Effect of Nano-Fertilizers on Alfalfa Plants Grown under Different Salt Stresses in Hydroponic System

 , , ,
, , , 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Salt Stress Response
2.1.1. Proline
2.1.2. Electrolyte Leakage
2.2. Antioxidant Enzymes
2.2.1. Catalase (CAT, EC 1.11.1.6)
2.2.2. Superoxide Dismutase (SOD, EC 1.15.1.1)
2.3. Statistical Analysis
3. Results
3.1. Plant Biomass
3.2. Physiological Effect
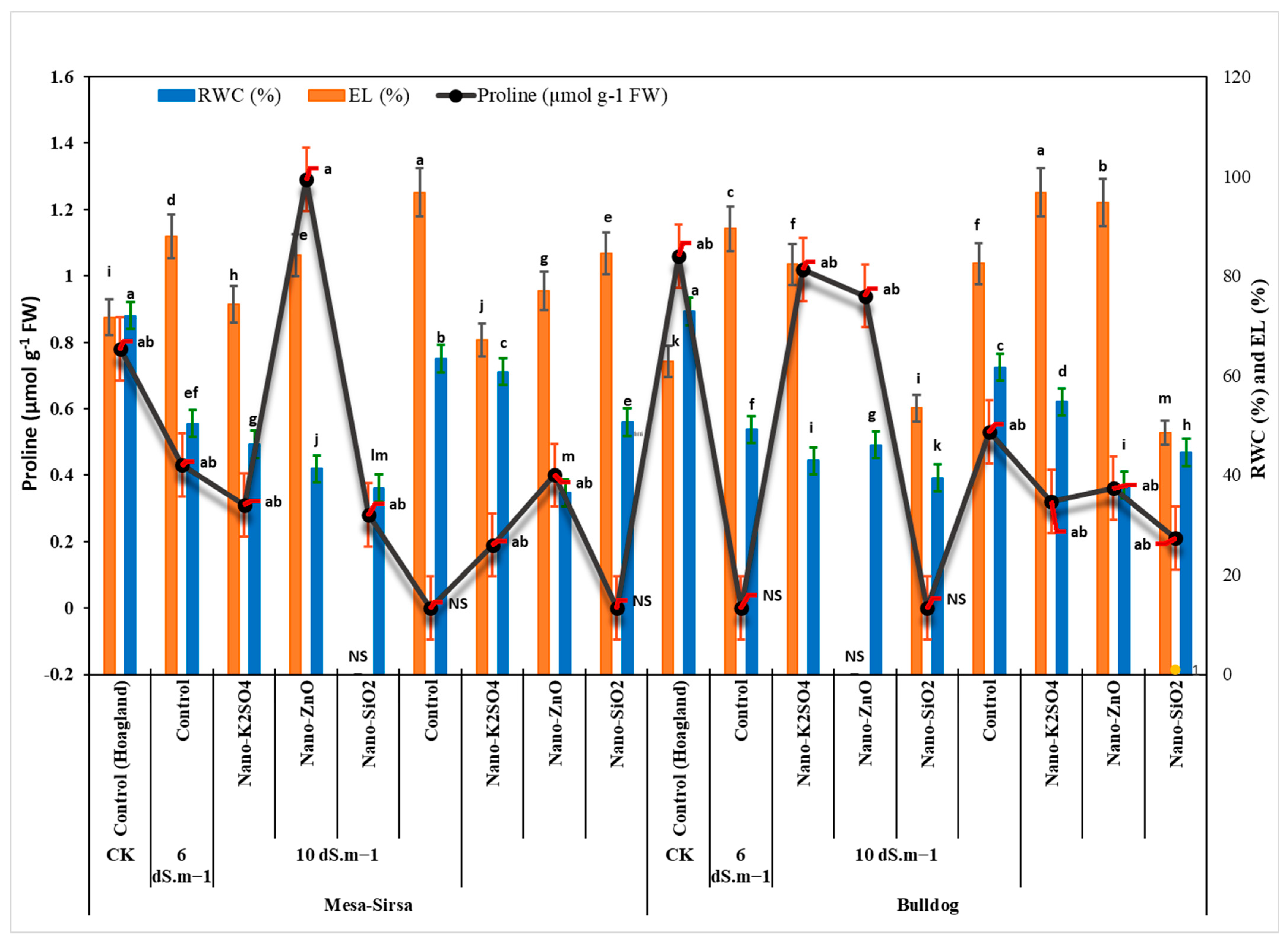
3.2.1. Relative Water Content (RWC)
3.2.2. Electrolyte Leakage (EL)
3.2.3. Proline
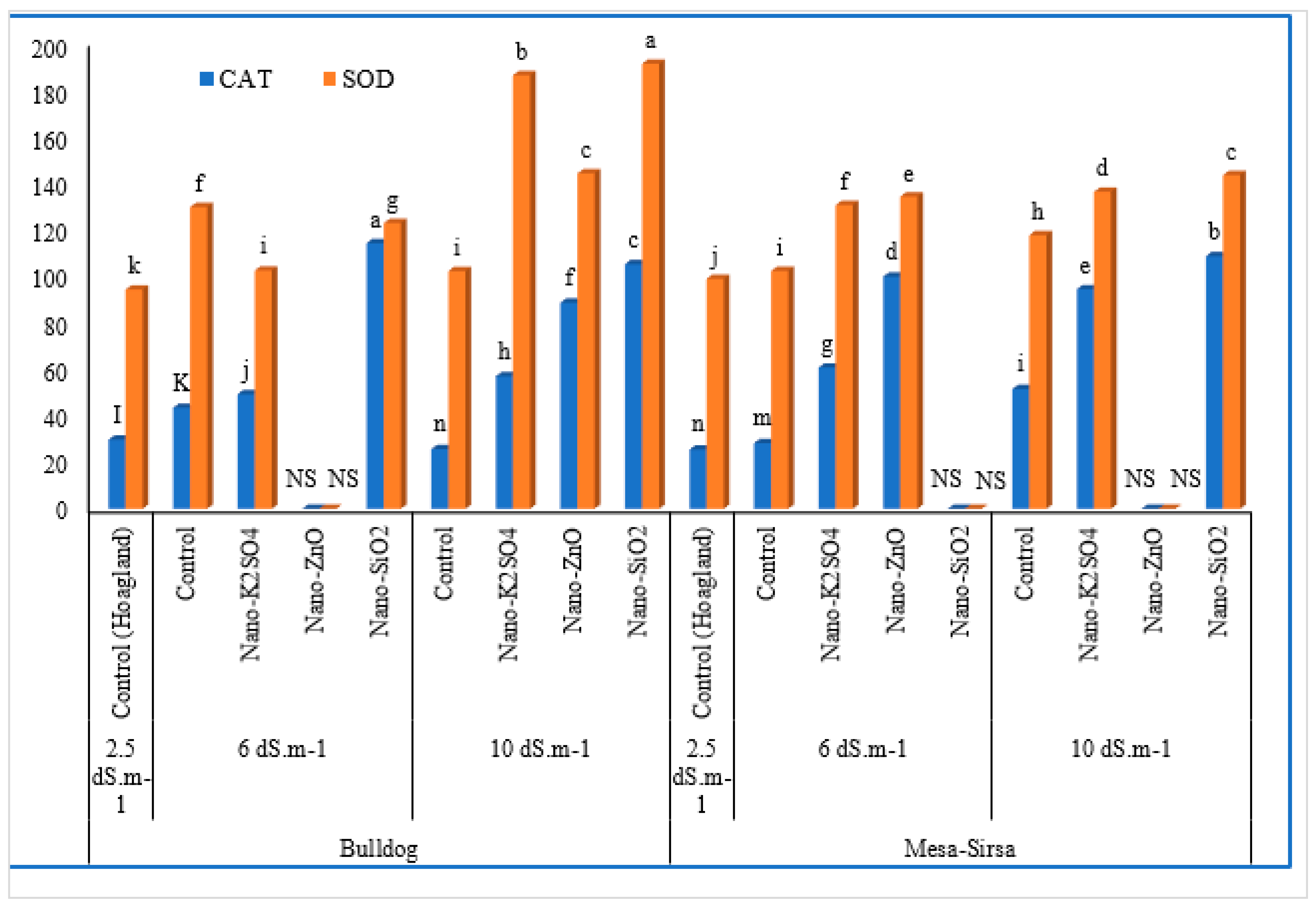
3.3. Antioxidant Enzymes
3.4. Plant Tissue Chemical Characteristics
4. Discussion
4.1. Plant Biomass
4.2. Physiological Effect
4.2.1. Relative Water Content
4.2.2. Electrolyte Leakage (EL)
4.2.3. Proline
4.3. Antioxidant Enzymes
4.4. Plant Tissue Chemical Characteristics
4.5. Correlation between Physiological and Phenotypic Responses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mora, O.; Le Mouël, C.; De Lattre-Gasquet, M.; Donnars, C.; Dumas, P.; Réchauchère, O.; Brunelle, T.; Manceron, S.; Marajo-Petitzon, E.; Moreau, C.; et al. Exploring the future of land use and food security: A new set of global scenarios. PLoS ONE 2020, 15, e0235597. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Cao, Y.; Gao, Y.; Li, J.; Tian, Y. Straw composts, gypsum and their mixtures enhance tomato yields under continuous saline water irrigation. Agric. Water Manag. 2019, 223, 105721. [Google Scholar] [CrossRef]
- Li, J.; Gao, Y.; Zhang, X.; Tian, P.; Li, J.; Tian, Y. Comprehensive comparison of different saline water irrigation strategies for tomato production: Soil properties, plant growth, fruit yield and fruit quality. Agric. Water Manag. 2018, 213, 521–533. [Google Scholar] [CrossRef]
- Jalali, V.; Kapourchal, S.A.; Homaee, M. Evaluating performance of macroscopic water uptake models at productive growth stages of durum wheat under saline conditions. Agric. Water Manag. 2017, 180, 13–21. [Google Scholar] [CrossRef]
- Yu, P.; Liu, S.; Yang, H.; Fan, G.; Zhou, D. Short-term land use conversions influence the profile distribution of soil salinity and sodicity in northeastern China. Ecol. Indic. 2018, 88, 79–87. [Google Scholar] [CrossRef]
- Badawy, E.M.; Khalid, K.A.; Heikal, A.A.M.; Nagdy, M.M. Effect of Salinity tress and Soil Types on Growth, Photosynthetic Pigments and Essential Oil of Artemisia annua L. Asian J. Crop Sci. 2018, 10, 40–47. [Google Scholar]
- Bertrand, A.; Dhont, C.; Bipfubusa, M.; Chalifour, F.P.; Drouin, P.; Beauchamp, C.J. Improving salt stress responses of the symbiosis in alfalfa using salt-tolerant cultivar and rhizobial strain. Appl. Soil Ecol. 2015, 87, 108–117. [Google Scholar] [CrossRef]
- FAO. Crop Water Management Alfalfa; FAO: Rome, Italy, 2002; Available online: http://www.fao.Org/AG/aglW/cropwater/alfalfa.stm (accessed on 1 January 2022).
- Lu, Q.; Ge, G.; Sa, D.; Wang, Z.; Hou, M.; Jia, Y.S. Effects of salt stress levels on nutritional quality and microorganisms of alfalfa-influenced soil. PeerJ 2021, 9, e11729. [Google Scholar] [CrossRef]
- Shan, W.X.; Long, R.H.; Wu, W.Z.; Wen, W.Y.; Bo, W.W. Effects of neutral salt and alkali on ion distributions in the roots, shoots, and leaves of two alfalfa cultivars with differing degrees of salt tolerance. J. Integr. Agric. 2017, 16, 1800–1807. [Google Scholar]
- Bhattarai, S.; Biswas, D.; Fu, Y.-B.; Biligetu, B. Morphological, Physiological, and Genetic Responses to Salt Stress in Alfalfa: A Review. Agronomy 2020, 10, 577. [Google Scholar] [CrossRef]
- Emam, Y.; Bijanzadeh, E.; Naderi, R.; Edalat, M. Effect of salt stress on vegetative growth and ion accumulation of two alfalfa (Medicago sativa L.) cultivars. Desert 2009, 14, 163–169. [Google Scholar]
- Shavrukov, Y.; Yusuf, G.; Julie, H. The use of hydroponics in abiotic stress tolerance research. In Hydroponics: A Standard Methodology for Plant Biological Researches; Asao, T., Ed.; InTech: Vienna, Austria, 2012; ISBN 978-953-51-0386-8. Available online: http://www.intechopen.com/books/hydroponics-a-standardmethodology-for-plantbiological-researches/the-use-of-hydroponics-in-abiotic-stress-tolerance-research (accessed on 1 January 2022).
- Cifuentes-Torres, L.; Mendoza-Espinosa, L.G.; Correa-Reyes, G.; Daesslé, L.W. Hydroponics with wastewater: A review of trends and opportunities. Water Environ. J. 2020, 35, 166–180. [Google Scholar] [CrossRef]
- Trejo-Téllez, L.I.; Gómez-Merino, F.C. Nutrient solutions for hydroponic systems. In Hydroponics: A Standard Methodology for Plant Biological Researches; InTech: Vienna, Austria, 2012; pp. 1–22. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. In Circular, 2nd ed.; California Agricultural Experiment Station: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lai, R. Synthetic apatite Nanoparticles as a phosphorus fertilizer for soybean (Glycine max). Sci. Rep. 2015, 4, 5686–5691. [Google Scholar] [CrossRef] [PubMed]
- Benzone, H.R.L.; Rubenecia, M.R.U.; Ultra, V.U.; Lee, S.C. Nanoparticles fertilizer affects the growth, development, and chemical properties of rice. Int. J. Agron. Agric. Res. 2015, 7, 105–117. [Google Scholar]
- Yaseen, R.; IS Ahmed, A.; M Omer, A.; KM Agha, M.; M Emam, T. Nano-fertilizers: Bio-fabrication, application and biosafety. Nov. Res. Microbiol. J. 2020, 4, 884–900. [Google Scholar]
- Laware, S.L.; Raskar, S.V. Influence of Zinc Oxide Nanoparticles on Growth, Flowering and Seed Productivity in Onion. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 874–881. [Google Scholar]
- Lin, D.H.; Xing, B.S. Root Uptake and Phytotoxicity of ZnO Nanoparticles. Environ. Sci. Technol. 2008, 42, 5580–5585. [Google Scholar] [CrossRef]
- Stampoulis, D.; Sinha, S.K.; White, J.C. Assay-Dependent Phytotoxicity of Nanoparticles to Plants. Environ. Sci. Technol. 2009, 43, 9473–9479. [Google Scholar] [CrossRef]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Prabu, P.; Rajendran, V.; Kannan, N. Growth and physiological responses of maize (Zea mays L.) to porous silica nanoparticles in soil. J. NanoPart. Res. 2012, 14, 1294. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress onCucurbita pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.G.; Abdelhamid, M.T.; Schmidhalter, U. Potassium fertiliser enhances the salt-tolerance of common bean (Phaseolus vulgaris L.). J. Hortic. Sci. Biotechnol. 2014, 89, 185–192. [Google Scholar] [CrossRef]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Kong, X.; Li, W.; Tang, W.; Zhang, D. Effects of plant density and nitrogen and potassium fertilization on cotton yield and uptake of major nutrients in two fields with varying fertility. Field Crop. Res. 2010, 119, 106–113. [Google Scholar] [CrossRef]
- Li, M.; Zhang, P.; Adeel, M.; Guo, Z.; Chetwynd, A.J.; Ma, C.; Bai, T.; Hao, Y.; Rui, Y. Physiological impacts of zero valent iron, Fe3O4 and Fe2O3 nanoparticles in rice plants and their potential as Fe fertilizers. Environ. Pollut. 2020, 269, 116134. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Ma, C.; Wang, K.; Hao, Y.; Chen, Q.; Mo, Y.; Rui, Y. Effects of cerium oxide on rice seedlings as affected by co-exposure of cadmium and salt. Environ. Pollut. 2019, 252, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, M.S.; El-Beshsbeshy, T.R.; Mahmoud, E.K.; Abdelkader, N.I.; Al-Shal, R.M.; Missaoui, A.M. Response of Alfalfa under Salt Stress to the Application of Potassium Sulfate Nanoparticles. Am. J. Plant Sci. 2017, 8, 1751–1773. [Google Scholar] [CrossRef]
- Turner, N.C. Techniques and experimental approaches for the measurement of plant water status. Plant Soil 1981, 58, 339–366. [Google Scholar] [CrossRef]
- Isla, R.; Aragüés, R. Response of alfalfa (Medicago sativa L.) to diurnal and nocturnal saline sprinkler irrigations. II: Shoot ion content and yield relationships. Irrig. Sci. 2009, 27, 507–513. [Google Scholar] [CrossRef][Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Gawehn, K. Methoden der Enzymatischen Analyse, 2nd ed.; Bergmeyer, H.U., Ed.; Verlag Chemie Weinheim: Weinheim, Germany, 1970; pp. 388–483. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef]
- Karimi, H.R.; Maleki, K.R. Evaluation of inter-specific hybrid of P. atlantica × P. vera cv. ‘Badami-Rize-Zarand’ as pistachio rootstock to salinity stress according to some growth indices, echo physiological and biochemical parameter. J. Stress Physiol. Biochem. 2014, 10, 5–17. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant growth under water/salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef]
- Shivay, Y.S.; Chen, J.H.; Ding, S. Effects of potassium on morphological and physiological parameters in barley (Hordeum vulgaris L.) genotype. Indian J. Plant Physiol. 2002, 7, 146–151. [Google Scholar]
- Clark, L.J.; Whalley, W.E.; Ellis-Jones, J.; Dent, K.; Rowse, H.R.; Finch- Savage, W.E.; Gatsai, T.; Jasi, L.; Kaseke, N.E.; Murungu, F.S.; et al. On farm seed priming in maize: A physiological evaluation. In Proceedings of the Seventh Eastern and Southern Africa Regional Maize Conference, Nairobi, Kenya, 5–11 February 2002; pp. 268–273. [Google Scholar]
- Raliya, R.; Tarafdar, J.C. ZnO nanoparticle biosynthesis and its effect on phosphorous-mobilizing enzyme secretion and gum contents in cluster bean (Cyamopsis tetragonoloba L.). Agric Res. 2013, 2, 48–57. [Google Scholar] [CrossRef]
- Raliya, R.; Nair, R.; Chavalmane, S.; Wang, W.-N.; Biswas, P. Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 2015, 7, 1584–1594. [Google Scholar] [CrossRef]
- Quary, F.X.; Leenhardt, F.; Remesy, C. Genetic variability and stability of grain Mg, Zn and Fe concentration in bread wheat. Eur. J. Agron. 2006, 25, 177–185. [Google Scholar]
- Hussein, M.M.; Abou-Baker, N.H. The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 2018, 5, 171809. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Shome, S.; Tewari, S.; Bhattacharya, M.K.; Panda, S.K. Effect of Zn nano-particles on growth responses of rice. In Nanotechnology: Novel Perspectives and Prospects; Singh, B., Kaushik, A., Mehta, S.K., Tripathi, S.K., Eds.; McGraw Hill Education: New Delhi, India, 2015; pp. 508–512. [Google Scholar]
- Babaei, K.; Sharifi, R.S.; Pirzad, A.; Khalilzadeh, R. Effects of bio fertilizer and nano Zn-Fe oxide on physiological traits, antioxidant enzymes activity and yield of wheat (Triticum aestivum L.) under salinity stress. J. Plant Interact. 2017, 12, 381–389. [Google Scholar] [CrossRef]
- Torabian, S.; Zahedi, M.; Khoshgoftarmanesh, A. Effect of foliar spray of zinc oxide on some antioxidant enzymes activity of sunflower under salt stress. J. Agric. Sci. Technol. 2016, 18, 1013–1025. [Google Scholar]
- González, L.; González-vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Reigosa, M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 207–212. [Google Scholar]
- Saeed, R.; Salma, M.; Rafiq, A. Electrolyte leakage and relative water content as affected by organic mulch in okra (Abelmoschus esculentus L.) moench grown under salinity. FUUAST J. Biol. 2014, 4, 221. [Google Scholar]
- Wrobel, J.; Mikiciuk, M. Water and ionic balance in the leaves of basket willow (Salix viminalis L.) cultivated in hydroponics with different salinity levels. Ecol. Chem. Eng. A 2010, 17, 1315–1321. [Google Scholar]
- Marschner, P.M. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–189. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40 (Suppl. 1), 326–345. [Google Scholar] [CrossRef]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ Nutrition in Plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.J.; A Leigh, R.; Miller, A.J. Potassium homeostasis in vacuolate plant cells. Proc. Natl. Acad. Sci. USA 1996, 93, 10510–10514. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica Nanoparticles on basil (Ocimum basilicum) under salinity stress. J. Chem. Health Risks 2014, 4, 49–55. [Google Scholar]
- Ashraf, M.; Ali, Q. Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ. Exp. Bot. 2008, 63, 266–273. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Munshi, A.D.; Kumar, R.; Pandey, R.N.; Arora, A.; Bhat, J.S.; Sureja, A.K. Effect of salt stress on cucumber: Na+–K+ ratio, osmolyte concentration, phenols and chlorophyll content. Acta Physiol. Plant. 2009, 32, 103–114. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Salama, K.H. Cellular basis of salinity tolerance in plants. Environ. Exp. Bot. 2004, 52, 113–122. [Google Scholar] [CrossRef]
- Agarie, S.; Hanaoka, N.; Ueno, O.; Miyazaki, A.; Kubota, F.; Agata, W.; Kaufman, P.B. Effects of silicon on tolerance to water deficit and heat stress in rice plants (Oryza sativa L.), monitored by electrolyte leakage. Plant Prod. Sci. 1998, 1, 96–103. [Google Scholar] [CrossRef]
- Liu, B.; Soundararajan, P.; Manivannan, A. Mechanisms of Silicon-Mediated Amelioration of Salt Stress in Plants. Plants 2019, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Tuna, L.; Higgs, D. Effect of Silicon on Plant Growth and Mineral Nutrition of Maize Grown under Water-Stress Conditions. J. Plant Nutr. 2006, 29, 1469–1480. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K.; Chen, G.; Wang, S.-M.; Zhang, C. Effects of Silicon on Growth of Wheat under Drought. J. Plant Nutr. 2003, 26, 1055–1063. [Google Scholar] [CrossRef]
- Gong, H.; Zhu, X.; Chen, K.; Wang, S.; Zhang, C. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 2005, 169, 313–321. [Google Scholar] [CrossRef]
- Eneji, E.; Inanaga, S.; Muranaka, S.; Li, J.; An, P.; Hattori, T.; Tsuji, W. Effect of calcium silicate on growth and dry matter yield of Chloris gayana and Sorghum sudanense under two soil water regimes. Grass Forage Sci. 2005, 60, 393–398. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Mazrou, Y.S.; Hafez, Y.M. Silicon Foliar Application Mitigates Salt Stress in Sweet Pepper Plants by Enhancing Water Status, Photosynthesis, Antioxidant Enzyme Activity and Fruit Yield. Plants 2020, 9, 733. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, M.S.; El-Beshbeshy, T.R.; Hassan, S.M.; Mahmoud, E.K.; Abdelkader, N.I.; Al-Shal, R.M.; Missaoui, A.M. Alleviating Salt Stress in Barley by Use of Plant Growth Stimulants and Potassium Sulfate. J. Agric. Sci. 2017, 9, 136. [Google Scholar] [CrossRef][Green Version]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide 2012, 27, 210–218. [Google Scholar] [CrossRef]
- Yu Lei, M.; Shaozheng, L. Research on salt tolerance of some tree species on muddy seashore of north China. Int. Semin. Prospect. Saline Agric. 2000, 5, 10–12. [Google Scholar]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- El-Metwally, I.M.; Doaa, M.R.; Abo-Basha, A.E.A.M.; Abd El-Aziz, M. Response of peanut plants to different foliar applications of nano-iron, manganese and zinc under sandy soil conditions. Middle East J. Appl. Sci. 2018, 8, 474–482. [Google Scholar]
- Safyan, N.; Reza, M.; Darbaghshahi, N.; Bahari, B. The effect of microelements spraying on growth, qualitative and quantitative grain corn in Iran. Int. Res. J. Basic Appl. Sci. 2012, 8, 34–57. [Google Scholar]
- Janmohammadi, M.; Amanzadeh, T.; Sabaghnia, N.; Dashti, S. Impact of foliar application of nano micronutrient fertilizers and titanium dioxide nanoparticles on the growth and yield components of barley under supplemental irrigation. Acta Agric. Slov. 2016, 107, 265–276. [Google Scholar] [CrossRef]
- Hertwig, B.; Streb, P.; Feierabend, J. Light Dependence of Catalase Synthesis and Degradation in Leaves and the Influence of Interfering Stress Conditions. Plant Physiol. 1992, 100, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, M.; Afifipour, Z.; Mozafarian, M. The effect of N-Si on tomato seed germination under salinity levels. J. Biol. Environ. Sci. 2012, 6, 87–90. [Google Scholar]
- Li, B.; Tao, G.; Xie, Y.; Cai, X. Physiological effects under the condition of spraying Nano SiO2 onto the Indocalamus barbatus McClure leaves. J. Nanjing Univ. (Nat. Sci. Ed.) 2012, 36, 161–164. [Google Scholar]
- Prakash, M.G.; Chung, I.M. Determination of zinc oxide nanoparticles toxicity in root growth in wheat (Triticum aestivum L.) seedlings. Acta Biol. Hung. 2016, 67, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, P.; Priyanka, N.; Manikandan, K.; Ganeshbabu, I.; Indiraarulselvi, P.; Geetha, N.; Muralikrishna, K.; Bhattacharya, R.; Tiwari, M.; Sharma, N.; et al. Enhanced plant growth promoting role of phycomolecules coated zinc oxide nanoparticles with P supplementation in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 110, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.S. (Ed.) Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Springer: Berlin, Germany, 2019; Volume 1. [Google Scholar] [CrossRef]
- Bala, R.; Kalia, A.; Dhaliwal, S.S. Evaluation of Efficacy of ZnO Nanoparticles as Remedial Zinc Nanofertilizer for Rice. J. Soil Sci. Plant Nutr. 2019, 19, 379–389. [Google Scholar] [CrossRef]
- Giordano, P.M.; Noggle, J.C.; Mortvedt, J.J. Zinc uptake by rice, as affected by metabolic inhibitors and competing cations. Plant Soil 1974, 41, 637–646. [Google Scholar] [CrossRef]
- Malvi, U. Interaction of micronutrients with major nutrients with special reference to potassium. Karnataka J. Agric. Sci. 2011, 24, 106–109. [Google Scholar]
- Sabir, A.; Yazar, K.; Sabir, F.; Kara, Z.; Yazici, M.A.; Goksu, N. Vine growth, yield, berry quality attributes and leaf nutrient content of grapevines as influenced by seaweed extract (Ascophyllum nodosum) and nanosize fertilizer pulverizations. Sci. Hortic. 2014, 175, 1–8. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Interactive effect of calcium and gibberellin on nickel tolerance in relation to antioxidant systems in Triticum aestivum L. Protoplasma 2010, 248, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Mohammad, F.; Khan, M.M.A.; Al-Whaibi, M.H. Cumulative effect of nitrogen and sulphur on Brassica juncea L. genotypes under NaCl stress. Protoplasma 2011, 249, 139–153. [Google Scholar] [CrossRef]
- Lynch, J.P.; Ho, M.D.; Phosphorus, L. Rhizoeconomics: Carbon costs of phosphorus acquisition. Plant Soil 2005, 269, 45–56. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genotype | Salt Conc. | Treatments | Shoot Dry Weight (gm) | Plant Height (cm) | No. of Flowers | No. of Tillers | Root Length (cm) | Root Fresh Weight (gm) | Root Dry Weight (gm) |
|---|---|---|---|---|---|---|---|---|---|
| Bulldog | 2.5 dS m−1 | Control (Hoagland) | 2.68 ± 0.31 b | 44.69 ± 4.82 b | 12.0 ± 0.01 cd | 3.67 ± 0.6 cd | 35.08 ± 14.38 f | 16.65 ± 1.34 f | 1.69 ± 0.11 bc |
| 6 dS m−1 | Control | 1.20 ± 0.1 cde | 40.23 ± 2.56 d | 12.3 ± 4.16 c | 4.0 ± 1.0 bc | 33.48 ± 1.96 g | 12.25 ± 6.97 g | 1.31 ± 0.6 bcdef | |
| Nano-K2SO4 | 2.55 ± 0.5 b | 22.89 ± 2.34 i | 1.0 ± 0.0 ij | 3.0 ± 0.0 cdef | 26.66 ± 1.04 k | 33.84 ± 14.6 a | 3.73 ± 1.51 a | ||
| Nano-ZnO | 0.54 ± 0.13 de | 10.97 ± 1.95 n | 0.0 ± 0.0 j | 2.00 ± 0.0 f | 26.1 ± 6.26 k | 20.2 ± 0.12 d | 3.65 ± 0.59 a | ||
| Nano-SiO2 | 0.20 ± 0.08 e | 17.31 ± 2.14 k | 0.0 ± 0.0 j | 3.3 ± 0.57 cde | 33.09 ± 9.96 gh | 10.36 ± 2.1 h | 1.65 ± 0.3 bcd | ||
| 10 dS m−1 | Control | 1.2 ± 0.1 cde | 35.74 ± 3.89 f | 5.3 ± 1.15 h | 4.0 ± 0.0 bc | 34.79 ± 8.06 f | 16.68 ± 5.74 f | 1.45 ± 0.43 bcde | |
| Nano-K2SO4 | 6.30 ± 0.3 a | 48.06 ± 2.97 a | 20.0 ± 8.89 a | 6.0 ± 1.0 a | 40.17 ± 12.09 d | 28.00 ± 0.34 b | 0.38 ± 0.04 def | ||
| Nano-ZnO | 0.36 ± 0.04 de | 18.15 ± 5.17 k | 2.0 ± 0.0 i | 2.7 ± 0.6 def | 30.47 ± 1.85 i | 10.7 ± 0.04 h | 0.12 ± 0.04 f | ||
| Nano-SiO2 | 2.3 ± 0.07 bc | 43.36 ± 1.55 c | 10.3 ± 4.5 e | 4.0 ± 0.0 bc | 55.42 ± 8.87 b | 18.9 ± 0.195 e | 0.24 ± 0.02 ef | ||
| Mesa-Sirsa | 2.5 dS m−1 | Control (Hoagland) | 2.16 ± 0.08 bc | 32.13 ± 4.07 g | 12.0 ± 0.0 cd | 3.3 ± 0.6 cde | 51.67 ± 5.93 c | 16.7 ± 4.2 f | 1.34 ± 0.01 bcdef |
| 6 dS m−1 | Control | 0.49 ± 0.08 de | 9.12 ± 0.55 o | 0.0 ± 0.0 j | 2.3 ± 0.6 ef | 20.36 ± 8.46 m | 10.4 ± 0.09 h | 0.14 ± 0.01 f | |
| Nano-K2SO4 | 2.23 ± 0.12 bc | 38.79 ± 0.78 e | 1.0 ± 0.0 ij | 5.0 ± 0.0 ab | 32.19 ± 1.29 h | 21.04 ± 3.97 d | 2.26 ± 0.09 b | ||
| Nano-ZnO | 1.43 ± 0.13 bcde | 28.64 ± 5.44 h | 6.7 ± 2.5 g | 4.0 ± 1.0 bc | 37.92 ± 5.54 e | 13.44 ± 1.03 g | 1.06 ± 0.02 bcdef | ||
| Nano-SiO2 | 1.5 ± 0.13 bcd | 33.07 ± 5.01 g | 0.0 ± 0.0 j | 5.7 ± 0.6 a | 27.21 ± 1.94 jk | 9.55 ± 0.15 h | 0.28 ± 0.07 ef | ||
| 10 dS m−1 | Control | 0.25 ± 0.00 de | 12.96 ± 3.76 m | 8.0 ± 3.0 f | 2.0 ± 0.0 f | 28.31 ± 5.0 j | 6.34 ± 0.43 i | 0.9 ± 0.2 cdef | |
| Nano-K2SO4 | 0.36 ± 0.04 de | 29.93 ± 2.45 j | 16.7 ± 9.8 b | 4.0 ± 0.0 bc | 29.71 ± 6.73 i | 18.2 ± 0.08 e | 0.21 ± 0.01 ef | ||
| Nano-ZnO | 0.87 ± 0.15 de | 21.05 ± 3.52 j | 0.0 ± 0.0 j | 2.3 ± 0.6 ef | 23.07 ± 9.64 l | 24.0 ± 0.28 c | 0.37 ± 0.03 ef | ||
| Nano-SiO2 | 0.41 ± 0.08 de | 14.33 ± 1.8 i | 10.7 ± 3.1 de | 2.0 ± 0.0 f | 58.85 ± 16.75 a | 12.86 ± 4.79 g | 1.71 ± 0.08 bc | ||
| LSD | 0.098 | 1.86 | 2.02 | 0.30 | 4.56 | 2.66 | 0.82 |
| Genotype | Salt Conc. | Treatments | RWC (%) | EL (%) | Proline (µmol g−1 FW) |
|---|---|---|---|---|---|
| Bulldog | 2.5 dS m−1 | Control (Hoagland) | 72.86 ± 10.57 | 62.9 ± 2.8 | NS |
| 6 dS m−1 | Control | 44.57 ± 1.77 | 63.5 ± 0.01 | 0.19 ± 0.01 | |
| Nano-K2SO4 | 61.61 ± 4.92 | 89.5 ± 2.6 | 0.4 ± 0.08 | ||
| Nano-ZnO | 54.69 ± 0.62 | ns * | ns | ||
| Nano-SiO2 | 49.17 ± 11.64 | 82.3 ± 7.6 | 0.28 ± 0.01 | ||
| 10 dS m−1 | Control | 37.97 ± 8.06 | 68.5 ± 1.1 | 0.29 ± 0.37 | |
| Nano-K2SO4 | 42.89 ± 4.15 | 96.8 ± 0.5 | 0.31 ± 0.01 | ||
| Nano-ZnO | 46.02 ± 0.39 | 94.7 ± 2.7 | 0.43 ± 0.o1 | ||
| Nano-SiO2 | 39.42 ± 0.63 | 82.5 ± 0.95 | 0.78 ± 0.071 | ||
| Mesa-Sirsa | 2.5 dS m−1 | Control (Hoagland) | 72.01 ± 2.5 | 64.7 ± 10.4 | 0.21 ± 0.03 |
| 6 dS m−1 | Control | 36.44 ± 1.78 | 74.1 ± 8.6 | 0.32 ± 0.01 | |
| Nano-K2SO4 | 63.39 ± 2.93 | 88.0 ± 4.0 | 0.36 ± 0.02 | ||
| Nano-ZnO | 60.71 ± 3.31 | 84.2 ± 3.3 | 0.53 ± 0.22 | ||
| Nano-SiO2 | 50.65 ± 1.02 | ns | ns | ||
| 10 dS m−1 | Control | 37.4 ± 6.24 | 74.4 ± 2.4 | 0.94 ± 0.01 | |
| Nano-K2SO4 | 50.36 ± 5.78 | 96.8 ± 2.2 | 1.02 ± 0.04 | ||
| Nano-ZnO | 46.16 ± 0.71 | 77.0 ± 1.5 | ns | ||
| Nano-SiO2 | 41.29 ± 1.75 | 84.5 ± 8.0 | 1.06 ± 0.06 | ||
| Genotype * Salt conc. | 173.17 * | 4.79 | 0.007 | ||
| Genotype * Treatments | 374.33 ** | 943.72 | 0.19 * | ||
| Salt Conc. * Treatment | 900.53 ** | 551.83 ** | 0.52 ** | ||
| Genotype * Salt Conc. * Treatments | 228.27 ** | 6920.94 ** | 0.52 ** | ||
| LSD | 2.03 | 2.50 | 0.005 | ||
| Genotype | Salt Conc. | Treatments | CAT | SOD |
|---|---|---|---|---|
| Bulldog 505 | 2.5 dS m−1 | Control (Hoagland) | 29.70 ± 2.39 | 94.19 ± 3.67 |
| 6 dS m−1 | Control | 35.68 ± 0.12 | 102.37 ± 15.54 | |
| Nano-K2SO4 | 49.21 ± 2.75 | 129.62 ± 4.03 | ||
| Nano-ZnO | NS * | NS | ||
| Nano-SiO2 | 114.14 ± 12.27 | 122.75 ± 6.87 | ||
| 10 dS m−1 | Control | 43.4 ± 9.9 | 102.13 ± 11.14 | |
| Nano-K2SO4 | 56.94 ± 4.11 | 186.21 ± 4.79 | ||
| Nano-ZnO | 88.58 ± 11.76 | 144.08 ± 9.32 | ||
| Nano-SiO2 | 105.14 ± 5.34 | 191.15 ± 13.71 | ||
| Mesa-Sirsa | 2.5 dS m−1 | Control (Hoagland) | 25.49 ± 3.66 | 98.80 ± 6.64 |
| 6 dS m−1 | Control | 38.24 ± 0.04 | 102.20 ± 18.31 | |
| Nano-K2SO4 | 60.66 ± 7.93 | 130.49 ± 6.81 | ||
| Nano-ZnO | 99.76 ± 4.1 | 134.12 ± 15.17 | ||
| Nano-SiO2 | NS | NS | ||
| 10 dS m−1 | Control | 51.47 ± 0.82 | 117.54 ± 9.48 | |
| Nano-K2SO4 | 94.32 ± 2.46 | 136.18 ± 10.77 | ||
| Nano-ZnO | NS | NS | ||
| Nano-SiO2 | 108.56 ± 9.83 | 143.46 ± 0.71 | ||
| Genotype * Salt conc. | 303.81 ** | 657.93 ** | ||
| Genotype * Treatments | 2114.10 ** | 315.66 * | ||
| Salt Conc.* Treatment | 1146.44 ** | 11,946.77 ** | ||
| Genotype * Salt Conc.* Treatments | 10,719.01 ** | 8393.59 ** | ||
| LSD | 10.46 | 7.73 | ||
| Genotype | Salt Conc. | Treatments | Na/K | Ca | Mg | P | S |
|---|---|---|---|---|---|---|---|
| % | mg. Kg−1 | ||||||
| Bulldog | 2.5 dS m−1 | Control (Hoagland) | 12.0 ± 1.5 i | 10,550.0 ± 2350.0 q | 2700.0 ± 100.0 f | 2250.0 ± 750.0 k | 2133.3 ± 122.2 o |
| 6 dS m−1 | Control | 13.8 ± 1.8 h | 42,233.3 ± 2442.0 f | 2533.3 ± 115.5 i | 1600.0 ± 173.2 p | 2500.0 ± 458.3 h | |
| Nano-K2SO4 | 7.6 ± 1.3 j | 28,600.0 ± 100.0 l | 2100.0 ± 100.0 k | 1800.0 ± 100.0 n | 2300.0 ± 100.0 l | ||
| Nano-ZnO | 18.13 ± 3.5 f | 19,350.0 ± 450.0 n | 7600.0 ± 10.0 a | 4600 ± 0.0 c | 7600.0 ± 15.0 b | ||
| Nano-SiO2 | 7.7 ± 1.6 j | 75,800.0 ± 0.0 a | 4200.0 ± 50.0 c | 2633.3 ± 155.0 h | 3233.3 ± 105.0 e | ||
| 10 dS m−1 | Control | 36.4 ± 3.5 b | 48,166.7 ± 405.1 c | 2566.7 ± 57.7 h | 1633.3 ± 152.8 o | 2366.7 ± 57.73 k | |
| Nano-K2SO4 | 20.8 ± 12.4 e | 36,300.0 ± 264.6 i | 1666.7 ± 152.8 n | 2733.3 ± 251.7 f | 2566.7 ± 115.5 g | ||
| Nano-ZnO | 30.1 ± 0.0 d | 43,950.0 ± 315.0 e | 2733.3 ± 351.2 e | 2400.0 ± 264.6 j | 2100 ± 624.5 p | ||
| Nano-SiO2 | 32.8 ± 8.4 c | 31,400.0 ± 1609.3 k | 1850.0 ± 50.0 m | 2000.0 ± 100.0 l | 2266.6 ± 115.4 m | ||
| Mesa-Sirsa | 2.5 dS m−1 | Control (Hoagland) | 13.03 ± 1.1 hi | 18,950.0 ± 6850.0 o | 6366.7 ± 1097.0 b | 4633.3 ± 251.4 b | 4566.7 ± 201.1 c |
| 6 dS m−1 | Control | 15.3 ± 1.1 g | 48,733.3 ± 850.5 b | 2266.7 ± 57.7 j | 2700 ± 173.2 g | 2466.7 ± 208.2 i | |
| Nano-K2SO4 | 6.9 ± 4.2 j | 32,116.7 ± 1675.1 j | 650.0 ± 35.0 q | 4250.0 ± 550.0 d | 3300.0 ± 900.0 d | ||
| Nano-ZnO | 7.09 ± 1.4 j | 39,050.0 ± 1550.0 g | 3450.0 ± 1250.0 d | 1833.3 ± 107.8 m | 1766.7 ± 101.1 q | ||
| Nano-SiO2 | 7.04 ± 1.6 j | 16,350.0 ± 5150.0 p | 2633.3 ± 251.7 g | 6633.3 ± 109.0 a | 8266.7 ± 134.5 a | ||
| 10 dS m−1 | Control | 21.3 ± 3.4 e | 46,891.4 ± 1370.8 d | 1909.1 ± 101.2 l | 2474.7 ± 198.7 i | 2151.2 ± 50.04 n | |
| Nano-K2SO4 | 15.7 ± 3.2 g | 37,066.7 ± 1285.8 h | 1500 ± 0.0 p | 3100 ± 100.0 e | 2633.3 ± 231.0 f | ||
| Nano-ZnO | 43.8 ± 0.0 a | 21,800.0 ± 0.0 m | 1600.0 ± 100.0 o | 900.0 ± 0.0 q | 1600.0 ± 0.0 r | ||
| Nano-SiO2 | 18.4 ± 4.1 f | 37,066.7 ± 1850.2 h | 2100.0 ± 200.0 k | 900.0 ± 100.0 q | 2400 ± 435.9 j | ||
| LSD | 16.8 | 4446.8 | 717.94 | 4335.4 | 5345.3 | ||
| Genotype | Salt Conc. | Treatments | Al | B | Cd | Cr | Cu | Fe | Mn | Ni | Si | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg. Kg−1 | ||||||||||||
| Bulldog | 2.5 dS m−1 | Control (Hoagland) | 12.9 ± 1.7 | 37.4 ± 2.8 | 1.6 ± 0.4 | 2.1 ± 0.8 | 8.2 ± 2.2 | 24.2 ± 2.4 | 74.2 ± 4.8 | 1.6 ± 0.4 | 31.03 ± 5.2 | 85.2 ± 8.7 |
| 6 dS m−1 | Control | 20.0 ± 0.0 | 53.6 ± 6.4 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 132.3 ± 7.1 | 140.6 ± 4.3 | 2.0 ± 0.0 | 164.9 ± 8.1 | 174.66 ± 24.7 | |
| Nano-K2SO4 | 12.9 ± 0.06 | 26.4 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.1 | 10.6 ± 0.1 | 34.2 ± 0.2 | 87.0 ± 0.1 | 1.3 ± 0.1 | 131.4 ± 0.2 | 28.9 ± 0.1 | ||
| Nano-ZnO | 40.0 ± 0.0 | 84.7 ± 0.0 | 3.1 ± 0.0 | 3.1 ± 0.0 | 15.5 ± 0.0 | 360.7 ± 0.0 | 188.2 ± 0.0 | 3.1 ± 0.0 | 160.7 ± 0.0 | 323.5 ± 75.1 | ||
| Nano-SiO2 | 31.7 ± 1.8 | 47.8 ± 3.3 | 2.7 ± 0.2 | 2.7 ± 1.0 | 13.7 ± 6.2 | 137.2 ± 6.3 | 105.2 ± 27.2 | 2.7 ± 1.2 | 399.5 ± 2.4 | 113.7 ± 6.8 | ||
| 10 dS m−1 | Control | 36.7 ± 2.8 | 68.0 ± 6.6 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 46.5 ± 2.6 | 156.0 ± 24.3 | 2.0 ± 0.0 | 47.0 ± 18.3 | 205.9 ± 1.9 | |
| Nano-K2SO4 | 20.0 ± 0.0 | 57.07 ± 8.2 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 73.7 ± 4.1 | 126.3 ± 6.1 | 2.0 ± 0.0 | 28.03 ± 9.8 | 159.2 ± 6.8 | ||
| Nano-ZnO | 17.8 ± 3.9 | 54.6 ± 13.6 | 1.8 ± 0.4 | 1.8 ± 0.4 | 8.9 ± 2.0 | 22.3 ± 2.6 | 102.9 ± 33.2 | 1.8 ± 0.4 | 47.6 ± 20.6 | 221.4 ± 56.3 | ||
| Nano-SiO2 | 20.0 ± 0.0 | 48.5 ± 4.8 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 35.2 ± 2.7 | 60.0 ± 5.3 | 2.0 ± 0.0 | 55.9 ± 16.1 | 87.0 ± 1.2 | ||
| Mesa-Sirsa | 2.5 dS m−1 | Control (Hoagland) | 80.8 ± 13.6 | 92.8 ± 4.2 | 2.0 ± 0.05 | 8.2 ± 1.1 | 15.9 ± 1.1 | 511.2 ± 76.7 | 162.8 ± 54.7 | 2.0 ± 0.07 | 428.7 ± 4.2 | 213.0 ± 13.2 |
| 6 dS m−1 | Control | 20.2 ± 0.3 | 50.1 ± 5.9 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 46.0 ± 2.6 | 114.8 ± 8.1 | 2.0 ± 0.0 | 49.0 ± 14.1 | 155.7 ± 8.5 | |
| Nano-K2SO4 | 18.7 ± 2.3 | 84.7 ± 4.1 | 1.9 ± 0.2 | 1.9 ± 0.2 | 9.7 ± 0.5 | 107.8 ± 6.2 | 127.0 ± 40.7 | 1.9 ± 0.2 | 176.0 ± 10.9 | 55.7 ± 1.6 | ||
| Nano-ZnO | 19.7 ± 0.6 | 42.3 ± 2.2 | 2.0 ± 0.06 | 2.0 ± 0.06 | 9.8 ± 0.3 | 19.7 ± 0.577 | 85.5 ± 25.1 | 2.0 ± 0.06 | 50.7 ± 17.3 | 189.0 ± 9.3 | ||
| Nano-SiO2 | 37.3 ± 14.8 | 112.2 ± 18.4 | 3.7 ± 1.4 | 3.7 ± 1.4 | 29.8 ± 2.6 | 160.4 ± 22.5 | 177.5 ± 25.6 | 3.7 ± 1.4 | 372.0 ± 50.7 | 138.8 ± 18.7 | ||
| 10 dS m−1 | Control | 20.0 ± 0.0 | 46.4 ± 1.3 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 35.5 ± 5.9 | 81.0 ± 10.6 | 2.0 ± 0.0 | 30.2 ± 11.0 | 163.3 ± 4.2 | |
| Nano-K2SO4 | 20.0 ± 0.0 | 60.4 ± 12.1 | 2.0 ± 0.0 | 2.0 ± 0..0 | 10.0 ± 0.0 | 70.0 ± 8.4 | 122.7 ± 25.3 | 2.0 ± 0.0 | 29.0 ± 1.8 | 160.0 ± 33.8 | ||
| Nano-ZnO | 17.8 ± 0.0 | 15.5 ± 0.0 | 1.7 ± 0.0 | 1.7 ± 0.0 | 8.9 ± 0.1 | 39.6 ± 0.0 | 12.2 ± 0.0 | 1.7 ± 0.0 | 102.6 ± 0.0 | 13.0 ± 0.0 | ||
| Nano-SiO2 | 35.0 ± 2.6 | 50.4 ± 3.2 | 2.0 ± 0.0 | 2.0 ± 0.0 | 10.0 ± 0.0 | 51.1 ± 2.3 | 56.6 ± 5.6 | 0.2 ± 0.0 | 65.7 ± 11.7 | 88.6 ± 7.7 | ||
| LSD | 5.33 | 7.68 | 0.8 | 4.3 | 1.14 | 3.06 | 11.1 | 4.0 | 2.74 | 11.42 | ||
| Shoot Dry Weight | Plant Height | No. of Flowers | No. of Tiller | Root Length | Root Fresh Weight | Root Dry Weight | RWC | Electrolyte Leakage | CAT | SOD | Proline | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot dry weight | 1.00 | 0.83 ** | 0.79 ** | 0.81 ** | 0.47 ** | 0.81 ** | 0.77 ** | 0.36 ** | 0.37 ** | 0.17 | 0.18 | 0.52 ** |
| Plant height | - | 1.00 | 0.79 ** | 0.7 ** | 0.48 ** | 0.85 ** | 0.81 ** | 0.26 | 0.34 * | 0.32 * | 0.43 ** | 0.45 ** |
| No. of Flower | - | - | 1.00 | 0.82 ** | 0.39 ** | 0.8 ** | 0.85 ** | 0.44 ** | 0.55 ** | 0.34 * | 0.34 * | 0.58 ** |
| No. of Tiller | - | - | - | 1.00 | 0.41 ** | 0.7 ** | 0.78 ** | 0.35 ** | 0.45 ** | 0.36 ** | 0.30 * | 0.58 ** |
| Root length | - | - | - | - | 1.00 | 0.48 ** | 0.53 ** | 0.53 ** | 0.2 | 0.32 * | 0.29 * | 0.34 * |
| Root fresh weight | - | - | - | - | - | 1.00 | 0.86 ** | 0.06 | 0.43 ** | 0.16 | 0.35 * | 0.53 ** |
| Root dry weight | - | - | - | - | - | - | 1.00 | 0.36 ** | 0.53 ** | 0.4 ** | 0.37 ** | 0.34 ** |
| RWC | - | - | - | - | - | - | - | 1.00 | -0.07 | 0.32 * | 0.01 | 0.23 |
| Electrolyte Leakage | - | - | - | - | - | - | - | - | 1.00 | 0.21 | 0.12 | 0.39 |
| CAT | - | - | - | - | - | - | - | - | - | 1.00 | 0.38 | 0.43 ** |
| SOD | - | - | - | - | - | - | - | - | - | - | 1.00 | 0.21 |
| Proline | - | - | - | - | - | - | - | - | - | - | - | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Shal, R.M.; El-Naggar, A.H.; El-Beshbeshy, T.R.; Mahmoud, E.K.; El-Kader, N.I.A.; Missaui, A.M.; Du, D.; Ghoneim, A.M.; El-Sharkawy, M.S. Effect of Nano-Fertilizers on Alfalfa Plants Grown under Different Salt Stresses in Hydroponic System. Agriculture 2022, 12, 1113. https://doi.org/10.3390/agriculture12081113
El-Shal RM, El-Naggar AH, El-Beshbeshy TR, Mahmoud EK, El-Kader NIA, Missaui AM, Du D, Ghoneim AM, El-Sharkawy MS. Effect of Nano-Fertilizers on Alfalfa Plants Grown under Different Salt Stresses in Hydroponic System. Agriculture. 2022; 12(8):1113. https://doi.org/10.3390/agriculture12081113
Chicago/Turabian StyleEl-Shal, Rania M., Ahmed H. El-Naggar, Talaat R. El-Beshbeshy, Esawy K. Mahmoud, Nasser I. Abd El-Kader, Ali M. Missaui, Daolin Du, Adel M. Ghoneim, and Mahmoud S. El-Sharkawy. 2022. "Effect of Nano-Fertilizers on Alfalfa Plants Grown under Different Salt Stresses in Hydroponic System" Agriculture 12, no. 8: 1113. https://doi.org/10.3390/agriculture12081113
APA StyleEl-Shal, R. M., El-Naggar, A. H., El-Beshbeshy, T. R., Mahmoud, E. K., El-Kader, N. I. A., Missaui, A. M., Du, D., Ghoneim, A. M., & El-Sharkawy, M. S. (2022). Effect of Nano-Fertilizers on Alfalfa Plants Grown under Different Salt Stresses in Hydroponic System. Agriculture, 12(8), 1113. https://doi.org/10.3390/agriculture12081113