
High Glucose Induces Oxidative Stress That Alters Glycocalyx Proteoglycan Levels in Primary Rat Retinal Microvascular Endothelial Cells and in Isolated Ophthalmic Arteries



Abstract
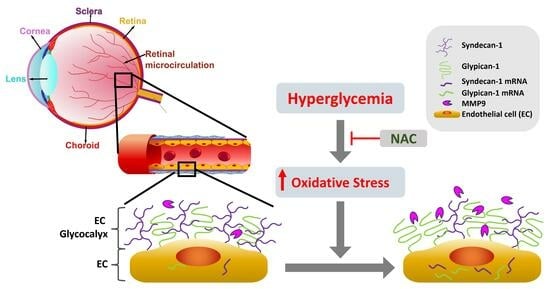
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Animals and Blood Vessel Tissue Culture
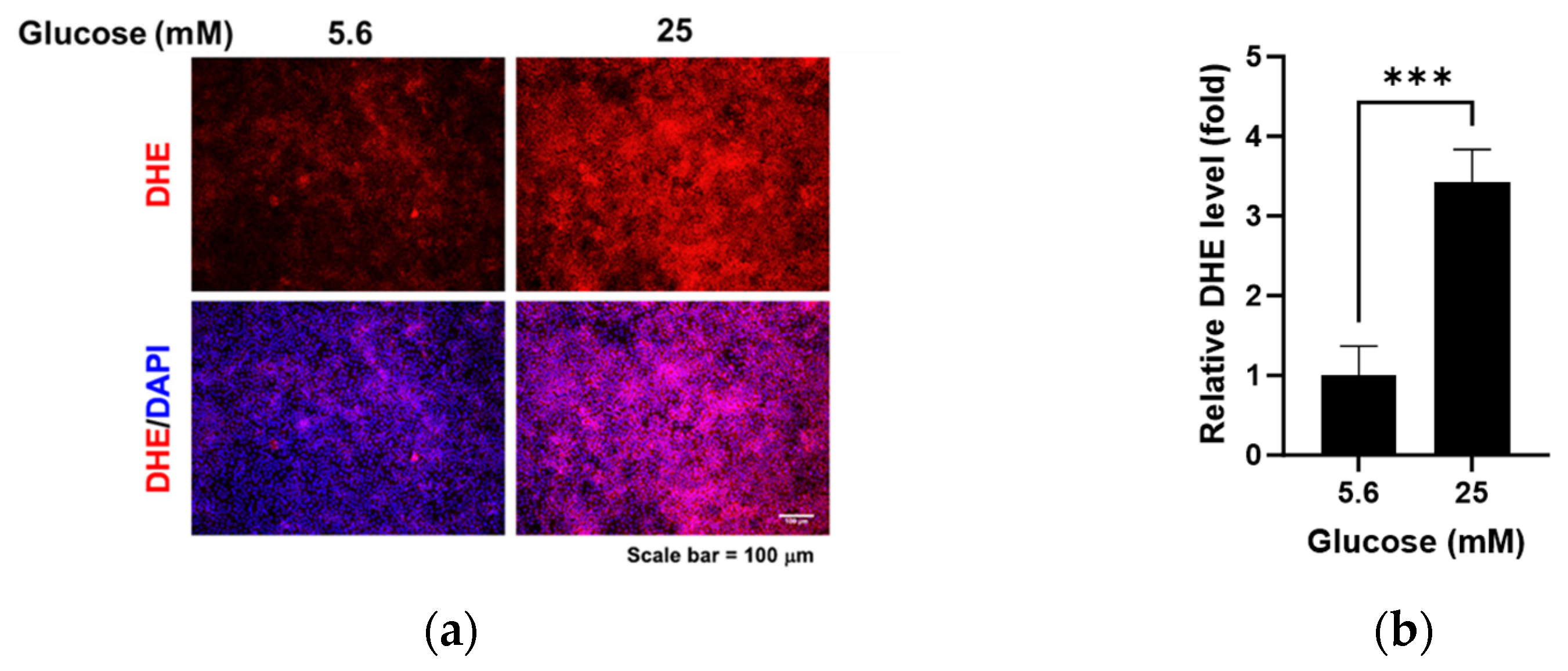
2.3. Measurement of Oxidative Stress
2.4. Immunoblot Analysis
2.5. Total RNA Isolation, Reverse Transcription, and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Statistics
3. Results
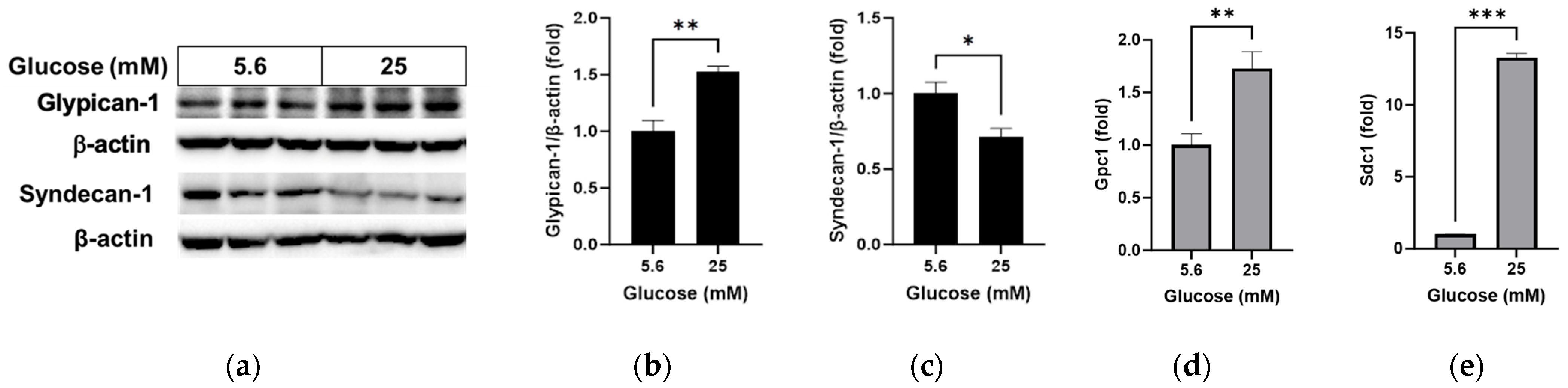
3.1. Effects of High Glucose on Glycocalyx Proteins and Oxidative Stress in RRMEC
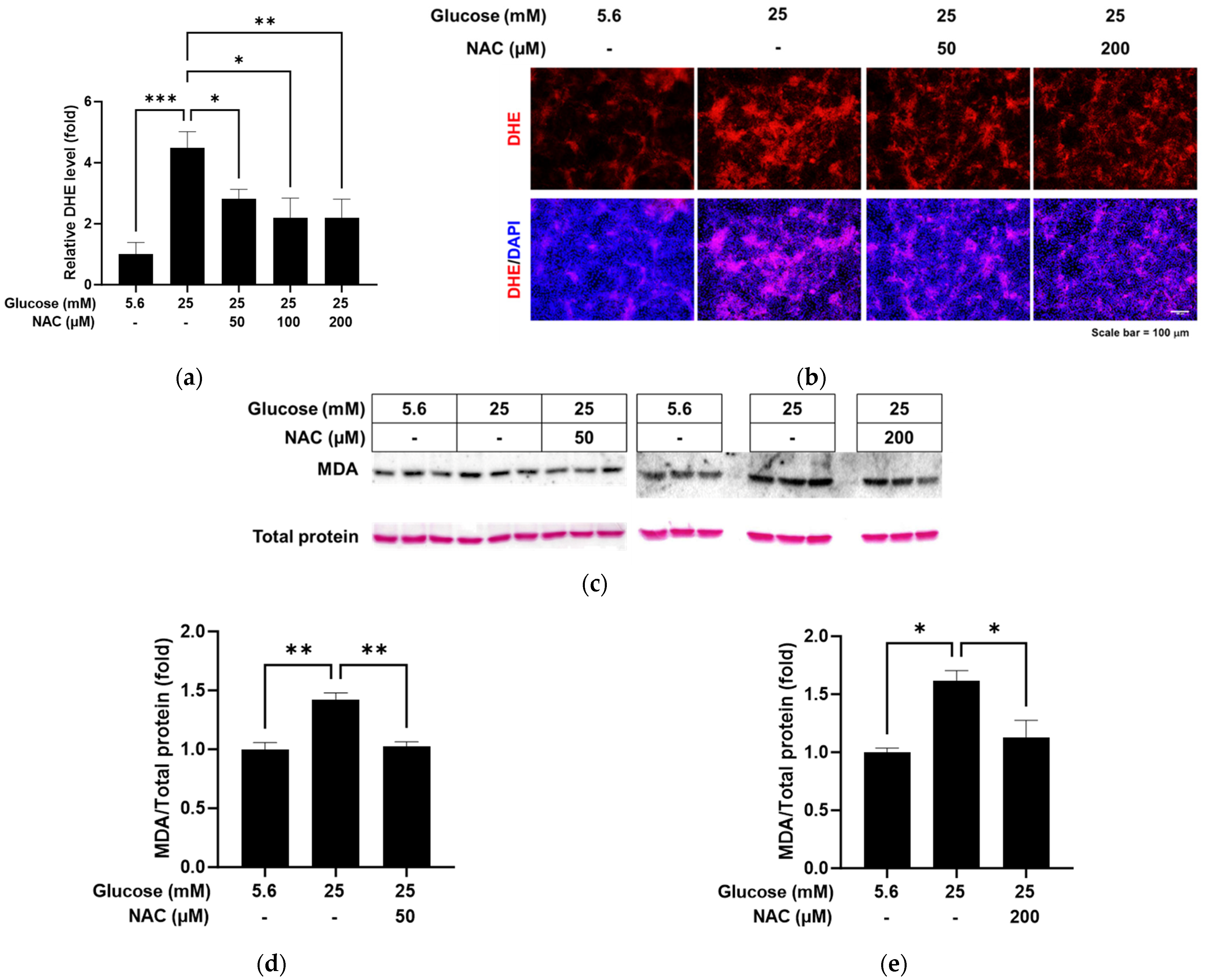
3.2. Effect of NAC on the High Glucose-Increased Oxidative Stress in RRMEC
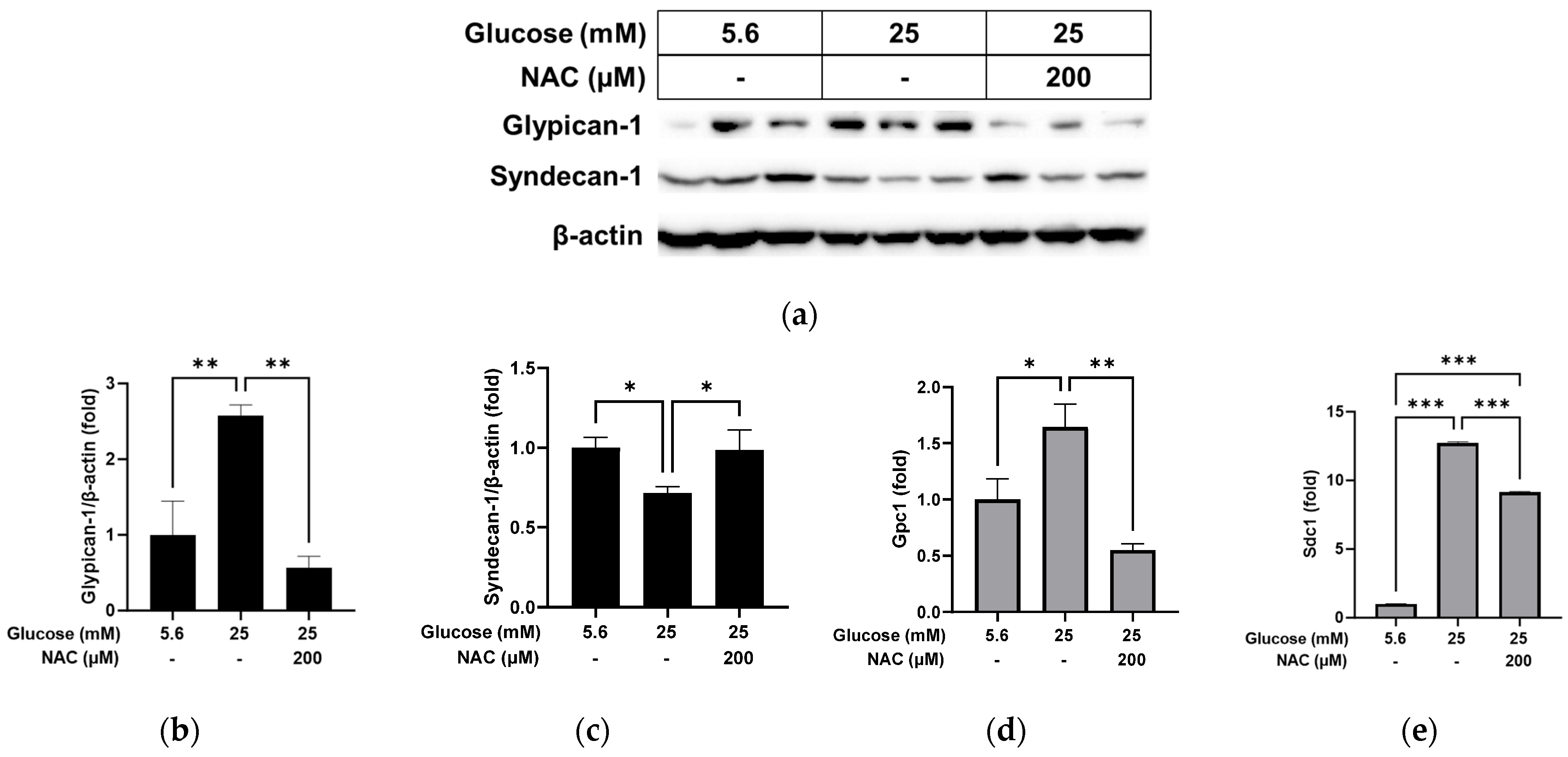
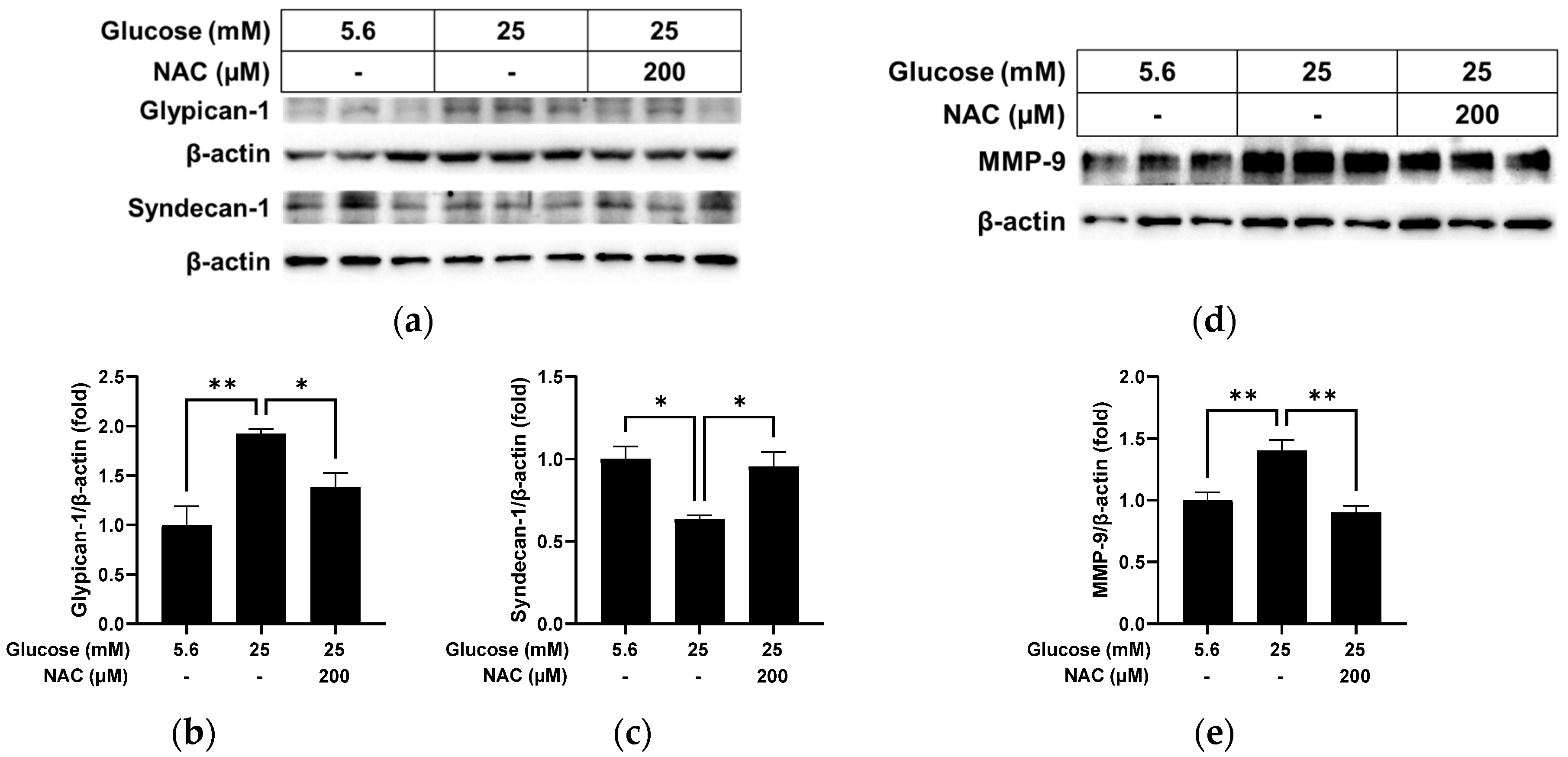
3.3. Effect of NAC on the Alterations Caused by High Glucose in RRMEC
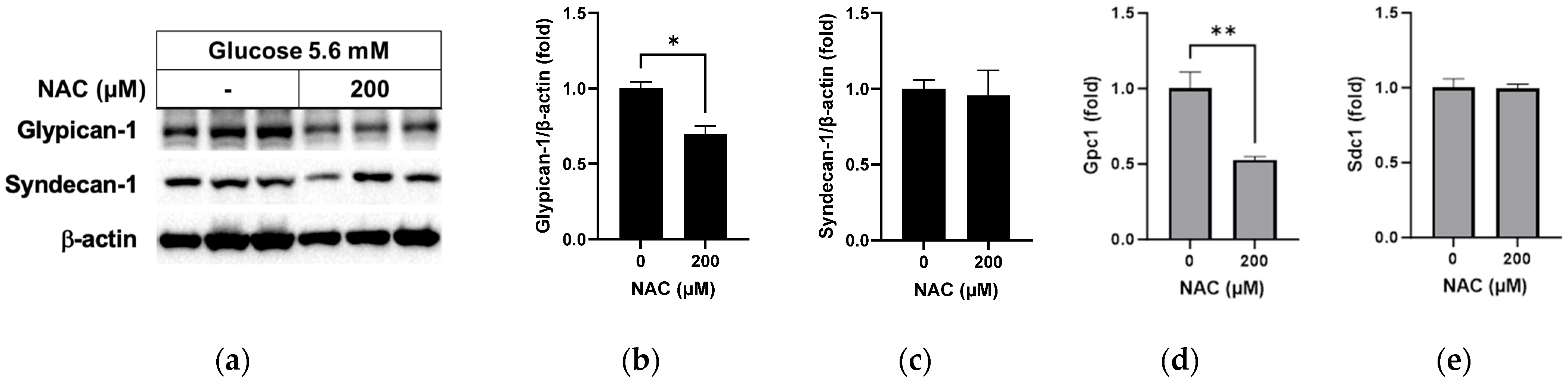
3.4. Effect of NAC on Protein Levels in RRMEC Incubated with Normal Glucose
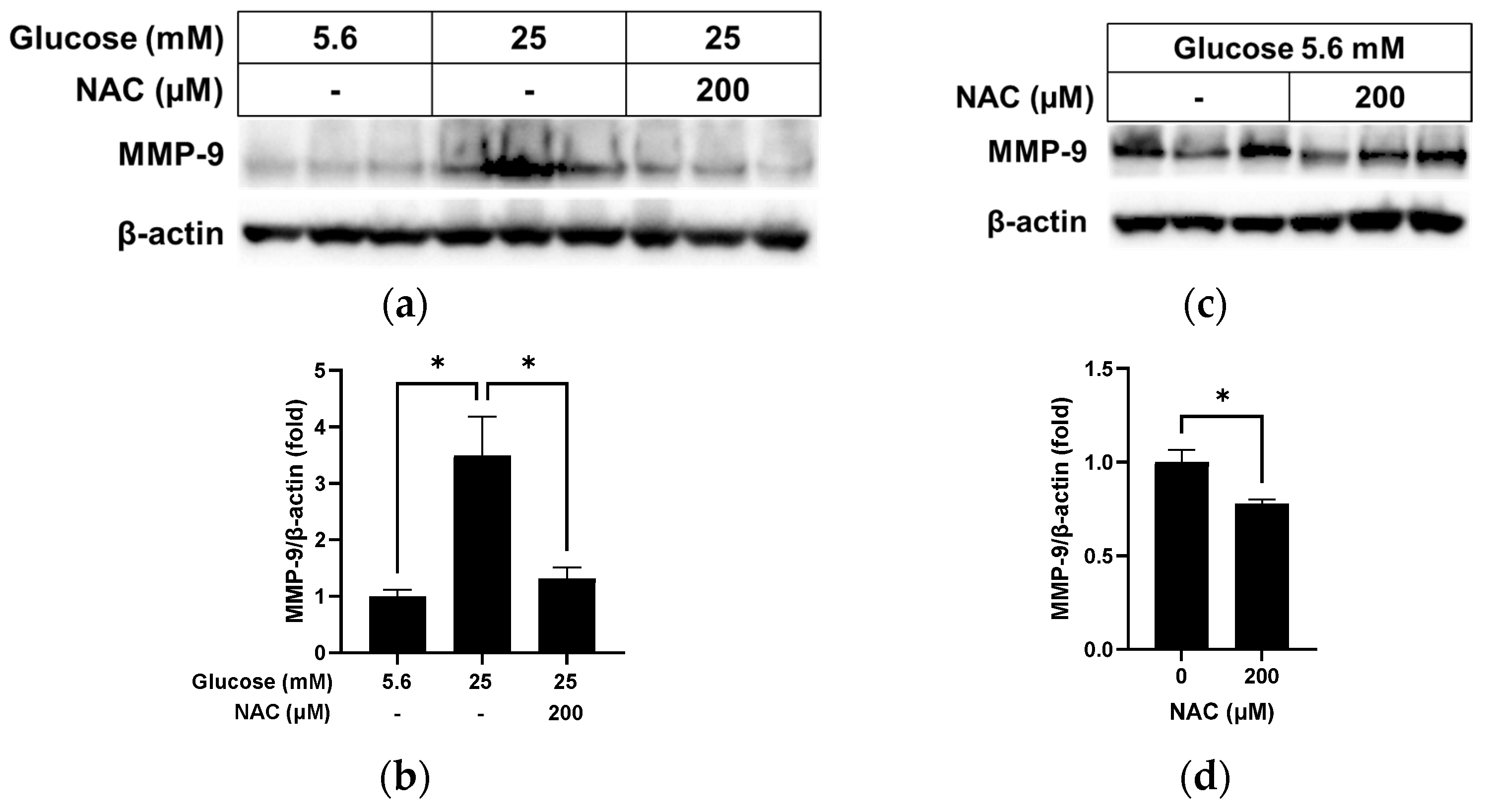
3.5. Effect of NAC on MMP-9 Levels in RRMEC Incubated with Normal and High Glucose
3.6. Effect of High Glucose and NAC on Ophthalmic Arteries
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, R.; Klein, B.E.; Moss, S.E.; Cruickshanks, K.J. Relationship of hyperglycemia to the long-term incidence and progression of diabetic retinopathy. Arch. Intern. Med. 1994, 154, 2169–2178. [Google Scholar] [CrossRef] [PubMed]
- Eshaq, R.S.; Aldalati, A.M.Z.; Alexander, J.S.; Harris, N.R. Diabetic retinopathy: Breaking the barrier. Pathophysiology 2017, 24, 229–241. [Google Scholar] [CrossRef]
- Leasher, J.L.; Bourne, R.R.; Flaxman, S.R.; Jonas, J.B.; Keeffe, J.; Naidoo, K.; Pesudovs, K.; Price, H.; White, R.A.; Wong, T.Y.; et al. Vision Loss Expert Group of the Global Burden of Disease Study, Global estimates on the number of people blind or visually impaired by diabetic retinopathy: A meta-analysis from 1990 to 2010. Diabetes Care 2016, 39, 1643–1649. [Google Scholar] [CrossRef] [PubMed]
- Antonetti, D.A.; Lieth, E.; Barber, A.J.; Gardner, T.W. Molecular mechanisms of vascular permeability in diabetic retinopathy. Semin. Ophthalmol. 1999, 14, 240–248. [Google Scholar] [CrossRef]
- Ishibazawa, A.; Nagaoka, T.; Yokota, H.; Takahashi, A.; Omae, T.; Song, Y.S.; Takahashi, T.; Yoshida, A. Characteristics of retinal neovascularization in proliferative diabetic retinopathy imaged by optical coherence tomography angiography. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6247–6255. [Google Scholar] [CrossRef]
- Schroder, S.; Palinski, W.; Schmid-Schonbein, G.W. Activated monocytes and granulocytes, capillary nonperfusion, and neovascularization in diabetic retinopathy. Am. J. Pathol. 1991, 139, 81–100. [Google Scholar]
- Ashton, N.; Harry, J. The pathology of cotton wool spots and cytoid bodies in hypertensive retinopathy and other diseases. Trans. Ophthalmol. Soc. U. K. 1963, 83, 91–114. [Google Scholar] [PubMed]
- Kaur, G.; Rogers, J.; Rashdan, N.A.; Cruz-Topete, D.; Pattillo, C.B.; Hartson, S.D.; Harris, N.R. Hyperglycemia-induced effects on glycocalyx components in the retina. Exp. Eye Res. 2021, 213, 108846. [Google Scholar] [CrossRef]
- Kaur, G.; Song, Y.; Xia, K.; McCarthy, K.; Zhang, F.; Linhardt, R.J.; Harris, N.R. Effect of high glucose on glycosaminoglycans in cultured retinal endothelial cells and rat retina. Glycobiology 2022, 32, 720–734. [Google Scholar] [CrossRef]
- Leskova, W.; Pickett, H.; Eshaq, R.S.; Shrestha, B.; Pattillo, C.B.; Harris, N.R. Effect of diabetes and hyaluronidase on the retinal endothelial glycocalyx in mice. Exp. Eye Res. 2019, 179, 125–131. [Google Scholar] [CrossRef]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting free radicals in oxidative stress-related human diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants—Quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: Initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Catala, A. Lipid peroxidation of membrane phospholipids in the vertebrate retina. Front. Biosci. Sch. Ed. 2011, 3, 52–60. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ceriello, A.; Testa, R.; Genovese, S. Clinical implications of oxidative stress and potential role of natural antioxidants in diabetic vascular complications. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Hikmah, N.; Dewi, A.; Shita, A.; Maulana, H. Rat Diabetic Blood Glucose Level Profile with Stratified Dose Streptozotocin (SD-STZ) and Multi Low Dose Streptozotocin (MLD-STZ) Induction Methods. J. Trop. Life Sci. 2015, 5, 30–34. [Google Scholar]
- Zhang, D.; Zhang, J.T.; Pan, Y.; Liu, X.F.; Xu, J.W.; Cui, W.J.; Qiao, X.R.; Dong, L. Syndecan-1 shedding by matrix metalloproteinase-9 signaling regulates alveolar epithelial tight junction in lipopolysaccharide-induced early acute lung injury. J. Inflamm. Res. 2021, 14, 5801–5816. [Google Scholar] [CrossRef] [PubMed]
- Eshaq, R.S.; Watts, M.N.; Carter, P.R.; Leskova, W.; Aw, T.Y.; Alexander, J.S.; Harris, N.R. Candesartan Normalizes Changes in Retinal Blood Flow and p22phox in the Diabetic Rat Retina. Pathophysiology 2021, 28, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, T.V.; Prioletta, A.; Zuo, P.; Folli, F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5695–5703. [Google Scholar] [CrossRef] [PubMed]
- Haidara, M.A.; Yassin, H.Z.; Rateb, M.; Ammar, H.; Zorkani, M.A. Role of oxidative stress in development of cardiovascular complications in diabetes mellitus. Curr. Vasc. Pharmacol. 2006, 4, 215–227. [Google Scholar] [CrossRef]
- Johansen, J.S.; Harris, A.K.; Rychly, D.J.; Ergul, A. Oxidative stress and the use of antioxidants in diabetes: Linking basic science to clinical practice. Cardiovasc. Diabetol. 2005, 4, 5. [Google Scholar] [CrossRef]
- Uemura, S.; Matsushita, H.; Li, W.; Glassford, A.J.; Asagami, T.; Lee, K.H.; Harrison, D.G.; Tsao, P.S. Diabetes mellitus enhances vascular matrix metalloproteinase activity: Role of oxidative stress. Circ. Res. 2001, 88, 1291–1298. [Google Scholar] [CrossRef]
- Sagara, M.; Satoh, J.; Wada, R.; Yagihashi, S.; Takahashi, K.; Fukuzawa, M.; Muto, G.; Muto, Y.; Toyota, T. Inhibition of development of peripheral neuropathy in streptozotocin-induced diabetic rats with N-acetylcysteine. Diabetologia 1996, 39, 263–269. [Google Scholar] [CrossRef]
- Gibson, K.R.; Winterburn, T.J.; Barrett, F.; Sharma, S.; MacRury, S.M.; Megson, I.L. Therapeutic potential of N-acetylcysteine as an antiplatelet agent in patients with type-2 diabetes. Cardiovasc. Diabetol. 2011, 10, 43. [Google Scholar] [CrossRef]
- Peng, N.; Geng, Y.; Ouyang, J.; Liu, S.; Yuan, F.; Wan, Y.; Chen, W.; Yu, B.; Tang, Y.; Su, L.; et al. Endothelial glycocalyx injury is involved in heatstroke-associated coagulopathy and protected by N-acetylcysteine. Front. Immunol. 2023, 14, 1159195. [Google Scholar] [CrossRef]
- Wu, W.Z.; Bai, Y.P. Endothelial GLUTs and vascular biology. Biomed. Pharmacother. 2023, 158, 114151. [Google Scholar] [CrossRef]
- Vink, H.; Constantinescu, A.A.; Spaan, J.A. Oxidized lipoproteins degrade the endothelial surface layer: Implications for platelet-endothelial cell adhesion. Circulation 2000, 101, 1500–1502. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; dello Russo, P.; Amstad, P.; Cerutti, P. High glucose induces antioxidant enzymes in human endothelial cells in culture. Evidence linking hyperglycemia and oxidative stress. Diabetes 1996, 45, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Marklund, S.L. Interactions between Human Extracellular Superoxide Dismutase C and Sulfated Polysaccharides. J. Biol. Chem. 1989, 264, 8537–8541. [Google Scholar] [CrossRef] [PubMed]
- Kliment, C.R.; Tobolewski, J.M.; Manni, M.L.; Tan, R.J.; Enghild, J.; Oury, T.D. Extracellular superoxide dismutase protects against matrix degradation of heparan sulfate in the lung. Antioxid. Redox Signal. 2008, 10, 261–268. [Google Scholar] [CrossRef]
- Adachi, T.; Yamada, H.; Yamada, Y.; Morihara, N.; Yamazaki, N.; Murakami, T.; Futenma, A.; Kato, K.; Hirano, K. Substitution of glycine for arginine-213 in extracellular-superoxide dismutase impairs affinity for heparin and endothelial cell surface. Biochem. J. 1996, 313 Pt 1, 235–239. [Google Scholar] [CrossRef]
- Sandstrom, J.; Nilsson, P.; Karlsson, K.; Marklund, S.L. 10-fold increase in human plasma extracellular superoxide dismutase content caused by a mutation in heparin-binding domain. J. Biol. Chem. 1994, 269, 19163–19166. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Abe, Y.; Takayama, M.; Adachi, T.; Okano, H.; Hirose, N.; Arai, Y. Association among extracellular superoxide dismutase genotype, plasma concentration, and comorbidity in the very old and centenarians. Sci. Rep. 2021, 11, 8539. [Google Scholar] [CrossRef] [PubMed]
- Rapraeger, A.C.; Ell, B.J.; Roy, M.; Li, X.; Morrison, O.R.; Thomas, G.M.; Beauvais, D.M. Vascular endothelial-cadherin stimulates syndecan-1-coupled insulin-like growth factor-1 receptor and cross-talk between alphaVbeta3 integrin and vascular endothelial growth factor receptor 2 at the onset of endothelial cell dissemination during angiogenesis. FEBS J. 2013, 280, 2194–2206. [Google Scholar] [PubMed]
- Drost, C.C.; Rovas, A.; Kumpers, P. Protection and rebuilding of the endothelial glycocalyx in sepsis—Science or fiction? Matrix Biol. Plus 2021, 12, 100091. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.P.; Yang, Y.; Janssen, W.J.; Gandjeva, A.; Perez, M.J.; Barthel, L.; Zemans, R.L.; Bowman, J.C.; Koyanagi, D.E.; Yunt, Z.X.; et al. The pulmonary endothelial glycocalyx regulates neutrophil adhesion and lung injury during experimental sepsis. Nat. Med. 2012, 18, 1217–1223. [Google Scholar] [CrossRef]
- Yang, X.; Meegan, J.E.; Jannaway, M.; Coleman, D.C.; Yuan, S.Y. A disintegrin and metalloproteinase 15-mediated glycocalyx shedding contributes to vascular leakage during inflammation. Cardiovasc. Res. 2018, 114, 1752–1763. [Google Scholar] [CrossRef] [PubMed]
- Jacqueminet, S.; Ben Abdesselam, O.; Chapman, M.J.; Nicolay, N.; Foglietti, M.J.; Grimaldi, A.; Beaudeux, J.L. Elevated circulating levels of matrix metalloproteinase-9 in type 1 diabetic patients with and without retinopathy. Clin. Chim. Acta 2006, 367, 103–107. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Accession No. | Gene Name | Primer Pair (5′ to 3′) |
|---|---|---|---|
| Gpc1 | NM_030828.2 | Glypican 1 | GCTGCTGGAATGGGATTT |
| ATGTCCACCTCCACTTCA | |||
| Ppia | NM_017101.1 | Peptidylprolyl isomerase A | TGTGGCCCTCCTACATAAA |
| AGTAGGAGACTAACCACGTG | |||
| Sdc1 | NM_013026.2 | Syndecan 1 | CCAAATCCGGACACCAAAGGGCACCAAACAGATAGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, I.A.; Lee, M.; Eshaq, R.S.; Leskova, W.; Harris, N.R. High Glucose Induces Oxidative Stress That Alters Glycocalyx Proteoglycan Levels in Primary Rat Retinal Microvascular Endothelial Cells and in Isolated Ophthalmic Arteries. Pathophysiology 2024, 31, 89-99. https://doi.org/10.3390/pathophysiology31010007
Alvarez IA, Lee M, Eshaq RS, Leskova W, Harris NR. High Glucose Induces Oxidative Stress That Alters Glycocalyx Proteoglycan Levels in Primary Rat Retinal Microvascular Endothelial Cells and in Isolated Ophthalmic Arteries. Pathophysiology. 2024; 31(1):89-99. https://doi.org/10.3390/pathophysiology31010007
Chicago/Turabian StyleAlvarez, Ivan A., Minsup Lee, Randa S. Eshaq, Wendy Leskova, and Norman R. Harris. 2024. "High Glucose Induces Oxidative Stress That Alters Glycocalyx Proteoglycan Levels in Primary Rat Retinal Microvascular Endothelial Cells and in Isolated Ophthalmic Arteries" Pathophysiology 31, no. 1: 89-99. https://doi.org/10.3390/pathophysiology31010007
APA StyleAlvarez, I. A., Lee, M., Eshaq, R. S., Leskova, W., & Harris, N. R. (2024). High Glucose Induces Oxidative Stress That Alters Glycocalyx Proteoglycan Levels in Primary Rat Retinal Microvascular Endothelial Cells and in Isolated Ophthalmic Arteries. Pathophysiology, 31(1), 89-99. https://doi.org/10.3390/pathophysiology31010007