
Salinity Tolerance and Ion Accumulation of Coastal and Inland Accessions of Clonal Climbing Plant Species Calystegia sepium in Comparison with a Coastal-Specific Clonal Species Calystegia soldanella



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Establishment of Stock Cultures
2.3. Plant Propagation for Experiments and Cultivation
2.4. Plant Treatment and Harvesting
2.5. Measurements
2.6. Data Analysis
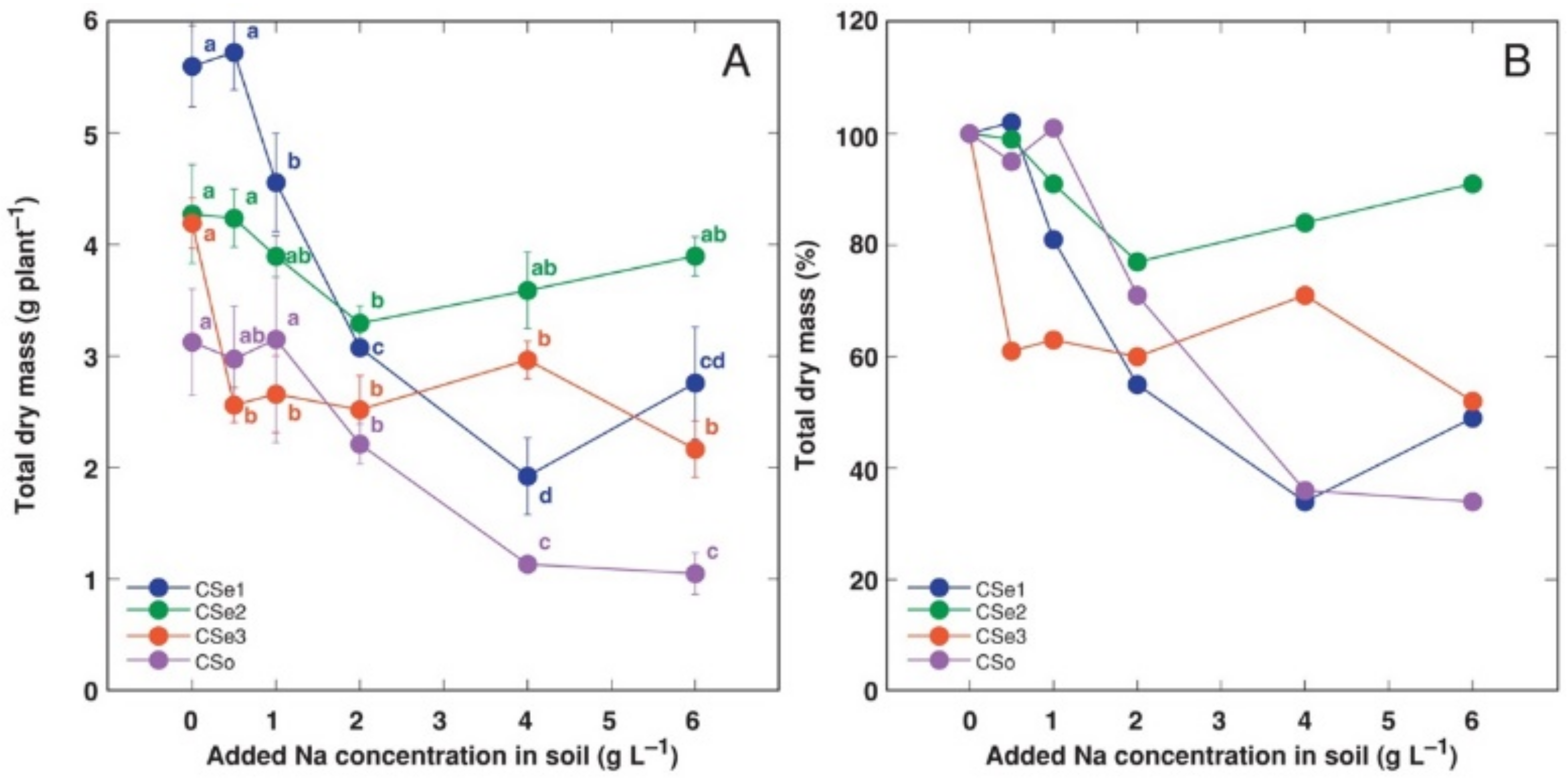
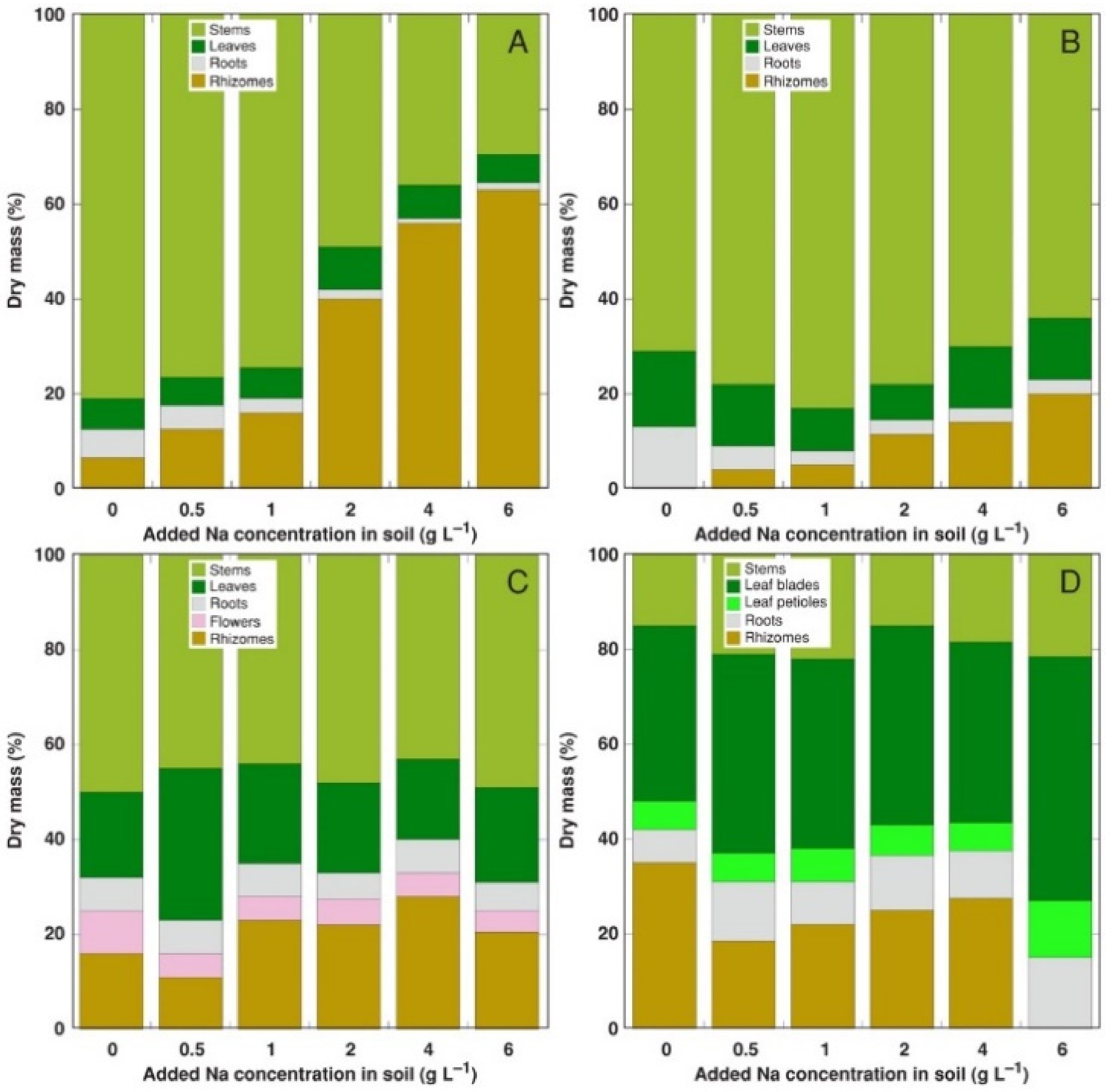
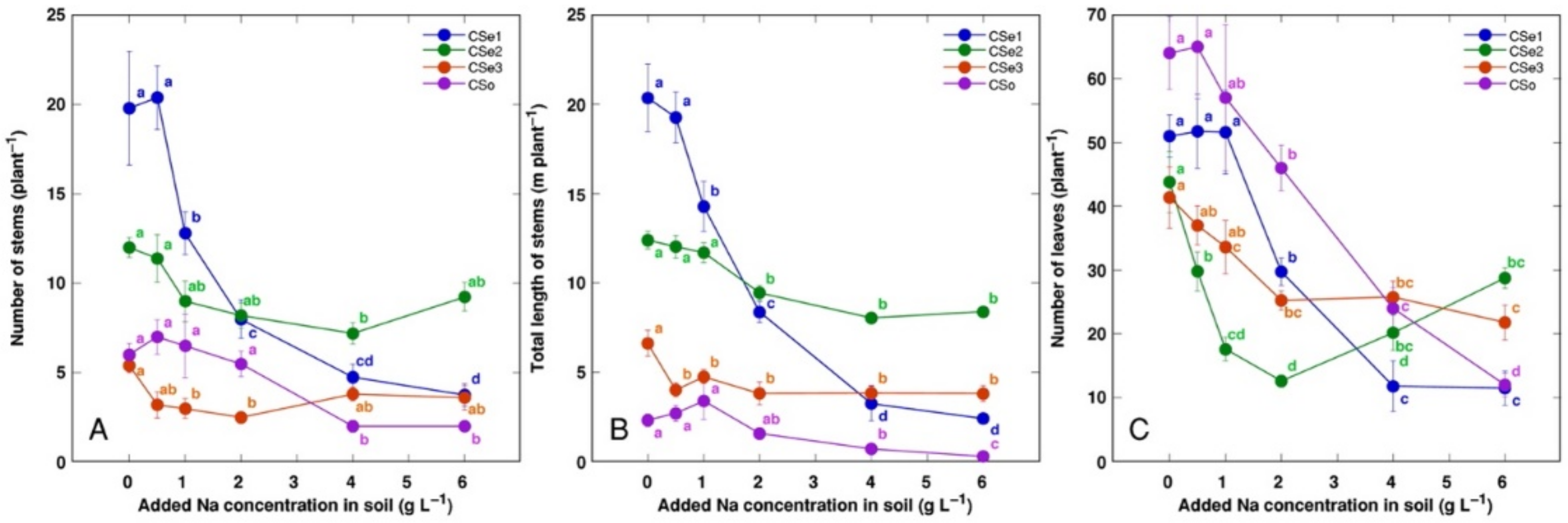
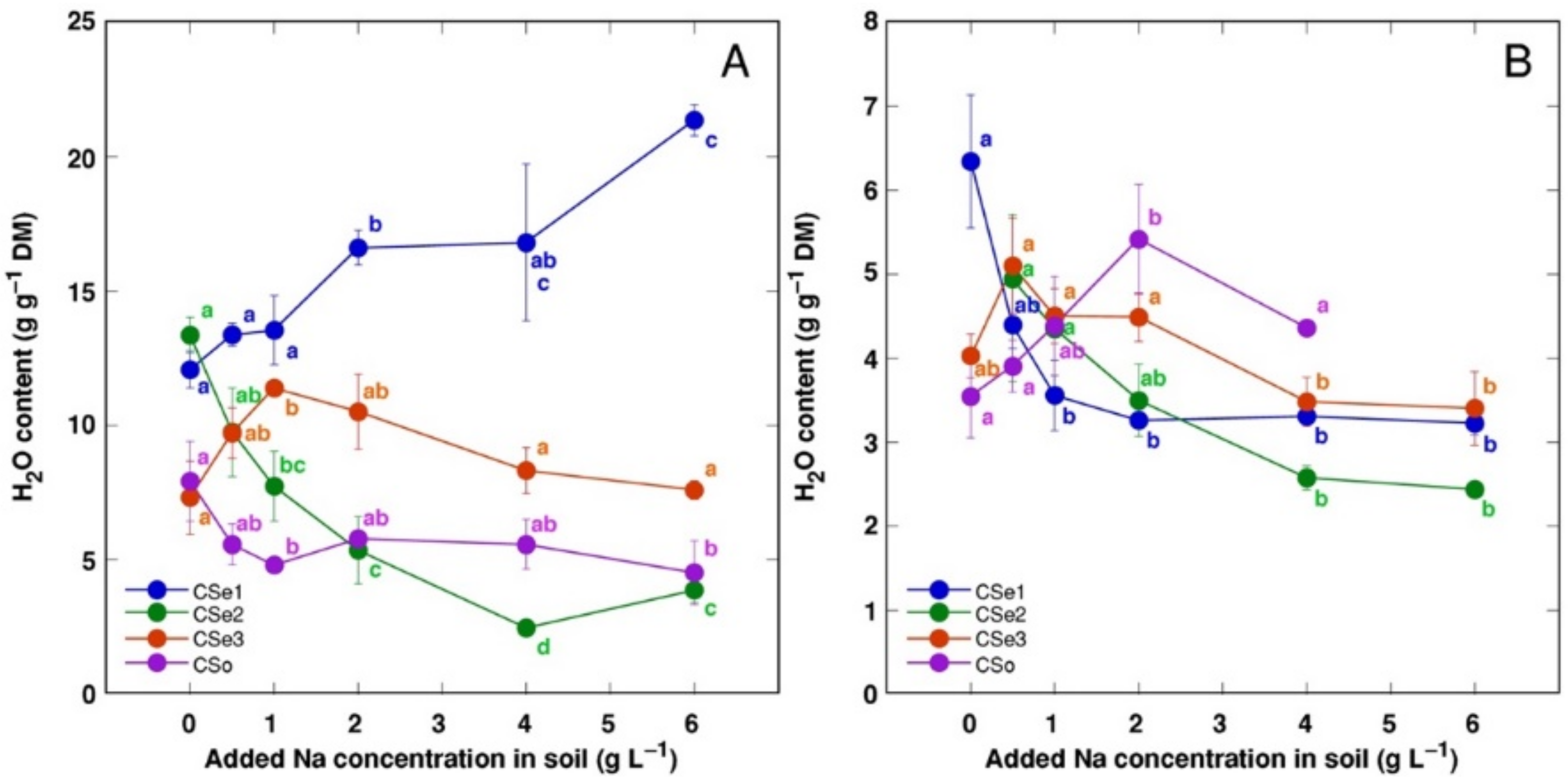
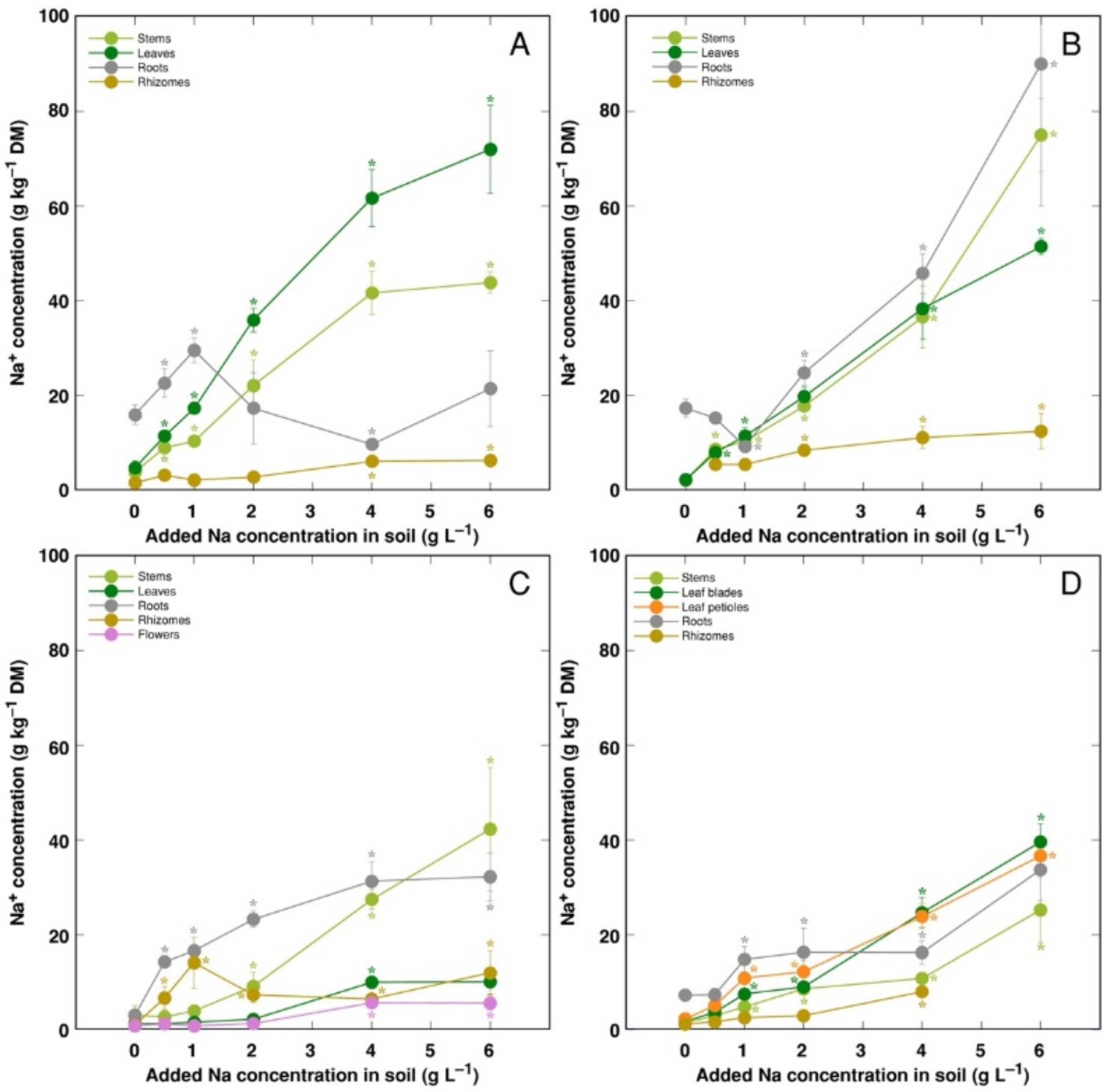
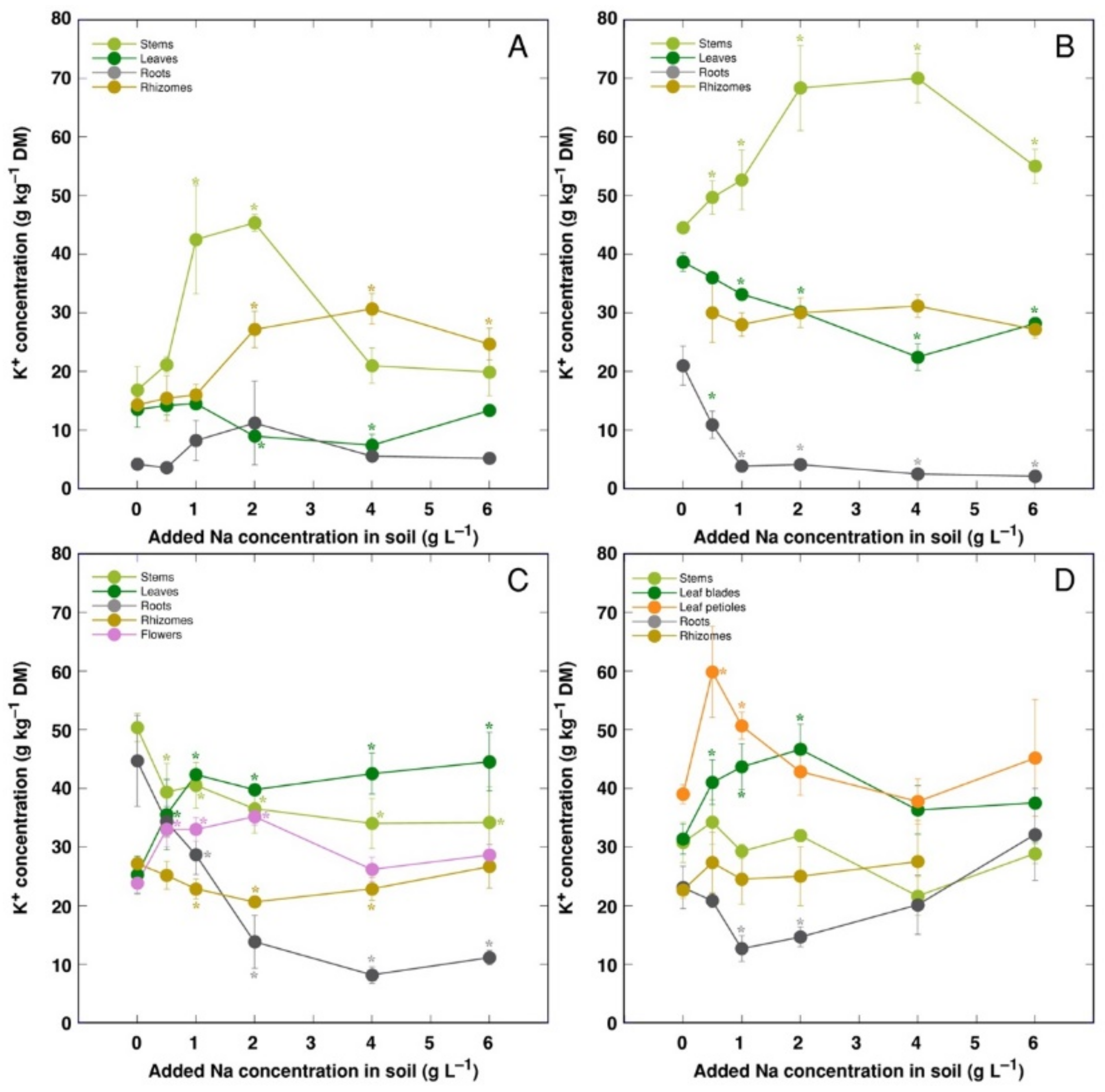
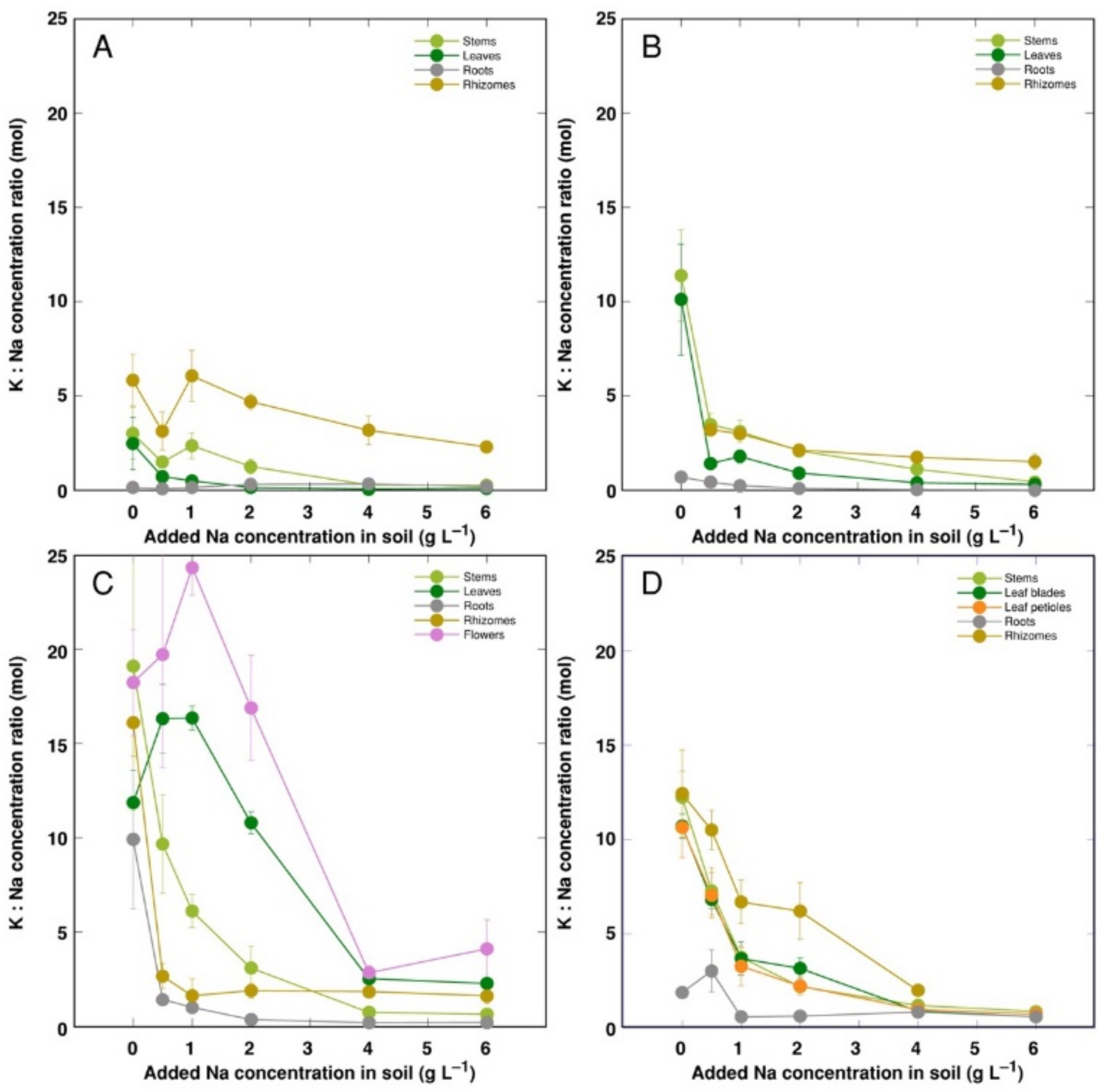
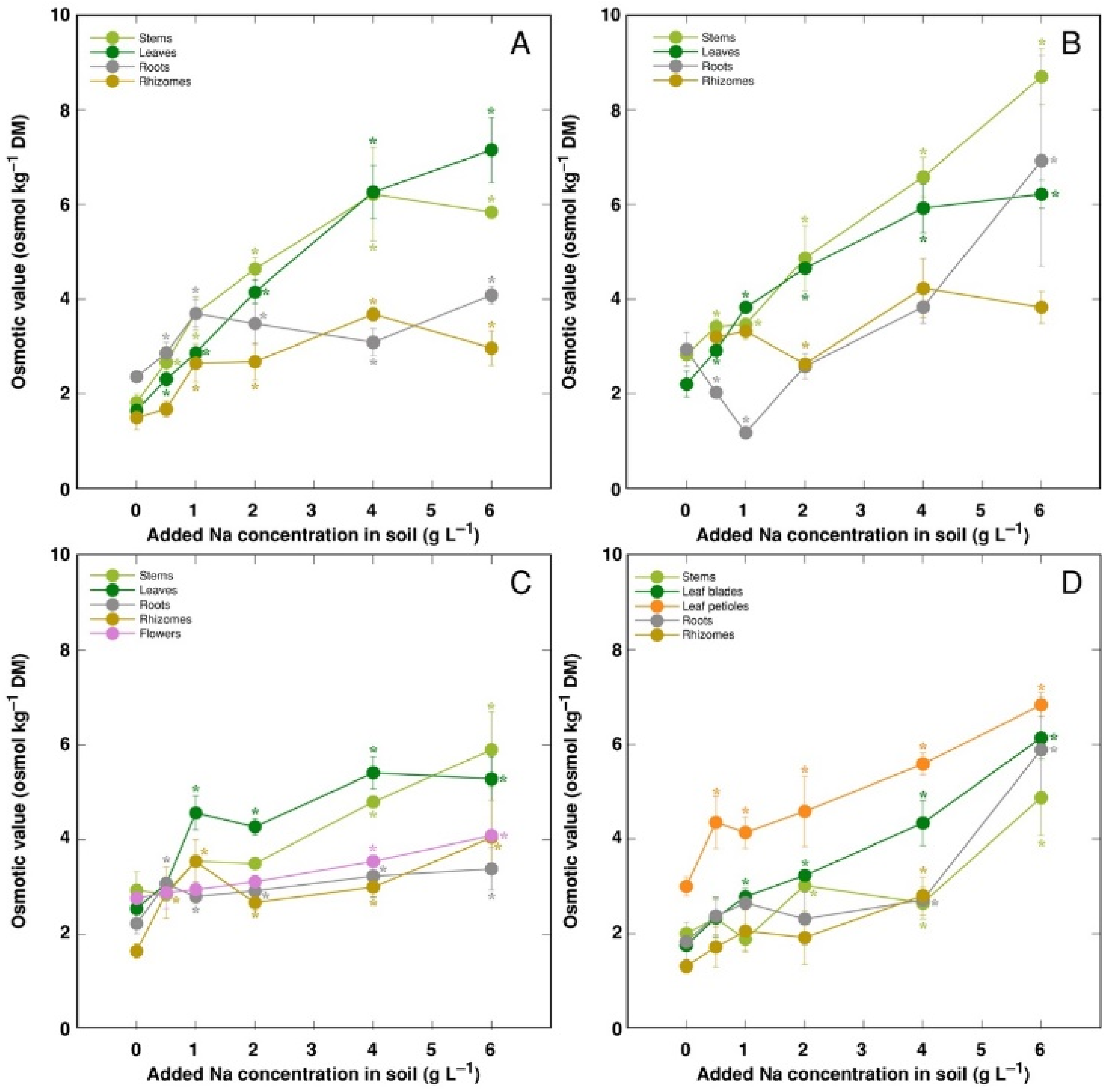
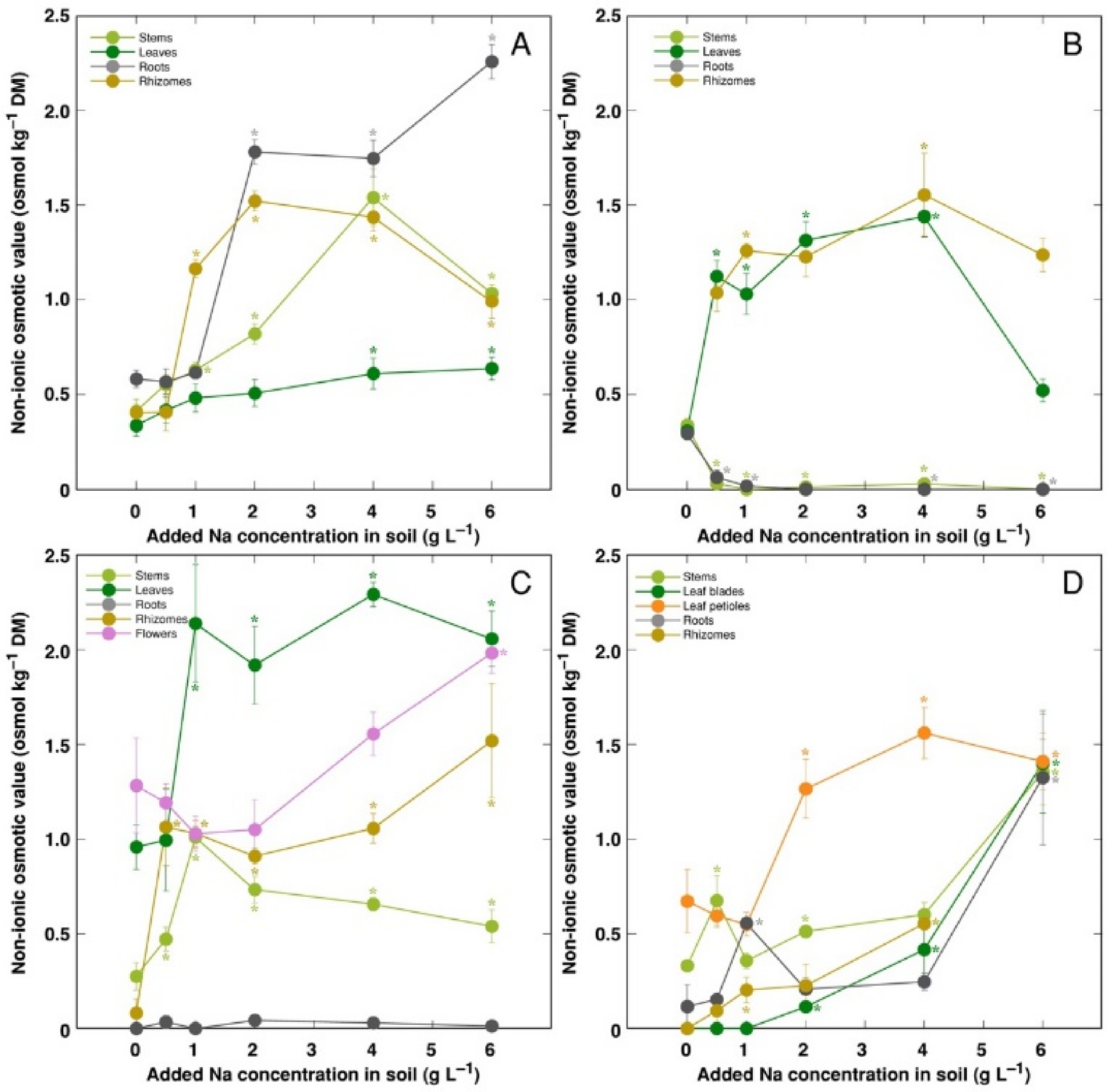
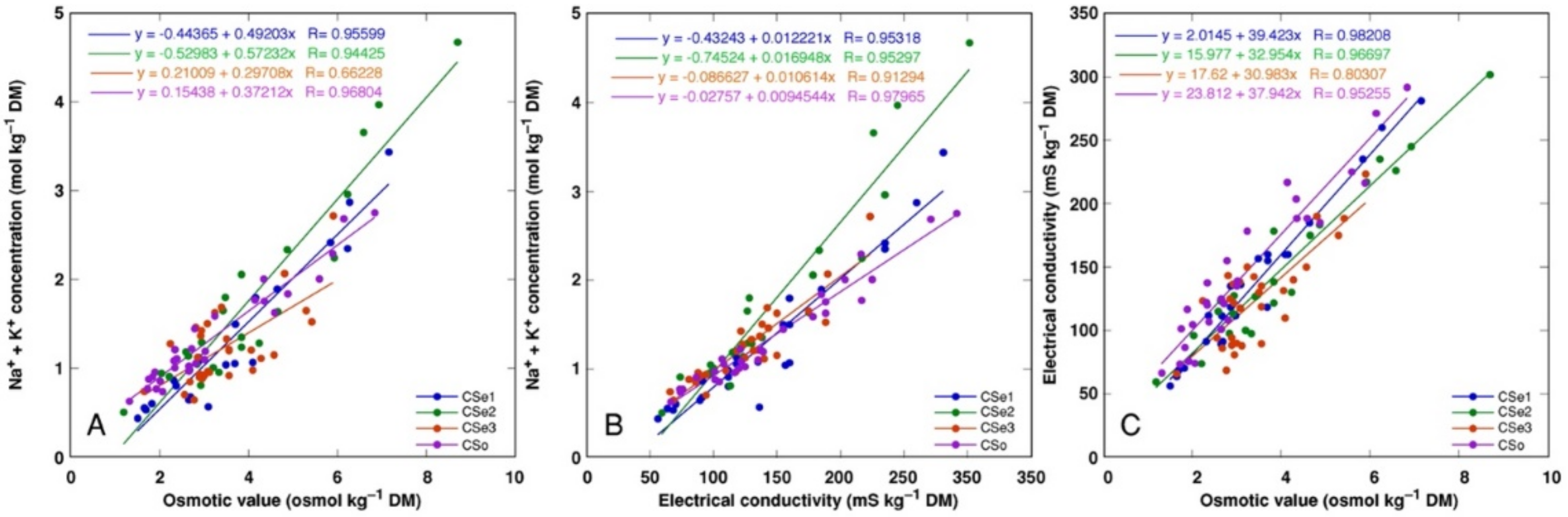
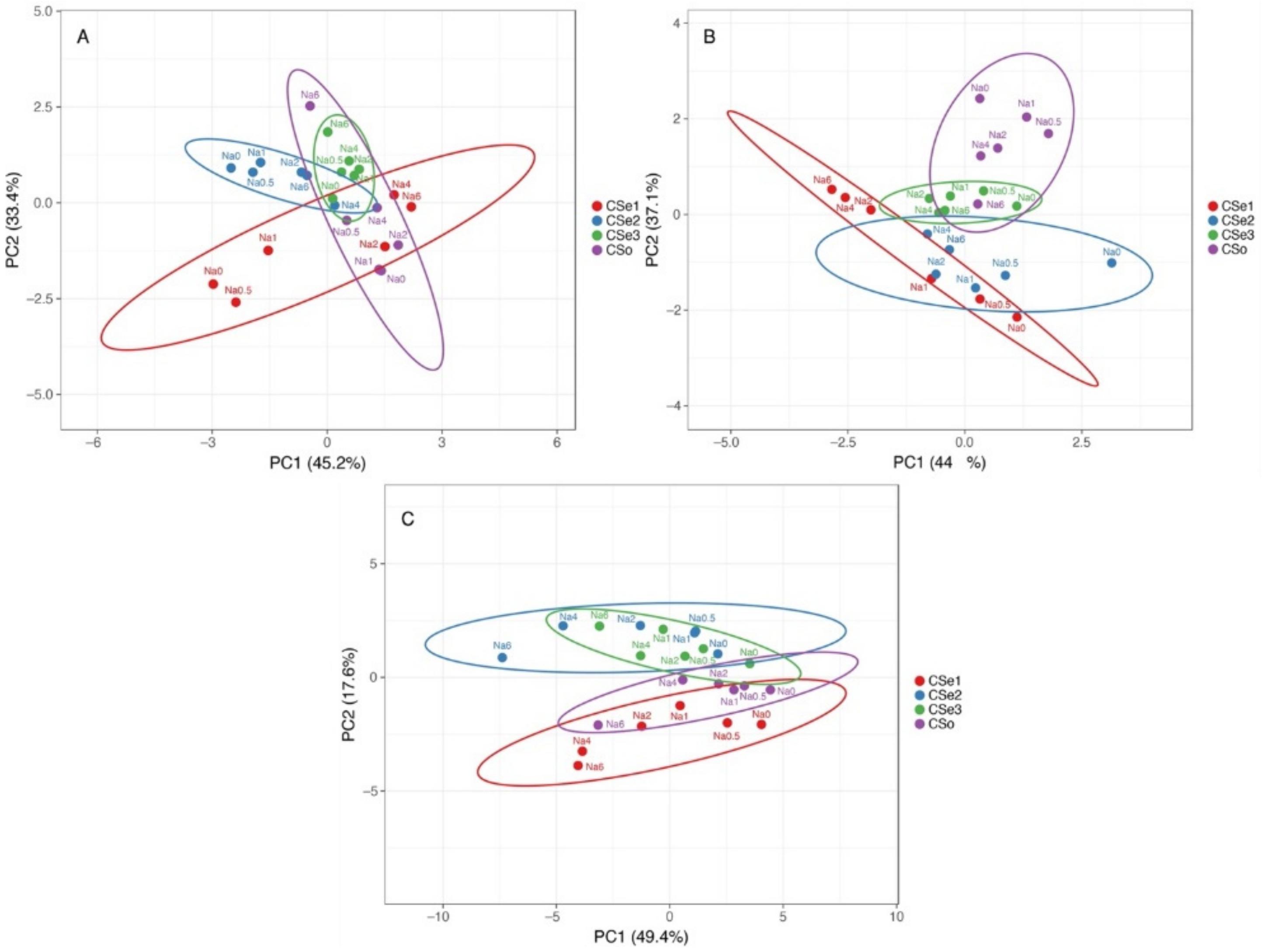
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rozema, J.; Schat, H. Salt tolerance of halophytes, research questions reviewed in the perspective of saline agriculture. Environ. Exp. Bot. 2013, 92, 83–95. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Ievinsh, G.; Andersone-Ozola, U.; Landorfa-Svalbe, Z.; Karlsons, A.; Osvalde, A. Wild plants from coastal habitats as a potential resource for soil remediation. In Soil Health; Giri, B., Varma, A., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 121–144. [Google Scholar]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Rūsiņa, S. 1230 Hydrophilous tall herb fringe communities of plains and the montane to alpine levels. In European Union Protected Habitats in Latvia; Auniņš, A., Ed.; Interpretation Manual. Riga, Latvian Fund for Nature, Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 186–189. [Google Scholar]
- Duvigneaud, J. La végétation nitrophile du bord des eaux (alliance du Calystegion sepium) dans la valée de la Moselle (Allemagne, France, Grand-Duché de Luxembourg). Bull. Soc. Roy. Bot. Belg. 1978, 111, 27–35. [Google Scholar]
- Grime, J.P.; Hodgson, J.G.; Hunt, R. Comparative Plant Ecology: A Functional Approach to Common British Species; Unwin Hyman: London, UK, 1988. [Google Scholar]
- Hulisz, P.; Piernik, A.; Mantilla-Contreras, J.; Elvisto, T. Main driving factors for seacoast vegetation in the Southern and Eastern Baltic. Wetlands 2016, 36, 909–919. [Google Scholar] [CrossRef]
- Klimeš, L.; Klimešová, J. Biomass allocation in a clonal vine: Effects of intraspecific competition and nutrient availability. Folia Geobot. Phytotax. 1994, 29, 237–244. [Google Scholar] [CrossRef]
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and traits values for Swedish vascular plants. Ecol. Indic. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Arafeh, R.; Kadereit, J.W. Long-distance seed dispersal, clone longevity and lack of phylogeographical structure in the European distributional range of the coastal Calystegia soldanella (L.) R. Br. (Convolvulaceae). J. Biogeogr. 2006, 33, 1461–1469. [Google Scholar] [CrossRef]
- Bae, C.-Y.; Hwang, J.-S.; Bae, J.-J.; Choi, S.-C.; Lim, S.-H.; Choi, D.-G.; Kim, J.-G.; Choo, Y.-S. Physiological responses of Calystegia soldanella under drought stress. J. Ecol. Environ. 2013, 36, 255–265. [Google Scholar] [CrossRef][Green Version]
- Noda, A.; Nomura, N.; Setoguchi, H. Chlorophyll fluorescence in response to salt stress in the coastal plant Calystegia soldanella: A comparison between coastal and freshwater populations. Bull. Nat. Mus. Nat. Sci. Ser. B 2009, 35, 123–129. [Google Scholar]
- Spanò, C.; Bruno, M.; Bottega, S. Calystegia soldanella: Dune versus laboratory plants to highlight key adaptive physiological traits. Acta Physiol. Plant. 2013, 35, 1329–11336. [Google Scholar] [CrossRef]
- Ishikawa, S.-I.; Oikawa, T.; Furukawa, A. Responses of photosynthesis, leaf conductance and growth to different salinities in three coastal dune plants. Ecol. Res. 1991, 6, 217–226. [Google Scholar] [CrossRef]
- Hasegawa, P.M. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot. 2013, 92, 19–31. [Google Scholar] [CrossRef]
- Maathuis, F.J.M. Sodium in plants: Perception, signalling and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849–858. [Google Scholar] [CrossRef]
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2014, 6, 873. [Google Scholar] [CrossRef]
- Tipirdamaz, R.; Gagneul, D.; Duhazé, C.; Aïnouche, A.; Monnier, C.; Özkum, D.; Larher, F. Clustering of halophytes from an inland salt marsh in Turkey according to their ability to accumulate sodium and nitrogenous osmolytes. Environ. Exp. Bot. 2006, 57, 139–153. [Google Scholar] [CrossRef]
- Matinzadeh, Z.; Breckle, S.-W.; Mirmaccoumi, M.; Akhani, H. Ionic relationships in some halophytic Iranian Chenopodiaceae and their rhizospheres. Plant Soil 2013, 372, 523–539. [Google Scholar] [CrossRef]
- Milić, D.; Luković, J.; Zorić, L.; Vasin, J.; Ninkov, J.; Zeremski, T.; Milić, S. Halophytes relations to soil ionic composition. J. Serb. Chem. Soc. 2013, 78, 1259–1268. [Google Scholar] [CrossRef]
- Matinzadeh, Z.; Akhani, H.; Abedi, M.; Palacio, S. The elemental composition of halophytes correlates with key morphological adaptations and taxonomic groups. Plant Physiol. Biochem. 2019, 141, 259–278. [Google Scholar] [CrossRef]
- Bendaly, A.; Messedi, D.; Smaoui, A.; Ksouri, R.; Bouchereau, A.; Abdelly, C. Physiological and leaf metabolome changes in the xerohalophyte species Atriplex halimus induced by salinity. Plant Physiol. Biochem. 2016, 103, 208–218. [Google Scholar] [CrossRef]
- Ievinsh, G. Coastal plant species as electrolytophytes: Effect of NaCl and light intensity on accumulation characteristics of Atriplex glabriuscula from coastal drift lines. Environ. Exp. Biol. 2020, 18, 95–105. [Google Scholar]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium, potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2021, 274, 151748. [Google Scholar] [CrossRef]
- Ko, J.-M.; Park, H.-J.; Min, B.-M.; Cha, H.-C. Effects of various pretreatments on seed germination of Calystegia soldanella (Convolvulaceae), a coastal sand dune plant. J. Plant Biol. 2004, 47, 396–400. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Bueno, M.; Lendinez, M.L.; Aparicio, C.; Cordovilla, M.P. Germination and growth of Atriplex prostrata and Plantago coronopus: Two strategies to survive in saline habitats. Flora 2017, 227, 56–63. [Google Scholar] [CrossRef]
- Tabot, P.T.; Adams, J.B. Ecophysiology of salt marsh plants and predicted responses to climate change in South Africa. Ocean Coast. Manag. 2013, 80, 89–99. [Google Scholar] [CrossRef]
- Al Hassan, M.; Pacurar, A.; López-Gresa, M.P.; Donat-Torres, M.P.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Effects of salt stress on three ecologically distinct Plantago species. PLoS ONE 2016, 11, e0160236. [Google Scholar]
- Ltaelf, H.B.; Sakhraoui, A.; González-Orenga, S.; Faz, A.L.; Boscaiu, M.; Vicente, O.; Rouz, S. Responses to salinity in four Plantago species from Tunisia. Plants 2021, 10, 1392. [Google Scholar]
- Al Hassan, M.; Estrelles, E.; Soriano, P.; López-Gresa, M.P.; Bellés, J.M.; Boscaiu, M.; Vicente, O. Unraveling salt tolerance mechanisms in halophytes: A comparative study on four Mediterranean Limonium species with different geographic patterns. Front. Plant Sci. 2017, 8, 1438. [Google Scholar] [CrossRef]
- Mi, P.; Yuan, F.; Guo, J.; Han, G.; Wang, B. Salt glans play a pivotal role in the salt resistance of four recretohalophyte Limonium Mill. species. Plant Biol. 2021, 23, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Kozminska, A.; Al Hassan, M.; Hanus-Fajerska, E.; Naranjo, M.A.; Boscaiu, M.; Vicente, O. Comparative analysis of water deficit and salt tolerance mechanisms in Silene. South Afr. J. Bot. 2017, 117, 193–206. [Google Scholar] [CrossRef]
- Ibraheem, F.; Al-Zahrani, A.; Mosa, A. Physiological adaptation of three wild halophytic Suaeda species: Salt tolerance strategies and metal accumulation capacity. Plants 2022, 11, 537. [Google Scholar] [CrossRef] [PubMed]
- Köhl, K.I. The effect of NaCl on growth, dry matter allocation and ion uptake in salt marsh and inland populations of Armeria maritima. New Phytol. 1997, 135, 213–225. [Google Scholar] [CrossRef]
- Köhl, K.I. NaCl homeostasis as a factor for the survival of the evergreen halophyte Armeria maritima (Mill.) Willd. under salt stress in winter. Plant Cell Environ. 1997, 20, 1253–1263. [Google Scholar] [CrossRef]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress om growth, imorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Envrion. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Y.; Mi, P.; Wang, B.; Yuan, F. Salt-tolerance screening in Limonium sinuatum varieties with different flower colors. Sci. Rep. 2021, 11, 14562. [Google Scholar] [CrossRef]
- Jēkabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Romanovs, M.; Ievinsh, G. Effect of salinity on growth, ion accumulation and mineral nutrition of different accessions of a crop wild relative legume species, Trifolium fragiferum. Plants 2022, 11, 797. [Google Scholar] [CrossRef]
- Zörb, C.; Sümer, A.; Sungur, A.; Flowers, T.J.; Özcan, H. Ranking of 11 coastal halophytes from salt marshes in northwest Turkey according their salt tolerance. Turk. J. Bot. 2013, 37, 1125–1133. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Bazihizina, N.; Barreett-Lennard, E.G.; Colmer, T.D. Plant growth and physiology under heterogeneous salinity. Plant Soil 2012, 354, 1–19. [Google Scholar] [CrossRef]
- Valenzuela, F.J.; Reineke, D.; Leventini, D.; Chen, C.C.L.; Barrett-Lennard, E.G.; Colmer, T.D.; Dodd, I.C.; Shabala, S.; Brown, P.; Bazihizina, N. Plant responses to heterogeneous salinity: Agronomic relevance and research priorities. Ann. Bot. 2022, 129, 499–518. [Google Scholar] [CrossRef] [PubMed]
- Haak, D.C.; Fukao, T.; Grene, R.; Hua, Z.; Ivanov, R.; Perrella, G.; Li, S. Multilevel regulation of abiotic stress responses in plants. Front. Plant Sci. 2017, 8, 1564. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.E.; Ripley, B.S. Resolving the differences in plant burial responses. Austral Ecol. 2010, 35, 53–59. [Google Scholar] [CrossRef]
- Neubauer, H.F. Die Auslaufertriebe von Calystegia sepium (L.) R. Br. (Convolvulaceae). Angew. Bot. 1988, 62, 221–224. [Google Scholar]
- Klimešová, J.; Martínková, J.; Herben, T. Horizontal growth: An overlooked dimension in plant trait space. Perspect. Plant Ecol. Evol. Syst. 2018, 32, 18–21. [Google Scholar] [CrossRef]
- White, A.C.; Colmer, T.D.; Cawthray, G.R.; Hanley, M.E. Variable response of three Trifolium repens ecotypes to soil flooding by seawater. Ann. Bot. 2014, 114, 347–355. [Google Scholar] [CrossRef]
- Liu, B.; Poolman, B.; Boersma, A.J. Ionic strength sensing in living cells. ACS Chem. Biol. 2017, 12, 2510–2514. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef]
- Belkheiri, O.; Mulas, M. The effects of salt stress on growth, water relations and ion accumulation in two halophyte Atriplex species. Environ. Exp. Bot. 2013, 86, 17–28. [Google Scholar] [CrossRef]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, D.R. Ion accumulation in halophytes. In Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; Hasanuzzaman, M., Shabala, S., Fujita, M., Eds.; CABI: Wallingford, UK, 2019; pp. 137–151. [Google Scholar]
- Pongrac, P.; Vogel-Mikuš, K.; Regvar, M.; Kaligarič, M.; Vavpetič, P.; Kelemen, M.; Grlj, N.; Shelef, O.; Golan-Goldhirsh, A.; Rachmilevitch, S.; et al. On the distribution and evaluation of Na, Mg and Cl in leaves of selected halophytes. Nucl. Instr. Methods Phys. Res. B 2013, 306, 144–149. [Google Scholar]
- Kavi Kishor, P.B.; Sangan, S.; Amrutha, R.N.; Sri Laxmi, P.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Shabala, S.; Shabala, L. Ion transport and osmotic adjustment in plants and bacteria. BiolMol Concepts 2011, 2, 407–419. [Google Scholar]
- Murakeözy, É.; Nagy, Z.; Duhazé, C.; Bouchereau, A.; Tuba, Z. Seasonal changes in the levels of compatible solutes in three halophytic species in inland saline vegetation in Hungary. J. Plant Physiol. 2003, 160, 395–401. [Google Scholar] [CrossRef]
- Karlsons, A.; Osvalde, A.; Ņečajeva, J.; Ievinsh, G. Changes of nutritional status of coastal plants Hydrocotyle vulgaris and Aster tripolium at elevated soil salinity. Acta Univ. Latv. 2008, 745, 165–177. [Google Scholar]
- Ohga, K.; Muroi, M.; Hayakawa, H.; Yokoyama, J.; Ito, K.; Tebayashi, S.-I.; Arakawa, R.; Fukuda, T. Coastal adaptation of Adenophora triphylla var. japonica. Am. J. Plant Sci. 2013, 4, 596–601. [Google Scholar]
- Bowsher, A.W.; Kearns, P.J.; Popovic, D.; Lowry, D.B.; Shade, A. Locally adapted Mimulus ecotypes differentially impact rhizosphere bacterial and archaeal communities in an environment-dependent manner. Phytobiomes J. 2020, 4, 53–63. [Google Scholar]
- Itoh, M. Phenotypic variation and adaptation in morphology and salt spray tolerance in coastal and inland populations of Setaria viridis in central Japan. Weed Res. 2021, 61, 199–209. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressfull “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- He, Y.; Li, Z. Epigenetic environmental memories in plants: Establishment, maintenance, and reprogramming. Trends Genet. 2018, 34, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Perrone, A.; Martinelli, F. Plant stress biology in epigenomic era. Plant Sci. 2020, 294, 110376. [Google Scholar] [CrossRef] [PubMed]
- Holeski, L.M.; Jander, G.; Agrawal, A.A. Transgenerational defense induction and epigenetic inheritance in plants. Trends Ecol. Evol. 2012, 27, 618–626. [Google Scholar] [CrossRef]
- Latzel, V.; Rendina González, A.P.; Rosenthal, J. Epigenetic memory as a basis for intelligent behavior in clonal plants. Front. Plant Sci. 2016, 7, 1354. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species, Accession | Code | Habitat | Soil Electrical Conductivity (mS m−1) | Establishment of Stock Culture/Propagation | Characteristics | Locality (Coordinates) |
|---|---|---|---|---|---|---|
| Calystegia sepium 1 | CSe1 | Coastal, sandy beach with perennial vegetation | 623 ± 105 | Rhizome fragments/stem explants with leaf | White flowers | Mērsrags, Latvia (N 57°21′57″ E 23°7′21″) |
| Calystegia sepium 2 | CSe2 | Inland, mesophytic vegetation on banks of pond | 86 ± 5 | Rhizome fragments/stem explants with leaf | White flowers | Salaspils, Latvia (N 56°51′32″ E 24°30′38″) |
| Calystegia sepium 3 | CSe3 | Inland, dry grassland vegetation on a steep riverbank | 83 ± 2 | Rhizome fragments/stem explants with leaf | Light pink flowers | Ogre, Latvia (N 56°48′59″ E 24°36′59″) |
| Calystegia soldanella | CSo | Coastal, sand dunes | NA | Seeds/stem explants with leaf | NA | NA |
| Parameter | C. sepium 1 | C. sepium 2 | C. sepium 3 |
|---|---|---|---|
| H2O (g g−1 DM) | 7.5 ± 0.8 a | 5.5 ± 0.1 b | 4.4 ± 0.2 c |
| Na+ (g kg−1 DM) | 26.33 ± 1.97 a | 0.95 ± 0.07 c | 2.23 ± 0.07 b |
| K+ (g kg−1 DM) | 32.8 ± 2.9 b | 26.6 ± 4.3 c | 41.0 ± 1.0 a |
| Na+ (mol L−1) | 0.156 ± 0.011 a | 0.008 ± 0.001 c | 0.022 ± 0.001 b |
| EC (mS cm−1 kg−1 DM) | 113 ± 6 a | 57 ± 5 b | 60 ± 1 b |
| K+ (mol L−1) | 0.115 ± 0.010 b | 0.123 ± 0.018 b | 0.237 ± 0.006 a |
| EC (mS cm−1 L−1) | 15.4 ± 0.9 a | 10.2 ± 0.7 b | 13.5 ± 0.1 a |
| K:Na (mol) | 0.8 ± 0.1 c | 17.2 ± 3.6 a | 10.8 ± 0.3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jēkabsone, A.; Ievinsh, G. Salinity Tolerance and Ion Accumulation of Coastal and Inland Accessions of Clonal Climbing Plant Species Calystegia sepium in Comparison with a Coastal-Specific Clonal Species Calystegia soldanella. Int. J. Plant Biol. 2022, 13, 381-399. https://doi.org/10.3390/ijpb13040032
Jēkabsone A, Ievinsh G. Salinity Tolerance and Ion Accumulation of Coastal and Inland Accessions of Clonal Climbing Plant Species Calystegia sepium in Comparison with a Coastal-Specific Clonal Species Calystegia soldanella. International Journal of Plant Biology. 2022; 13(4):381-399. https://doi.org/10.3390/ijpb13040032
Chicago/Turabian StyleJēkabsone, Astra, and Gederts Ievinsh. 2022. "Salinity Tolerance and Ion Accumulation of Coastal and Inland Accessions of Clonal Climbing Plant Species Calystegia sepium in Comparison with a Coastal-Specific Clonal Species Calystegia soldanella" International Journal of Plant Biology 13, no. 4: 381-399. https://doi.org/10.3390/ijpb13040032
APA StyleJēkabsone, A., & Ievinsh, G. (2022). Salinity Tolerance and Ion Accumulation of Coastal and Inland Accessions of Clonal Climbing Plant Species Calystegia sepium in Comparison with a Coastal-Specific Clonal Species Calystegia soldanella. International Journal of Plant Biology, 13(4), 381-399. https://doi.org/10.3390/ijpb13040032