
Gut Microbiome Changes in Anorexia Nervosa: A Comprehensive Review



Abstract
1. Introduction
- The qualitative and quantitative changes in gut microbiome composition in patients with AN, compared to healthy populations (HC) or patients with other psychiatric disorders;
- The relationship between changes in gut microbiome composition and the risk of refeeding syndrome in AN patients.
2. Materials and Methods
3. Results
3.1. Research Question 1 Findings
3.1.1. AN Patients Compared to Healthy Controls (HCs)
3.1.2. AN Patients Compared to Patients with Other Psychiatric Disorders
4. Discussion
4.1. Changes in Gut Microbiome Composition
4.1.1. Alpha Diversity
4.1.2. Decreased Faecalibacterium prausnitzii
4.1.3. Decreased Roseburia inulinivorans
4.1.4. Increased Methanobrevibacter smithii
4.1.5. Comparison to Other Existing Pathophysiological Models
4.2. Refeeding Syndrome
4.3. Future Directions
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Saier, M.H., Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef] [PubMed]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Kelly, J.R.; Borre, Y.; O’Brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G.; et al. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T.; et al. Longitudinal multi-omics of host–microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Hawinkel, S.; Mattiello, F.; Bijnens, L.; Thas, O. A broken promise: Microbiome differential abundance methods do not control the false discovery rate. Brief. Bioinform. 2017, 20, 210–221. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing, Inc.: Arlington, VA, USA, 2013. [Google Scholar]
- Arcelus, J.; Mitchell, A.J.; Wales, J.; Nielsen, S. Mortality Rates in Patients With Anorexia Nervosa and Other Eating Disorders: A Meta-analysis of 36 Studies. Arch. Gen. Psychiatry 2011, 68, 724–731. [Google Scholar] [CrossRef]
- Hans-Christoph Steinhausen, M.D. The Outcome of Anorexia Nervosa in the 20th Century. Am. J. Psychiatry 2002, 159, 1284–1293. [Google Scholar] [CrossRef]
- Zipfel, S.; Giel, K.E.; Bulik, C.M.; Hay, P.; Schmidt, U. Anorexia nervosa: Aetiology, assessment, and treatment. Lancet Psychiatry 2015, 2, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.J.; Yilmaz, Z.; Thornton, L.M.; Hübel, C.; Coleman, J.R.I.; Gaspar, H.A.; Bryois, J.; Hinney, A.; Leppä, V.M.; Mattheisen, M.; et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 2019, 51, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Barakat, S.; McLean, S.A.; Bryant, E.; Le, A.; Marks, P.; Aouad, P.; Barakat, S.; Boakes, R.; Brennan, L.; Bryant, E.; et al. Risk factors for eating disorders: Findings from a rapid review. J. Eat. Disord. 2023, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Bulik, C.M.; Flatt, R.; Abbaspour, A.; Carroll, I. Reconceptualizing anorexia nervosa. Psychiatry Clin. Neurosci. 2019, 73, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Hrncir, T. Gut Microbiota Dysbiosis: Triggers, Consequences, Diagnostic and Therapeutic Options. Microorganisms 2022, 10, 578. [Google Scholar] [CrossRef] [PubMed]
- NICE. Nutrition Support for Adults: Oral Nutrition Support, Enteral Tube Feeding and Parenteral Nutrition; National Institute for Health and Care Excellence (NICE): London, UK, 2017. [Google Scholar]
- Kruizenga, H.M.; Seidell, J.C.; de Vet, H.C.; Wierdsma, N.J.; van Bokhorst-de van der Schueren, M.A. Development and validation of a hospital screening tool for malnutrition: The short nutritional assessment questionnaire (SNAQ). Clin. Nutr. 2005, 24, 75–82. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.S.V.; Seres, D.S.; Sabino, K.; Adams, S.C.; Berdahl, G.J.; Citty, S.W.; Cober, M.P.; Evans, D.C.; Greaves, J.R.; Gura, K.M.; et al. ASPEN Consensus Recommendations for Refeeding Syndrome. Nutr. Clin. Pract. 2020, 35, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Heuft, L.; Voigt, J.; Selig, L.; Stumvoll, M.; Schlögl, H.; Kaiser, T. Refeeding Syndrome. Dtsch. Arztebl. Int. 2023, 120, 107–114. [Google Scholar] [CrossRef]
- Ponzo, V.; Pellegrini, M.; Cioffi, I.; Scaglione, L.; Bo, S. The Refeeding Syndrome: A neglected but potentially serious condition for inpatients. A narrative review. Intern. Emerg. Med. 2021, 16, 49–60. [Google Scholar] [CrossRef]
- Carbone, E.A.; D’Amato, P.; Vicchio, G.; De Fazio, P.; Segura-Garcia, C. A systematic review on the role of microbiota in the pathogenesis and treatment of eating disorders. Eur. Psychiatry J. Assoc. Eur. Psychiatr. 2020, 64, e2. [Google Scholar] [CrossRef]
- Ghenciulescu, A.; Park, R.J.; Burnet, P.W.J. The Gut Microbiome in Anorexia Nervosa: Friend or Foe? Front. Psychiatry 2020, 11, 611677. [Google Scholar] [CrossRef] [PubMed]
- Di Lodovico, L.; Mondot, S.; Dore, J.; Mack, I.; Hanachi, M.; Gorwood, P. Anorexia nervosa and gut microbiota: A systematic review and quantitative synthesis of pooled microbiological data. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110114. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.; Gutierrez, E. Anorexia nervosa and microbiota: Systematic review and critical appraisal. Eat. Weight. Disord. 2023, 28, 1. [Google Scholar] [CrossRef] [PubMed]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef]
- Borgo, F.; Riva, A.; Benetti, A.; Casiraghi, M.C.; Bertelli, S.; Garbossa, S.; Anselmetti, S.; Scarone, S.; Pontiroli, A.E.; Morace, G.; et al. Microbiota in anorexia nervosa: The triangle between bacterial species, metabolites and psychological tests. PLoS ONE 2017, 12, e0179739. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Stoving, R.K.; Berreira Ibraim, S.; Hyotylainen, T.; Thirion, F.; Arora, T.; Lyu, L.; Stankevic, E.; Hansen, T.H.; Dechelotte, P.; et al. The gut microbiota contributes to the pathogenesis of anorexia nervosa in humans and mice. Nat. Microbiol. 2023, 8, 787–802. [Google Scholar] [CrossRef]
- Fouladi, F.; Bulik-Sullivan, E.C.; Glenny, E.M.; Thornton, L.M.; Reed, K.K.; Thomas, S.; Kleiman, S.; Watters, A.; Oakes, J.; Huh, E.-Y.; et al. Reproducible changes in the anorexia nervosa gut microbiota following inpatient therapy remain distinct from non-eating disorder controls. Gut Microbes 2022, 14, 2143217. [Google Scholar] [CrossRef]
- Hanachi, M.; Manichanh, C.; Schoenenberger, A.; Pascal, V.; Levenez, F.; Cournede, N.; Dore, J.; Melchior, J.-C. Altered host-gut microbes symbiosis in severely malnourished anorexia nervosa (AN) patients undergoing enteral nutrition: An explicative factor of functional intestinal disorders? Clin. Nutr. 2019, 38, 2304–2310. [Google Scholar] [CrossRef]
- Kleiman, S.C.; Carroll, I.M.; Tarantino, L.M.; Bulik, C.M. Gut feelings: A role for the intestinal microbiota in anorexia nervosa? Int. J. Eat. Disord. 2015, 48, 449–451. [Google Scholar] [CrossRef]
- Mack, I.; Cuntz, U.; Gramer, C.; Niedermaier, S.; Pohl, C.; Schwiertz, A.; Zimmermann, K.; Zipfel, S.; Enck, P.; Penders, J. Weight gain in anorexia nervosa does not ameliorate the faecal microbiota, branched chain fatty acid profiles, and gastrointestinal complaints. Sci. Rep. 2016, 6, 26752. [Google Scholar] [CrossRef]
- Million, M.; Angelakis, E.; Maraninchi, M.; Henry, M.; Giorgi, R.; Valero, R.; Vialettes, B.; Raoult, D. Correlation between body mass index and gut concentrations of Lactobacillus reuteri, Bifidobacterium animalis, Methanobrevibacter smithii and Escherichia coli. Int. J. Obes. 2013, 37, 1460–1466. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, A.M.; Troisi, J.; Fasano, A.; Dalle Grave, R.; Marciello, F.; Serena, G.; Calugi, S.; Scala, G.; Corrivetti, G.; Cascino, G.; et al. Multi-omics data integration in anorexia nervosa patients before and after weight regain: A microbiome-metabolomics investigation. Clin. Nutr. 2021, 40, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Morita, C.; Tsuji, H.; Hata, T.; Gondo, M.; Takakura, S.; Kawai, K.; Yoshihara, K.; Ogata, K.; Nomoto, K.; Miyazaki, K.; et al. Gut Dysbiosis in Patients with Anorexia Nervosa. PLoS ONE 2015, 10, e0145274. [Google Scholar] [CrossRef] [PubMed]
- Morkl, S.; Lackner, S.; Muller, W.; Gorkiewicz, G.; Kashofer, K.; Oberascher, A.; Painold, A.; Holl, A.; Holzer, P.; Meinitzer, A.; et al. Gut microbiota and body composition in anorexia nervosa inpatients in comparison to athletes, overweight, obese, and normal weight controls. Int. J. Eat. Disord. 2017, 50, 1421–1431. [Google Scholar] [CrossRef]
- Prochazkova, P.; Roubalova, R.; Dvorak, J.; Kreisinger, J.; Hill, M.; Tlaskalova-Hogenova, H.; Tomasova, P.; Pelantova, H.; Cermakova, M.; Kuzma, M.; et al. The intestinal microbiota and metabolites in patients with anorexia nervosa. Gut Microbes 2021, 13, 1902771. [Google Scholar] [CrossRef] [PubMed]
- Schulz, N.; Belheouane, M.; Dahmen, B.; Ruan, V.A.; Specht, H.E.; Dempfle, A.; Herpertz-Dahlmann, B.; Baines, J.F.; Seitz, J. Gut microbiota alteration in adolescent anorexia nervosa does not normalize with short-term weight restoration. Int. J. Eat. Disord. 2021, 54, 969–980. [Google Scholar] [CrossRef]
- Yuan, R.; Yang, L.; Yao, G.; Geng, S.; Ge, Q.; Bo, S.; Li, X. Features of gut microbiota in patients with anorexia nervosa. Chin. Med. J. 2022, 135, 1993–2002. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, J. Alpha Diversity. In Bioinformatic and Statistical Analysis of Microbiome Data: From Raw Sequences to Advanced Modeling with QIIME 2 and R; Xia, Y., Sun, J., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 289–333. [Google Scholar]
- Kelly, J.; Kennedy, P.; Cryan, J.; Dinan, T.; Clarke, G.; Hyland, N. Breaking Down the Barriers: The Gut Microbiome, Intestinal Permeability and Stress-related Psychiatric Disorders. Front. Cell. Neurosci. 2015, 9, 392. [Google Scholar] [CrossRef]
- Shade, A. Diversity is the question, not the answer. ISME J. 2017, 11, 1–6. [Google Scholar] [CrossRef]
- Coretti, L.; Paparo, L.; Riccio, M.P.; Amato, F.; Cuomo, M.; Natale, A.; Borrelli, L.; Corrado, G.; De Caro, C.; Comegna, M.; et al. Gut Microbiota Features in Young Children with Autism Spectrum Disorders. Front. Microbiol. 2018, 9, 3146. [Google Scholar] [CrossRef]
- Yin, J.; Liao, S.X.; He, Y.; Wang, S.; Xia, G.H.; Liu, F.T.; Zhu, J.J.; You, C.; Chen, Q.; Zhou, L.; et al. Dysbiosis of Gut Microbiota With Reduced Trimethylamine-N-Oxide Level in Patients With Large-Artery Atherosclerotic Stroke or Transient Ischemic Attack. J. Am. Heart Assoc. 2015, 4, e002699. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.T.; Marotz, C.; Washburne, A.; Silverman, J.; Zaramela, L.S.; Edlund, A.; Zengler, K.; Knight, R. Establishing microbial composition measurement standards with reference frames. Nat. Commun. 2019, 10, 2719. [Google Scholar] [CrossRef] [PubMed]
- Allaband, C.; McDonald, D.; Vázquez-Baeza, Y.; Minich, J.J.; Tripathi, A.; Brenner, D.A.; Loomba, R.; Smarr, L.; Sandborn, W.J.; Schnabl, B.; et al. Microbiome 101: Studying, Analyzing, and Interpreting Gut Microbiome Data for Clinicians. Clin. Gastroenterol. Hepatol. 2019, 17, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Lee, G.; Son, H.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.H. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front. Microbiol. 2022, 13, 1103836. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Halder, C.V.; Faria, A.V.d.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte metabolism shapes the gut microbiota. Science 2018, 362, eaat9076. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef]
- Aizawa, E.; Tsuji, H.; Asahara, T.; Takahashi, T.; Teraishi, T.; Yoshida, S.; Ota, M.; Koga, N.; Hattori, K.; Kunugi, H. Possible association of Bifidobacterium and Lactobacillus in the gut microbiota of patients with major depressive disorder. J. Affect. Disord. 2016, 202, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Misra, M.; Klibanski, A. Endocrine consequences of anorexia nervosa. Lancet Diabetes Endocrinol. 2014, 2, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Martin, J.C.; Campbell, G.; Mayer, C.-D.; Flint, H.J. Whole-Genome Transcription Profiling Reveals Genes Up-Regulated by Growth on Fucose in the Human Gut Bacterium “Roseburia inulinivorans”. J. Bacteriol. 2006, 188, 4340–4349. [Google Scholar] [CrossRef]
- Pingitore, A.; Chambers, E.S.; Hill, T.; Maldonado, I.R.; Liu, B.; Bewick, G.; Morrison, D.J.; Preston, T.; Wallis, G.A.; Tedford, C.; et al. The diet-derived short chain fatty acid propionate improves beta-cell function in humans and stimulates insulin secretion from human islets in vitro. Diabetes Obes. Metab. 2017, 19, 257–265. [Google Scholar] [CrossRef] [PubMed]
- van de Pol, J.A.; van Best, N.; Mbakwa, C.A.; Thijs, C.; Savelkoul, P.H.; Arts, I.C.; Hornef, M.W.; Mommers, M.; Penders, J. Gut Colonization by Methanogenic Archaea Is Associated with Organic Dairy Consumption in Children. Front. Microbiol. 2017, 8, 355. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, T.; Massoubre, C.; Hanachi, M.; Doré, J.; Lambert, C.; Germain, N.; Galusca, B.; Paul, S. Association of gut-specific non-inflammatory T lymphocytes with chronic anorexia nervosa and constitutional thinness. Eur. Eat. Disord. Rev. 2023, 31, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, I.A.K.; Millischer, V.; Göteson, A.; Hübel, C.; Thornton, L.M.; Bulik, C.M.; Schalling, M.; Landén, M. Aberrant inflammatory profile in acute but not recovered anorexia nervosa. Brain Behav. Immun. 2020, 88, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Dalton, B.; Bartholdy, S.; Robinson, L.; Solmi, M.; Ibrahim, M.A.A.; Breen, G.; Schmidt, U.; Himmerich, H. A meta-analysis of cytokine concentrations in eating disorders. J. Psychiatr. Res. 2018, 103, 252–264. [Google Scholar] [CrossRef]
- Solmi, F.; Bulik, C.M.; De Stavola, B.L.; Dalman, C.; Khandaker, G.M.; Lewis, G. Longitudinal associations between circulating interleukin-6 and C-reactive protein in childhood, and eating disorders and disordered eating in adolescence. Brain Behav. Immun. 2020, 89, 491–500. [Google Scholar] [CrossRef]
- Himmerich, H.; Bentley, J.; Kan, C.; Treasure, J. Genetic risk factors for eating disorders: An update and insights into pathophysiology. Ther. Adv. Psychopharmacol. 2019, 9, 2045125318814734. [Google Scholar] [CrossRef]
- Breton, J.; Déchelotte, P.; Ribet, D. Intestinal microbiota and Anorexia Nervosa. Clin. Nutr. Exp. 2019, 28, 11–21. [Google Scholar] [CrossRef]
- Fetissov, S.O.; Hökfelt, T. On the origin of eating disorders: Altered signaling between gut microbiota, adaptive immunity and the brain melanocortin system regulating feeding behavior. Curr. Opin. Pharmacol. 2019, 48, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Garber, A.K.; Sawyer, S.M.; Golden, N.H.; Guarda, A.S.; Katzman, D.K.; Kohn, M.R.; Le Grange, D.; Madden, S.; Whitelaw, M.; Redgrave, G.W. A systematic review of approaches to refeeding in patients with anorexia nervosa. Int. J. Eat. Disord. 2016, 49, 293–310. [Google Scholar] [CrossRef]
- Mosuka, E.M.; Murugan, A.; Thakral, A.; Ngomo, M.C.; Budhiraja, S.; St Victor, R. Clinical Outcomes of Refeeding Syndrome: A Systematic Review of High vs. Low-Calorie Diets for the Treatment of Anorexia Nervosa and Related Eating Disorders in Children and Adolescents. Cureus 2023, 15, e39313. [Google Scholar] [CrossRef] [PubMed]
- The Society for Adolescent Health and Medicine. Refeeding Hypophosphatemia in Hospitalized Adolescents with Anorexia Nervosa. J. Adolesc. Health 2022, 71, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Jowik, K.; Tyszkiewicz-Nwafor, M.; Słopień, A. Anorexia Nervosa—What Has Changed in the State of Knowledge about Nutritional Rehabilitation for Patients over the Past 10 Years? A Review of Literature. Nutrients 2021, 13, 3819. [Google Scholar] [CrossRef]
- Ruusunen, A.; Rocks, T.; Jacka, F.; Loughman, A. The gut microbiome in anorexia nervosa: Relevance for nutritional rehabilitation. Psychopharmacology 2019, 236, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef]
- O’Connor, G.; Nicholls, D. Refeeding hypophosphatemia in adolescents with anorexia nervosa: A systematic review. Nutr. Clin. Pract. 2013, 28, 358–364. [Google Scholar] [CrossRef]
- Zaneveld, J.R.; McMinds, R.; Vega Thurber, R. Stress and stability: Applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2017, 2, 17121. [Google Scholar] [CrossRef]
- Prochazkova, P.; Roubalova, R.; Dvorak, J.; Tlaskalova-Hogenova, H.; Cermakova, M.; Tomasova, P.; Sediva, B.; Kuzma, M.; Bulant, J.; Bilej, M.; et al. Microbiota, Microbial Metabolites, and Barrier Function in A Patient with Anorexia Nervosa after Fecal Microbiota Transplantation. Microorganisms 2019, 7, 338. [Google Scholar] [CrossRef] [PubMed]
- de Clercq, N.C.; Frissen, M.N.; Davids, M.; Groen, A.K.; Nieuwdorp, M. Weight gain after fecal microbiota transplantation in a patient with recurrent underweight following clinical recovery from anorexia nervosa. Psychother. Psychosom. 2019, 88, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; Montes-Borrego, M.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Tinahones, F.J.; Landa, B.B.; et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J. Clin. Endocrinol. Metab. 2016, 101, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Cussotto, S.; Strain, C.R.; Fouhy, F.; Strain, R.G.; Peterson, V.L.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Differential effects of psychotropic drugs on microbiome composition and gastrointestinal function. Psychopharmacology 2019, 236, 1671–1685. [Google Scholar] [CrossRef]
- Bohnert, J.A.; Szymaniak-Vits, M.; Schuster, S.; Kern, W.V. Efflux inhibition by selective serotonin reuptake inhibitors in Escherichia coli. J. Antimicrob. Chemother. 2011, 66, 2057–2060. [Google Scholar] [CrossRef] [PubMed]
- Boulund, U.; Bastos, D.M.; Ferwerda, B.; van den Born, B.-J.; Pinto-Sietsma, S.-J.; Galenkamp, H.; Levin, E.; Groen, A.K.; Zwinderman, A.H.; Nieuwdorp, M. Gut microbiome associations with host genotype vary across ethnicities and potentially influence cardiometabolic traits. Cell Host Microbe 2022, 30, 1464–1480.e1466. [Google Scholar] [CrossRef]
- Rausch, P.; Rühlemann, M.; Hermes, B.M.; Doms, S.; Dagan, T.; Dierking, K.; Domin, H.; Fraune, S.; von Frieling, J.; Hentschel, U.; et al. Comparative analysis of amplicon and metagenomic sequencing methods reveals key features in the evolution of animal metaorganisms. Microbiome 2019, 7, 133. [Google Scholar] [CrossRef]
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the Gut Microbiome Using 16S or Shotgun Metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef]
- Laudadio, I.; Fulci, V.; Palone, F.; Stronati, L.; Cucchiara, S.; Carissimi, C. Quantitative Assessment of Shotgun Metagenomics and 16S rDNA Amplicon Sequencing in the Study of Human Gut Microbiome. Omics 2018, 22, 248–254. [Google Scholar] [CrossRef] [PubMed]
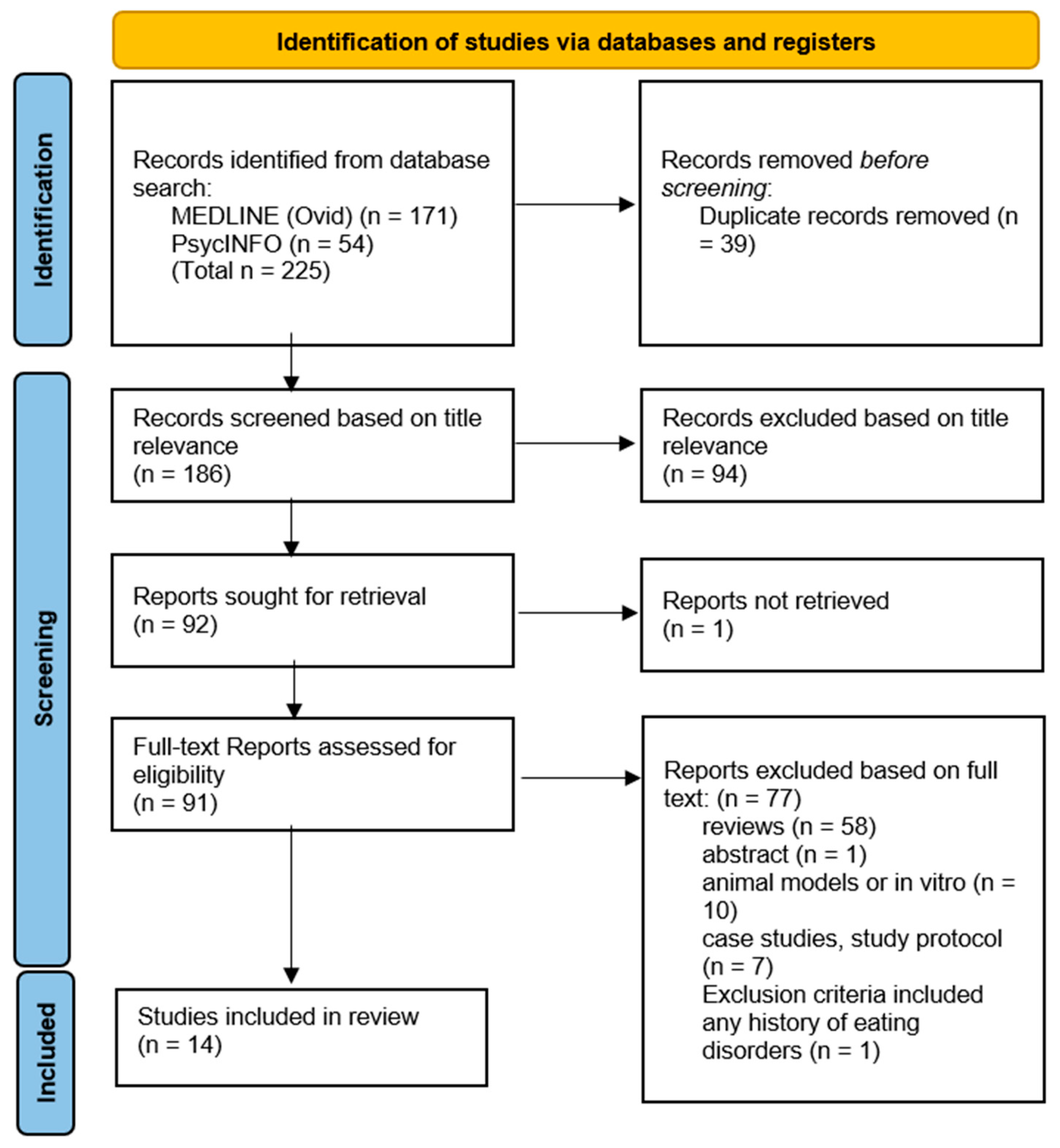


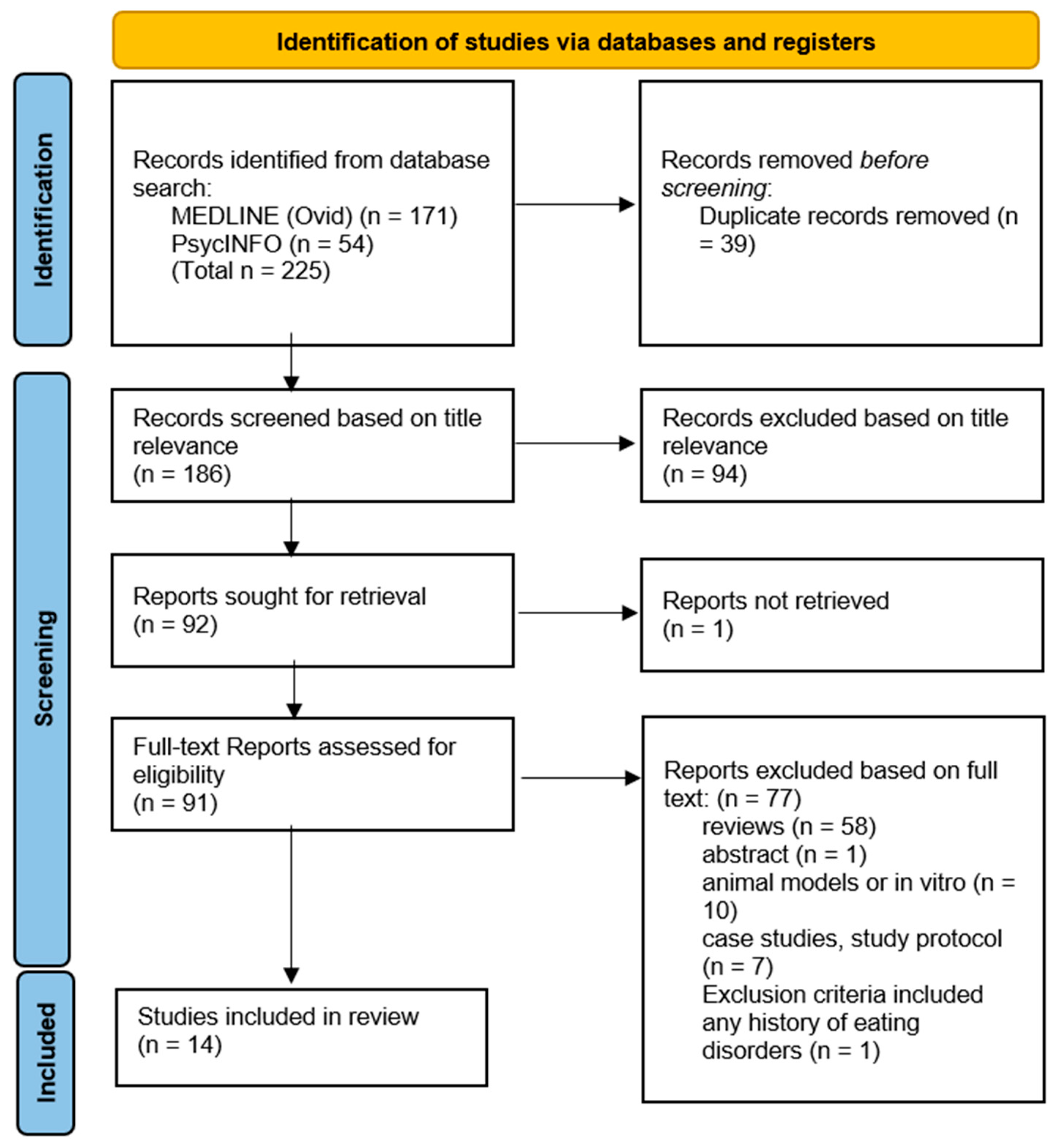{kind=link}
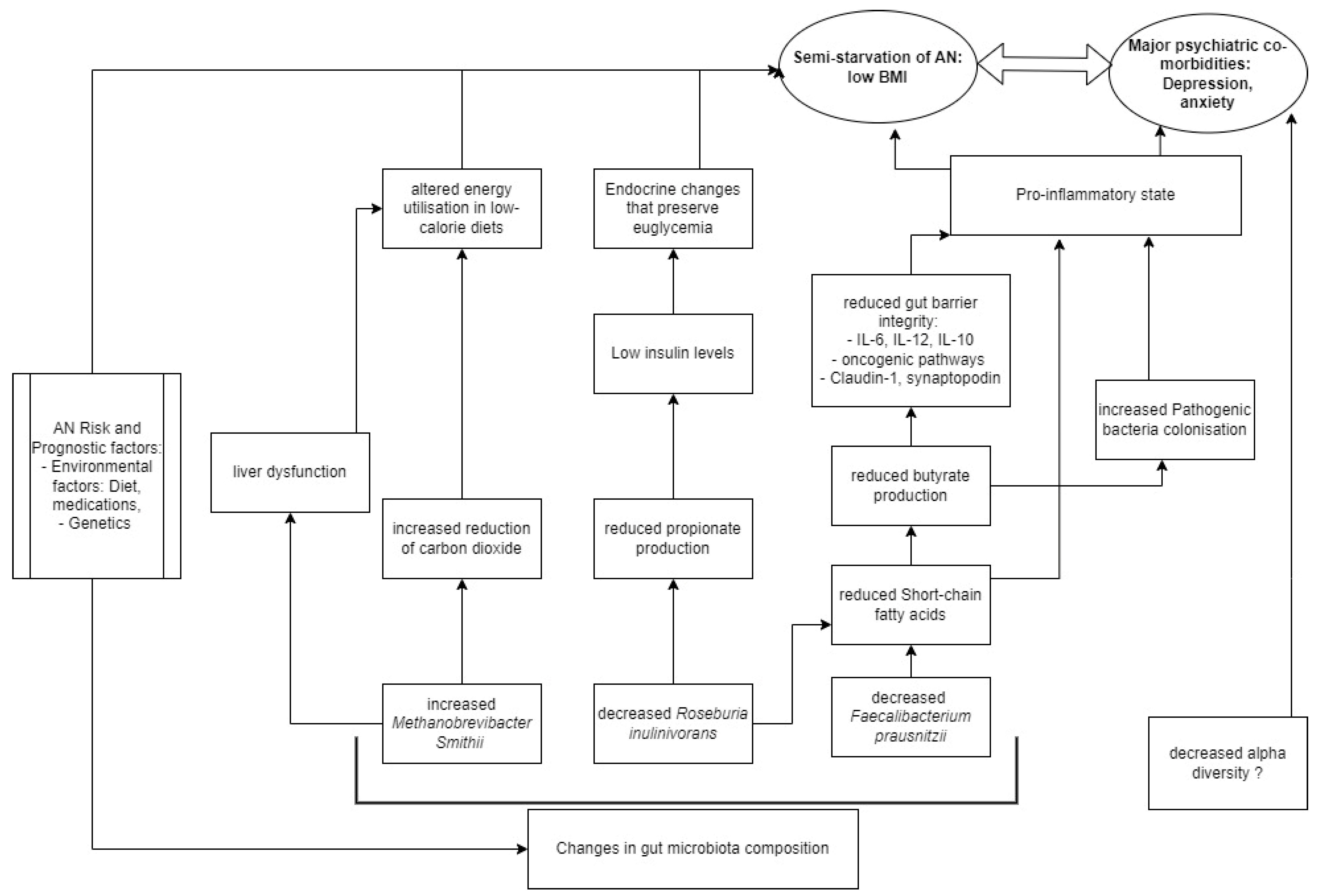{kind=link}
| (P)opulation | Patients diagnosed with AN according to DSM-IV, DSM-V, ICD-10, EDI-3 or other, patients of both sexes and all ethnicities, aged > 12 years old and BMI < 18.5 kg/m2. |
| (I)nvestigation | Assessment of gut microbiota composition. |
| (C)omparators | HC (healthy-weight control) group or patients with other psychiatric disorders. |
| (O)utcomes | Analysis of the gut microbiota derived from stool samples using shotgun metagenomic sequencing or 16S rRNA sequencing techniques and/or real-time polymerase chain reaction (rt-PCR). Measures for microbiota composition in patients with AN and HC:
|
| (S)tudy design | Cross-sectional or longitudinal studies. |
| References | Study Design | Sample | Analysis Technique for Gut Microbiome Composition | Changes in Gut Microbiome Composition in AN Patients Compared to Healthy Controls (Statistically Significant p < 0.05) | Changes in Gut Microbiome Composition in AN Patients Compared to Other Psychiatric Disorders | Therapeutic Interventions | Other Parameters | ||
|---|---|---|---|---|---|---|---|---|---|
| Qualitative Differences in Bacteria | Quantitative Differences in Bacteria | Metabolite Concentrations | |||||||
| Armougom et al. 2009 [26] | CS | 9 AN (BMI 12.73 kg/m2 ± 1.602) 20 HC (BMI 20.68 kg/m2 ± 2.014) N/A PSY | qPCR | ↑ M. smithii ↔ Firmicutes ↔ Bacteroidetes ↔ Lactobacillus | N/A | N/A | N/A | N/A | Positive correlation between BMI and Lactobacillus. Negative correlation between BMI and M. smithii. |
| Borgo et al. 2017 [27] | CS | 15 AN (BMI 13.9 ± 2.1 kg/m2) 15 HC (BMI 22.1 ± 2.6 kg/m2) N/A PSY | 16S rRNA qPCR | ↑ Proteobacteria ↑ Enterobacteriaceae ↑ M. smithii ↓ Firmicutes ↓ Ruminococcus ↓ Roseburia ↓ Clostridium ↓ Ruminococcaceae | ↔ alpha diversity ↔ beta diversity | ↓ total SCFA ↓ butyrate ↓ propionate ↔ iso-valerate ↔ iso-butyrate | All 11 patients with depression (assessed by BDI) were included in the AN patient sample. All 15 AN patients had an STAI-trait score > 40, 2 of HC. 7 AN patients had an STAI-state score > 40, 1 of HC. Negative correlation between BDI depression score and Clostridium spp. Negative correlation between faecal butyrate concentration and depression and anxiety scores. | N/A | Negative correlation between Bacteroides uniformis and BMI. Positive correlation between insulin and Roseburia inulinivorans. |
| Fan et al. 2023 [28] | CS, RCT (mice studies) | 77 AN (BMI 15.6 ± 2.5 kg/m2) 70 HC (BMI 21.8 ± 1.9 kg/m2) N/A PSY | Shotgun metagenomics | ↑ Christensenellales ↑ Clostridium paraputrificum ↑ Lactobacillus ↑ Ruminococcaceae-enterotype ↓ Bacteroidota ↓ Actinobacteriota ↓ Roseburia intestinalis, inulinivorans | ↑ beta diversity ↔ richness | ↑ indoxyl sulphate | N/A | Day 21 germ-free mice with FMT from AN showed a larger initial decrease in body weight and slower weight gain over time (compared with HC FMT mice) | Positive correlation between eating disorder scores and Clostridium spp. Negative correlation between eating disorder scores and Lactococcus acidophilus and Faecalibacterium prausnitzii. |
| Fouladi et al. 2022 [29] | L | 93 AN (BMI 14.6 ± 2.12 kg/m2) 98 HC (BMI 21.98 ± 2.13 kg/m2, BMI 22.56 ± 1.60 kg/m2) N/A PSY | Shotgun metagenomics | ↓ Bifidobacterium adolescentis ↓ Faecalibacterium prausnitzii | ↓ alpha diversity | N/A | N/A | Post-weight-restoration patients (compared to before): ↑ alpha diversity | Increase in the relative abundance of fermentation pathways in AN compared to HC. |
| Hanachi et al. 2019 [30] | CS | 33 AN (BMI 11.7 ± 1.5 kg/m2) 22 HC (BMI 21 ± 2 kg/m2) N/A PSY | 16S rRNA | ↑ Turicibacter ↑ Anaerotruncus ↑ Ruminococcus ↑ Salmonella ↑ Klebsiella ↓ Eubacterium ↓ Roseburia | ↓ alpha diversity ↓ richness | N/A | N/A | N/A | Negative correlation between BMI and Verrucomicrobiaceae and Ruminococcacea. Positive correlation between BMI and Clostridiales, Turicibacteraceae, and Eubacteriaceae. |
| Kleiman et al. 2015 [31] | L | 15 AN (BMI 16.2 ± 1.5 kg/m2) 14 HC (BMI 21.5 ± 1.9 kg/m2) N/A PSY | 16S rRNA | ↑ Bacilli ↑ Coriobacteriales ↓ Clostridia ↓ Anaerostipes ↓ Faecalibacterium | ↓ alpha diversity ↓ richness | N/A | 12 AN patients had at least mild depression (BDI), and 10 AN patients had at least mild anxiety (BAI). Greater levels of depression were negatively associated with alpha diversity. | Post-weight-restoration patients (compared to before), ↔ alpha diversity (compared to HCs): ↓ alpha diversity ↓ richness | Negative correlation between alpha diversity and levels of depression, eating disorder psychopathology. |
| Mack et al. 2016 [32] | L | 55 AN (BMI 15.3 ± 1.4 kg/m2) 55 HC (BMI 21.6 ± 2.0 kg/m2) N/A PSY | 16S rRNA | ↑ Firmicutes ↑ Actinobacteria ↑ Verrucomicrobia (mucin-degraders) ↑ Clostridium clusters I, XI, XVIII ↑ Methanobrevibacter ↓ Bacteroidetes ↓ Roseburia | ↑ beta diversity ↔ alpha diversity ↔ richness | ↑ Branched-chain fatty acids (isobutyrate and isovalerate) ↓ butyrate ↔ total SCFA | N/A | Post-weight-restoration patients (compared to before): ↑ beta diversity ↑ Firmicutes ↑ richness ↓ Verrucomicrobia ↓ Bacteroidetes | N/A |
| Million et al. 2013 [33] | CS | 15 AN (BMI 13.5 kg/m2) 76 HC (BMI 22.4 kg/m2) N/A PSY | qPCR | ↑ Escherichia coli ↑ M. smithii ↓ Lactobacillus reuteri, ↓ Lactobacillus plantarum | N/A | N/A | N/A | N/A | Negative correlation between BMI and Methanobrevibacter smithii, Escherichia coli, Bifidobacterium animalis. Positive correlation between BMI and Lactobacillus reuteri. |
| Monteleone et al. 2021 [34] | L | 21 AN (BMI 14.6 ± 1.3 kg/m2) 20 AN (BMI 20.3 ± 1.4 kg/m2) N/A PSY | 16S rRNA | ↑ Bacteroidetes ↑ Actinobacteria ↑ Weissella ↑ Coprococcus ↑ Bacteroidetes-to-Firmicutes abundance ratio ↓ Firmicutes ↓ Coriobacteriales ↓ Oxalobacteraceae ↓ Parabacterioides | ↓ alpha diversity ↔ beta diversity | ↓ sugars-derived metabolites | AN patients had significantly higher BSI scores at study entry compared to at discharge. AN at study entry mean a BSI global severity index of 22.1, compared to 12 at discharge. Positive correlation with BSI scores and negative with EDE scores: Coprococcus, Clostridium IV, Roseburia, Termsporobacter. | Post-weight-restoration patients (compared to HCs): ↓ alpha diversity (↑ compared to patients before weight restoration) ↑ Leuconostocaceae ↓ Actinobacteria ↓ Coriobacteriales ↓ Catabacteriaceae ↓ Collinsella ↓ Parabacteirodes ↓ Catabacter | Positive correlation between EDE scores and Bifidobacterium and Enterococcus. Negative correlation between BMI and Clostridium XVIII. |
| Morita et al. 2015 [35] | CS | 25 AN (BMI 12.8 ± 1.3 kg/m2) 21 HC (BMI 20.5 ± 2.1 kg/m2) N/A PSY | 16S or 23S rRNA -qPCR | ↓ Clostridium coccoides ↓ Clostridium leptum ↓ Bacteroides fragilis ↓ Streptococcus ↓ Lactobacillus plantarum | ↓ total abundance | ↓ acetate ↓ propionate | N/A | N/A | N/A |
| Morkl et al. 2017 [36] | CS | 18 AN (BMI 15.29 ± 1.28 kg/m2) 26 HC (BMI 21.89 ± 1.73 kg/m2) N/A PSY | 16S rRNA | ↑ Coriobacteriaceae | ↓ alpha diversity ↓ richness | N/A | AN patients had a mean BDI score of 21.72 and a HAMD score of 18.22. HCs had a mean BDI of 3.19, and HAMD of 2.65. Negative correlation between alpha diversity and levels of depression. | N/A | N/A |
| Prochazkova et al. 2021 [37] | CS | 51 AN (BMI 14.4 kg/m2) 67 HC (BMI 21.9 kg/m2) N/A PSY | 16S rRNA | ↑ Alistipes ↑ Clostridiales ↑ Christensenellaceae ↑ Ruminococcaceae ↓ Faecalibacterium, ↓ Agathobacter, ↓ Bacteroides, ↓ Blautia, ↓ Lachnospira. | ↑ beta diversity ↔ alpha diversity | ↓ butyrate ↓ acetate | N/A | Post-weight-restoration patients: ↔ gut microbiome composition ↔ beta diversity ↔ SCFA levels | Alpha diversity was not associated with BMI and EDE-Q score changes. |
| Schulz et al. 2021 [38] | L | 19 AN (BMI 15.76 ± 2.03 kg/m2) 20 HC (BMI 20.31 ± 2.35 kg/m2) N/A PSY | 16S rRNA | ↑ Anaerostipes ↓ Romboutsia | ↔ alpha diversity | N/A | AN patients at admission had a BDI 2 mean score of 22.68, compared to at discharge mean of 17.0, compared to the HCs’ mean of 5.65. | Post-weight-restoration patients (compared to HCs): ↑ Firmicutes ↑ Lachnospiraceae ↑ Fusicatenibacter ↓ Romboutsia | A higher abundance of unclassified Lachnospiraceae in AN patients at admission is associated with a shorter duration of treatment. |
| Yuan et al. 2022 [39] | CS | 30 AN (BMI 14.92 ± 2.54 kg/m2) 30 HC (BMI 20.89 ± 2.14 kg/m2) N/A PSY | 16S rRNA | ↑ Lachnospiraceae ↑ Eubacterium hallii ↓ Ruminococcaceae ↓ Faecalibacterium ↓ Subdoligranulum | ↑ beta diversity ↔ alpha diversity | N/A | AN patients had an HAMD mean score of 5, compared to the HCs’ mean of 2. | N/A | Negative correlation between BMI and Bacteroidota, Bacteroides, and Proteobacteria. Positive correlation between BMI and Subdoligranulum, Firmicutes. |
| Armougom | Borgo | Fan | Fouladi | Hanachi | Kleiman | Mack | Million | Monteleone | Morita | Morkl | Prochazkova | Schulz | Yuan | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phylum | ||||||||||||||
| Bacterioides | ↔ | ↓ | ↓ | ↑ | ||||||||||
| Firmicutes | ↔ | ↓ | ↑ | ↓ | ||||||||||
| Actinobacteria | ↓ | ↑ | ↑ | |||||||||||
| Verrucomicrobia | ↑ | |||||||||||||
| Proteobacteria | ↑ | |||||||||||||
| Family | ||||||||||||||
| Coriobacteriaceae | ↑ | ↓ | ↑ | |||||||||||
| Ruminococcaceae | ↓ | ↑ | ↓ | |||||||||||
| Enterobacteriaceae | ↑ | |||||||||||||
| Peptostreptococcaceae | ||||||||||||||
| Oxalobacteraceae | ↓ | |||||||||||||
| Genus | ||||||||||||||
| Ruminococcus | ↓ | ↑ | ↑ | |||||||||||
| Roseburia | ↓ | ↓ | ↓ | ↓ | ||||||||||
| Clostridium | ↓ | ↑ | ↓ | ↑ | ↓ | ↑ | ||||||||
| Bacterioides fragilis | ↓ | |||||||||||||
| Streptococcus | ↓ | |||||||||||||
| Lactobacillus | ↔ | ↑ | ↓ | ↓ | ||||||||||
| Eubacterium | ↓ | ↑ | ||||||||||||
| Anaerostipes | ↓ | ↑ | ||||||||||||
| Turicibacter | ↑ | |||||||||||||
| Anaerotruncus | ↑ | |||||||||||||
| Salmonella | ↑ | |||||||||||||
| Klebsiella | ↑ | |||||||||||||
| Methanobrevibacter | ↑ | ↑ | ↑ | ↑ | ||||||||||
| Gemmiger | ||||||||||||||
| Bifidobacterium | ↓ | |||||||||||||
| Faecalibacterium | ↓ | ↓ | ↓ | ↓ | ||||||||||
| Christensenella | ↑ | ↑ | ||||||||||||
| Escherichia | ↑ | |||||||||||||
| Weissella | ↑ | |||||||||||||
| Coprococcus | ↑ | |||||||||||||
| Parabacterioides | ↓ | |||||||||||||
| Alistipes | ↑ | |||||||||||||
| Agathobacter | ↓ | |||||||||||||
| Lachnospira | ↓ | ↑ | ||||||||||||
| Romboutsia | ↓ | |||||||||||||
| Bacterial metabolites | ||||||||||||||
| Acetate | ↓ | ↓ | ||||||||||||
| Butyrate | ↓ | ↓ | ↓ | |||||||||||
| Propionate | ↓ | ↓ | ||||||||||||
| Iso-Valerate | ↔ | ↑ | ||||||||||||
| Iso-butyrate | ↔ | ↑ |
| References | Significant Changes in Gut Microbiome Composition among AN Patients with Psychiatric Comorbidities (Depression or Anxiety) |
|---|---|
| Borgo | Negative correlation between BDI depression score and Clostridium spp. Negative correlation between faecal butyrate concentration and depression and anxiety scores. |
| Kleiman | Greater levels of depression were negatively associated with alpha diversity. |
| Monteleone | Positive correlation with BSI scores: Coprococcus, Clostridium IV, Roseburia, Termsporobacter. |
| Morkl | Negative correlation between alpha diversity and levels of depression. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Kodancha, P.; Das, S. Gut Microbiome Changes in Anorexia Nervosa: A Comprehensive Review. Pathophysiology 2024, 31, 68-88. https://doi.org/10.3390/pathophysiology31010006
Zhao W, Kodancha P, Das S. Gut Microbiome Changes in Anorexia Nervosa: A Comprehensive Review. Pathophysiology. 2024; 31(1):68-88. https://doi.org/10.3390/pathophysiology31010006
Chicago/Turabian StyleZhao, Wendi, Prabhath Kodancha, and Soumitra Das. 2024. "Gut Microbiome Changes in Anorexia Nervosa: A Comprehensive Review" Pathophysiology 31, no. 1: 68-88. https://doi.org/10.3390/pathophysiology31010006
APA StyleZhao, W., Kodancha, P., & Das, S. (2024). Gut Microbiome Changes in Anorexia Nervosa: A Comprehensive Review. Pathophysiology, 31(1), 68-88. https://doi.org/10.3390/pathophysiology31010006