Moringa oleifera Extract Extenuates Echis ocellatus Venom-Induced Toxicities, Histopathological Impairments and Inflammation via Enhancement of Nrf2 Expression in Rats

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Snake Venom and Antivenom
2.2. Ethical Consideration
2.2.1. Experimental Animals
2.2.2. Evaluation of Lethality Dose (LD50) of E. ocellatus Venom
2.3. Collection, Identification and Extraction Procedure of Plant Materials
2.4. Anti-Ophidic Experiment
Experimental Design, Envenoming and Treatment Procedures
2.5. Measurement of Haematological Indices
2.6. Blood Chemistry Analysis
2.7. Antioxidant Enzyme Profile
2.8. Measurement of Inflammatory Cytokines in Serum and Heart Tissues
2.8.1. Tissue Preparation
2.8.2. Measurement of Pro-Inflammatory Cytokines and NRF2 Levels in Serum and Heart Tissue
2.9. Anti-Immunoglobulin G Assay
2.10. In Vitro Assays
2.10.1. Anti-Hemorrhagic Test
2.10.2. Anti-Hemolytic Assay
2.10.3. Anticoagulant Assay
2.11. Histopathological Studies
3. Statistical Analysis
4. Results
4.1. Lethal Dose Test of E. ocellatusVenom
4.2. Mortality and Behavioural Responses of Experimental Rats
4.3. Effects of Treatment with M. oleifera Extract on Haematological Parameters of Envenomed Rats
White Blood Cell and Differentials of the Envenomed Treated Rats
4.4. Effect of Treatment with M. oleifera Extract on Blood Electrolytes of Envenomed Treated Rats
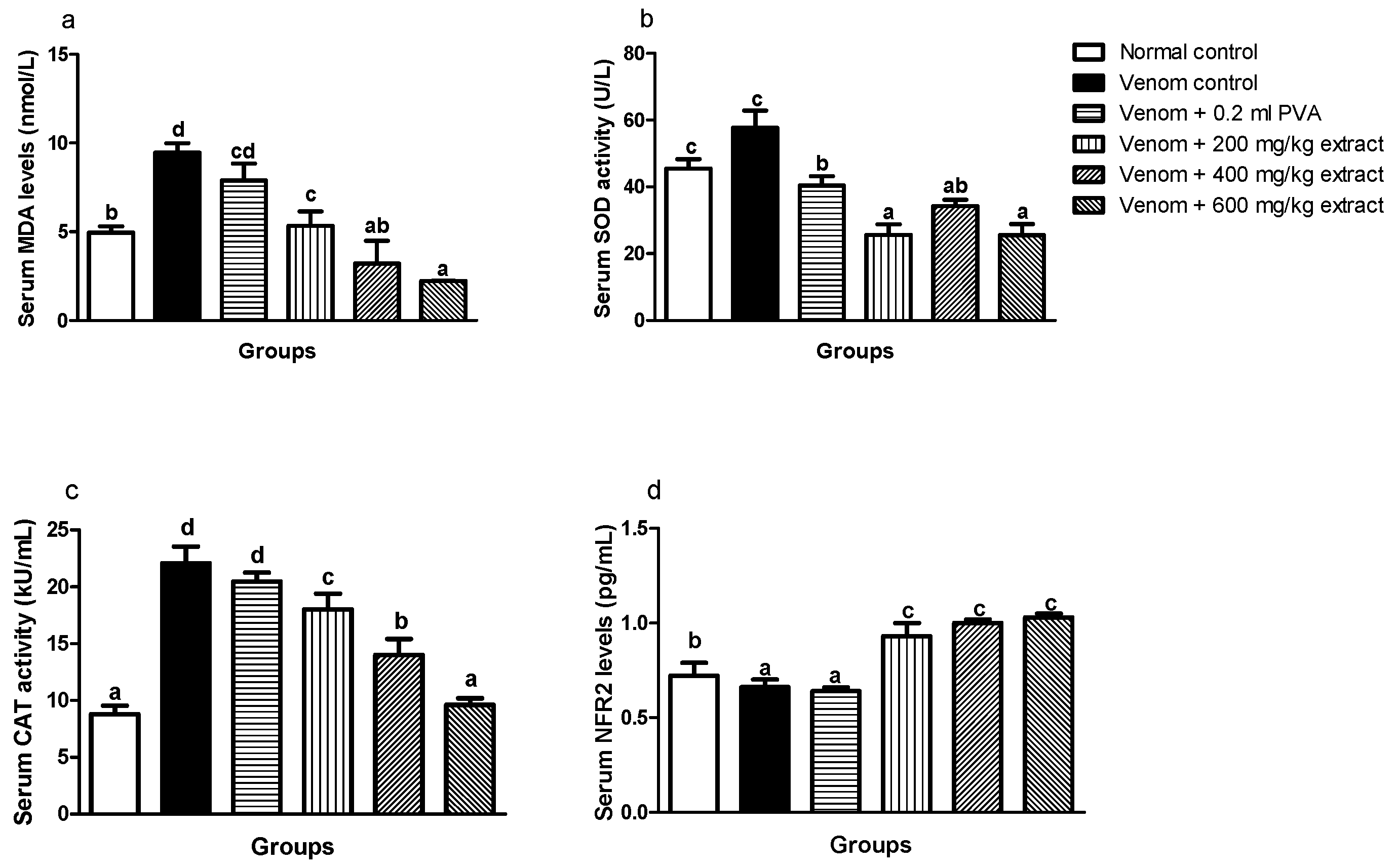
4.5. Effects of M. oleifera Extract on Serum and Heart Oxidative Stress Index, Antioxidant Enzymes Activity and Nrf2 Expression of Envenomed Treated Rats
4.6. Effects of M. oleiferaExtract on Serum and Heart Tumor Necrosis Factor-Alpha (TNF-α) and Interleukin 1–Beta (IL1-β) Expression in Envenomed Treated Rats
4.7. IgG Antibody Titre of Envenomed Rats Treated with M. oleiferaExtract
4.8. In Vitro Studies
4.8.1. Anti-Hemorrhagic Activity of M. oleifera Extract
4.8.2. Anticoagulant Studies
4.8.3. Antihemolytic Activity of M. oleifera Leave Extract
4.9. Histological Studies
4.9.1. Histopathological Studies of the Brain
4.9.2. Histopathological Studies of the Heart
4.9.3. Histopathological Studies of the Kidney
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gutierrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A. The Global Burden of Snakebite, A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths. PLoS Med. 2017, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- The Lancet. Snake-bite envenoming, A priority neglected tropical disease. Lancet 2017, 390, 2. [Google Scholar] [CrossRef]
- Chippaux, J.P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef] [PubMed]
- Onyeama, H.P.; Ebong, P.E.; Eteng, M.U.; Igile, G.O.; Ofemile, P.Y.; Ibekwe, H.A. Histological Reponses of the heart, liver and kidney to Calliandra portoricensis extracts in Wistar rats challenged with venom of Echis ocellatus. J. Appl. Pharm. Sci. 2012, 2, 164–171. [Google Scholar]
- Warrell, D.A.; Arnett, C. The importance of bites by the saw-scaled or carpet viper (Echiscarinatus). Epidemiological studies in Nigeria and a review of the world literature. Acta Trop. 1976, 33, 307–341. [Google Scholar] [PubMed]
- Wever, E.G. The Reptile Ear; Princeton University Press: Princeton, NJ, USA, 2019; Volume 5346. [Google Scholar]
- Wagstaff, S.; Harrison, R. Venom gland EST analysis of the saw-scaled viper, Echis ocellatus, reveals novel α β integrin-binding motifs in venom metalloproteinases and a new group of putative toxins, renin-like aspartic proteases. Gene 2006, 377, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Hasson, S.S.; Al-Jabri, A.A.; Sallam, T.A.; Al-Balushi, M.S.; Mothana, R.A.A. Antisnake venom activity of Hibiscus aethiopicus L. against Echis ocellatus and Naja n. nigricollis. J. Toxicol. 2010, 2010, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chaves, F.; Teixeira, C.F.P.; Gutiérrez, J.M. Role of TNF-α, IL-1β and IL-6 in the local tissue damage induced by Bothrops asper snake venom, An experimental assessment in mice. Toxicon 2005, 45, 171–178. [Google Scholar] [CrossRef]
- Zamuner, S.R.; Zuliani, J.P.; Fernandes, C.M.; Gutiérrez, J.M.; Teixeira, C.F.P. Inflammation induced by Bothrops asper venom, Release of proinflammatory cytokines and eicosanoids, and role of adhesion molecules in leukocyte infiltration. Toxicon 2005, 46, 806–813. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Dkhil, M.A.; Moneim, A.E.A. Hepatotoxicity and oxidative stress induced by Najahaje crude venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 42. [Google Scholar] [CrossRef]
- Rushmore, T.H.; Morton, M.R.; Pickett, C.B. The antioxidant responsive element. Activation by oxidative stress and identification of the DNA consensus sequence required for functional activity. J. Biol. Chem. 1991, 266, 11632–11639. [Google Scholar] [CrossRef]
- Onyeama, H.P.; Igile, G.O.; Mgbeje, B.I.; Eteng, M.U.; Ebong, P.E. Evaluation of the biochemical and anti-snake venom effects of Calliandra portoricensis extract fractions in wistar rat models challenged with venom of carpet viper (Echis ocellatus). Biochemistry 2013, 3, 441–449. [Google Scholar]
- Zailani, A.H.; Magaji, P.K.; Sarkiyayi, S.; Wurochekke, A.U. Anti-snake Venom Activity of Aqueous and Ethanolic Extracts of Crinum jagus Bulb. Asian J. Res. Biochem. 2018, 2, 1–9. [Google Scholar] [CrossRef]
- Gutierrez, J.M. Clinical Toxicology of Snakebite in Central America. In Handbook of Clinical Toxicology of Animal Venoms and Poison; Meier, J.W., Ed.; CRC Press: Florida, FL, USA, 1995; pp. 645–665. [Google Scholar]
- de Silva, H.A.; Ryan, N.M.; de Silva, H.J. Adverse reactions to snake anti-venom, and their prevention and treatment. Br. J. Clin. Pharmacol. 2016, 81, 446–452. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Leon, G.; Rojas, G.; Lomonte, B.; Rucavado, A.; Chaves, T. Neutralization of local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon 1998, 36, 1529–1538. [Google Scholar] [CrossRef]
- Adeyi, A.O.; Ajisebiola, B.S.; Adeyi, O.E.; Alimba, G.C.; Okorie, U.G. Anti-venom activity of Moringa oleifera leave against pathophysiological alterations, somatic mutation and biological activities of Naja nigricollis venom. Sci. Afr. 2020, 8, e00356. [Google Scholar]
- Ajisebiola, B.S.; Solomon, R.; Anwar, U.; Adeyi, A.O. Neutralization of Bitis arietans venom-induced pathophysiological disorder, biological activities and genetic alterations by Moringa oleifera leaves. Toxin Rev. 2020. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Spearman-Karber, P. Alternative Methods of Analysis for Quantal Responses. In Statistical Method in Biological Assay; Finney and Griffin: London, UK, 1978; pp. 1–89. [Google Scholar]
- World Health Organization. WHO Monographs on the Selected Medicinal Plants; WHO: Geneva, Switzerland, 1998. [Google Scholar]
- Rowett, H.G.O. Dissecting Guides of Rats with Notes on Mouse. In 111, Bulter and Tanner; LTD: London, UK, 1997; pp. 5–23. [Google Scholar]
- Baker, F.T.; Silverton, R.E. Introduction to Medical Laboratory Technology; Butterworth: London, UK, 1985; p. 408. ISBN 0407732527. [Google Scholar]
- Terri, A.E.; Sesin, P.G. Potassium concentration determination by Teco diagnostics reagent kits. Am. J. Clin. Pathol. 1958, 61, 544. [Google Scholar]
- Henry, R.J. Sodium Concentration Determination. In CE Human Reagents Kits; Clinical Chemistry Harper & Row New York, Ed.; Clinical Chemistry Harper & Row New York: New York, NY, USA, 1974; p. 643. [Google Scholar]
- Schonfeld, R.G.; Lowellen, C.S. Agappe Diagnostic Kits. Clin. Chem. 1994, 10, 533. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Satoh, K. Serum lipid peroxide in cerebrovascular disorders determined by a new colorimetric method. Clin. Chim. Acta 1978, 90, 37–43. [Google Scholar]
- Yacoub, T.; Rima, M.; Sadek, R.; Hleihel, W.; Fajloun, Z.; Karam, M. Montivipera bornmuelleri venom has immunomodulatory effects mainly up-regulating pro-inflammatory cytokines in the spleens of mice. Toxicol. Rep. 2018, 5, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Theakston, R.D.G.; Reid, H.A. Development of simple standard assay procedures for the characterisation of snake venoms. Bull. WHO 1983, 61, 949–956. [Google Scholar] [PubMed]
- Gomes, A.; Pallabi, D.; Hannahpep, A. Novel Fibrinolytic Peptide from the Indian King Cobra (Ophiophagus hannah) Venom. Biochem. Biophys. Res. Commun. 1999, 266, 488–491. [Google Scholar] [CrossRef]
- Carleton, H.M.; Drury, R.A.B.; Wallington, E.A. Caleton’s Histological Techniques, 5th ed.; Oxford University Press: New York, NY, USA, 1980; pp. 147–148. [Google Scholar]
- Calvete, J.J. Anti-venomics and venom phenotyping, A marriage of convenience to address the performance and range of clinical use of anti-venoms. Toxicon 2010, 56, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M. Timeline of key events in snake venom metalloproteinase research. J. Proteom. 2009, 72, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Escalante, T.; Rucavado, A.; Fox, J.W.; Gutiérrez, J.M. Key events in microvascular damage induced by snake venom hemorrhagic metalloproteinases. J. Proteom. 2011, 74, 1781–1794. [Google Scholar] [CrossRef]
- Kini, R.M. The intriguing world of prothrombin activators from snake venom. Toxicon 2005, 45, 1133–1145. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Chaves, F.; Bolano, R. Estudio comparativo de venenos de ejemplares recien nacidos y adultos de Bothrops asper. Rev. Biol. Trop. 2010, 28, 341–351. [Google Scholar]
- Moura-da-Silva, A.M.; Serrano, S.M.T.; Fox, J.W.; Gutiérrez, J.M. Snake venom metalloproteinases. Structure, function and effects on snake bite pathology. In Animal Toxins, State of the Art. Perspectives in Health and Biotechnology; de Lima, M.E., Pimenta, A.M.C., Martin-Euclaire, M.F., Zingali, M.F., Rochat, H., Eds.; UFM: Belo Horizonte, Brazil, 2009; pp. 525–546. ISBN 978-85-7041-735-0. [Google Scholar]
- Onyeama, H.P.; Ebong, P.E.; Eteng, M.U.; Igile, G.O.; Ibekwe, H.A.; Atangwho, I.J. Effects of Calliandra Portoricensis Extracts on the Haematological Indices of Wistar Rats Challenged with Venom of Echis ocellatus. J. Appl. Pharm. Sci. 2012, 2, 140–144. [Google Scholar]
- Munim, A.; Puteri, M.U.; Purna, S.S.; Azizahwati, A. Anti-Anemia Effect of Standardized Extract of Moringa oleifera Lam. Leaves on Aniline Induced Rats. Pharmacogn. Res. 2016, 8, 255–258. [Google Scholar]
- Meier, J.; Stocker, K. Effect of snake venoms on homeostasis. Toxicology 1991, 21, 171–1820. [Google Scholar]
- Zuliani, J.P.; Soares, A.M.; Gutiérrez, J.M. Polymorphonuclear neutrophil leukocytes in snakebite envenoming. Toxicon 2020, 18, 188–197. [Google Scholar] [CrossRef]
- El Shanawany, E.E.; Fouad, E.A.; Keshta, H.G.; Hassan, S.E.; Hegazi, A.G.; Abdel-Rahman, E.H. Immunomodulatory effects of Moringa oleifera leaves aqueous extract in sheep naturally co-infected with Fasciola gigantica and Clostridium novyi. J. Parasit. Dis. 2019, 43, 583–591. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Boeno, C.N.; Lopes, J.A.; da Rosa, A.E.D.S.; Pires, W.L.; Pontes, A.S.; Zuliani, J.P. Role of l-amino acid oxidase isolated from Calloselasma rhodostoma venom on neutrophil NADPH oxidase complex activation. Toxicon 2018, 145, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D. Mechanistic studies of the Nrf2-Keap1 signaling pathway. Drug Metab. Rev. 2006, 38, 769–789. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Chen, Q.; Tao, J. Astaxanthin Promotes Nrf2/ARE Signaling to Inhibit HG-Induced Renal Fibrosis in GMCs. Mar. Drugs 2018, 16, 117. [Google Scholar]
- Kangralkar, V.A.; Patil, S.D.; Bandivadekar, R.M. Oxidative stress and diabetes, A review. Intl. J. Pharm. Appl. 2010, 1, 38–45. [Google Scholar]
- Santhosh, M.S.; Sundaram, M.S.; Sunitha, K.; Kemparaju, K.; Girish, K.S. Viper venom-induced oxidative stress and activation of inflammatory cytokines:A therapeutic approach for overlooked issues of snakebite management. Inflamm. Res. 2013, 62, 721–731. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar]
- Hageman, J.J.; Bast, A.; Vermeulen, N.P. Monitoring of oxidative free radical damage in vivo: Analytical aspects. Chem. Biol. Interact. 1992, 82, 243–293. [Google Scholar] [CrossRef]
- Somade, O.T.; Ajayi, B.O.; Adeyi, O.E.; Adeshina, A.A.; James, A.S.; Ayodele, P.F. Ethylene glycol monomethyl ether-induced testicular oxidative stress and time-dependent up-regulation of apoptotic, pro-inflammatory, and oncogenic markers in rats. Metab. Open 2020, 7, 100051. [Google Scholar] [CrossRef] [PubMed]
- Cherian, D.A.; Peter, T.; Narayanan, A.; Madhavan, S.S.; Achammada, S.; Vynat, G.P. Malondialdehyde as a Marker of Oxidative Stress in Periodontitis Patients. J. Pharm. Bioallied Sci. 2019, 11 (Suppl. S2), S297–S300. [Google Scholar] [CrossRef] [PubMed]
- Suphachai, C. Antioxidant and anticancer activities of Moringa oleifera leaves. J. Med. Plant. Res. 2014, 8, 318–325. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesth. Clin. 2004, 45, 27–37. [Google Scholar] [CrossRef]
- Zuliani, J.P.; Fernandes, C.M.; Zamuner, S.R.; Gutie´rrez, J.M.; Teixeira, C.F. Inflammatory events induced by Lys-49 and Asp-49 phospholipases A2 isolated from Bothrops asper snake venom, Role of catalytic activity. Toxicon 2005, 45, 335–346. [Google Scholar] [CrossRef]
- Petricevich, V.L.; Teixeira, C.F.P.; Tambourgi, D.V.; Gutie´rrez, J.M. Increments in cytokine and nitric oxide serum levels in mice injected with Bothrops asper and Bothrops jararaca snake venoms. Toxicon 2000, 38, 1253–1266. [Google Scholar] [CrossRef]
- Kooltheat, N.; Sranujit, R.P.; Chumark, P.; Potup, P.; Laytragoon-Lewin, N.; Usuwanthim, K. An ethyl acetate fraction of Moringa oleifera Lam. inhibits human macrophage cytokine production induced by cigarette smoke. Nutrients 2014, 6, 697–710. [Google Scholar] [CrossRef]
- Mittal, A.; Sharma, M.; David, A.; Vishwakarma, P.; Saini, M.; Goel, M.; Saxena, K.K. An experimental study to evaluate the anti-inflammatory effect of Moringa oleifera leaves in animal models. Int. J. Basic Clin. Pharmacol. 2017, 6, 452. [Google Scholar] [CrossRef]
- Scott-Taylor, T.H.; Axinia, S.C.; Amin, S.; Pettengell, R. Immunoglobulin G.; structure and functional implications of different subclass modifications in initiation and resolution of allergy. Immun. Inflamm. Dis. 2018, 6, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Angulo, Y.; Calderón, L. An overview of lysine-49 phospholipase A2 myotoxins from crotalid snake venoms and their structural determinants of myotoxic action. Toxicon 2003, 42, 885–901. [Google Scholar] [CrossRef]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Sajevic, T.; Leonardi, A.; Križaj, I. Haemostatically active proteins in snake venoms. Toxicon 2011, 57, 627–645. [Google Scholar] [CrossRef]
- Bardaweel, S.K.; Gul, M.; Alzweiri, M.; Ishaqat, A.; AL Salamat, H.A.; Bashatwah, R.M. Reactive Oxygen Species: The Dual Role in Physiological and Pathological Conditions of the Human Body. Eurasian J. Med. 2018, 50, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Jimenez, M.; Almatrafi, M.M.; Fernandez, M.L. Bioactive Components in Moringa oleifera Leaves Protect against Chronic Disease. Antioxidants 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.I.; Gomes, A. Viper venom-induced inflammation and inhibition of free radical formation by pure compound (2-hydroxy-4-methoxy benzoic acid) isolated from anantamul (Hemidesmus indicus, R. BR) root extract. Toxicon 1998, 36, 207–215. [Google Scholar] [CrossRef]
- Chugh, K.S. Snake-bite-induced acute renal failure in India. Kidney Int. 1989, 35, 891–907. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | GROUP 1 | GROUP 2 | GROUP 3 | GROUP 4 | GROUP 5 | GROUP 6 |
|---|---|---|---|---|---|---|
| PCV (%) | 46.00 ± 2.00 e | 40.33 ± 0.58 d | 53.00 ± 0.00 b | 48.00 ± 1.00 a | 58.00 ± 1.00 c | 63.00 ± 2.00 b |
| Hb (g/dl) | 15.23 ± 0.82 b | 13.43 ± 0.06 c | 15.37 ± 0.15 b | 17.70 ± 0.20 a | 17.80 ± 0.10 a | 19.80 ± 0.10 b |
| RBC (cell/L) | 7.92 ± 0.35 b | 6.77 ± 0.08 c | 8.62 ± 0.08 b | 8.15 ± 0.02 b | 9.83 ± 0.01 a | 9.75 ± 0.05 a |
| PLATELET (×105 cell/L) | 1.31 ± 0.20 c | 1.44 ± 0.40 b | 1.72 ± 0.30 a | 1.57 ± 0.10 b | 1.29 ± 0.01 c | 1.44 ± 0.29 b |
| MCV (fl) | 58.1 ± 1.21 c | 59.5 ± 0.22 a | 61.5 ± 0.52 a | 58.9 ± 1.43 ab | 59.0 ± 1.12 b | 64.6 ± 1.81 a |
| MCH (pg/cell) | 19.2 ± 0.21 a | 19.8 ± 0.11 a | 20.2 ± 0.21 b | 19.3 ± 0.22 a | 18.1 ± 0.13 b | 20.3 ± 0.22 a |
| MCHC (g/dl) | 33.1 ± 0.70 a | 33.3 ± 0.62 a | 32.8 ± 0.32 ab | 32.7 ± 1.11 ab | 30.7 ± 0.40 c | 31.5 ± 1.11 b |
| Parameters | GROUP 1 | GROUP 2 | GROUP 3 | GROUP 4 | GROUP 5 | GROUP 6 |
|---|---|---|---|---|---|---|
| WBC (×103 cell/L) | 6.5 ± 0.46 a | 5.30 ± 0.10 c | 5.97 ± 0.33 ab | 6.7 ± 0.05 a | 6.0 ± 0.05 ab | 6.6 ± 1.00 a |
| LYM (%) | 76.0 ± 3.01 b | 65.0 ± 1.00 c | 75.0 ± 1.00 b | 64.3 ± 0.60 c | 75.0 ± 1.00 b | 83.0 ± 1.00 a |
| NEUT (%) | 32.3 ± 3.50 a | 23.0 ± 1.00 c | 22.0 ± 2.00 c | 30.0 ± 1.00 a | 28.0 ± 1.00 ab | 32.7 ± 0.61 a |
| MONO (%) | 1.7 ± 0.62 ab | 1.0 ± 0.00 c | 1.7 ± 0.60 ab | 3.0 ± 1.00 a | 3.0 ± 0.00 a | 2.3 ± 0.62 ab |
| EO (%) | 2.0 ± 1.00 b | 1.0 ± 0.00 c | 2.0 ± 1.00 b | 2.0 ± 0.00 b | 2.0 ± 0.00 b | 2.7 ± 0.60 a |
| Treatment Groups | Percentage Inhibition (%) |
|---|---|
| Group 1 | - |
| Group 2 | 0 (No inhibition) |
| Group 3 | 58.00 ± 8.00 c |
| Group 4 | 72.00 ± 7.00 b |
| Group 5 | 84.00 ± 4.00 ab |
| Group 6 | 95.00 ± 11.72 a |
| GROUPS | % Inhibition |
|---|---|
| 1 | 0 (No inhibition) |
| 2 | 28.00 ± 4.00 c |
| 3 | 80.00 ± 10.00 b |
| 4 | 65.00 ± 5.00 ab |
| 5 | 71.00 ± 1.00 b |
| 6 | 87.00 ± 7.00 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeyi, A.O.; Adeyemi, S.O.; Effiong, E.-O.P.; Ajisebiola, B.S.; Adeyi, O.E.; James, A.S. Moringa oleifera Extract Extenuates Echis ocellatus Venom-Induced Toxicities, Histopathological Impairments and Inflammation via Enhancement of Nrf2 Expression in Rats. Pathophysiology 2021, 28, 98-115. https://doi.org/10.3390/pathophysiology28010009
Adeyi AO, Adeyemi SO, Effiong E-OP, Ajisebiola BS, Adeyi OE, James AS. Moringa oleifera Extract Extenuates Echis ocellatus Venom-Induced Toxicities, Histopathological Impairments and Inflammation via Enhancement of Nrf2 Expression in Rats. Pathophysiology. 2021; 28(1):98-115. https://doi.org/10.3390/pathophysiology28010009
Chicago/Turabian StyleAdeyi, Akindele O., Sodiq O. Adeyemi, Enoh-Obong P. Effiong, Babafemi S. Ajisebiola, Olubisi E. Adeyi, and Adewale S. James. 2021. "Moringa oleifera Extract Extenuates Echis ocellatus Venom-Induced Toxicities, Histopathological Impairments and Inflammation via Enhancement of Nrf2 Expression in Rats" Pathophysiology 28, no. 1: 98-115. https://doi.org/10.3390/pathophysiology28010009
APA StyleAdeyi, A. O., Adeyemi, S. O., Effiong, E.-O. P., Ajisebiola, B. S., Adeyi, O. E., & James, A. S. (2021). Moringa oleifera Extract Extenuates Echis ocellatus Venom-Induced Toxicities, Histopathological Impairments and Inflammation via Enhancement of Nrf2 Expression in Rats. Pathophysiology, 28(1), 98-115. https://doi.org/10.3390/pathophysiology28010009