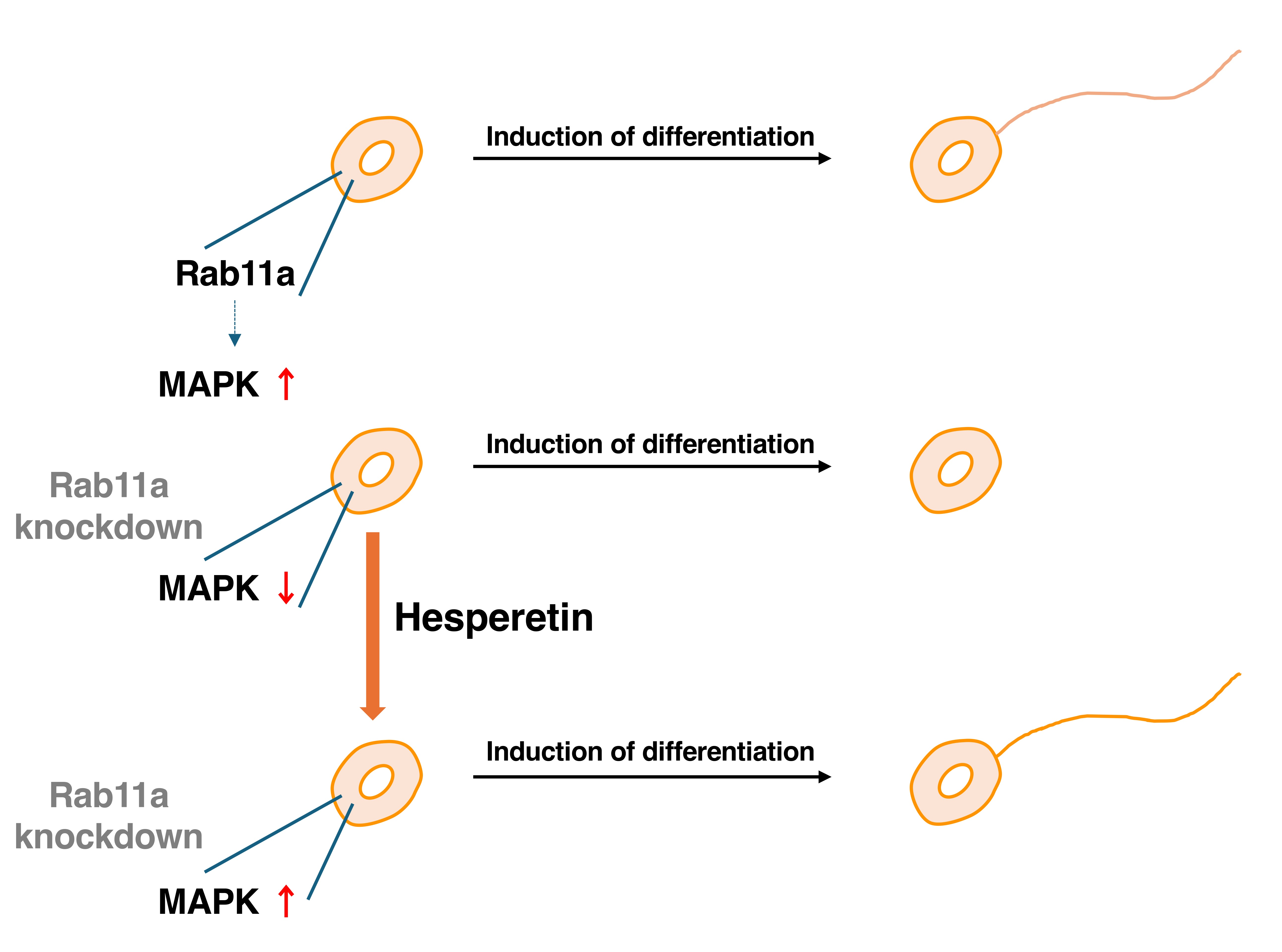
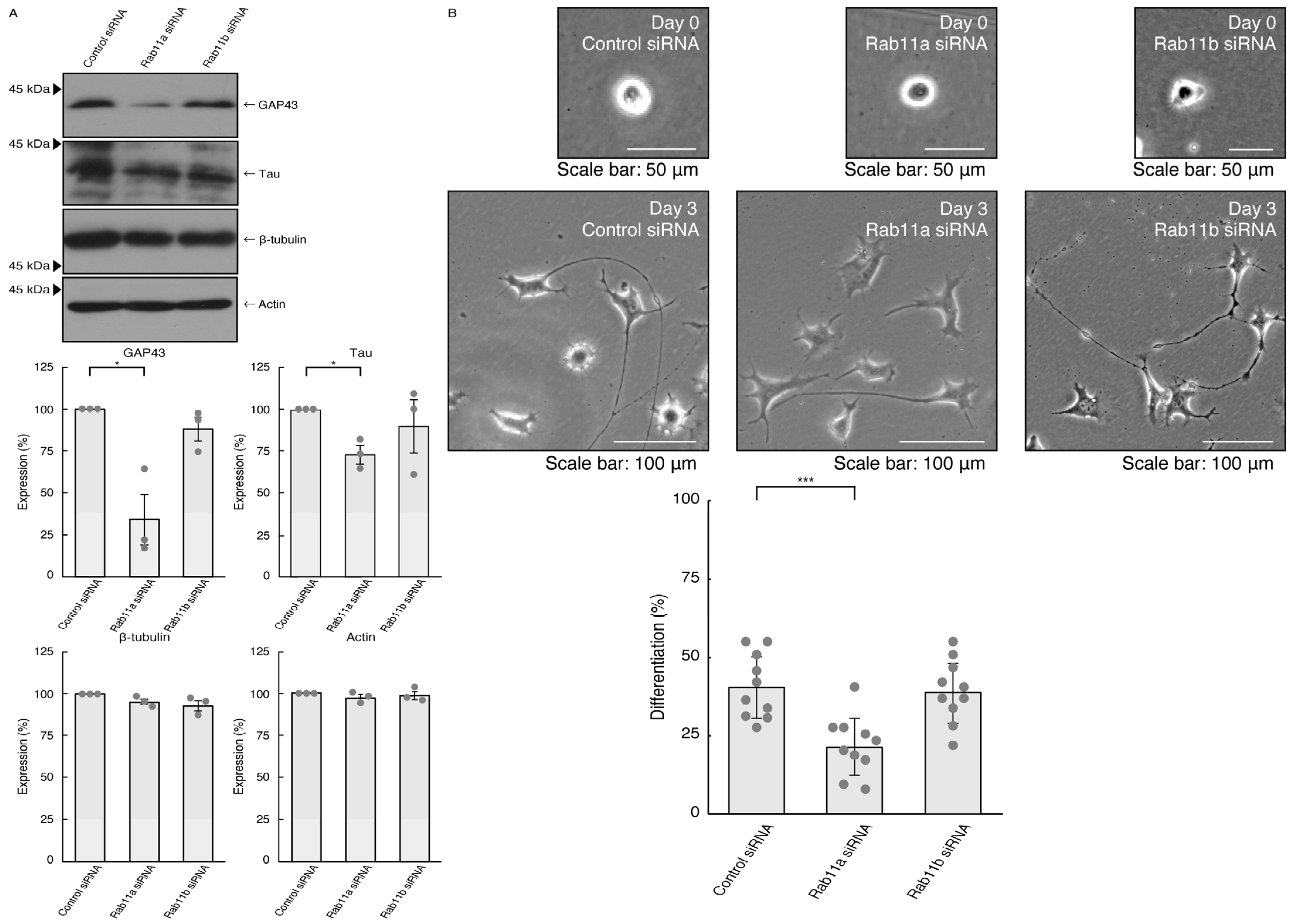
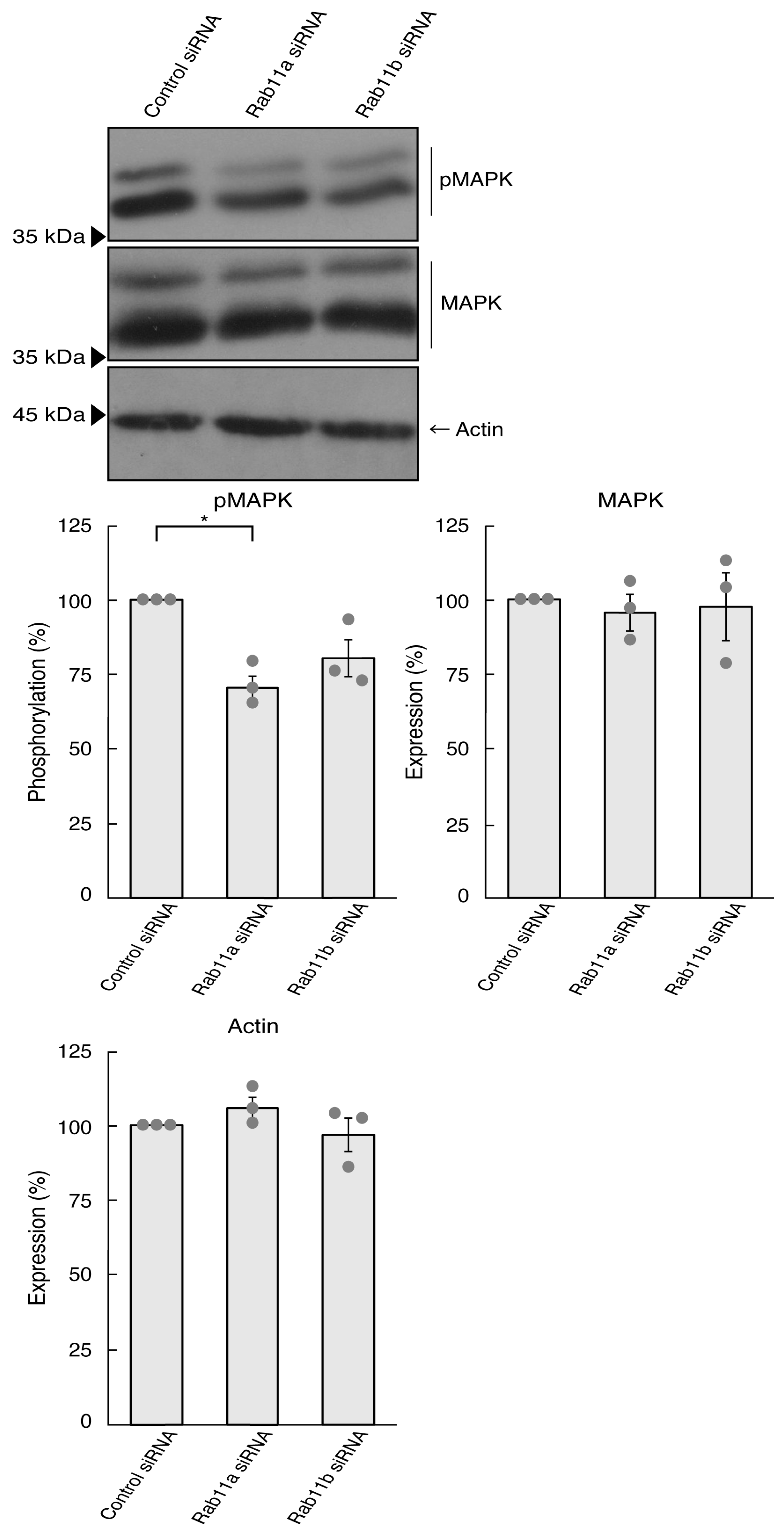
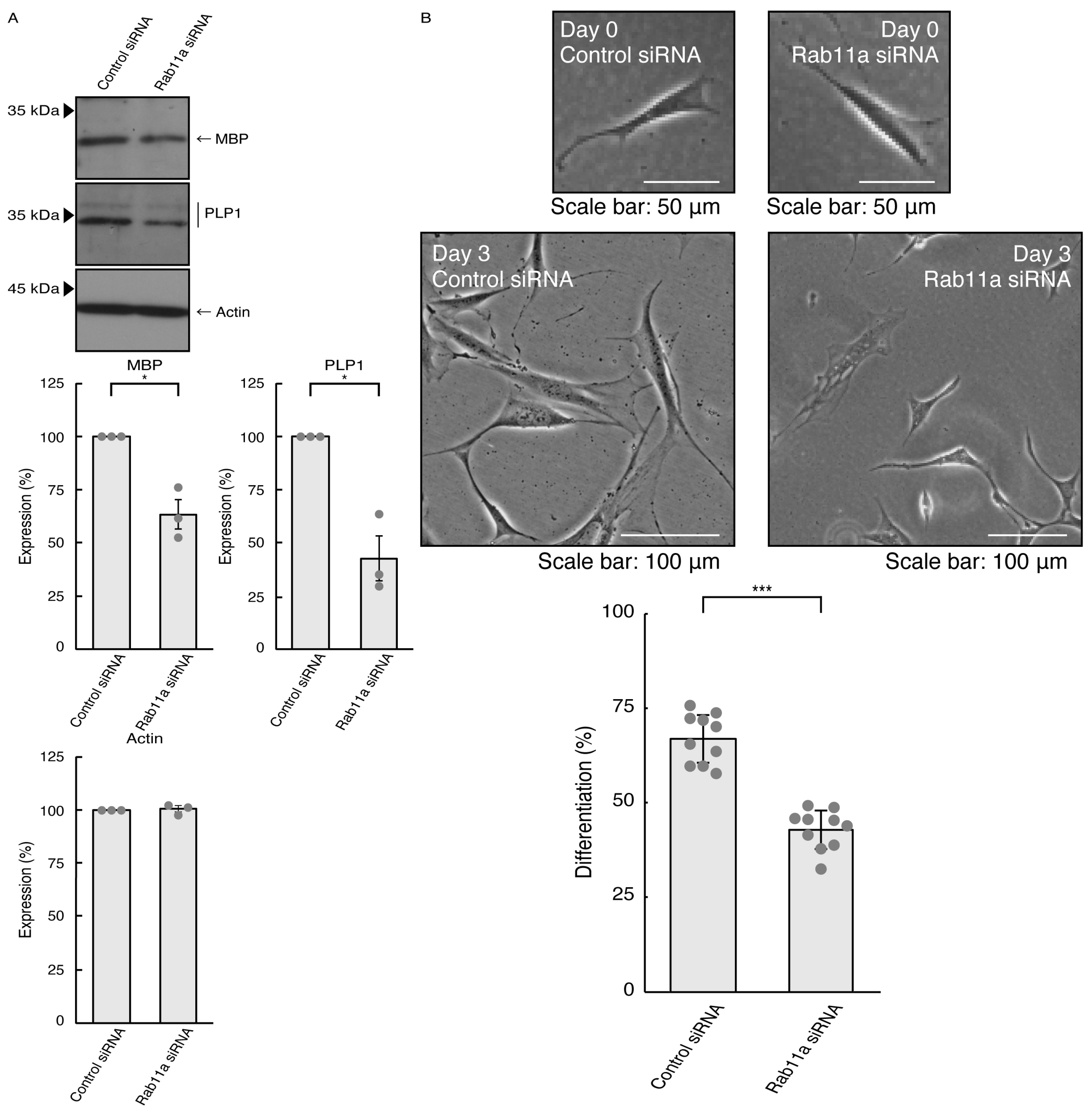
3.1. Rab11a Positively Regulates Neuronal Cell Morphological Differentiation
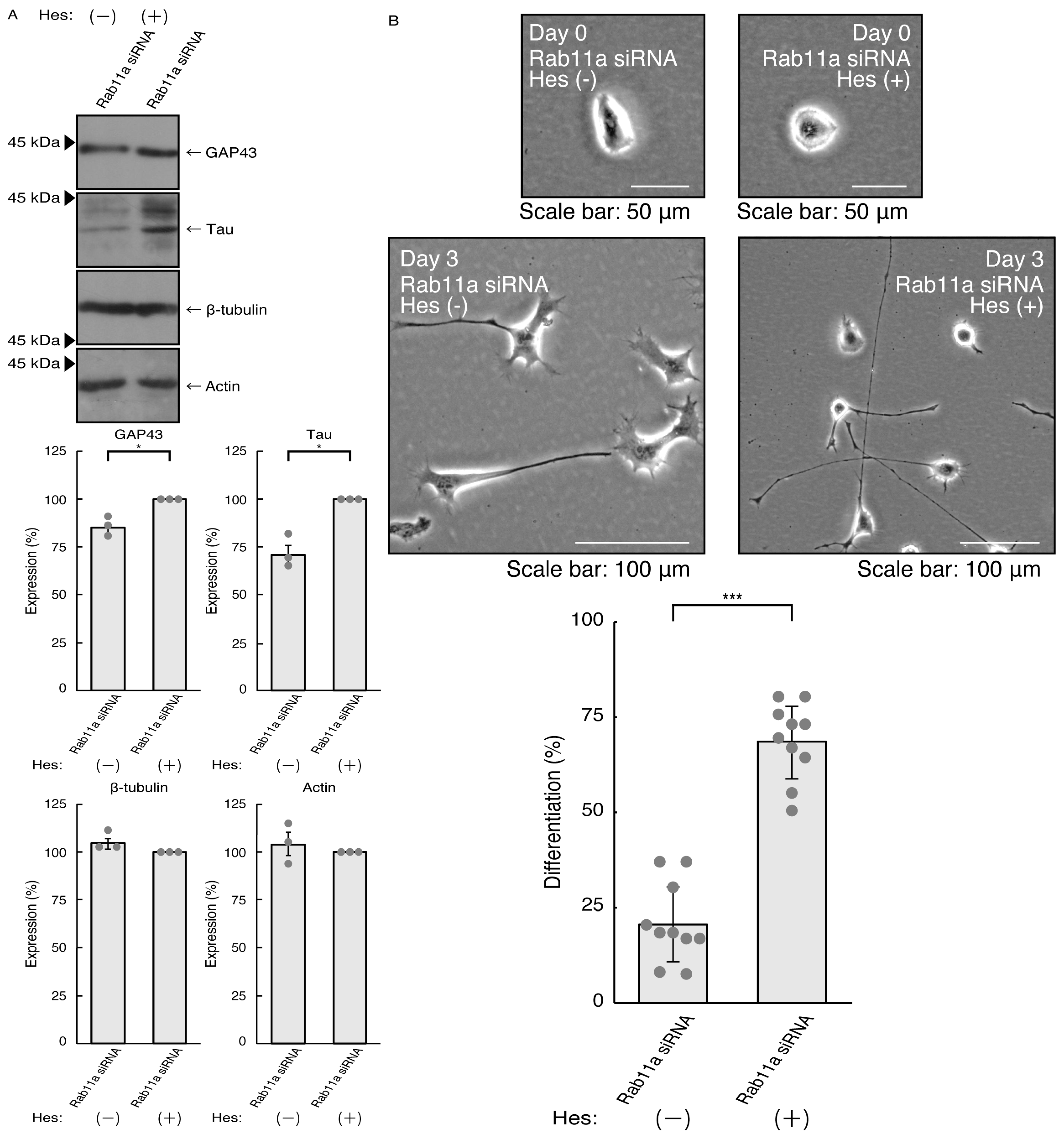
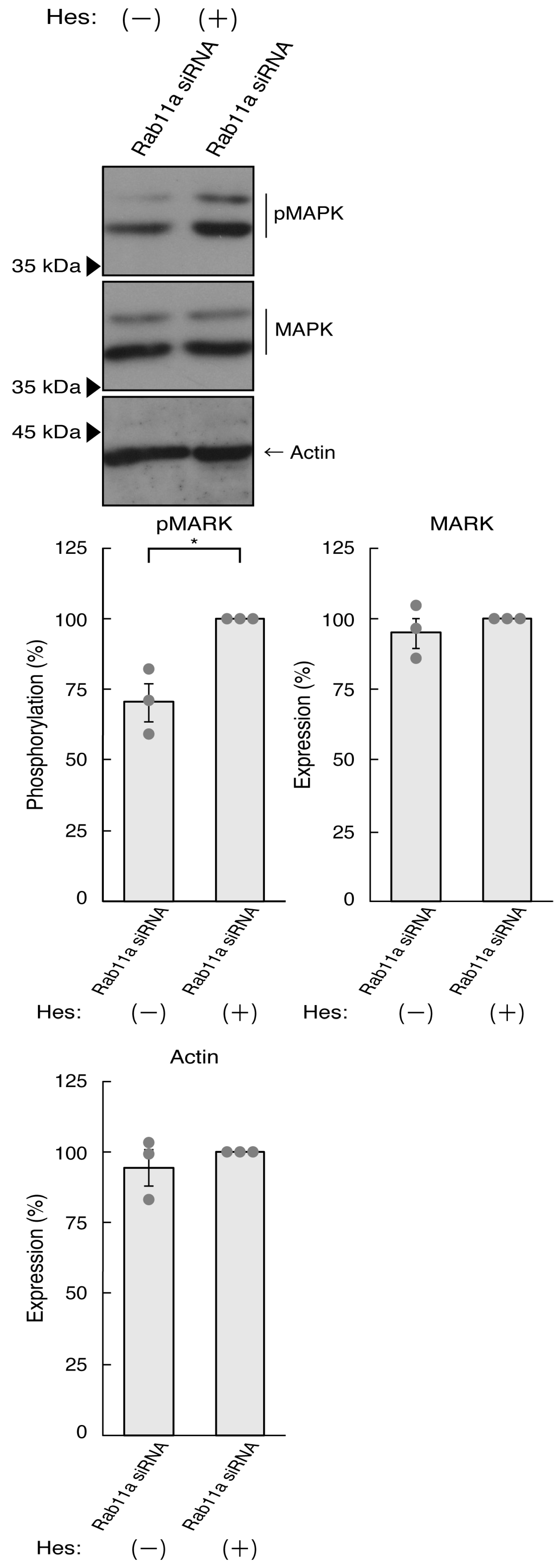
3.2. Hesperetin Recovers Cellular Phenotypes Induced by Rab11a Knockdown
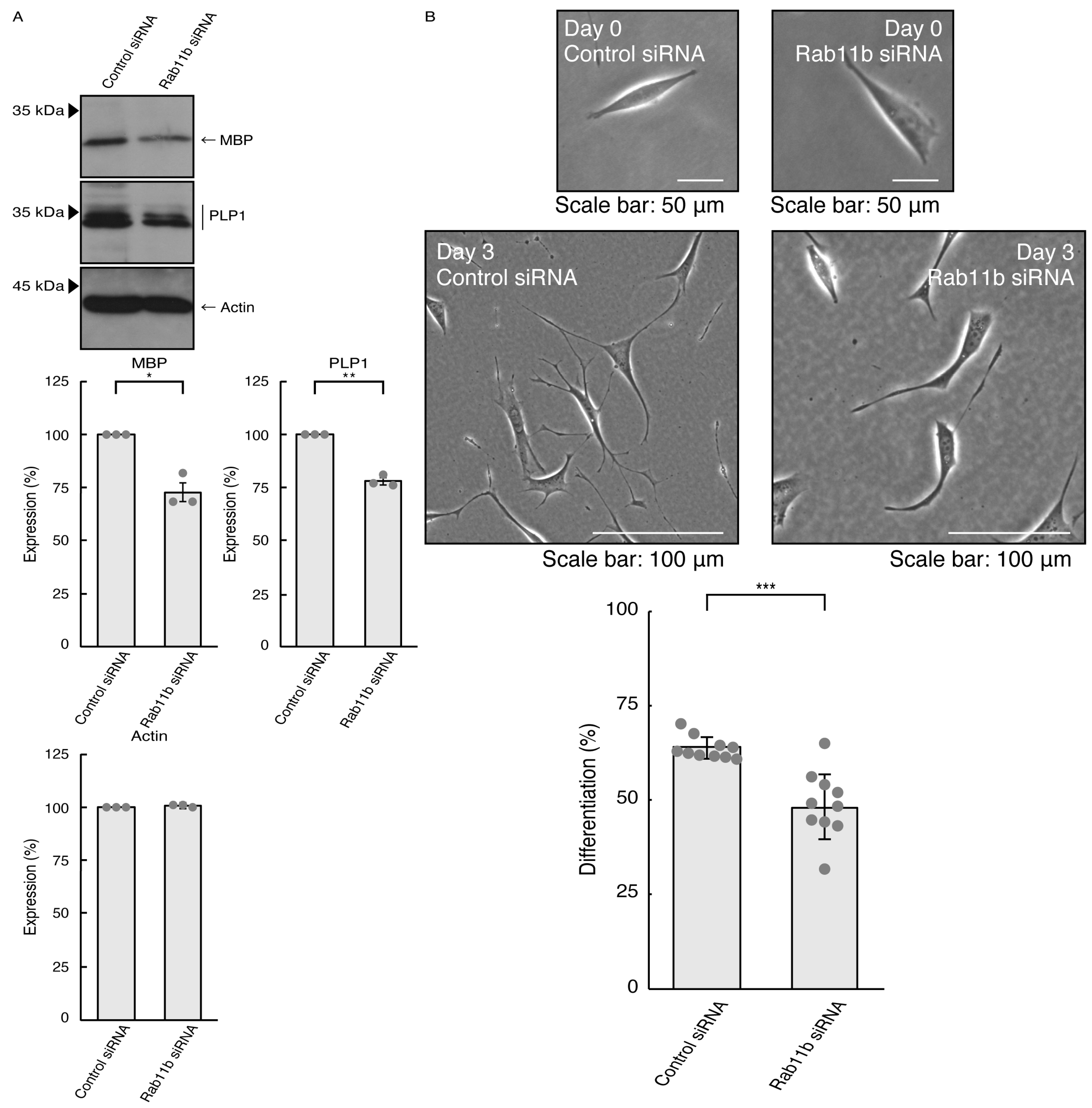
3.3. Rab11a and Rab11b Also Positively Regulate Oligodendroglial Cell Morphological Differentiation
3.4. Knockdown of Rab14 Does Not Significantly Affect Cell Morphogenesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, D. Surface movements during the growth of single explanted neurons. Proc. Natl. Acad. Sci. USA 1970, 65, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Bray, D. Actin and myosin in neurons: A first review. Biochimie 1977, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.M.; Banker, G. Neuronal polarity. Annu. Rev. Neurosci. 1994, 17, 267–310. [Google Scholar] [CrossRef]
- da Silva, J.S.; Dotti, C.G. Breaking the neuronal sphere: Regulation of the actin cytoskeleton in neuritogenesis. Nat. Rev. Neurosci. 2002, 3, 694–704. [Google Scholar] [CrossRef]
- Arimura, N.; Kaibuchi, K. Neuronal polarity: From extracellular signals to intracellular mechanisms. Nat. Rev. Neurosci. 2007, 8, 194–205. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Olney, N.T.; Spina, S.; Miller, B.L. Frontotemporal dementia. Neurol. Clin. 2017, 35, 339–374. [Google Scholar] [CrossRef]
- Balendra, R.; Isaacs, A.M. C9orf72-mediated ALS and FTD: Multiple pathways to disease. Nat. Rev. Neurol. 2018, 14, 544–558. [Google Scholar] [CrossRef]
- McCauley, M.E.; Baloh, R.H. Inflammation in ALS/FTD pathogenesis. Acta Neuropathol. 2019, 137, 715–730. [Google Scholar] [CrossRef]
- Smeyers, J.; Banchi, E.G.; Latouche, M. FC9ERF72: What it is, what it does, and why it matters. Front. Cell Neurosci. 2021, 15, 661447. [Google Scholar] [CrossRef]
- Pang, W.; Hu, F. Cellular and physiological functions of C9ORF72 and implications for ALS/FTD. J. Neurochem. 2021, 157, 334–350. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.A.; Lally, C.; Kupelian, V.; Flanders, W.D. Estimated prevalence and incidence of amyotrophic lateral sclerosis and SOD1 and C9orf72 genetic variants. Neuroepidemiology 2021, 5, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Greaves, C.V.; Rohrer, J.D. An update on genetic frontotemporal dementia. J. Neurol. 2019, 266, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Root, J.; Merino, P.; Nuckols, A.; Johnson, M.; Kukar, T. Lysosome dysfunction as a cause of neurodegenerative diseases: Lessons from frontotemporal dementia and amyotrophic lateral sclerosis. Neurobiol. Dis. 2021, 154, 105360. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Hipp, M.S.; Kasturi, P.; Hartl, F.U. The proteostasis network and its decline in ageing. Nat. Rev. Mol. Cell Biol. 2019, 20, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.R. Rab GTPase regulation of membrane identity. Curr. Opin. Cell Biol. 2013, 25, 414–419. [Google Scholar] [CrossRef]
- Homma, Y.; Hiragi, S.; Fukuda, M. Rab family of small GTPases: An updated view on their regulation and functions. FEBS J. 2021, 288, 36–55. [Google Scholar] [CrossRef]
- Raghavan, S.; Brishti, M.A.; Leo, M.D. Rab GTPases as modulators of vascular function. Cells 2022, 11, 3061. [Google Scholar] [CrossRef]
- Hirose, M.; Ishizaki, T.; Watanabe, N.; Uehata, M.; Kranenburg, O.; Moolenaar, W.H.; Matsumura, F.; Maekawa, M.; Bito, H.; Narumiya, S. Molecular dissection of the Rho-associated protein kinase (p160ROCK)-regulated neurite remodeling in neuroblastoma N1E-115 cells. J. Cell Biol. 1998, 141, 1625–1636. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Torii, T.; Yamamori, N.; Ogata, T.; Tanoue, A.; Yamauchi, J. Akt and PP2A reciprocally regulate the guanine nucleotide exchange factor Dock6 to control axon growth of sensory neurons. Sci. Signal. 2013, 6, ra15. [Google Scholar] [CrossRef] [PubMed]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Neuropharmacological properties and pharmacokinetics of the citrus flavonoids hesperidin and hesperetin—A mini review. Life Sci. 2014, 113, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, H.; Seiwa, C.; Yoshioka, N.; Mizoguchi, K.; Yamamoto, M.; Asou, H.; Aiso, S. An extract of Chinpi, the dried peel of the citrus fruit Unshiu, enhances axonal remyelination via promoting the proliferation of oligodendrocyte progenitor cells. Evid. Based Complement. Alternat. Med. 2016, 2016, 86. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef] [PubMed]
- William Raja, T.R.; Duraipandiyan, V.; Ignacimuthu, S.; Janakiraman, U.; Packiam, S.M. Role of polyphenols in alleviating Alzheimer’s disease: A review Curr. Med. Chem. 2023, 30, 4032–4047. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef]
- Stupack, J.; Xiong, X.P.; Jiang, L.L.; Zhang, T.; Zhou, L.; Campos, A.; Ranscht, B.; Mobley, W.; Pasquale, E.B.; Xu, H.; et al. Soluble SORLA enhances neurite outgrowth and regeneration through activation of the EGF Receptor/ERK signaling axis. J. Neurosci. 2020, 40, 5908–5921. [Google Scholar] [CrossRef]
- Kato, Y.; Shirai, R.; Ohbuchi, K.; Oizumi, H.; Yamamoto, M.; Miyata, W.; Iguchi, T.; Mimaki, Y.; Miyamoto, Y.; Yamauchi, J. Hesperetin ameliorates inhibition of neuronal and oligodendroglial cell differentiation phenotypes induced by knockdown of Rab2b, an autism spectrum disorder-associated gene product. Neurol. Int. 2023, 15, 371–391. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Yamauchi, J.; Chan, J.R.; Okada, A.; Tomooka, Y.; Hisanaga, S.; Tanoue, A. Cdk5 regulates differentiation of oligodendrocyte precursor cells through the direct phosphorylation of paxillin. J. Cell Sci. 2007, 120, 4355–4366. [Google Scholar] [CrossRef]
- Numakawa, T.; Yokomaku, D.; Kiyosue, K.; Adachi, N.; Matsumoto, T.; Numakawa, Y.; Taguchi, T.; Hatanaka, H.; Yamada, M. Basic fibroblast growth factor evokes a rapid glutamate release through activation of the MAPK pathway in cultured cortical neurons. J. Biol. Chem. 2002, 277, 28861–28869. [Google Scholar] [CrossRef]
- Hwang, S.; Lee, S.E.; Ahn, S.G.; Lee, G.H. Psoralidin stimulates expression of immediate-early genes and synapse development in primary cortical neurons. Neurochem. Res. 2018, 43, 2460–2472. [Google Scholar] [CrossRef]
- Zhao, L.; Ji, X.; Zhang, X.; Li, L.; Jin, Y.; Liu, W. FLCN is a novel Rab11A-interacting protein that is involved in the Rab11A-mediated recycling transport. J. Cell Sci. 2018, 131, jcs218792. [Google Scholar] [CrossRef]
- Rinaldi, B.; Villa, R.; Sironi, A.; Garavelli, L.; Finelli, P.; Bedeschi, M.F. Smith-Magenis syndrome-clinical review, biological background and related disorders. Genes 2022, 13, 335. [Google Scholar] [CrossRef]
- Poisson, A.; Nicolas, A.; Bousquet, I.; Raverot, V.; Gronfier, C.; Demily, C. Smith-Magenis Syndrome: Molecular basis of a genetic-driven melatonin circadian secretion disorder. Int. J. Mol. Sci. 2019, 20, 3533. [Google Scholar] [CrossRef]
- Scotter, E.L.; Chen, H.J.; Shaw, C.E. TDP-43 proteinopathy and ALS: Insights into disease mechanisms and therapeutic targets. Neurotherapeutics 2015, 12, 352–363. [Google Scholar] [CrossRef]
- Feneberg, E.; Gray, E.; Ansorge, O.; Talbot, K.; Turner, M.R. Towards a TDP-43-based biomarker for ALS and FTLD. Mol. Neurobiol. 2018, 55, 7789–7801. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Bharathi, V.; Sivalingam, V.; Girdhar, A.; Patel, B.K. Molecular mechanisms of TDP-43 misfolding and pathology in amyotrophic lateral sclerosis. Front. Mol. Neurosci. 2019, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, Y.; Urushitani, M. Molecular Dissection of TDP-43 as a leading cause of ALS/FTLD. Int. J. Mol. Sci. 2022, 23, 12508. [Google Scholar] [CrossRef]
- Xia, Q.; Wang, G.; Wang, H.; Hu, Q.; Ying, Z. Folliculin, a tumor suppressor associated with Birt-Hogg-Dubé (BHD) syndrome, is a novel modifier of TDP-43 cytoplasmic translocation and aggregation. Hum. Mol. Genet. 2016, 25, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.L.; Margaria, J.P.; Stariha, J.T.B.; Hoffmann, R.M.; McPhail, J.A.; Hamelin, D.J.; Boulanger, M.J.; Hirsch, E.; Burke, J.E. Structural determinants of Rab11 activation by the guanine nucleotide exchange factor SH3BP5. Nat. Commun. 2018, 9, 3772. [Google Scholar] [CrossRef]
- Goto-Ito, S.; Morooka, N.; Yamagata, A.; Sato, Y.; Sato, K.; Fukai, S. Structural basis of guanine nucleotide exchange for Rab11 by SH3BP5. Life Sci. Alliance 2019, 2, e201900297. [Google Scholar] [CrossRef]
- Takeshita, Y.; Hashimoto, Y.; Nawa, M.; Uchino, H.; Matsuoka, M. SH3-binding protein 5 mediates the neuroprotective effect of the secreted bioactive peptide humanin by inhibiting c-Jun NH2-terminal kinase. J. Biol. Chem. 2013, 288, 24691–24704. [Google Scholar] [CrossRef]
- Horgan, C.P.; Hanscom, S.R.; McCaffrey, M.W. GRAB is a binding partner for the Rab11a and Rab11b GTPases. Biochem. Biophys. Res. Commun. 2013, 441, 214–219. [Google Scholar] [CrossRef]
- Van Bergen, N.J.; Ahmed, S.M.; Collins, F.; Cowley, M.; Vetro, A.; Dale, R.C.; Hock, D.H.; de Caestecker, C.; Menezes, M.; Massey, S.; et al. Mutations in the exocyst component EXOC2 cause severe defects in human brain development. J. Exp. Med. 2020, 217, e20192040. [Google Scholar] [CrossRef] [PubMed]
- Coulter, M.E.; Musaev, D.; DeGennaro, E.M.; Zhang, X.; Henke, K.; James, K.N.; Smith, R.S.; Hill, R.S.; Partlow, J.N.; Muna, A.S.; et al. Regulation of human cerebral cortical development by EXOC7 and EXOC8, components of the exocyst complex, and roles in neural progenitor cell proliferation and survival. Genet. Med. 2020, 22, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Read, D.E.; Gorman, A.M. Involvement of Akt in neurite outgrowth. Cell Mol. Life Sci. 2009, 66, 2975–2984. [Google Scholar]
- Chacón, M.R.; Fazzari, P. FAK: Dynamic integration of guidance signals at the growth cone. Cell Adh. Migr. 2011, 5, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Fu, L.; Hao, J.; Lin, X.; Dong, Q. Rab11a is overexpressed in gastric cancer and regulates FAK/AKT signaling. J. Oncol. 2020, 2020, 3494396. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.; Balasubramanian, I.; D’Agostino, L.; Bandyopadhyay, S.; Patel, R.; Avasthi, S.; Yu, S.; Goldenring, J.R.; Bonder, E.M.; Gao, N. RAB11A-mediated YAP localization to adherens and tight junctions is essential for colonic epithelial integrity. J. Biol. Chem. 2021, 297, 100848. [Google Scholar] [CrossRef]
- Lorente Pons, A.; Higginbottom, A.; Cooper-Knock, J.; Alrafiah, A.; Alofi, E.; Kirby, J.; Shaw, P.J.; Wood, J.D.; Highley, J.R. Oligodendrocyte pathology exceeds axonal pathology in white matter in human amyotrophic lateral sclerosis. J. Pathol. 2020, 251, 262–271. [Google Scholar] [CrossRef]
- Sirisi, S.; Querol-Vilaseca, M.; Dols-Icardo, O.; Pegueroles, J.; Montal, V.; Muñoz, L.; Torres, S.; Ferrer-Raventós, P.; Iulita, M.F.; Sánchez-Aced, É.; et al. Myelin loss in C9orf72 hexanucleotide expansion carriers. J. Neurosci. Res. 2022, 100, 1862–1875. [Google Scholar] [CrossRef]
- Marian, O.C.; Teo, J.D.; Lee, J.Y.; Song, H.; Kwok, J.B.; Landin-Romero, R.; Halliday, G.; Don, A.S. Disrupted myelin lipid metabolism differentiates frontotemporal dementia caused by GRN and C9orf72 gene mutations. Acta Neuropathol. Commun. 2023, 11, 52. [Google Scholar] [CrossRef]
- De Marchi, F.; Tondo, G.; Corrado, L.; Menegon, F.; Aprile, D.; Anselmi, M.; D’Alfonso, S.; Comi, C.; Mazzini, L. Neuroinflammatory pathways in the ALS-FTD continuum: A focus on genetic variants. Genes 2023, 14, 1658. [Google Scholar] [CrossRef]
- Bryant, D.M.; Datta, A.; Rodríguez-Fraticelli, A.E.; Peränen, J.; Martín-Belmonte, F.; Mostov, K.E. A molecular network for de novo generation of the apical surface and lumen. Nat. Cell Biol. 2010, 12, 1035–1145. [Google Scholar] [CrossRef]
- Fielding, A.B.; Schonteich, E.; Matheson, J.; Wilson, G.; Yu, X.; Hickson, G.R.; Srivastava, S.; Baldwin, S.A.; Prekeris, R.; Gould, G.W. Rab11-FIP3 and FIP4 interact with Arf6 and the exocyst to control membrane traffic in cytokinesis. EMBO J. 2005, 24, 3389–3899. [Google Scholar] [CrossRef]
- Welz, T.; Wellbourne-Wood, J.; Kerkhoff, E. Orchestration of cell surface proteins by Rab 11. Trends Cell Biol. 2014, 24, 407–415. [Google Scholar] [CrossRef]
- Escrevente, C.; Bento-Lopes, L.; Ramalho, J.S.; Barral, D.C. Rab11 is required for lysosome exocytosis through the interaction with Rab3a, Sec15 and GRAB. J. Cell Sci. 2021, 134, jcs246694. [Google Scholar] [CrossRef]
- Niu, M.; Zheng, N.; Wang, Z.; Gao, Y.; Luo, X.; Chen, Z.; Fu, X.; Wang, Y.; Wang, T.; Liu, M.; et al. RAB39B deficiency impairs learning and memory partially through compromising autophagy. Front. Cell Dev. Biol. 2020, 8, 598622. [Google Scholar] [CrossRef]
- Tang, B.L. RAB39B’s role in membrane traffic, autophagy, and associated neuropathology. J. Cell Physiol. 2021, 236, 1579–1592. [Google Scholar] [CrossRef]
- Mignogna, M.L.; Musardo, S.; Ranieri, G.; Gelmini, S.; Espinosa, P.; Marra, P.; Belloli, S.; Murtaj, V.; Moresco, R.M.; Bellone, C.; et al. RAB39B-mediated trafficking of the GluA2-AMPAR subunit controls dendritic spine maturation and intellectual disability-related behavior. Mol. Psychiatry 2021, 26, 6531–6549. [Google Scholar] [CrossRef] [PubMed]
- Mignogna, M.L.; Ficarella, R.; Gelmini, S.; Marzulli, L.; Ponzi, E.; Gabellone, A.; Peschechera, A.; Alessio, M.; Margari, L.; Gentile, M.; et al. Clinical characterization of a novel RAB39B nonstop mutation in a family with ASD and severe ID causing RAB39B downregulation and study of a Rab39b knock down mouse model. Hum. Mol. Genet. 2022, 31, 1389–1406. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.M.; Zhou, X.; Robins, A.M.; Paushter, D.H.; Kim, D.; Smolka, M.B.; Hu, F. The ALS/FTLD associated protein C9orf72 associates with SMCR8 and WDR41 to regulate the autophagy-lysosome pathway. Acta Neuropathol. Commun. 2016, 4, 51. [Google Scholar] [CrossRef]
- Ho, W.Y.; Tai, Y.K.; Chang, J.C.; Liang, J.; Tyan, S.H.; Chen, S.; Guan, J.L.; Zhou, H.; Shen, H.M.; Koo, E.; et al. The ALS-FTD-linked gene product, C9orf72, regulates neuronal morphogenesis via autophagy. Autophagy 2019, 15, 827–842. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents or Sources | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| Antibodies | ||||
| Ant-growth associated protein 43 (GAP43) | Santa Cruz Biotechnology (Santa Cruz, CA, USA) | sc-17790 | J0920 | Immunoblotting (IB), 1:20,000 |
| Anti-tau | Santa Cruz Biotechnology | sc-21796 | G1222 | IB, 1:500 |
| Anti-tubulin (beta type) | Santa Cruz Biotechnology | sc-80005 | A1821 | IB, 1:20,000 |
| Anti-actin (also called beta-type actin) | MBL (Tokyo, Japan) | M177-3 | 007 | IB, 1:5000 |
| Anti-phospho-p44/42 MAPK (pErk1/2) | Cell Signaling Technology (Danvers, MA, USA) | 4370S | 5 | IB, 1:2000 |
| Anti-p44/42 MAPK (Erk1/2) | Cell Signaling Technology | 4695T | 35 | IB, 1:1000 |
| Anti-myelin basic protein (MBP) | BioLegend (San Diego, CA, USA) | 836504 | B225469 | IB, 1:500 |
| Anti-proteolipid protein 1 (PLP1) | Atlas Antibodies (Bromma, Sweden) | HPA004128 | 8115828 | IB, 1:500 |
| Anti-IgG (H+L chain) (mouse) pAb-HRP | MBL (Tokyo, Japan) | 330 | 365 | IB, 1:5000 |
| Anti-IgG (H+L chain) (rabbit) pAb-HRP | MBL | 458 | 353 | IB, 1:5000 |
| Key chemicals | ||||
| Hesperitin (Hes) | FUJIFILM Wako Pure Chemical Corporation (Tokyo, Japan) | 087-10001 | DLK5755 | 15 miclomoler (final concentration) |
| Dimethyl sulfoxide (DMSO) | FUJIFILM Wako Pure Chemical Corporation | 047-29353 | CDN0170 | Less than 0.1% (final concentration) |
| Cell lines | ||||
| FBD-102b cells (mouse cells) | Dr. Yasuhiro Tomo-oka (Tokyo University of Science, Chiba, Japan and Riken, Saitama, Japan) | N/A | N/A | |
| N1E-115 cells (mouse cells) | Dr. Daisuke Shiokawa (Tokyo University of Science, Chiba, Japan) | N/A | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukatsu, S.; Sashi, H.; Shirai, R.; Takagi, N.; Oizumi, H.; Yamamoto, M.; Ohbuchi, K.; Miyamoto, Y.; Yamauchi, J. Rab11a Controls Cell Shape via C9orf72 Protein: Possible Relationships to Frontotemporal Dementia/Amyotrophic Lateral Sclerosis (FTDALS) Type 1. Pathophysiology 2024, 31, 100-116. https://doi.org/10.3390/pathophysiology31010008
Fukatsu S, Sashi H, Shirai R, Takagi N, Oizumi H, Yamamoto M, Ohbuchi K, Miyamoto Y, Yamauchi J. Rab11a Controls Cell Shape via C9orf72 Protein: Possible Relationships to Frontotemporal Dementia/Amyotrophic Lateral Sclerosis (FTDALS) Type 1. Pathophysiology. 2024; 31(1):100-116. https://doi.org/10.3390/pathophysiology31010008
Chicago/Turabian StyleFukatsu, Shoya, Hinami Sashi, Remina Shirai, Norio Takagi, Hiroaki Oizumi, Masahiro Yamamoto, Katsuya Ohbuchi, Yuki Miyamoto, and Junji Yamauchi. 2024. "Rab11a Controls Cell Shape via C9orf72 Protein: Possible Relationships to Frontotemporal Dementia/Amyotrophic Lateral Sclerosis (FTDALS) Type 1" Pathophysiology 31, no. 1: 100-116. https://doi.org/10.3390/pathophysiology31010008
APA StyleFukatsu, S., Sashi, H., Shirai, R., Takagi, N., Oizumi, H., Yamamoto, M., Ohbuchi, K., Miyamoto, Y., & Yamauchi, J. (2024). Rab11a Controls Cell Shape via C9orf72 Protein: Possible Relationships to Frontotemporal Dementia/Amyotrophic Lateral Sclerosis (FTDALS) Type 1. Pathophysiology, 31(1), 100-116. https://doi.org/10.3390/pathophysiology31010008