Abstract
Iron is the most abundant mineral in the human body and plays essential roles in sustaining life, such as the transport of oxygen to systemic organs. The Fenton reaction is the reaction between iron and hydrogen peroxide, generating hydroxyl radical, which is highly reactive and highly toxic to living cells. “Ferroptosis”, a programmed cell death in which the Fenton reaction is closely involved, has recently received much attention. Furthermore, various applications of the Fenton reaction have been reported in the medical and nutritional fields, such as cancer treatment or sterilization. Here, this review summarizes the recent growing interest in the usefulness of iron and its biological relevance through basic and practical information of the Fenton reaction and recent reports.
Keywords:
antioxidants; cancer; Fenton reaction; hydrogen peroxide; hygiene; iron; nanomedicine; oxidative stress; polyphenol; vitamin C 1. Introduction
It has only been around 100 years since people started research on food in terms of “modern nutritional science” [1]. In the latter part of the 1800s, Lavoisier investigated chemical oxidation in living things, which is thought to be the beginning of nutritional science. In the early 1900s, individual food components began to be found. For example, vitamin B1 (oryzanin) was found in trials to overcome the deficiencies of military patients [2]. Vitamin E was isolated from wheat germ oil, which was found to be involved in rat reproduction [3]. In contrast, the discovery of minerals dates back to 6000 B.C, but it took centuries to recognize their roles in biological systems. Among the various minerals, iron is one of the well-known and -utilized minerals from ancient times and was estimated to be present in the blood in the 1700s [4]. Iron is the most abundant transition metal on Earth’s surface, with 3–4 g of iron in the body of a healthy adult human [5,6]. Since iron is the most abundant transition metal in the human body, its contribution toward various biological activities has long been the focus of growing attention [7]. For example, anemia is one of the major manifestations of iron deficiency. In 1925, Fontès and Thivolle found that iron-deficient horses had lower serum iron concentrations [8]. In the human body, most extracellular iron is bound to iron-binding proteins (such as transferrin and lactoferrin) [9]. Heme proteins in red blood cells play an important role in transporting oxygen to organs [10].
On the other hand, iron is involved not only in the delivery of oxygen in our body but also in DNA synthesis and/or repair [11], indicating this mineral is essential for the survival of living things. Iron also works as a cofactor to facilitate various enzymes, such as catalase and cytochromes. The roles of iron in the body are particularly involved in redox reactions due to its preferable affinity to oxygen. In the 1890s, Henry John Horstman Fenton found the redox reaction between iron (II) and hydrogen peroxide (H2O2) to produce hydroxyl radical (OH•), called the Fenton reaction [12]. This reaction potentially occurs in the human body and is thought to regulate complicated systems, which is related to homeostasis. The products of the Fenton reaction OH• is highly reactive particles that induce oxidative damage to cells, but this is also an aspect that can be a therapeutic strategy for cancer patients. Several medicines (e.g., doxorubicin (DOX) [13], β-lapachone [14], and cisplatin [15]) include mechanisms of reaction that have applied the Fenton reaction to generate the poison, OH•, to cancer cells. Additionally, numerous food components daily consumed have beneficial effects in the human body, such as on chronic diseases and on immune systems [16,17,18]. The antioxidant reaction is one of the major properties of such food components (e.g., polyphenols and vitamins). Considering reactive oxygen species (ROS) generated by vitamin C [19] and chelating metal iron by flavonoids [20], iron potentially affects the bioactivities of absorbed and metabolized food components in the body. Furthermore, there is growing interest in programed death “ferroptosis” related to the Fenton reaction [21]. Against these backgrounds, this review summarizes the recent growing interest in the usefulness of iron and its biological relevance through basic and practical information of the Fenton reaction and recent reports.
2. Fenton Reaction
The Fenton reaction is the reaction of iron (II) with H2O2, reported by Henry John Horstman Fenton in 1894 [12]. In 1876, his student found that a mixture of H2O2, tartaric acid, ferrous salt and water turned a violet color. This is known as the Fenton reaction (Reaction (1)):
Fe2+ + H2O2 → Fe3+ + OH− + OH•
While Fenton speculated the mechanism of oxidation by H2O2 and iron (II), some researchers doubted the formation of OH• in one-electron reduction by iron (II). In 1931, Haber and Wilstatter mentioned the hydroxy radical in radical chain mechanisms (Reaction (2) and (3)) [22]. They described that chain reactions are initiated by enzymes, specifically catalase:
OH + H2O2 → H2O + O2H
O2H + H2O2 → O2 + H2O + OH
Thereafter, Harber and Weiss explained the decomposition of H2O2 by iron (II) using Reaction (4) to (6), where the Fenton reaction initiates and Reaction (6) terminates the chain reactions [23]:
OH• + H2O2 → H2O + O2•− + H+
O2•− + H+ + H2O2 → O2 + H2O + OH•
Fe2+ + HO + H+ → Fe3+ + H2O
In 1937, Weiss explained the reaction mechanism of catalase: an anion H2O2 reduces iron (III) to iron (II), and then iron (II) reduces H2O2 to OH• and water, followed by chain reaction (5, 6), which is collectively referred to as the Haber–Weiss reaction. The mechanism of the Fenton reaction has studied and discussed, among which detailed equilibrium principles have been well summarized by Stanbury [24]. The Fenton reaction is affected by the environmental pH and concentration of iron. The major ROS generated from the Fenton reaction are oxoiron (IV) species at pH > 3, and OH• at more acidic conditions [25,26,27]. Two mechanisms, the “radical mechanism” and the “complex mechanism”, contribute to the iron-catalyzed disproportionation of H2O2 and the Fenton reaction. The products obtained from these reactions are different. In the “radical mechanism”, Fe2+ and Fe3+ react with H2O2 to produce OH• and superoxide, respectively. In the “complex mechanism”, Fe2+ and Fe3+ react with H2O2 to produce FeO2+ and FeO3 +, respectively. In 2013, more than 100 years after the Fenton reaction was proposed, successful detection of Fe(IV) was reported [28]. Additionally, it has been suggested that fellyl ion controls iron cycling by the Fenton reaction in a cloud as well as Fe2+ and Fe3+ [29]. Such reports indicate how great the impact and complexity of this reaction is. The use of other transition metals such as copper leads to a reaction similar to the Fenton reaction, called the Fenton-like reaction. Although the Fenton reaction initially began to be used for analytical purposes, chelation or sequestration of transition metals involving Fenton and Fenton-like reactions have been found to play important roles in the internal and external environments of living things.
3. Fenton Reaction in Body
3.1. Iron as a Nutrient
Nutrients are essential for living things, among which proteins, fats, and carbohydrates are three major nutrients. In the context of the human diet, minerals are elements, except H, C, N, and O (the main components of three major nutrients: organic compounds), that maintain or regulate biological systems, and account for approximately 4% of the human body. Of these, 16 types of elements (Na, N, P, K, S, Ca, Mg, iodine, Se, Cr, Co, Fe, Mn, Zn, Cu, and Mo) are thought to play particularly important roles. They are classified into two groups based on the required amount (more than 100 mg/day: Na, N, P, K, S, Ca, and Mg; less than 100 mg/day: iodine, Se, Cr, Co, Fe, Mn, Zn, Cu, and Mo).
Iron is present in all human cells, with an average of 2.4 g in women and 3.8 g in men, with daily losses of 1–2 mg [30] (Figure 1). Examples of iron-rich foods include oysters, clams, mussels, beef or chicken liver, and poultry while non-heme iron is contained in beans, spinach, nuts, and seeds. One of the most important roles of iron is to transport oxygen in hemoglobin (Hb). This protein, consisting of 96% of blood cells [31], provides oxygen to the whole body from the lungs or other airway organs and supports metabolism. Hb iron binds up to four oxygen molecules in the form of Fe2+ or Fe3+ [32]. Additionally, other oxygen storage protein and enzymes bind to iron (hemoglobin, 2500 mg iron; myoglobin, 130 mg iron; enzymes, 150 mg iron) [33]. Anemia due to a general iron deficiency (Hb <13 g/dL in males, <12 g/dL in females, <11 g/dL during pregnancy) is mainly due to biological mechanisms (e.g., iron deficiency, hemolytic anemia, and anemia of inflammation) and/or erythrocyte morphology. Iron deficiency occurs when there is an insufficient supply of iron against the amount needed such as during periods of high iron requirements (e.g., infancy and pregnancy) and/or iron loss exceeds intake. Iron is absorbed via human intestinal mucosa in heme and non-heme forms, whereas heme-iron is reported to be more readily absorbed through the folate transporter [34]. Non-heme iron, Fe2+ and Fe3 +, is transported into the duodenal cytoplasm via divalent metal iron transporter-1 (DMT-1) in Fe2+ [35], where Fe3+ is previously reduced to Fe2+ by cytochrome b reductase and/or other reductants. After being transported into cells, iron either binds to ferritin for storage or is transported into the blood stream via ferroportin as Fe2 +. Iron is then oxidized by membrane-bound ferroxidase hephaestin and ceruloplasmin to be incorporated into transferrin to form the transferrin–Fe3+ complex. Hepcidin is a peptide hormone excreted from the liver that binds to ferroportin, the only iron efflux transporter in the blood, and regulates iron homeostasis by promoting internalization and degradation of the transporter [36]. Hepcidin completely occludes the iron pathway by binding ferroportin with an outward-open conformation [37]. While this section only presented limited information on iron absorption and metabolism, more detailed clinical characteristics of iron deficiency are described by Camaschella et al. and Pasricha et al. [33,38].
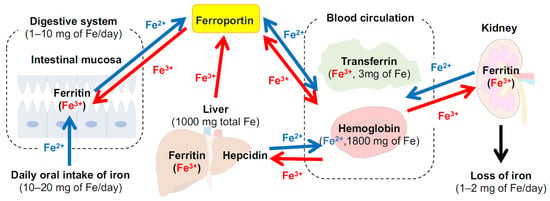
Figure 1.
Distribution of iron in the body and the main organs involved in the regulation of iron metabolism (modified from permission from [39] under the Creative Commons CC BY 4.0 license, https://creativecommons.org/licenses/by/4.0/ (accessed on 28 July 2022)). Values data for iron levels are obtained from Lesjac et al. [40]).
As mentioned above, iron is absorbed in the intestine as either heme or non-heme forms, but other food-derived components are also absorbed in the intestine, suggesting that interactions with them may affect iron absorption (Figure 2). For example, quercetin has been reported to inhibit intestinal iron absorption by different mechanisms, through chelation in an acute duodenal injection study and by suppressing ferroportin expression in an oral administration study in rats [41]. Tea consumption also reduces the bioavailability of iron, possibly due to polyphenols such as tannins [42,43]. It has also been reported that the intake of a high-fat diet inhibits intestinal iron absorption, causing iron deficiency [44], and that the amount of absorbed iron in overweight women was two-thirds of the normal value [45]. In contrast, ascorbic acid is well known to increase iron absorption related to iron reduction and the intake of ascorbic acid attenuates the above inhibitory effect of polyphenols [46]. The major peptide hormone, hepcidin, is also affected by flavonoids; myricetin significantly suppresses the expression of this hormone [47]. Higher concentrations and lower clearance of hepcidin due to chronic kidney disease suppress iron absorption, resulting in iron deficiency [48]. On the contrary, 17β-estradiol possibly promotes iron absorption by inhibiting hepcidin expression through an estrogen-responsive element half-site in the promoter region of the hepcidin gene [49], indicating that increased iron might be caused by another mechanism in postmenopausal women.
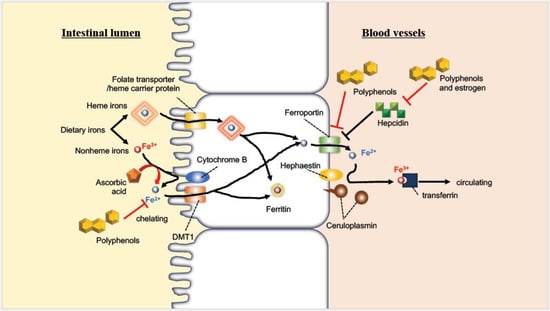
Figure 2.
Absorption behavior of iron in the intestine and the interaction with molecules derived from food (modified with permission from [50] under the Creative Commons CC BY 4.0 license, https://creativecommons.org/licenses/by/4.0/ (accessed on 28 July 2022)).
3.2. Fenton Reaction under Biological Environment
Iron is transferred from binding in transferrin as Fe3+ form and transported into cells via the transferrin receptor. Most transition metals, including iron, are involved in the generation of various free radicals due to their redox features. As “second-messengers”, ROS play essential roles in cellular life cycles, such as proliferation [51] and gene expression [52] (Figure 3). Observation with fluorescent reagents (e.g., dihydrorhodamine, coumarin-3-calboxylic acid, and endoplasmic reticulum (ER)-targeting OH• probe) has revealed intracellular localization of the generated OH•. Such studies have reported that ER by the Fenton reaction [53,54] regulates hypoxia-inducible gene expression. ER stress has been reported to regulate more than one-third of all proteins made in the cell in synthesis, folding, and structural maturation [55]. Additionally, H2O2, one type of ROS generated extracellularly, penetrates the cell membrane easily to react with intracellular iron (Fe2+ and Fe3 +), producing OH• through the Fenton and Fenton-like reaction. OH• reacts most strongly with biomolecules shorter than 1 ns [3,56], which results in the most severe damage to biological systems among ROS. It involves the induction of the oxidation of molecules. OH• produced through the Fenton reaction has been reported to induce DNA damage [57]. Iron released from Hb is also known to promote the degradation of deoxyribose, inducing lipid peroxidation [58,59]. Additionally, Fenton-type chemistry (e.g., peroxidases, free heme, and metal ions) is involved in the tyrosine nitration observed within tyrosine residues in proteins and used as a signature for peroxynitrite [60].
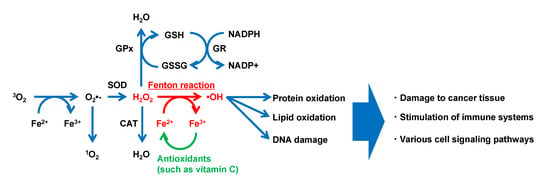
Figure 3.
Typical model of reactive oxygen species generation via the Fenton reaction in a biological environment. CAT, catalase; GPx, glutathione peroxidase; GR, glutathione reductase; GSH, reduced glutathione; GSSG, oxidized glutathione; NADPH, reduced form of nicotinamide adenine dinucleotide phosphate; NADP +, oxidized form of nicotinamide adenine dinucleotide phosphate; SOD, superoxide dismutase (modified with permission from [61] under the Creative Commons CC BY 3.0 license, https://creativecommons.org/licenses/by/3.0/ (accessed on 28 July 2022)).
In heme proteins, the transition of iron is essential for the performance of their functions. Among them, cytochrome P450 is one of the largest enzyme families, in which as many as 18,000 P450s have been identified [62] and is well known to work in detoxification of drugs or other xenobiotics. This enzyme is made up of 40–50 kDa single polypeptides with a long I helix and H-bond between Cys, and a peptide NH group is regarded as the key factor to heme iron redox. Fe2+ centered in the enzyme binds to O2 to form oxy complex followed by the second electron transfer and heterolytic cleavage, during which ROS can be produced. Heme degradation catalyzed by heme oxygenases also generates ROS by non-heme iron.
Recently, ferritinophagy and ferroptosis have attracted attention as iron-dependent cell death. Ferritinophagy consists of the autophagic degradation of ferritin to regulate iron homeostasis [63]. Increased intracellular iron levels following the release from ferritin promotes ROS production, leading to cell death; radiation is reported to induce autophagic iron-dependent death in cancer cells, which is a promising therapeutic strategy [64]. “Ferroptosis”, coined by Brent Roark Stockwell and Scott Dixon in 2012 [65], is one type of regulated cell death dependent on iron or ROS, which is distinct from other types such as apoptosis, necrosis, and autophagic death at the morphological, biochemical, and genetic levels. Excessive iron in cell lines (harboring RAS mutations with increased iron uptake and decreased iron storage) induces ferroptosis, which is regulated by suppression of the master transcription factor of iron metabolism [66], indicating that ferroptosis is iron dependent. Although the correlation between autophagy and ferroptosis is not well understood, Park et al. elucidated that ROS-induced autophagy plays an important role in ferritin degradation and transferrin receptor 1 expression during ferroptosis [67]. More details on ferroptosis are beyond the scope of this review and are reviewed and described by Xie et al. [68], Chen et al. [69], and Bebber et al. [70].
4. Use of the Fenton Reaction for Drugs
ROS are regarded to cause intracellular lipid peroxidation, leading to ferroptosis. Therefore, the Fenton reaction has been challenged for use in directly attacking cancer cells, but it is difficult to treat them because of the low amounts of generated OH• [71]. In recent years, various nanoparticles that enhance the effectiveness of Fenton reactions for drug applications (nanomedicines) have been reported. A simple scheme is depicted in Figure 4. Previous reports on such an approach have already been well reviewed by Meng et al. [72], Ranji-Burachaloo et al. [73], and Miyazawa et al. [74], so the present review focuses on very recent reports (from 2020) on nanomedicines using the Fenton reaction.
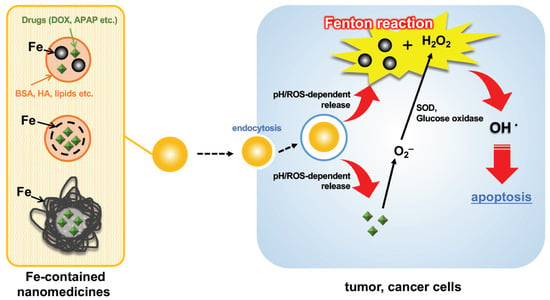
Figure 4.
Depicted representative scheme of cell apoptosis by Fe-containing nanomedicines via the Fenton reaction. DOX, doxorubicin; APAP, amionoacetophen; BSA, bovine serum albumin; HA, hyaluronic acid; SOD, superoxide dismutase; ROS, reactive oxygen species.
Xing et al. prepared an iron-loaded liposome using hollow mesoporous Prussian blue co-delivering iron, unsaturated lipids, and a photothermal converter. Controlled passive targeting enabled efficient photothermal effects and ferroptosis of these liposomes with low toxicity [75]. Tian et al. prepared ultra-small ellagic acid-Fe-bovine serum albumin nanoparticles and showed acceleration of Fe3+/Fe2+ transformation by strong reduction of endogenous H2S [76]. Sang et al. first prepared PZIF-67 nanoparticles with SOD (super oxide dismutase)-like activity and an OH -generating ability [77]. Gao et al. prepared the nanoparticles encapsulating light-responsive CO prodrugs by self-assembly of photoresponsive polymers. These nanoparticles accumulated in mitochondria and light-responsively released CO and the prodrugs, followed by the Fenton reaction, which generated high levels of ROS to decrease cell viability. Actually, intravenous injection of the nanoparticle significantly suppressed the tumor growth with an increase in ROS [78]. You et al. combined NIR irradiation with nanoparticles. Functional nanoparticles with internally encapsulated functional benzothiazole complexes (eTB2) and the photosensitizer indocyanine green induced FeTB2 release and Fenton reaction under NIR irradiation [79]. Wu et al. prepared hollow porous carbon coated with FeS2-based nanoparticles. Prepared hollow porous carbon revealed that the conversion of NIR heat into an effective temperature rise by the carbon shell and the reduction of Fe3+ to Fe2+ by tannic acid promoted the Fenton reaction [80]. Fu et al. reported that DOX and glucose oxidase-gallic acid/iron complexes were encapsulated into zeolitic imidazole framework-8 nano particles, which induced cancer cell death by the Fenton reaction with gallic acid/iron complexes under an acidic microenvironment [81]. Chen et al. reported the preparation of biodegradable nanoparticles of Fe3O4 bound with protocatechuic acid and human serum albumin loaded with β-lapachone [82].
Correlating pH or ROS sensitivity with the Fenton reaction enables more effective and selective attack of tumor cells, which is also being used as an approach. Sun et al. prepared synergistically therapeutic nanoparticles that encapsulated acetaminophen (APAP). Sun et al. prepared nanoparticles with the ability to induce the Fenton reaction in a weakly acidic tumor microenvironment. The prepared nanoparticles showed the conversion of APAP to the toxic metabolite NAPQ1, leading to GSH depletion and accelerating the effect of the Fenton reaction [83]. Zhong et al. prepared pH-responsive nanoparticles using BSA-derived albumin as carrier nanoparticles and encapsulating triphenylphosphine-modified DOX, which could be used to target tumor mitochondria [84]. Meng et al. prepared a metal-phenolic network-based multifunctional nanocomposite coated with Fe–tannic acid complexes and reported that Fe–tannic acid was degraded by laser irradiation (808 nm) and the acidic pH of the tumor environment, resulting in drug release and the Fenton reaction, promoting the effect of tannic acid [85]. Lei et al. prepared pH-responsive nanoparticles co-encapsulated with DOX and APAP, which were released at 56.5% and 61.8%, respectively, under an acidic endosomal/lysosomal environment, synergistically promoting OH• generation by the Fenton reaction [86]. Cho et al. prepared dual (pH- and redox-)responsive magnetic nanoparticles that promote drug release under low pH and high GSH concentrations [87], and Chen et al. developed a pH/ROS-responsive multifunctional nanoplatform that inhibits tumor through chemo/photodynamic/chemodynamic combinations [88]. Jia et al. prepared multifunctional nanoparticles with a core-shell structure encapsulating Fe3O4 and demonstrated that simultaneous photothermal and chemodynamic therapy is possible [89]. In addition to tannic acid, several food components have been used as effective applications for anticancer therapy as follows: the generation of ROS by vitamin C based on the Fenton reaction of Fe3O4 nanoparticles in cells [90]; promotion of lipid peroxidation and induction of ferroptosis in anaplastic thyroid carcinoma produced by vitamin C via the Fenton reaction [91]; enhancement of linomycin release by the Fenton reaction using tea polyphenols [92].
In addition to Fenton reactions, approaches utilizing the Fenton-like reaction have also been utilized. Cheng et al. reported that the Cu2+ and polymersome complex efficiently induced the Fenton-like reaction and promoted the oxidation of iminoboronates [93]. Wang et al. reported that conjugation of nanoparticles composed of glucose oxidase, Cu2-xSe, and a membrane of 4T1 cells promoted the Fenton reaction by increasing H2O2 under NIR-II irradiation [94]. Sun et al. prepared nanotubes composed of SiO and Cu, which is advantageous for the combination of photodynamic therapy and photothermal therapy (PTT). The prepared nanotubes effectively promoted the generation of ROS by the reaction between Cu2+ with H2O2 in the Fenton-like reaction, PTT effect, and porous structure of the nanotubes [95].
5. Fenton Reaction in Food
It is known that complex interactions occur between metal ions (or protein–metal ion complexes) and food components. Research on the relationship between the Fenton reaction and food components is relatively advanced in terms of flavonoids. Flavonoids are known to have antioxidative effects and are regarded as candidates that modulate the Fenton reaction. Among the flavonoids, the antioxidant/prooxidant properties of luteolin or kaempferol in Fenton-like reactions have been reported. For example, it has been reported that coordination of luteolin or kaempferol to Cu(II) significantly suppresses the generation of hydroxyl and superoxide radicals by 80% in the Fenton-like reaction [96,97]. These Cu-flavonoid complexes are considered to have intercalation activity towards DNA, which have potential applications for disorders associated with oxidative damage. Perron et al. measured the oxidation rate of Fe2+ when several polyphenol compounds were bound and found that galloyl groups oxidize iron faster than catechol groups, suggesting that a single iron-binding moiety contributes to the protective effects of polyphenols against oxidative damage [98]. Proteins are also affected by the Fenton reaction at their amino acid residues; cysteine and methionine residues are especially easily oxidized [99,100]. Bochi et al. investigated the effects of Fenton reaction-generated advanced oxidation protein products on the gene transcription in HEK293 cells [101]. As a result, it activated the gene transcription of inflammatory genes (NF-κB, COX-2, and IL-6), possibly mediating inflammation in the kidneys. Ishikawa et al. reported that phosphoprotein phosvitin, known as iron-career in egg yolk, chelated iron more effectively than other iron-binding proteins such as ferritin and transferrin, and accelerated the oxidation of Fe2+ to inhibit the Fenton reaction [102]. In some cases, the Fenton reaction may play a role in improving food quality as an effective tool. Voltea et al. used the Fenton reaction to accelerate the oxidative brewing of white wines, enabling rapid testing to assess the susceptibility, appropriate levels of flavanols and total free sulfhydryls for subsequent processes [103]. Gharib-Bibalan et al. showed that the oxidation process via the Fenton reaction modified the color and total polyphenols, improving the quality indexes of the purified juice [104]. Blank et al. reported that the Fenton-type reaction has significant effects on the aroma of coffee beverages [105]. Yeung et al. hydrolyzed okra pectin by the Fenton reaction to obtain pectic oligosaccharides with low molecular weights (1.79–6.09 kDa) and improved bioactivity (antioxidant and anti-inflammatory) [106]. Food components should also interact with metal ions in the body, but there are few reports on this.
One of the most important concerns regarding commercial food is their safety. As the Fenton reaction generates strong toxic radicals, it is used to kill bacteria that cause food poisoning. Shi et al. developed the Fenton reaction-assisted photodynamic inactivation method, a simple system that combines calcinated melamine sponges and Fe2+ to inactivate Salmonella under light illumination [107]. Morikawa et al. developed two “green” iron catalysts with reducing and chelating ability using tea leaves and coffee grounds [108]. This system with the catalytic Fenton reaction enhanced the degradation of the contaminants into harmless compounds and disinfection of Escherichia coli. In contrast, the Fenton reaction can also work as a protective system for certain microbes. Calhoun et al. reported that Dps, a ferritin-like protein with DNA-binding properties, protects Salmonella enterica serotype Enteritidis against the common killing mechanism of bactericidal antibiotics through the Fenton reaction [109]. Since oxidation leads to food deterioration, the monitoring of food conditions is essential and several approaches using the Fenton reaction have been reported. Abbas et al. developed a simple and highly sensitive fluorometric method based on the Fenton reaction system to assess H2O2 in foods [110]. Additionally, Wang et al. developed a novel colorimetric and fluorescent ELISA based on the Fenton reaction triggered by glucose oxidation was constructed to quantitatively and qualitatively measure danofloxacin in milk [111].
6. Conclusions
This review summarized the Fenton reaction from the basic principle to the bioavailability of iron, and the latest applications in the medical and nutritional fields. In the medical fields, it appears that various nanomedicines utilize the intracellular Fenton reaction as the generating system of strongly toxic OH• to enhance their selectivity and efficiency. In the nutritional fields, the Fenton reaction has been used to kill microorganisms that cause food spoilage, and this redox system has also been applied to food processing. The Fenton reaction is also useful in the synthesis of bioethanol [112,113] and the removal of pollutants derived from drugs or food additives [114,115,116,117]. Through this review, it can be inferred that the Fenton reaction can be used as a useful technology in both the medical and nutritional fields, though the mechanism is partially unknown. Additionally, the biosafety of Fenton-reaction-based nanomedicines is insufficient and unclear. Most papers regard the Fenton reaction as being useful, with less side effects than other drugs because the reaction is regulated by H2O2 and pH. However, for example, hypoxia, related to H2O2 generation, is a typical feature of solid tumors of cancer. Further investigation into the biosafety of Fenton-reaction-based treatment is warranted. It is expected that new technologies utilizing the Fenton reaction will continue to be developed.
On the contrary, it was found that the Fenton reaction of absorbed food components has been little examined to date. For example, vitamin C is one of the well-known antioxidants in the body, but it can also act as a pro-oxidant through the Fenton reaction [118]. Simultaneously, vitamin C changes into its oxidized form, dehydroascorbic acid. The mechanism by which high-level vitamin C kills cancer cells has been the subject of much debate [119], and recent studies have described the potential contribution of dehydroascorbic acid to cancer cell destruction [120,121]. This indicates the free-iron and Fenton reaction are involved in the functions of compounds with reduction properties, but their interaction has rarely been examined. As the redox system is too complicated in the body system, a comprehensive understanding might be necessary to elucidate the rules of their bioactivities. As science and technology advance in general, there will be a demand for a more sufficient understanding of the effects of these food components and Fenton reactions. More progress is expected in the near future.
Author Contributions
Conceptualization, C.A. and T.M. (Taiki Miyazawa); writing—original draft preparation, C.A.; writing—review and editing, T.M. (Taiki Miyazawa); supervision, T.M. (Teruo Miyazawa). All authors have read and agreed to the published version of the manuscript.
Funding
This research received no funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ozawa, H.; Miyazawa, T.; Burdeos, G.C.; Miyazawa, T. Biological Functions of Antioxidant Dipeptides. J. Nutr. Sci. Vitaminol. 2022, 68, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, S.; Kakinuma, G.; Kawashima, K.; Tanikawa, K.; Ochiai, S.; Miyata, R.; Fujisaki, K.; Kanao, R.; Noguchi, K.; Aoki, K.; et al. The Etiology of Beriberi. J. Am. Med. Assoc. 1929, 92, 1325–1327. [Google Scholar] [CrossRef]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory Redox Interactions. IUBMB Life 2019, 71, 430–441. [Google Scholar] [CrossRef]
- Poskitt, E.M.E. Early History of Iron Deficiency. Br. J. Haematol. 2003, 122, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Biswas, J.P.; Paul, S.; Paik, A.; Maiti, D. Organic Synthesis with the Most Abundant Transition Metal–Iron: From Rust to Multitasking Catalysts. Chem. Soc. Rev. 2021, 50, 243–472. [Google Scholar] [CrossRef]
- Andrews, N.C. Iron Metabolism: Iron Deficiency and Iron Overload. Annu. Rev. Genomics Hum. Genet. 2000, 1, 75–98. [Google Scholar] [CrossRef]
- Diaz-Ochoa, V.E.; Jellbauer, S.; Klaus, S.; Raffatellu, M. Transition Metal Ions at the Crossroads of Mucosal Immunity and Microbial Pathogenesis. Front. Cell. Infect. Microbiol. 2014, 4, 2. [Google Scholar] [CrossRef]
- Sheftel, A.D.; Mason, A.B.; Ponka, P. The Long History of Iron in the Universe and in Health and Disease. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 161–187. [Google Scholar] [CrossRef]
- Kell, D.B.; Heyden, E.L.; Pretorius, E. The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria. Front. Immunol. 2020, 11, 1221. [Google Scholar] [CrossRef]
- Jensen, F.B. The Dual Roles of Red Blood Cells in Tissue Oxygen Delivery: Oxygen Carriers and Regulators of Local Blood Flow. J. Exp. Biol. 2009, 212, 3387–3393. [Google Scholar] [CrossRef]
- Puig, S.; Ramos-Alonso, L.; Romero, A.M.; Martínez-Pastor, M.T. The Elemental Role of Iron in DNA Synthesis and Repair. Metallomics 2017, 9, 1483–1500. [Google Scholar] [CrossRef] [PubMed]
- Fenton, H.J.H. LXXIII.—Oxidation of Tartaric Acid in Presence of Iron. J. Chem. Soc. Trans. 1894, 65, 899–910. [Google Scholar] [CrossRef]
- Ruan, L.; Wang, M.; Zhou, M.; Lu, H.; Zhang, J.; Gao, J.; Chen, J.; Hu, Y. Doxorubicin–Metal Coordinated Micellar Nanoparticles for Intracellular Codelivery and Chemo/Chemodynamic Therapy In Vitro. ACS Appl. Bio Mater. 2019, 2, 4703–4707. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, M.; Cheng, J.; Yin, J.; Huang, C.; Cui, H.; Zhang, X.; Zhao, G. Acidity-Triggered Tumor-Targeted Nanosystem for Synergistic Therapy via a Cascade of ROS Generation and NO Release. ACS Appl. Mater. Interfaces 2020, 12, 28975–28984. [Google Scholar] [CrossRef]
- Yang, N.; Zhang, T.; Cao, C.; Mao, G.; Shao, J.; Song, X.; Wang, W.; Mou, X.; Dong, X. BSA Stabilized Photothermal-Fenton Reactor with Cisplatin for Chemo/Chemodynamic Cascade Oncotherapy. Nano Res. 2022, 15, 2235–2243. [Google Scholar] [CrossRef]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current Evidence on the Effect of Dietary Polyphenols Intake on Chronic Diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef]
- Duffuler, P.; Bhullar, K.S.; de Campos Zani, S.C.; Wu, J. Bioactive Peptides: From Basic Research to Clinical Trials and Commercialization. J. Agric. Food Chem. 2022, 70, 3585–3595. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.H.; Parihar, A.; Doseff, A.I. Dietary Flavonoids for Immunoregulation and Cancer: Food Design for Targeting Disease. Antioxidants 2019, 8, 202. [Google Scholar] [CrossRef]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Wagner, B.A.; Cramer-Morales, K.L.; Furqan, M.; Sandhu, S.; Carlisle, T.L.; Smith, M.C.; Abu Hejleh, T.; et al. O2− and H2O2-Mediated Disruption of Fe Metabolism Causes the Differential Susceptibility of NSCLC and GBM Cancer Cells to Pharmacological Ascorbate. Cancer Cell 2017, 31, 487–500. [Google Scholar] [CrossRef]
- Puppo, A. Effect of Flavonoids on Hydroxyl Radical Formation by Fenton-Type Reactions; Influence of the Iron Chelator. Phytochemistry 1992, 31, 85–88. [Google Scholar] [CrossRef]
- Xu, G.; Wang, H.; Li, X.; Huang, R.; Luo, L. Recent Progress on Targeting Ferroptosis for Cancer Therapy. Biochem. Pharmacol. 2021, 190, 114584. [Google Scholar] [CrossRef] [PubMed]
- Haber, F.; Willstätter, R. Unpaarigkeit Und Radikalketten Im Reaktionsmechanismus Organischer Und Enzymatischer Vorgänge. Ber. Dtsch. Chem. Ges. 1931, 64, 2844–2856. [Google Scholar] [CrossRef]
- Haber, F.; Weiss, J.; Pope, W.J. The Catalytic Decomposition of Hydrogen Peroxide by Iron Salts. Proc. R. Soc. London. Ser. A Math. Phys. Sci. 1934, 147, 332–351. [Google Scholar] [CrossRef]
- Stanbury, D.M. The Principle of Detailed Balancing, the Iron-Catalyzed Disproportionation of Hydrogen Peroxide, and the Fenton Reaction. Dalton Trans. 2022, 51, 2135–2157. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-F.; Chen, H.-F.; Kao, C.-L.; Chao, I.; Chen, H.-Y. A Computational Study of the Fenton Reaction in Different PH Ranges. Phys. Chem. Chem. Phys. 2018, 20, 22890–22901. [Google Scholar] [CrossRef] [PubMed]
- Kremer, M.L. The Fenton Reaction. Dependence of the Rate on PH. J. Phys. Chem. A 2003, 107, 1734–1741. [Google Scholar] [CrossRef]
- Chen, H.-Y. Why the Reactive Oxygen Species of the Fenton Reaction Switches from Oxoiron(IV) Species to Hydroxyl Radical in Phosphate Buffer Solutions? A Computational Rationale. ACS Omega 2019, 4, 14105–14113. [Google Scholar] [CrossRef]
- Shinichi, E.; Yosuke, S.; Colussi, A.J. Fenton Chemistry at Aqueous Interfaces. Proc. Natl. Acad. Sci. USA 2014, 111, 623–628. [Google Scholar] [CrossRef]
- Deguillaume, L.; Leriche, M.; Chaumerliac, N. Impact of Radical versus Non-Radical Pathway in the Fenton Chemistry on the Iron Redox Cycle in Clouds. Chemosphere 2005, 60, 718–724. [Google Scholar] [CrossRef]
- Prasad, R.; Majumdar, K.; Shivay, Y.; Kapil, U. Minerals in Plant and Human Nutrition and Health; International Plant Nutrition Institute (IPNI) Peachtree Corners: Peachtree Corners, GA, USA, 2016. [Google Scholar]
- Weed, R.I.; Reed, C.F.; Berg, G. Is Hemoglobin an Essential Structural Component of Human Erythrocyte Membranes? J. Clin. Investig. 1963, 42, 581–588. [Google Scholar] [CrossRef]
- Thomas, C.; Lumb, A.B. Physiology of Haemoglobin. Contin. Educ. Anaesth. Crit. Care Pain 2012, 12, 251–256. [Google Scholar] [CrossRef]
- Pasricha, S.-R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron Deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef]
- Laftah, A.H.; Latunde-Dada, G.O.; Fakih, S.; Hider, R.C.; Simpson, R.J.; McKie, A.T. Haem and Folate Transport by Proton-Coupled Folate Transporter/Haem Carrier Protein 1 (SLC46A1). Br. J. Nutr. 2008, 101, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and Characterization of a Mammalian Proton-Coupled Metal-Ion Transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The Iron Exporter Ferroportin/Slc40a1 Is Essential for Iron Homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef]
- Billesbølle, C.B.; Azumaya, C.M.; Kretsch, R.C.; Powers, A.S.; Gonen, S.; Schneider, S.; Arvedson, T.; Dror, R.O.; Cheng, Y.; Manglik, A. Structure of Hepcidin-Bound Ferroportin Reveals Iron Homeostatic Mechanisms. Nature 2020, 586, 807–811. [Google Scholar] [CrossRef]
- Camaschella, C. Iron Deficiency. Blood 2019, 133, 30–39. [Google Scholar] [CrossRef]
- Yiannikourides, A.; Latunde-Dada, G.O. A Short Review of Iron Metabolism and Pathophysiology of Iron Disorders. Medicines 2019, 6, 85. [Google Scholar] [CrossRef]
- Lesjak, M.; Srai, S.K.S. Role of Dietary Flavonoids in Iron Homeostasis. Pharmaceuticals 2019, 12, 119. [Google Scholar] [CrossRef]
- Lesjak, M.; Hoque, R.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.S.; Sharp, P.A. Quercetin Inhibits Intestinal Iron Absorption and Ferroportin Transporter Expression In Vivo and In Vitro. PLoS ONE 2014, 9, e102900. [Google Scholar] [CrossRef]
- Lazrak, M.; El Kari, K.; Stoffel, N.U.; Elammari, L.; Al-Jawaldeh, A.; Loechl, C.U.; Yahyane, A.; Barkat, A.; Zimmermann, M.B.; Aguenaou, H. Tea Consumption Reduces Iron Bioavailability from NaFeEDTA in Nonanemic Women and Women with Iron Deficiency Anemia: Stable Iron Isotope Studies in Morocco. J. Nutr. 2021, 151, 2714–2720. [Google Scholar] [CrossRef] [PubMed]
- Shubham, K.; Anukiruthika, T.; Dutta, S.; Kashyap, A.V.; Moses, J.A.; Anandharamakrishnan, C. Iron Deficiency Anemia: A Comprehensive Review on Iron Absorption, Bioavailability and Emerging Food Fortification Approaches. Trends Food Sci. Technol. 2020, 99, 58–75. [Google Scholar] [CrossRef]
- Sonnweber, T.; Ress, C.; Nairz, M.; Theurl, I.; Schroll, A.; Murphy, A.T.; Wroblewski, V.; Witcher, D.R.; Moser, P.; Ebenbichler, C.F.; et al. High-Fat Diet Causes Iron Deficiency via Hepcidin-Independent Reduction of Duodenal Iron Absorption. J. Nutr. Biochem. 2012, 23, 1600–1608. [Google Scholar] [CrossRef]
- Cepeda-Lopez, A.C.; Melse-Boonstra, A.; Zimmermann, M.B.; Herter-Aeberli, I. In Overweight and Obese Women, Dietary Iron Absorption Is Reduced and the Enhancement of Iron Absorption by Ascorbic Acid Is One-Half That in Normal-Weight Women. Am. J. Clin. Nutr. 2015, 102, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Villaño, D.; Vilaplana, C.; Medina, S.; Algaba-Chueca, F.; Cejuela-Anta, R.; Martínez-Sanz, J.M.; Ferreres, F.; Gil-Izquierdo, A. Relationship between the Ingestion of a Polyphenol-Rich Drink, Hepcidin Hormone, and Long-Term Training. Molecules 2016, 21, 1333. [Google Scholar] [CrossRef]
- Mu, M.; An, P.; Wu, Q.; Shen, X.; Shao, D.; Wang, H.; Zhang, Y.; Zhang, S.; Yao, H.; Min, J.; et al. The Dietary Flavonoid Myricetin Regulates Iron Homeostasis by Suppressing Hepcidin Expression. J. Nutr. Biochem. 2016, 30, 53–61. [Google Scholar] [CrossRef]
- Zaritsky, J.; Young, B.; Wang, H.-J.; Westerman, M.; Olbina, G.; Nemeth, E.; Ganz, T.; Rivera, S.; Nissenson, A.R.; Salusky, I.B. Hepcidin—A Potential Novel Biomarker for Iron Status in Chronic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Jian, J.; Katz, S.; Abramson, S.B.; Huang, X. 17β-Estradiol Inhibits Iron Hormone Hepcidin Through an Estrogen Responsive Element Half-Site. Endocrinology 2012, 153, 3170–3178. [Google Scholar] [CrossRef]
- Dasa, F.; Abera, T. Factors Affecting Iron Absorption and Mitigation Mechanisms: A Review. Int. J. Agric. Sci. Food Technol. 2018, 4, 24–30. [Google Scholar] [CrossRef]
- Diebold, L.; Chandel, N.S. Mitochondrial ROS Regulation of Proliferating Cells. Free Radic. Biol. Med. 2016, 100, 86–93. [Google Scholar] [CrossRef]
- Locato, V.; Cimini, S.; De Gara, L. ROS and Redox Balance as Multifaceted Players of Cross-Tolerance: Epigenetic and Retrograde Control of Gene Expression. J. Exp. Bot. 2018, 69, 3373–3391. [Google Scholar] [CrossRef] [PubMed]
- Qing, L.; Utta, B.-P.; Ulrike, M.; Martina, B.; Christoph, W.; Helmut, A.; Kurt, J.; Thomas, K. A Fenton Reaction at the Endoplasmic Reticulum Is Involved in the Redox Control of Hypoxia-Inducible Gene Expression. Proc. Natl. Acad. Sci. USA 2004, 101, 4302–4307. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, H.; Chai, Z.; Shi, W.; Li, X.; Ma, H. An Endoplasmic Reticulum-Targeting Fluorescent Probe for Imaging ˙OH in Living Cells. Chem. Commun. 2020, 56, 6344–6347. [Google Scholar] [CrossRef]
- Anelli, T.; Sitia, R. Protein Quality Control in the Early Secretory Pathway. EMBO J. 2008, 27, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Sung, G.; Kim, G.; Park, J.; Jin, B.; Kim, S.K. Effect of Various Intercalators on the Fenton-Type Oxidative Cleavage of Double-Stranded DNA. Chem. Asian J. 2014, 9, 1341–1348. [Google Scholar] [CrossRef]
- Imlay, J.A.; Chin, S.M.; Linn, S. Toxic DNA Damage by Hydrogen Peroxide Through the Fenton Reaction in Vivo and in Vitro. Science 1988, 240, 640–642. [Google Scholar] [CrossRef]
- Gutteridge, J.M.C. Iron Promoters of the Fenton Reaction and Lipid Peroxidation Can Be Released from Haemoglobin by Peroxides. FEBS Lett. 1986, 201, 291–295. [Google Scholar] [CrossRef]
- Miyazawa, T. Lipid Hydroperoxides in Nutrition, Health, and Diseases. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2021, 97, 161–196. [Google Scholar] [CrossRef]
- Thomas, D.D.; Michael, E.G.; Vitek, M.P.; Miranda, K.M.; Wink, D.A. Protein Nitration Is Mediated by Heme and Free Metals through Fenton-Type Chemistry: An Alternative to the NO/O2− Reaction. Proc. Natl. Acad. Sci. USA 2002, 99, 12691–12696. [Google Scholar] [CrossRef]
- Murakami, K.; Shimizu, T.; Irie, K. Formation of the 42-Mer and Amyloid β Radical and the Therapeutic Role of Superoxide Dismutase in Alzheimer’s Disease. J. Amino Acids 2011, 2011, 654207. [Google Scholar] [CrossRef][Green Version]
- Hachimoto, S.; Tatsuno, Y.; Kitagawa, T. Resonance Raman Evidence for the Presence of the FeIV=O Bond in Horseradish Peroxidase Compound II. Proc. Jpn. Acad. Ser. B 1984, 60, 345–348. [Google Scholar] [CrossRef]
- Carlomagno, F. Ferritinophagy and Cell Cycle Control. Blood 2016, 128, SCI-20. [Google Scholar] [CrossRef]
- Ma, S.; Fu, X.; Liu, L.; Liu, Y.; Feng, H.; Jiang, H.; Liu, X.; Liu, R.; Liang, Z.; Li, M.; et al. Iron-Dependent Autophagic Cell Death Induced by Radiation in MDA-MB-231 Breast Cancer Cells. Front. Cell Dev. Biol. 2021, 9, 723801. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Stockwell, B.R. Synthetic Lethal Screening Identifies Compounds Activating Iron-Dependent, Nonapoptotic Cell Death in Oncogenic-RAS-Harboring Cancer Cells. Chem. Biol. 2008, 15, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Chung, S.W. ROS-Mediated Autophagy Increases Intracellular Iron Levels and Ferroptosis by Ferritin and Transferrin Receptor Regulation. Cell Death Dis. 2019, 10, 822. [Google Scholar] [CrossRef]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and Function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef]
- Chen, X.; Li, J.; Kang, R.; Klionsky, D.J.; Tang, D. Ferroptosis: Machinery and Regulation. Autophagy 2021, 17, 2054–2081. [Google Scholar] [CrossRef]
- Bebber, C.M.; Müller, F.; Prieto Clemente, L.; Weber, J.; von Karstedt, S. Ferroptosis in Cancer Cell Biology. Cancers 2020, 12, 164. [Google Scholar] [CrossRef]
- Young, I.S.; Woodside, J.V. Antioxidants in Health and Disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, X.; Liu, M.; Cai, B.; He, N.; Wang, Z. Fenton Reaction-Based Nanomedicine in Cancer Chemodynamic and Synergistic Therapy. Appl. Mater. Today 2020, 21, 100864. [Google Scholar] [CrossRef]
- Ranji-Burachaloo, H.; Gurr, P.A.; Dunstan, D.E.; Qiao, G.G. Cancer Treatment through Nanoparticle-Facilitated Fenton Reaction. ACS Nano 2018, 12, 11819–11837. [Google Scholar] [CrossRef]
- Miyazawa, T.; Itaya, M.; Burdeos, G.C.; Nakagawa, K.; Miyazawa, T. A Critical Review of the Use of Surfactant-Coated Nanoparticles in Nanomedicine and Food Nanotechnology. Int. J. Nanomed. 2021, 16, 3937–3999. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Liu, X.-Y.; Zhou, T.-J.; Wan, X.; Wang, Y.; Jiang, H.-L. Photothermal Nanozyme-Ignited Fenton Reaction-Independent Ferroptosis for Breast Cancer Therapy. J. Control. Release 2021, 339, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; An, L.; Tian, Q.; Lin, J.; Yang, S. Ellagic Acid-Fe@BSA Nanoparticles for Endogenous H2S Accelerated Fe(III)/Fe(II) Conversion and Photothermal Synergistically Enhanced Chemodynamic Therapy. Theranostics 2020, 10, 4101–4115. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Cao, F.; Li, W.; Zhang, L.; You, Y.; Deng, Q.; Dong, K.; Ren, J.; Qu, X. Bioinspired Construction of a Nanozyme-Based H2O2 Homeostasis Disruptor for Intensive Chemodynamic Therapy. J. Am. Chem. Soc. 2020, 142, 5177–5183. [Google Scholar] [CrossRef]
- Gao, F.; Wang, F.; Nie, X.; Zhang, Z.; Chen, G.; Xia, L.; Wang, L.-H.; Wang, C.-H.; Hao, Z.-Y.; Zhang, W.-J.; et al. Mitochondria-Targeted Delivery and Light Controlled Release of Iron Prodrug and CO to Enhance Cancer Therapy by Ferroptosis. New J. Chem. 2020, 44, 3478–3486. [Google Scholar] [CrossRef]
- You, C.; Wu, H.; Gao, Z.; Chen, F.; Ning, L.; Zhang, Y.; Dong, Y.; Sun, B.; Wang, F. Enhanced Reactive Oxygen Species Levels by an Active Benzothiazole Complex-Mediated Fenton Reaction for Highly Effective Antitumor Therapy. Mol. Pharm. 2019, 16, 4929–4939. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, Q.; Zhang, M.; Sun, B.; She, Z.; Ge, M.; Lu, T.; Chu, X.; Wang, Y.; Wang, J.; et al. Hollow Porous Carbon Coated FeS2-Based Nanocatalysts for Multimodal Imaging-Guided Photothermal, Starvation, and Triple-Enhanced Chemodynamic Therapy of Cancer. ACS Appl. Mater. Interfaces 2020, 12, 10142–10155. [Google Scholar] [CrossRef]
- Fu, X.; Zhang, Y.; Zhang, G.; Li, X.; Ni, S.; Cui, J. Targeted Delivery of Fenton Reaction Packages and Drugs for Cancer Theranostics. Appl. Mater. Today 2022, 26, 101353. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, J.; Chen, Z.; Luo, Q.; Xu, J.; Song, G. Tumor-Specific Expansion of Oxidative Stress by Glutathione Depletion and Use of a Fenton Nanoagent for Enhanced Chemodynamic Therapy. ACS Appl. Mater. Interfaces 2019, 11, 30551–30565. [Google Scholar] [CrossRef]
- Sun, F.; Peng, Y.; Li, Y.; Xu, M.; Cai, T. Fenton-Reaction-Triggered Metabolism of Acetaminophen for Enhanced Cancer Therapy. Chin. Chem. Lett. 2022, in press. [Google Scholar] [CrossRef]
- Zhong, X.; Bao, X.; Zhong, H.; Zhou, Y.; Zhang, Z.; Lu, Y.; Dai, Q.; Yang, Q.; Ke, P.; Xia, Y.; et al. Mitochondrial Targeted Drug Delivery Combined with Manganese Catalyzed Fenton Reaction for the Treatment of Breast Cancer. Int. J. Pharm. 2022, 622, 121810. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Chen, L.; Lv, R.; Liu, M.; He, N.; Wang, Z. A Metal–Phenolic Network-Based Multifunctional Nanocomposite with PH-Responsive ROS Generation and Drug Release for Synergistic Chemodynamic/Photothermal/Chemo-Therapy. J. Mater. Chem. B 2020, 8, 2177–2188. [Google Scholar] [CrossRef]
- Lei, M.; Chen, G.; Zhang, M.; Lei, J.; Li, T.; Li, D.; Zheng, H. A PH-Sensitive Drug Delivery System Based on Hyaluronic Acid Co-Deliver Doxorubicin and Aminoferrocene for the Combined Application of Chemotherapy and Chemodynamic Therapy. Colloids Surf. B Biointerfaces 2021, 203, 111750. [Google Scholar] [CrossRef]
- Chi, H.; Zhu, G.; Yin, Y.; Diao, H.; Liu, Z.; Sun, S.; Guo, Z.; Xu, W.; Xu, J.; Cui, C.; et al. Dual-Responsive Multifunctional “Core–Shell” Magnetic Nanoparticles Promoting Fenton Reaction for Tumor Ferroptosis Therapy. Int. J. Pharm. 2022, 622, 121898. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yang, J.; Zhou, L.; Hu, X.; Wang, C.; Chai, K.; Li, R.; Feng, L.; Sun, Y.; Dong, C.; et al. Dual-Responsive and ROS-Augmented Nanoplatform for Chemo/Photodynamic/Chemodynamic Combination Therapy of Triple Negative Breast Cancer. ACS Appl. Mater. Interfaces 2022, 14, 57–68. [Google Scholar] [CrossRef]
- Jia, C.; Wu, H.; Luo, K.; Hao, W.; Wang, S.; Huang, M. Magnetic Silica Nanosystems With NIR-Responsive and Redox Reaction Capacity for Drug Delivery and Tumor Therapy. Front. Chem. 2020, 8, 567652. [Google Scholar] [CrossRef]
- Pal, S.; Jana, N.R. Pharmacologic Vitamin C-Based Cell Therapy via Iron Oxide Nanoparticle-Induced Intracellular Fenton Reaction. ACS Appl. Nano Mater. 2020, 3, 1683–1692. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Zhang, L.; Cheng, X.; Yu, H.; Bao, J.; Lu, R. Vitamin C Induces Ferroptosis in Anaplastic Thyroid Cancer Cells by Ferritinophagy Activation. Biochem. Biophys. Res. Commun. 2021, 551, 46–53. [Google Scholar] [CrossRef]
- Ouyang, Q.; Kou, F.; Zhang, N.; Lian, J.; Tu, G.; Fang, Z. Tea Polyphenols Promote Fenton-like Reaction: PH Self-Driving Chelation and Reduction Mechanism. Chem. Eng. J. 2019, 366, 514–522. [Google Scholar] [CrossRef]
- Cheng, R.; Li, G.; Fan, L.; Jiang, J.; Zhao, Y. Therapeutic Iminoboronate-Based Polymersomes with a Cu(II)-Mediated Fenton Reaction-Enhanced ROS-Response. Chem. Commun. 2020, 56, 12246–12249. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, H.; Liu, H.; Yuan, Q.; Ren, F.; Han, Y.; Sun, Q.; Li, Z.; Gao, M. Boosting H2O2-Guided Chemodynamic Therapy of Cancer by Enhancing Reaction Kinetics through Versatile Biomimetic Fenton Nanocatalysts and the Second Near-Infrared Light Irradiation. Adv. Funct. Mater. 2020, 30, 1906128. [Google Scholar] [CrossRef]
- Sun, M.; Yang, D.; Sun, Q.; Jia, T.; Kuang, Y.; Gai, S.; He, F.; Zhang, F.; Yang, P. A Porous Material Excited by Near-Infrared Light for Photo/Chemodynamic and Photothermal Dual-Mode Combination Therapy. J. Mater. Chem. B 2020, 8, 10559–10576. [Google Scholar] [CrossRef]
- Jomova, K.; Hudecova, L.; Lauro, P.; Simunková, M.; Barbierikova, Z.; Malcek, M.; Alwasel, S.H.; Alhazza, I.M.; Rhodes, C.J.; Valko, M. The Effect of Luteolin on DNA Damage Mediated by a Copper Catalyzed Fenton Reaction. J. Inorg. Biochem. 2022, 226, 111635. [Google Scholar] [CrossRef]
- Simunkova, M.; Barbierikova, Z.; Jomova, K.; Hudecova, L.; Lauro, P.; Alwasel, S.H.; Alhazza, I.; Rhodes, C.J.; Valko, M. Antioxidant vs. Prooxidant Properties of the Flavonoid, Kaempferol, in the Presence of Cu(II) Ions: A ROS-Scavenging Activity, Fenton Reaction and DNA Damage Study. Int. J. Mol. Sci. 2021, 22, 1619. [Google Scholar] [CrossRef]
- Perron, N.R.; Wang, H.C.; DeGuire, S.N.; Jenkins, M.; Lawson, M.; Brumaghim, J.L. Kinetics of Iron Oxidation upon Polyphenol Binding. Dalt. Trans. 2010, 39, 9982–9987. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Edwards, A.M.; Ruiz, M.; Silva, E.; Lissi, E. Lysozyme Modification by the Fenton Reaction and Gamma Radiation. Free Radic. Res. 2002, 36, 277–284. [Google Scholar] [CrossRef]
- Bochi, G.V.; Torbitz, V.D.; Santos, R.C.V.; Cubillos-Rojas, M.; López, J.L.R.; Siebel, A.M.; Gomes, P.; de Oliveira, J.R.; Moresco, R.N. Fenton Reaction-Generated Advanced Oxidation Protein Products Induces Inflammation in Human Embryonic Kidney Cells. Inflammation 2016, 39, 1285–1290. [Google Scholar] [CrossRef]
- Ishikawa, S.; Yano, Y.; Arihara, K.; Itoh, M. Egg Yolk Phosvitin Inhibits Hydroxyl Radical Formation from the Fenton Reaction. Biosci. Biotechnol. Biochem. 2004, 68, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Voltea, S.; Karabagias, I.K.; Roussis, I.G. Use of Fe (II) and H2O2 along with Heating for the Estimation of the Browning Susceptibility of White Wine. Appl. Sci. 2022, 12, 4422. [Google Scholar] [CrossRef]
- Gharib-Bibalan, S.; Keramat, J.; Hamdami, N. Better Lime Purification of Raw Sugar Beet Juice by Advanced Fenton Oxidation Process. Ozone Sci. Eng. 2018, 40, 54–63. [Google Scholar] [CrossRef]
- Blank, I.; Pascual, E.C.; Devaud, S.; Fay, L.B.; Stadler, R.H.; Yeretzian, C.; Goodman, B.A. Degradation of the Coffee Flavor Compound Furfuryl Mercaptan in Model Fenton-Type Reaction Systems. J. Agric. Food Chem. 2002, 50, 2356–2364. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Y.K.; Kang, Y.-R.; So, B.R.; Jung, S.K.; Chang, Y.H. Structural, Antioxidant, Prebiotic and Anti-Inflammatory Properties of Pectic Oligosaccharides Hydrolyzed from Okra Pectin by Fenton Reaction. Food Hydrocoll. 2021, 118, 106779. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, J.J.; Chen, L.; Peng, Z.; Zeng, Q.-H.; Zhu, Y.; Zhao, Y. Fenton Reaction-Assisted Photodynamic Inactivation of Calcined Melamine Sponge against Salmonella and Its Application. Food Res. Int. 2022, 151, 110847. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, C.K. A New Green Approach to Fenton’s Chemistry Using Tea Dregs and Coffee Grounds as Raw Material. Green Process. Synth. 2014, 3, 117–125. [Google Scholar] [CrossRef]
- Calhoun, L.N.; Kwon, Y.M. The Ferritin-like Protein Dps Protects Salmonella Enterica Serotype Enteritidis from the Fenton-Mediated Killing Mechanism of Bactericidal Antibiotics. Int. J. Antimicrob. Agents 2011, 37, 261–265. [Google Scholar] [CrossRef]
- Abbas, M.E.; Luo, W.; Zhu, L.; Zou, J.; Tang, H. Fluorometric Determination of Hydrogen Peroxide in Milk by Using a Fenton Reaction System. Food Chem. 2010, 120, 327–331. [Google Scholar] [CrossRef]
- Wang, S.; Fang, B.; Yuan, M.; Wang, Z.; Peng, J.; Lai, W. Dual-Mode Immunoassay System Based on Glucose Oxidase-Triggered Fenton Reaction for Qualitative and Quantitative Detection of Danofloxacin in Milk. J. Dairy Sci. 2020, 103, 7826–7833. [Google Scholar] [CrossRef]
- Wu, K.; Ying, W.; Shi, Z.; Yang, H.; Zheng, Z.; Zhang, J.; Yang, J. Fenton Reaction-Oxidized Bamboo Lignin Surface and Structural Modification to Reduce Nonproductive Cellulase Binding and Improve Enzyme Digestion of Cellulose. ACS Sustain. Chem. Eng. 2018, 6, 3853–3861. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, M.-J. Enhancing Enzymolysis and Fermentation Efficiency of Sugarcane Bagasse by Synergistic Pretreatment of Fenton Reaction and Sodium Hydroxide Extraction. Bioresour. Technol. 2016, 214, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Midassi, S.; Bedoui, A.; Bensalah, N. Efficient Degradation of Chloroquine Drug by Electro-Fenton Oxidation: Effects of Operating Conditions and Degradation Mechanism. Chemosphere 2020, 260, 127558. [Google Scholar] [CrossRef]
- Marchetti, M.D.; Bessa Azevedo, E. Degradation of NSAIDs by Optimized Photo-Fenton Process Using UV-LEDs at near-Neutral PH. J. Water Process Eng. 2020, 35, 101171. [Google Scholar] [CrossRef]
- Barros, W.R.P.; Franco, P.C.; Steter, J.R.; Rocha, R.S.; Lanza, M.R.V. Electro-Fenton Degradation of the Food Dye Amaranth Using a Gas Diffusion Electrode Modified with Cobalt (II) Phthalocyanine. J. Electroanal. Chem. 2014, 722–723, 46–53. [Google Scholar] [CrossRef]
- Shin, J.; Bae, S.; Chon, K. Fenton Oxidation of Synthetic Food Dyes by Fe-Embedded Coffee Biochar Catalysts Prepared at Different Pyrolysis Temperatures: A Mechanism Study. Chem. Eng. J. 2021, 421, 129943. [Google Scholar] [CrossRef]
- Chen, Q.; Espey, M.G.; Sun, A.Y.; Pooput, C.; Kirk, K.L.; Krishna, M.C.; Khosh, D.B.; Drisko, J.; Levine, M. Pharmacologic Doses of Ascorbate Act as a Prooxidant and Decrease Growth of Aggressive Tumor Xenografts in Mice. Proc. Natl. Acad. Sci. USA 2008, 105, 11105–11109. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.; Van Riper, J.M.; Cantley, L.C.; Yun, J. Targeting Cancer Vulnerabilities with High-Dose Vitamin C. Nat. Rev. Cancer 2019, 19, 271–282. [Google Scholar] [CrossRef]
- Yun, J.; Mullarky, E.; Lu, C.; Bosch, K.N.; Kavalier, A.; Rivera, K.; Roper, J.; Chio, C.I.I.; Giannopoulou, E.G.; Rago, C.; et al. Vitamin C Selectively Kills KRAS and BRAF Mutant Colorectal Cancer Cells by Targeting GAPDH. Science 2015, 350, 1391–1396. [Google Scholar] [CrossRef]
- Abe, C.; Higuchi, O.; Matsumoto, A.; Miyazawa, T. Determination of Intracellular Ascorbic Acid Using Tandem Mass Spectrometry. Analyst 2022, 147, 2640–2643. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).