Physical Activity and Depression in Breast Cancer Patients: Mechanisms and Therapeutic Potential

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Biological Mechanisms of Depression and the Crucial Role of Physical Activity
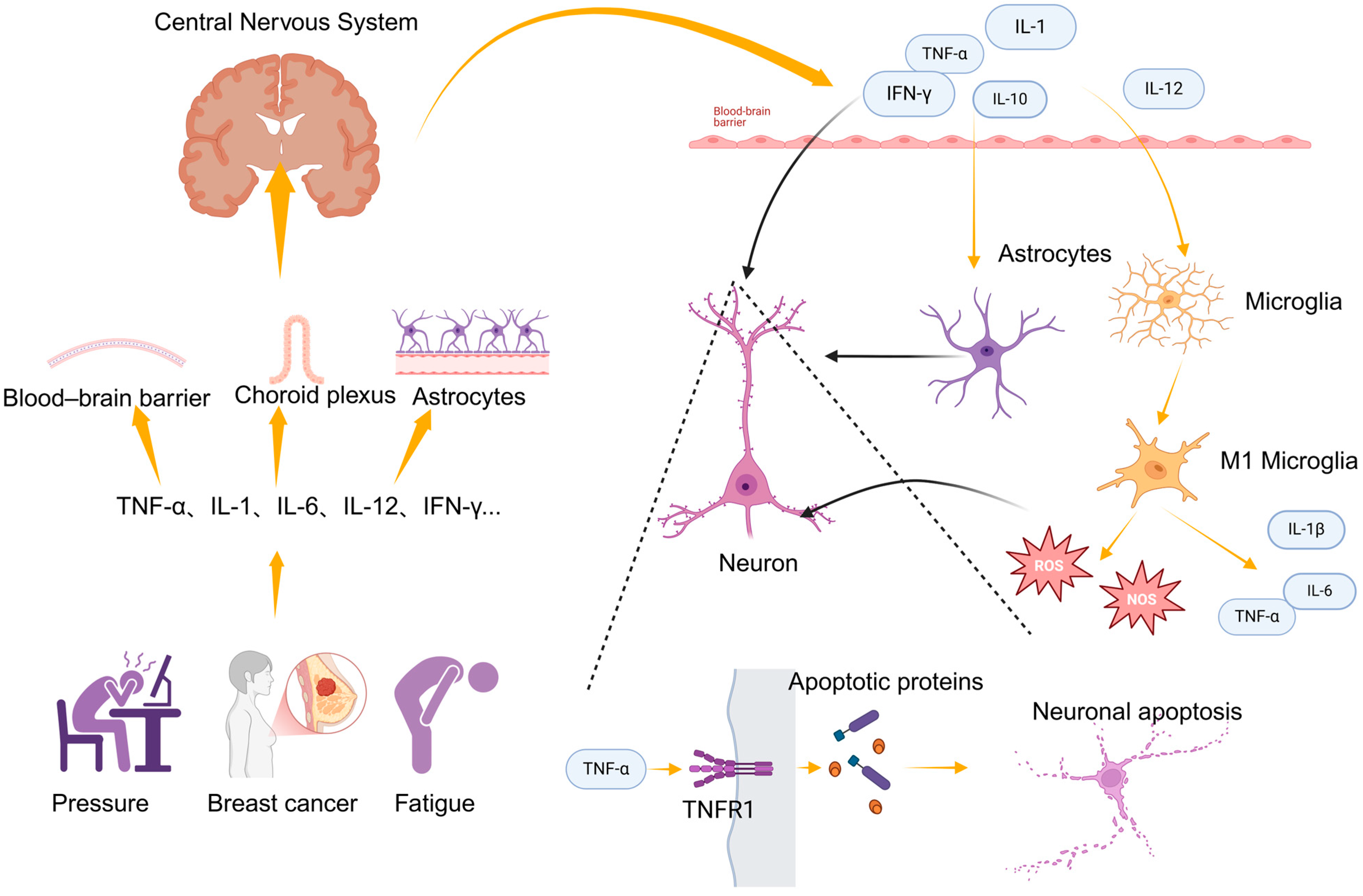
2.1. Inflammation
2.2. Neuroplasticity
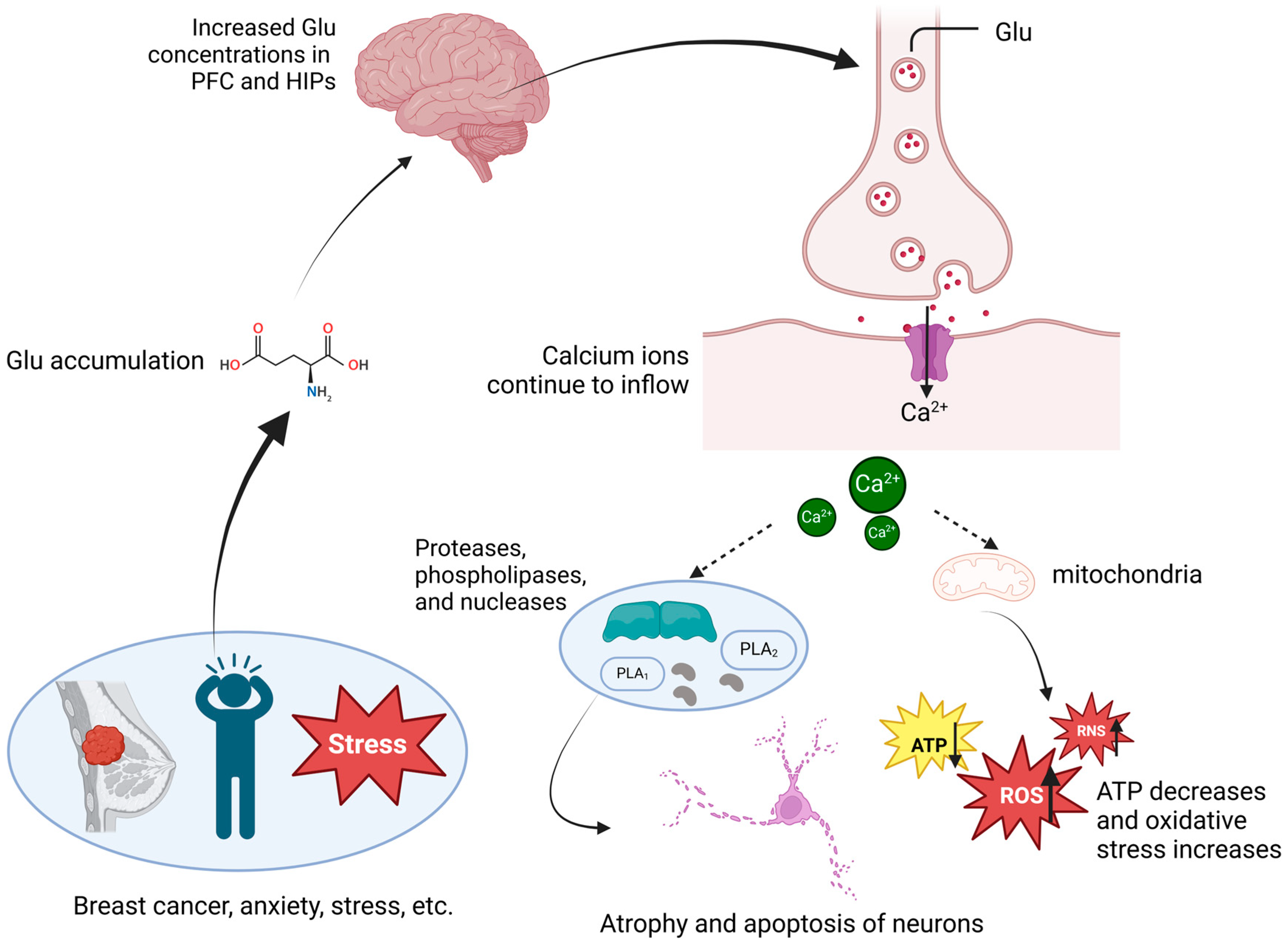
2.3. Glutamate Metabolism
2.4. Other Factors
2.4.1. Oxidative Stress
2.4.2. Neuroendocrine System
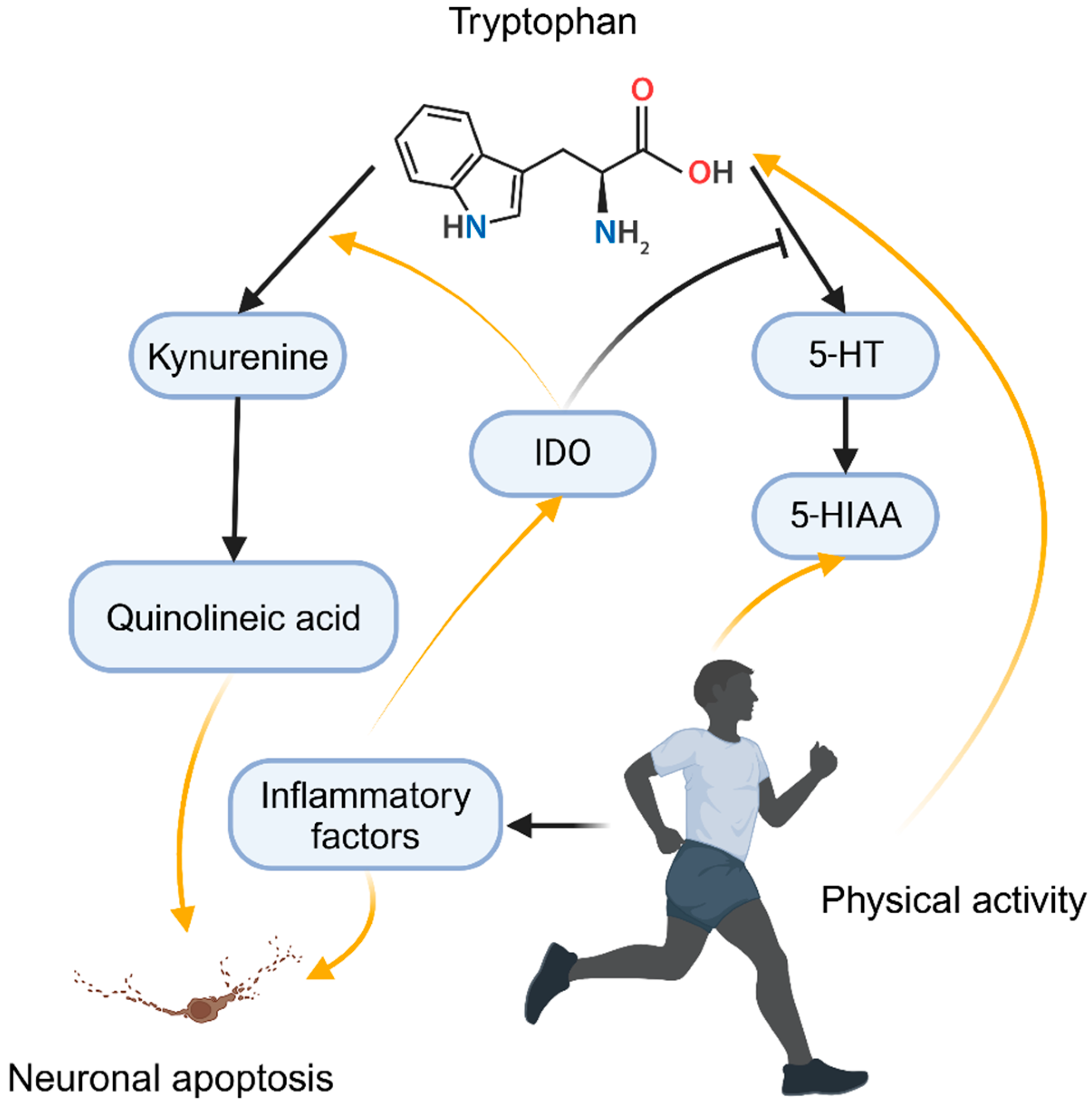
2.4.3. Monoamine Neurotransmitters
3. Discussion
4. Conclusions
5. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Park, H.; Kim, K. Impact of Psycho-Social Factors on Fatigue among Breast Cancer Patients Who Are Currently Undergoing Radiotherapy. Int. J. Environ. Res. Public Health 2020, 17, 6092. [Google Scholar] [CrossRef] [PubMed]
- Mausbach, B.T.; Schwab, R.B.; Irwin, S.A. Depression as a predictor of adherence to adjuvant endocrine therapy (AET) in women with breast cancer: A systematic review and meta-analysis. Breast Cancer Res. Treat. 2015, 152, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Aitken, L.-A.; Hossan, S.Z. The Psychological Distress and Quality of Life of Breast Cancer Survivors in Sydney, Australia. Healthcare 2022, 10, 2017. [Google Scholar] [CrossRef]
- Pilevarzadeh, M. Global prevalence of depression among breast cancer patients: A systematic review and meta-analysis. Breast Cancer Res. Treat. 2019, 176, 519–533. [Google Scholar] [CrossRef] [PubMed]
- García-Montero, C.; Ortega, M.A.; Alvarez-Mon, M.A.; Fraile-Martinez, O.; Romero-Bazán, A.; Lahera, G.; Montes-Rodríguez, J.M.; Molina-Ruiz, R.M.; Mora, F.; Rodriguez-Jimenez, R.; et al. The Problem of Malnutrition Associated with Major Depressive Disorder from a Sex-Gender Perspective. Nutrients 2022, 14, 1107. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, N.; Pariante, C.M.; Zunszain, P.A. Immune mechanisms linked to depression via oxidative stress and neuroprogression. Immunology 2015, 144, 365–373. [Google Scholar] [CrossRef]
- McFarland, D.C.; Doherty, M.; Atkinson, T.M.; O’Hanlon, R.; Breitbart, W.; Nelson, C.J.; Miller, A.H. Cancer-Related Inflammation and Depressive Symptoms: Systematic Review and Meta-Analysis. Cancer 2022, 128, 2504–2519. [Google Scholar] [CrossRef]
- McFarland, D.C.; Riba, M.; Grassi, L. Clinical Implications of Cancer Related Inflammation and Depression: A Critical Review. Clin. Pract. Epidemiol. Ment. Health 2021, 17, 287–294. [Google Scholar] [CrossRef]
- Hofmann, S.G.; Asnaani, A.; Vonk, I.J.J.; Sawyer, A.T.; Fang, A. The Efficacy of Cognitive Behavioral Therapy: A Review of Meta-analyses. Cognit. Ther. Res. 2012, 36, 427–440. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Alsuwaidan, M.; Baune, B.T.; Berk, M.; Demyttenaere, K.; Goldberg, J.F.; Gorwood, P.; Ho, R.; Kasper, S.; Kennedy, S.H.; et al. Treatment-resistant depression: Definition, prevalence, detection, management, and investigational interventions. World Psychiatry 2023, 22, 394–412. [Google Scholar] [CrossRef]
- Cuijpers, P.; Quero, S.; Dowrick, C.; Arroll, B. Psychological Treatment of Depression in Primary Care: Recent Developments. Curr. Psychiatry Rep. 2019, 21, 129. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.R. The NIMH Research Domain Criteria (RDoC) Project: Precision medicine for psychiatry. Am. J. Psychiatry 2014, 171, 395–397. [Google Scholar] [CrossRef]
- Schuch, F.; Vancampfort, D.; Firth, J.; Rosenbaum, S.; Ward, P.; Silva, E.S.; Hallgren, M.; de Leon, A.P.D.P.; Dunn, A.L.; Deslandes, A.; et al. Physical Activity and Incident Depression: A Meta-Analysis of Prospective Cohort Studies. Am. J. Psychiatry 2018, 175, 631–648. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.W.; Chen, C.-Y.; Stein, M.B.; Klimentidis, Y.C.; Wang, M.-J.; Koenen, K.C.; Smoller, J.W.; Major Depressive Disorder Working Group of the Psychiatric Genomics Consortium. Assessment of Bidirectional Relationships Between Physical Activity and Depression Among Adults: A 2-Sample Mendelian Randomization Study. JAMA Psychiatry 2019, 76, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.W.; Zheutlin, A.B.; Karlson, R.A.; Wang, M.-J.; Dunn, E.C.; Stein, M.B.; Karlson, E.W.; Smoller, J.W. Physical activity offsets genetic risk for incident depression assessed via electronic health records in a biobank cohort study. Depress. Anxiety 2020, 37, 106–114. [Google Scholar] [CrossRef]
- Patsou, E.D.; Alexias, G.D.; Anagnostopoulos, F.G.; Karamouzis, M.V. Effects of physical activity on depressive symptoms during breast cancer survivorship: A meta-analysis of randomised control trials. ESMO Open 2017, 2, e000271. [Google Scholar] [CrossRef] [PubMed]
- Fisher, H.; Jacobs, J.M.; Taub, C.; Lechner, S.; Lewis, J.; Carver, C.; Blomberg, B.; Antoni, M. How changes in physical activity relate to fatigue interference, mood, and quality of life during treatment for non-metastatic breast cancer. Gen. Hosp. Psychiatry 2017, 49, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Cannioto, R.A.; Hutson, A.; Dighe, S.; McCann, W.; McCann, S.E.; Zirpoli, G.R.; Barlow, W.; Kelly, K.M.; DeNysschen, C.A.; Hershman, D.L.; et al. Physical Activity Before, During, and After Chemotherapy for High-Risk Breast Cancer: Relationships With Survival. J. Natl. Cancer Inst. 2021, 113, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Baumann, F.T.; Bloch, W.; Weissen, A.; Brockhaus, M.; Beulertz, J.; Zimmer, P.; Streckmann, F.; Zopf, E.M. Physical Activity in Breast Cancer Patients during Medical Treatment and in the Aftercare—A Review. Breast Care 2013, 8, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Tranchita, E.; Murri, A.; Grazioli, E.; Cerulli, C.; Emerenziani, G.P.; Ceci, R.; Caporossi, D.; Dimauro, I.; Parisi, A. The Beneficial Role of Physical Exercise on Anthracyclines Induced Cardiotoxicity in Breast Cancer Patients. Cancers 2022, 14, 2288. [Google Scholar] [CrossRef] [PubMed]
- Boyle, C.; Ganz, P.; Dyk, K.; Bower, J. Inflammation and attentional bias in breast cancer survivors. Brain Behav. Immun. 2017, 66, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Negishi, H.; Taniguchi, T.; Yanai, H. The Interferon (IFN) Class of Cytokines and the IFN Regulatory Factor (IRF) Transcription Factor Family. Cold Spring Harb. Perspect. Biol. 2018, 10, a028423. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.F.; Andrade, C. Interferon-Related Depression: A Primer on Mechanisms, Treatment, and Prevention of a Common Clinical Problem. Curr. Neuropharmacol. 2016, 14, 743–748. [Google Scholar] [CrossRef]
- Chiu, W.-C.; Su, Y.-P.; Su, K.-P.; Chen, P.-C. Recurrence of depressive disorders after interferon-induced depression. Transl. Psychiatry 2017, 7, e1026. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Zhang, H.; Baloch, Z. Pathogenetic and Therapeutic Applications of Tumor Necrosis Factor-α (TNF-α) in Major Depressive Disorder: A Systematic Review. Int. J. Mol. Sci. 2016, 17, 733. [Google Scholar] [CrossRef]
- Brymer, K.J.; Romay-Tallon, R.; Allen, J.; Caruncho, H.J.; Kalynchuk, L.E. Exploring the Potential Antidepressant Mechanisms of TNFα Antagonists. Front. Neurosci. 2019, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Toups, M.; Nemeroff, C. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.; Derry, H.; Fagundes, C. Inflammation: Depression fans the flames and feasts on the heat. Am. J. Psychiatry 2015, 172, 1075–1091. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H.; Maletić, V.; Raison, C. Inflammation and Its Discontents: The Role of Cytokines in the Pathophysiology of Major Depression. Biol. Psychiatry 2009, 65, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Berk, M.; Williams, L.; Jacka, F.; O’Neil, A.; Pasco, J.; Moylan, S.; Allen, N.; Stuart, A.; Hayley, A.; Byrne, M.; et al. So depression is an inflammatory disease, but where does the inflammation come from? Bmc Med. 2013, 11, 200. [Google Scholar] [CrossRef]
- Raison, C.; Miller, A.H. Is Depression an Inflammatory Disorder? Curr. Psychiatry Rep. 2011, 13, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Hussain, B.; Chang, J. Peripheral inflammation and blood–brain barrier disruption: Effects and mechanisms. CNS Neurosci. Ther. 2020, 27, 36–47. [Google Scholar] [CrossRef]
- Shimada, A.; Hasegawa-Ishii, S. Increased cytokine expression in the choroid plexus stroma and epithelium in response to endotoxin-induced systemic inflammation in mice. Toxicol. Rep. 2021, 8, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Argaw, A.T.; Asp, L.; Zhang, J.; Navrazhina, K.; Pham, T.; Mariani, J.N.; Mahase, S.; Dutta, D.J.; Seto, J.; Kramer, E.G.; et al. Astrocyte-derived VEGF-A drives blood-brain barrier disruption in CNS inflammatory disease. J. Clin. Investig. 2012, 122, 2454–2468. [Google Scholar] [CrossRef]
- Lopez-Ortiz, A.O.; Eyo, U.B. Astrocytes and microglia in the coordination of CNS development and homeostasis. J. Neurochem. 2024, 168, 3599–3614. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, J.; Bao, J.; Bai, Q.; Wang, G. Interaction of Microglia and Astrocytes in the Neurovascular Unit. Front. Immunol. 2020, 11, 514779. [Google Scholar] [CrossRef]
- Chadwick, W.; Magnus, T.; Martin, B.; Keselman, A.; Mattson, M.P.; Maudsley, S. Targeting TNF-α receptors for neurotherapeutics. Trends Neurosci. 2008, 31, 504–511. [Google Scholar] [CrossRef]
- Jayaraman, A.; Htike, T.T.; James, R.; Picon, C.; Reynolds, R. TNF-mediated neuroinflammation is linked to neuronal necroptosis in Alzheimer’s disease hippocampus. Acta Neuropathol. Commun. 2021, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Haroon, E.; Raison, C.L.; Miller, A.H. Psychoneuroimmunology meets neuropsychopharmacology: Translational implications of the impact of inflammation on behavior. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2012, 37, 137–162. [Google Scholar] [CrossRef]
- Ida, T.; Hara, M.; Nakamura, Y.; Kozaki, S.; Tsunoda, S.; Ihara, H. Cytokine-induced enhancement of calcium-dependent glutamate release from astrocytes mediated by nitric oxide. Neurosci. Lett. 2008, 432, 232–236. [Google Scholar] [CrossRef]
- Matute, C.; Domercq, M.; Sánchez-Gómez, M.-V. Glutamate-mediated glial injury: Mechanisms and clinical importance. Glia 2006, 53, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Hanna, M.; Dumas, I.; Orain, M.; Jacob, S.; Têtu, B.; Diorio, C. Association between physical activity and the expression of mediators of inflammation in normal breast tissue among premenopausal and postmenopausal women. Cytokine 2017, 102, 151–160. [Google Scholar] [CrossRef]
- Fischer, C.P.; Berntsen, A.; Perstrup, L.B.; Eskildsen, P.; Pedersen, B.K. Plasma levels of interleukin-6 and C-reactive protein are associated with physical inactivity independent of obesity. Scand. J. Med. Sci. Sports 2007, 17, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, K. Biological functions of tumor necrosis factor cytokines and their receptors. Cytokine Growth Factor Rev. 2003, 14, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Mathur, N.; Pedersen, B.K. Exercise as a mean to control low-grade systemic inflammation. Mediat. Inflamm. 2008, 2008, 109502. [Google Scholar] [CrossRef]
- Matthys, P.; Mitera, T.; Heremans, H.; Van Damme, J.; Billiau, A. Anti-gamma interferon and anti-interleukin-6 antibodies affect staphylococcal enterotoxin B-induced weight loss, hypoglycemia, and cytokine release in D-galactosamine-sensitized and unsensitized mice. Infect. Immun. 1995, 63, 1158–1164. [Google Scholar] [CrossRef]
- Freeman, B.D.; Buchman, T.G. Interleukin-1 receptor antagonist as therapy for inflammatory disorders. Expert Opin. Biol. Ther. 2001, 1, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.L.; Weaver, C.T. Diversity in the contribution of interleukin-10 to T-cell-mediated immune regulation. Immunol. Rev. 2008, 226, 219–233. [Google Scholar] [CrossRef]
- Svensson, M.; Lexell, J.; Deierborg, T. Effects of Physical Exercise on Neuroinflammation, Neuroplasticity, Neurodegeneration, and Behavior: What We Can Learn From Animal Models in Clinical Settings. Neurorehabilit. Neural Repair 2015, 29, 577–589. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 2011, 11, 607–615. [Google Scholar] [CrossRef]
- Nieman, D.C.; Henson, D.A.; Gusewitch, G.; Warren, B.J.; Dotson, R.C.; Butterworth, D.E.; Nehlsen-Cannarella, S.L. Physical activity and immune function in elderly women. Med. Sci. Sports Exerc. 1993, 25, 823–831. [Google Scholar] [CrossRef]
- Shinkai, S.; Kohno, H.; Kimura, K.; Komura, T.; Asai, H.; Inai, R.; Oka, K.; Kurokawa, Y.; Shephard, R. Physical activity and immune senescence in men. Med. Sci. Sports Exerc. 1995, 27, 1516–1526. [Google Scholar] [CrossRef]
- Spielmann, G.; McFarlin, B.K.; O’Connor, D.P.; Smith, P.J.W.; Pircher, H.; Simpson, R.J. Aerobic fitness is associated with lower proportions of senescent blood T-cells in man. Brain Behav. Immun. 2011, 25, 1521–1529. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Bruunsgaard, H. Possible beneficial role of exercise in modulating low-grade inflammation in the elderly. Scand. J. Med. Sci. Sports 2003, 13, 56–62. [Google Scholar] [CrossRef]
- Bigley, A.B.; Rezvani, K.; Pistillo, M.; Reed, J.; Agha, N.; Kunz, H.; O’Connor, D.P.; Sekine, T.; Bollard, C.M.; Simpson, R.J. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and augments cytotoxicity against lymphoma and multiple myeloma target cells. Part II: Impact of latent cytomegalovirus infection and catecholamine sensitivity. Brain Behav. Immun. 2015, 49, 59–65. [Google Scholar] [CrossRef]
- Pedersen, L.; Idorn, M.; Olofsson, G.H.; Lauenborg, B.; Nookaew, I.; Hansen, R.H.; Johannesen, H.H.; Becker, J.C.; Pedersen, K.S.; Dethlefsen, C.; et al. Voluntary Running Suppresses Tumor Growth through Epinephrine- and IL-6-Dependent NK Cell Mobilization and Redistribution. Cell Metab. 2016, 23, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Karpe, F.; Lafontan, M.; Frayn, K. Physical activity and exercise in the regulation of human adipose tissue physiology. Physiol. Rev. 2012, 92, 157–191. [Google Scholar] [CrossRef]
- Flynn, M.G.; McFarlin, B.K.; Phillips, M.D.; Stewart, L.K.; Timmerman, K.L. Toll-like receptor 4 and CD14 mRNA expression are lower in resistive exercise-trained elderly women. J. Appl. Physiol. 2003, 95, 1833–1842. [Google Scholar] [CrossRef]
- Guo, X.; Le, Y. The triangular relationship of physical activity, depression, and inflammatory markers: A large cross-sectional analysis with NHANES data. J. Affect. Disorders 2024, 367, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, E.M.; Loukov, D.; Bowdish, D.M.E.; Heisz, J.J. Exercise reduces depression and inflammation but intensity matters. Biol. Psychol. 2018, 133, 79–84. [Google Scholar] [CrossRef]
- Bremner, J.D.; Vermetten, E.; Mazure, C.M. Development and preliminary psychometric properties of an instrument for the measurement of childhood trauma: The Early Trauma Inventory. Depress. Anxiety 2000, 12, 1–12. [Google Scholar] [CrossRef]
- Mervaala, E.; Föhr, J.; Könönen, M.; Valkonen-Korhonen, M.; Vainio, P.; Partanen, K.; Partanen, J.; Tiihonen, J.; Viinamäki, H.; Karjalainen, A.K.; et al. Quantitative MRI of the hippocampus and amygdala in severe depression. Psychol. Med. 2000, 30, 117–125. [Google Scholar] [CrossRef]
- Sheline, Y.I.; Wang, P.W.; Gado, M.H.; Csernansky, J.G.; Vannier, M.W. Hippocampal atrophy in recurrent major depression. Proc. Natl. Acad. Sci. USA 1996, 93, 3908–3913. [Google Scholar] [CrossRef] [PubMed]
- Carroll, B.J. Untreated depression and hippocampal volume loss. Am. J. Psychiatry 2003, 160, 1516–1518. [Google Scholar] [CrossRef] [PubMed]
- Stockmeier, C.A.; Mahajan, G.J.; Konick, L.C.; Overholser, J.C.; Jurjus, G.J.; Meltzer, H.Y.; Uylings, H.B.M.; Friedman, L.; Rajkowska, G. Cellular changes in the postmortem hippocampus in major depression. Biol. Psychiatry 2004, 56, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.D.; Vythilingam, M.; Vermetten, E.; Nazeer, A.; Adil, J.; Khan, S.; Staib, L.H.; Charney, D.S. Reduced volume of orbitofrontal cortex in major depression. Biol. Psychiatry 2002, 51, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Drevets, W.C.; Price, J.L.; Simpson, J.R.; Todd, R.D.; Reich, T.; Vannier, M.; Raichle, M.E. Subgenual prefrontal cortex abnormalities in mood disorders. Nature 1997, 386, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Cotter, D.R.; Pariante, C.M.; Everall, I.P. Glial cell abnormalities in major psychiatric disorders: The evidence and implications. Brain Res. Bull. 2001, 55, 585–595. [Google Scholar] [CrossRef]
- Rajkowska, G.; Miguel-Hidalgo, J.J.; Wei, J.; Dilley, G.; Pittman, S.D.; Meltzer, H.Y.; Overholser, J.C.; Roth, B.L.; Stockmeier, C.A. Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol. Psychiatry 1999, 45, 1085–1098. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; Miguel-Hidalgo, J.J.; O’Dwyer, G.; Stockmeier, C.A.; Rajkowska, G. Age-dependent reductions in the level of glial fibrillary acidic protein in the prefrontal cortex in major depression. Neuropsychopharmacology 2004, 29, 2088–2096. [Google Scholar] [CrossRef]
- Hamidi, M.; Drevets, W.C.; Price, J.L. Glial reduction in amygdala in major depressive disorder is due to oligodendrocytes. Biol. Psychiatry 2004, 55, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Player, M.; Taylor, J.L.; Weickert, C.; Alonzo, A.; Sachdev, P.; Martin, D.; Mitchell, P.; Loo, C. Neuroplasticity in Depressed Individuals Compared with Healthy Controls. Neuropsychopharmacol 2013, 38, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Karkare, S. Stress, Depression and Neuroplasticity. arXiv: Neurons and Cognition 2017. Available online: https://consensus.app/papers/stress-depression-neuroplasticity-singh/c49f321aa4e754288485ec784639dfb5/ (accessed on 14 September 2024).
- Player, M.; Taylor, J.L.; Weickert, C.; Alonzo, A.; Sachdev, P.; Martin, D.; Mitchell, P.; Loo, C. Increase in PAS-induced neuroplasticity after a treatment course of transcranial direct current stimulation for depression. J. Affect. Disord. 2014, 167, 140–147. [Google Scholar] [CrossRef]
- Brunoni, A.; Lopes, M.; Fregni, F. A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: Implications for the role of neuroplasticity in depression. Int. J. Neuropsychopharmacol. 2008, 11, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Dooley, L.; Ganz, P.; Cole, S.; Crespi, C.; Bower, J. Val66Met BDNF polymorphism as a vulnerability factor for inflammation-associated depressive symptoms in women with breast cancer. J. Affect. Disord. 2016, 197, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Kim, C.-J.; Kwak, H.; No, M.-H.; Heo, J.-W.; Kim, T.-W. Physical exercise prevents cognitive impairment by enhancing hippocampal neuroplasticity and mitochondrial function in doxorubicin-induced chemobrain. Neuropharmacology 2018, 133, 451–461. [Google Scholar] [CrossRef]
- Müller, P.; Duderstadt, Y.; Lessmann, V.; Müller, N. Lactate and BDNF: Key Mediators of Exercise Induced Neuroplasticity? J. Clin. Med. 2020, 9, 1136. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Ying, Z.; Roy, R.; Molteni, R.; Edgerton, V. Voluntary exercise induces a BDNF-mediated mechanism that promotes neuroplasticity. J. Neurophysiol. 2002, 88, 2187–2195. [Google Scholar] [CrossRef] [PubMed]
- Mellow, M.; Goldsworthy, M.; Coussens, S.; Smith, A.E. Acute aerobic exercise and neuroplasticity of the motor cortex: A systematic review. J. Sci. Med. Sport 2019, 23, 408–414. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.; Buckley, J.; Opie, G.; Ridding, M.; Semmler, J. A single bout of aerobic exercise promotes motor cortical neuroplasticity. J. Appl. Physiol. 2013, 114, 1174–1182. [Google Scholar] [CrossRef]
- Hötting, K.; Röder, B. Beneficial effects of physical exercise on neuroplasticity and cognition. Neurosci. Biobehav. Rev. 2013, 37, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Luo, R.; Valaris, S.; Haley, E.B.; Takase, H.; Chen, Y.I.; Dickerson, B.; Schon, K.; Arai, K.; Nguyen, C.; et al. Diffusion tensor-MRI detects exercise-induced neuroplasticity in the hippocampal microstructure in mice. Brain Plast. 2020, 5, 147–159. [Google Scholar] [CrossRef]
- Real, C.; Garcia, P.C.; Britto, L.; Pires, R.S. Different protocols of treadmill exercise induce distinct neuroplastic effects in rat brain motor areas. Brain Res. 2015, 1624, 188–198. [Google Scholar] [CrossRef]
- Brüchle, W.; Schwarzer, C.; Berns, C.; Scho, S.; Schneefeld, J.; Koester, D.; Schack, T.; Schneider, U.; Rosenkranz, K. Physical Activity Reduces Clinical Symptoms and Restores Neuroplasticity in Major Depression. Front. Psychiatry 2021, 12, 660642. [Google Scholar] [CrossRef] [PubMed]
- Tolahunase, M.; Sagar, R.; Faiq, M.; Dada, R. Yoga- and meditation-based lifestyle intervention increases neuroplasticity and reduces severity of major depressive disorder: A randomized controlled trial. Restor. Neurol. Neurosci. 2018, 36, 423–442. [Google Scholar] [CrossRef]
- Aguiar, A.; Stragier, E.; Scheffer, D.; Remor, A.; Oliveira, P.A.; Prediger, R.; Latini, A.; Raisman-Vozari, R.; Mongeau, R.; Lanfumey, L. Effects of exercise on mitochondrial function, neuroplasticity and anxio-depressive behavior of mice. Neuroscience 2014, 271, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, C.; Duman, R. Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacol 2008, 33, 88–109. [Google Scholar] [CrossRef]
- Schuch, F.; Deslandes, A.; Stubbs, B.; Gosmann, N.P.; da Silva, C.T.B.; Fleck, M. Neurobiological effects of exercise on major depressive disorder: A systematic review. Neurosci. Biobehav. Rew. 2016, 61, 1–11. [Google Scholar] [CrossRef]
- Sanacora, G.; Treccani, G.; Popoli, M. Towards a glutamate hypothesis of depression An emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology 2012, 62, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Madeira, C.; Vargas-Lopes, C.; Brandão, C.O.; Reis, T.; Laks, J.; Panizzutti, R.; Ferreira, S. Elevated Glutamate and Glutamine Levels in the Cerebrospinal Fluid of Patients With Probable Alzheimer’s Disease and Depression. Front. Psychiatry 2018, 9, 561. [Google Scholar] [CrossRef] [PubMed]
- Reznikov, L.R.; Fadel, J.; Reagan, L. Glutamate-Mediated Neuroplasticity Deficits in Mood Disorders. In Neuroplasticity; Springer: Tarporley, UK, 2011; pp. 13–26. [Google Scholar]
- Nakamura, T.; Lipton, S. Preventing Ca2+-mediated nitrosative stress in neurodegenerative diseases: Possible pharmacological strategies. Cell Calcium 2010, 47, 190–197. [Google Scholar] [CrossRef]
- Nicholls, D. Oxidative Stress and Energy Crises in Neuronal Dysfunction. Ann. N. Y. Acad. Sci. 2008, 1147, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Verwer, R.; Wamelen, D.J.; Qi, X.-R.; Gao, S.-F.; Lucassen, P.; Swaab, D. Prefrontal changes in the glutamate-glutamine cycle and neuronal/glial glutamate transporters in depression with and without suicide. J. Psychiatr. Res. 2016, 82, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Valentine, G.W.; Sanacora, G. Targeting glial physiology and glutamate cycling in the treatment of depression. Biochem. Pharmacol. 2009, 78, 431–439. [Google Scholar] [CrossRef]
- Murrough, J.; Abdallah, C.; Mathew, S. Targeting glutamate signalling in depression: Progress and prospects. Nat. Rev. Drug Discov. 2017, 16, 472–486. [Google Scholar] [CrossRef]
- Berman, R.; Cappiello, A.; Anand, A.; Oren, D.; Heninger, G.; Charney, D.; Krystal, J. Antidepressant effects of ketamine in depressed patients. Biol. Psychiatry 2000, 47, 351–354. [Google Scholar] [CrossRef]
- Xiao, K.; Luo, Y.; Liang, X.; Tang, J.; Wang, J.; Xiao, Q.; Qi, Y.; Li, Y.; Zhu, P.; Yang, H.; et al. Beneficial effects of running exercise on hippocampal microglia and neuroinflammation in chronic unpredictable stress-induced depression model rats. Transl. Psychiatry 2021, 11, 461. [Google Scholar] [CrossRef]
- Mattson, M.P. Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N. Y. Acad. Sci. 2008, 1144, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Haroon, E.; Miller, A.H. Inflammation Effects on Brain Glutamate in Depression: Mechanistic Considerations and Treatment Implications. Curr. Top. Behav. Neurosci. 2017, 31, 173–198. [Google Scholar] [PubMed]
- Świątkiewicz, M.; Fiedorowicz, M.; Orzeł, J.; Wełniak-Kamińska, M.; Bogorodzki, P.; Langfort, J.; Grieb, P. Increases in Brain 1H-MR Glutamine and Glutamate Signals Following Acute Exhaustive Endurance Exercise in the Rat. Front. Physiol. 2017, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.; Mantese, C.E.; Porciúncula, L.; Ghisleni, G.; Vinadé, L.; Souza, D.; Portela, L. Exercise affects glutamate receptors in postsynaptic densities from cortical mice brain. Brain Res. 2005, 1065, 20–25. [Google Scholar] [CrossRef]
- Vivar, C.; van Praag, H. Running Changes the Brain: The Long and the Short of It. Physiology 2017, 32, 410–424. [Google Scholar] [CrossRef]
- Baskerville, R.; McGrath, T.; Castell, L. The effects of physical activity on glutamate neurotransmission in neuropsychiatric disorders. Front. Sports Act Living 2023, 5, 1147384. [Google Scholar] [CrossRef] [PubMed]
- Greenamyre, J.T. The role of glutamate in neurotransmission and in neurologic disease. Arch. Neurol. 1986, 43, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Ramonet, D.; Rodríguez, M.J.; Fredriksson, K.; Bernal, F.; Mahy, N. In vivo neuroprotective adaptation of the glutamate/glutamine cycle to neuronal death. Hippocampus 2004, 14, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Han, C.; Zeng, Z.; Liu, L.; Wang, H.; Xu, J.; Feng, Z.-P.; Little, P.J.; Quirion, R.; Zheng, W. Glutamate Attenuates the Survival Property of IGFR through NR2B Containing N-Methyl-D-aspartate Receptors in Cortical Neurons. Oxid. Med. Cell. Longev. 2020, 2020, 5173184. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C. URI in athletes: Are mucosal immunity and cytokine responses key risk factors? Exerc. Sport Sci. Rev. 2013, 41, 148–153. [Google Scholar] [CrossRef]
- Haroon, E.; Fleischer, C.C.; Felger, J.C.; Chen, X.; Woolwine, B.J.; Patel, T.; Hu, X.P.; Miller, A.H. Conceptual convergence: Increased inflammation is associated with increased basal ganglia glutamate in patients with major depression. Mol. Psychiatry 2016, 21, 1351–1357. [Google Scholar] [CrossRef]
- Moylan, S.; Berk, M.; Dean, O.; Samuni, Y.; Williams, L.; O’Neil, A.; Hayley, A.; Pasco, J.; Anderson, G.; Jacka, F.; et al. Oxidative & nitrosative stress in depression: Why so much stress? Neurosci. Biobehav. R 2014, 45, 46–62. [Google Scholar]
- Liu, T.; Zhong, S.; Liao, X.; Chen, J.; He, T.; Lai, S.; Jia, Y. A Meta-Analysis of Oxidative Stress Markers in Depression. PLoS ONE 2015, 10, e0138904. [Google Scholar] [CrossRef] [PubMed]
- Ștefănescu, C.; Ciobica, A. The relevance of oxidative stress status in first episode and recurrent depression. J. Affect. Disord. 2012, 143, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Michalakeas, C.; Parissis, J.; Douzenis, A.; Nikolaou, M.; Varounis, C.; Andreadou, I.; Antonellos, N.; Markantonis-Kiroudis, S.; Paraskevaidis, I.; Ikonomidis, I.; et al. Effects of sertraline on circulating markers of oxidative stress in depressed patients with chronic heart failure: A pilot study. J. Card. Fail. 2011, 17, 748–754. [Google Scholar] [CrossRef]
- Chung, C.; Schmidt, D.; Stein, C.; Morrow, J.; Salomon, R. Increased oxidative stress in patients with depression and its relationship to treatment. Psychiatry Res. 2013, 206, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Tas, F.; Hansel, H.; Belce, A.; İlvan, Ş.; Argon, A.; Çamlıca, H.; Topuz, E. Oxidative stress in breast cancer. Med. Oncol. 2005, 22, 11–15. [Google Scholar] [CrossRef]
- Sastre-Serra, J.; Valle, A.; Company, M.; Garau, I.; Oliver, J.; Roca, P. Estrogen down-regulates uncoupling proteins and increases oxidative stress in breast cancer. Free. Radic. Biol. Med. 2010, 48, 506–512. [Google Scholar] [CrossRef]
- Binsaleh, N.K.; Sherwani, S.; Eltayeb, R.; Qanash, H.; Bazaid, A.S.; Althobiti, M.; Hazzazi, M.S.; Rajendrasozhan, S. Increased inflammatory cytokines and oxidative stress enhanced antibody production in breast and prostate cancer patients with COVID-19 related depression. Front. Chem. 2023, 11, 1192074. [Google Scholar] [CrossRef] [PubMed]
- Longobucco, Y.; Masini, A.; Marini, S.; Barone, G.; Fimognari, C.; Bragonzoni, L.; Dallolio, L.; Maffei, F. Exercise and Oxidative Stress Biomarkers among Adult with Cancer: A Systematic Review. Oxid. Med. Cell. Longev. 2022, 2022, 2097318. [Google Scholar] [CrossRef]
- Knop, K.; Schwan, R.; Bongartz, M.; Bloch, W.; Brixius, K.; Baumann, F. Sport and oxidative stress in oncological patients. Int. J. Sports Med. 2011, 32, 960–964. [Google Scholar] [CrossRef]
- Luca, M.; Luca, A. Oxidative Stress-Related Endothelial Damage in Vascular Depression and Vascular Cognitive Impairment: Beneficial Effects of Aerobic Physical Exercise. Oxid. Med. Cell. Longev. 2019, 2019, 8067045. [Google Scholar] [CrossRef] [PubMed]
- Agudelo, L.Z.; Femenía, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal muscle PGC-1α1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef]
- Santos, T.M.; Kolling, J.; Siebert, C.; Biasibetti, H.; Bertó, C.; Grun, L.; Dalmaz, C.; Barbé-Tuana, F.; Wyse, A. Effects of previous physical exercise to chronic stress on long-term aversive memory and oxidative stress in amygdala and hippocampus of rats. Int. J. Dev. Neurosci. 2017, 56, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhou, C. Corticosterone reduces brain mitochondrial function and expression of mitofusin, BDNF in depression-like rodents regardless of exercise preconditioning. Psychoneuroendocrino 2012, 37, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.D.; Lopez, J.F.; Lyons, D.M.; Burke, S.; Wallace, M.; Schatzberg, A.F. Glucocorticoid and mineralocorticoid receptor mRNA expression in squirrel monkey brain. J. Psychiatr. Res. 2000, 34, 383–392. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Reul, J.M. Feedback action and tonic influence of corticosteroids on brain function: A concept arising from the heterogeneity of brain receptor systems. Psychoneuroendocrinology 1987, 12, 83–105. [Google Scholar] [CrossRef] [PubMed]
- de Kloet, E.R.; Oitzl, M.S.; Joëls, M. Stress and cognition: Are corticosteroids good or bad guys? Trends Neurosci. 1999, 22, 422–426. [Google Scholar] [CrossRef]
- Ali, F.; Nemeroff, C. Neuroendocrine Alterations in Major Depressive Disorder. In Major Depressive Disorder; Elsevier: Amsterdam, The Netherlands, 2020; pp. 63–74. [Google Scholar]
- Iob, E.; Kirschbaum, C.; Steptoe, A. Persistent depressive symptoms, HPA-axis hyperactivity, and inflammation: The role of cognitive-affective and somatic symptoms. Mol. Psychiatry 2019, 25, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, F.; Capoccia, S.; Berry, A.; Raggi, C.; Vomero, M.; Ortona, E.; Fabi, A.; Pugliese, P. Increased Cortisol Secretion, Immune Activation and Mood Changes in Breast Cancer Patients Following Surgery and Adjuvant Chemotherapy. Eur. Psychiatry 2015, 30, 28–31. [Google Scholar] [CrossRef]
- Hsiao, F.-H.; Jow, G.; Kuo, W.; Chang, K.-J.; Liu, Y.-F.; Ho, R.; Ng, S.; Chan, C.; Lai, Y.; Chen, Y.-T. The Effects of Psychotherapy on Psychological Well-Being and Diurnal Cortisol Patterns in Breast Cancer Survivors. Psychother. Psychosom. 2012, 81, 173–182. [Google Scholar] [CrossRef]
- Schumacher, M.M.; Santambrogio, J. Cortisol and the Dexamethasone Suppression Test as a Biomarker for Melancholic Depression: A Narrative Review. J. Pers. Med. 2023, 13, 837. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.J.; Chen, C.; Scullen, T.; Zsombok, A.; Salahudeen, A.A.; Di, S.; Herman, J.P.; Tasker, J.G. Sensitization of the Hypothalamic-Pituitary-Adrenal Axis in a Male Rat Chronic Stress Model. Endocrinology 2016, 157, 2346–2355. [Google Scholar] [CrossRef]
- Saleem, P.T. Dexamethasone Suppression Test in Depressive Illness: Its Relation to Anxiety Symptoms. Br. J. Psychiatry 1984, 144, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Herbert, J. Cortisol and depression: Three questions for psychiatry. Psychol. Med. 2013, 43, 449–469. [Google Scholar] [CrossRef] [PubMed]
- Germain, A.; Kupfer, D.J. Circadian rhythm disturbances in depression. Hum. Psychopharmacol. 2008, 23, 571–585. [Google Scholar] [CrossRef]
- Leal-Cerro, A.; Gippini, A.; Amaya, M.J.; Lage, M.; Mato, J.A.; Diéguez, C.; Casanueva, F. Mechanisms underlying the neuroendocrine response to physical exercise. J. Endocrinol. Investig. 2003, 26, 879–885. [Google Scholar] [CrossRef]
- Tsatsoulis, A.; Fountoulakis, S. The Protective Role of Exercise on Stress System Dysregulation and Comorbidities. Ann. N. Y. Acad. Sci. 2006, 1083, 196–213. [Google Scholar] [CrossRef]
- Marsh, J.; Scanes, C. Neuroendocrine-Immune Interactions. J. Neuroimmunol. 1994, 124, 1049–1061. [Google Scholar] [CrossRef]
- Rao, R.; Vadiraja, H.; Nagaratna, R.; Gopinath, K.; Patil, S.; Diwakar, R.; Shahsidhara, H.P.; Ajaikumar, B.; Nagendra, H. Effect of Yoga on Sleep Quality and Neuroendocrine Immune Response in Metastatic Breast Cancer Patients. Indian J. Palliat. Care 2017, 23, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Saxton, J.; Scott, E.; Daley, A.; Woodroofe, M.; Mutrie, N.; Crank, H.; Powers, H.; Coleman, R. Effects of an exercise and hypocaloric healthy eating intervention on indices of psychological health status, hypothalamic-pituitary-adrenal axis regulation and immune function after early-stage breast cancer: A randomised controlled trial. Breast Cancer Res. BCR 2014, 16, R39. [Google Scholar] [CrossRef] [PubMed]
- Nabkasorn, C.; Miyai, N.; Sootmongkol, A.; Junprasert, S.; Yamamoto, H.; Arita, M.; Miyashita, K. Effects of physical exercise on depression, neuroendocrine stress hormones and physiological fitness in adolescent females with depressive symptoms. Eur. J. Public Health 2006, 16, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Liu, Y.; Li, W.; Yang, B.; Chen, D.; Wang, X.; Jiang, Z.; Wang, H.; Wang, Z.; Cornelisson, G.; et al. Beneficial effects of exercise and its molecular mechanisms on depression in rats. Behav. Brain Res. 2006, 168, 47–55. [Google Scholar] [CrossRef]
- Jauhar, S.; Cowen, P.J.; Browning, M. Fifty years on: Serotonin and depression. J. Psychopharmacol. 2023, 37, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Hüfner, K.; Oberguggenberger, A.; Kohl, C.; Geisler, S.; Gamper, E.; Meraner, V.; Egeter, J.; Hubalek, M.; Beer, B.; Fuchs, D.; et al. Levels in neurotransmitter precursor amino acids correlate with mental health in patients with breast cancer. Psychoneuroendocrinology 2015, 60, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Parrott, J.M.; Redus, L.; Santana-Coelho, D.; Morales, J.; Gao, X.; O’Connor, J. Neurotoxic kynurenine metabolism is increased in the dorsal hippocampus and drives distinct depressive behaviors during inflammation. Transl. Psychiatry 2016, 6, e918. [Google Scholar] [CrossRef] [PubMed]
- Labadie, B.W.; Bao, R.; Luke, J.J. Reimagining IDO Pathway Inhibition in Cancer Immunotherapy via Downstream Focus on the Tryptophan-Kynurenine-Aryl Hydrocarbon Axis. Clin. Cancer Res. 2019, 25, 1462–1471. [Google Scholar] [CrossRef]
- Garrison, A.; Parrott, J.M.; Tuñon, A.; Delgado, J.; Redus, L.; O’Connor, J. Kynurenine pathway metabolic balance influences microglia activity: Targeting kynurenine monooxygenase to dampen neuroinflammation. Psychoneuroendocrino 2018, 94, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Asghar, K.; Loya, A.; Rana, I.A.; Tahseen, M.; Ishaq, M.; Farooq, A.; Bakar, M.A.; Masood, I. Indoleamine 2,3-dioxygenase expression and overall survival in patients diagnosed with breast cancer in Pakistan. Cancer Manag. Res. 2019, 11, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Dill, E.; Dillon, P.; Bullock, T.; Mills, A. IDO expression in breast cancer: An assessment of 281 primary and metastatic cases with comparison to PD-L1. Mod. Pathol. 2018, 31, 1513–1522. [Google Scholar] [CrossRef]
- Caperuto, E.C.; Santos, R.D.; Mello, M.; Rosa, L.F.B.P.C. Effect of endurance training on hypothalamic serotonin concentration and performance. Clin. Exp. Pharmacol. Physiol. 2009, 36, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Chaouloff, F. Effects of acute physical exercise on central serotonergic systems. Med. Sci. Sports Exerc. 1997, 29, 58–62. [Google Scholar] [CrossRef]
- Flora, R.; Theodorus, T.; Zulkarnain, M.; Juliansyah, R.A.; Syokumawena, S. Effect Of Anaerobic And Aerobic Exercise Toward Serotonin In Rat Brain Tissue. J. Neurobehav. Sci. 2016, 3, 3–6. [Google Scholar] [CrossRef]
- Kondo, M.; Shimada, S. Serotonin and exercise-induced brain plasticity. Neurotransmitter 2015, 2, e793. [Google Scholar] [CrossRef]
- Yuan, T.; Paes, F.; Arias-Carrión, Ó.; Rocha, N.B.F.; de Sá Filho, A.S.; Machado, S. Neural Mechanisms of Exercise: Anti-Depression, Neurogenesis, and Serotonin Signaling. CNS Neurol. Disord. Drug Targets 2015, 14, 1307–1311. [Google Scholar] [CrossRef]
- Payne, J.; Held, J.; Thorpe, J.; Shaw, H. Effect of exercise on biomarkers, fatigue, sleep disturbances, and depressive symptoms in older women with breast cancer receiving hormonal therapy. Oncol. Nurs. Forum 2008, 35, 635–642. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Tang, W.; Tang, W.; Kao, X.-L.; Zhang, C.-G.; Wong, X.-T. Exercise Intervention May Prevent Depression. Int. J. Sports Med. 2012, 33, 525–530. [Google Scholar] [CrossRef]
- Liu, W.; Sheng, H.; Xu, Y.; Liu, Y.; Lu, J.; Ni, X. Swimming exercise ameliorates depression-like behavior in chronically stressed rats: Relevant to proinflammatory cytokines and IDO activation. Behav. Brain Res. 2013, 242, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Buch, A.M.; Liston, C. Dissecting diagnostic heterogeneity in depression by integrating neuroimaging and genetics. Neuropsychopharmacol 2021, 46, 156–175. [Google Scholar]
- Solleiro-Villavicencio, H.; Rivas-Arancibia, S. Effect of Chronic Oxidative Stress on Neuroinflammatory Response Mediated by CD4+T Cells in Neurodegenerative Diseases. Front. Cell Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, Y.; Hunter, R.; Wei, Y.; Blumenthal, R.; Falke, C.; Khairova, R.; Zhou, R.; Yuan, P.; Machado-Vieira, R.; et al. Dynamic regulation of mitochondrial function by glucocorticoids. Proc. Natl. Acad. Sci. USA 2009, 106, 3543–3548. [Google Scholar] [CrossRef] [PubMed]
- Spiers, J.G.; Chen, H.-J.C.; Sernia, C.; Lavidis, N.A. Activation of the hypothalamic-pituitary-adrenal stress axis induces cellular oxidative stress. Front. Neurosci. 2014, 8, 456. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Adcock, I.M. Oxidative stress and redox regulation of lung inflammation in COPD. Eur. Respir. J. 2006, 28, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Ribaudo, G.; Bortoli, M.; Witt, C.E.; Parke, B.; Mena, S.; Oselladore, E.; Zagotto, G.; Hashemi, P.; Orian, L. ROS-Scavenging Selenofluoxetine Derivatives Inhibit In Vivo Serotonin Reuptake. ACS Omega 2022, 7, 8314–8322. [Google Scholar] [CrossRef]
- Correia, A.S.; Cardoso, A.; Vale, N. Significant Differences in the Reversal of Cellular Stress Induced by Hydrogen Peroxide and Corticosterone by the Application of Mirtazapine or L-Tryptophan. Int. J. Transl. Med. 2022, 2, 482–505. [Google Scholar] [CrossRef]
- Liu, B.; Liu, J.; Wang, M.; Zhang, Y.; Li, L. From Serotonin to Neuroplasticity: Evolvement of Theories for Major Depressive Disorder. Front. Cell Neurosci. 2017, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Hassamal, S. Chronic stress, neuroinflammation, and depression: An overview of pathophysiological mechanisms and emerging anti-inflammatories. Front. Psychiatry 2023, 14, 1130989. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, N.A.L.; Del Ángel, D.S.; Brizuela, N.O.; Peraza, A.V.; Olguín, H.J.; Soto, M.P.; Guzmán, D.C. Inflammatory Process and Immune System in Major Depressive Disorder. Int. J. Neuropsychopharmacol. 2022, 25, 46–53. [Google Scholar] [CrossRef]
- Guglielmo, R.; de Filippis, R.; Ouanes, S.; Hasler, G. Editorial: The glutamate hypothesis of mood disorders: Neuroplasticity processes, clinical features, treatment perspectives. Front. Psychiatry 2022, 13, 1054887. [Google Scholar] [CrossRef]
- Han, K.-M.; Ham, B.-J. How Inflammation Affects the Brain in Depression: A Review of Functional and Structural MRI Studies. J. Clin. Neurol. 2021, 17, 503–515. [Google Scholar] [CrossRef]
- Wang, X.; Wang, N.; Zhong, L.L.D.; Su, K.; Wang, S.; Zheng, Y.; Yang, B.; Zhang, J.; Pan, B.; Yang, W.; et al. Development and Validation of a Risk Prediction Model for Breast Cancer Prognosis Based on Depression-Related Genes. Front. Oncol. 2022, 12, 879563. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, N.; Zhong, L.; Wang, S.; Zheng, Y.; Yang, B.; Zhang, J.; Lin, Y.; Wang, Z. Prognostic value of depression and anxiety on breast cancer recurrence and mortality: A systematic review and meta-analysis of 282,203 patients. Mol. Psychiatry 2020, 25, 3186–3197. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Liu, H.; Gao, J. Side Effects of Endocrine Therapy Are Associated with Depression and Anxiety in Breast Cancer Patients Accepting Endocrine Therapy: A Cross-Sectional Study in China. Front. Psychol. 2022, 13, 905459. [Google Scholar] [CrossRef]
- Castro-Figueroa, E.M.; Acevedo, K.I.; Peña-Vargas, C.I.; Torres-Blasco, N.; Flores, I.; Colón-Echevarria, C.B.; Maldonado, L.; Rodríguez, Z.; Aquino-Acevedo, A.N.; Jim, H.; et al. Depression, Anxiety, and Social Environmental Adversity as Potential Modulators of the Immune Tumor Microenvironment in Breast Cancer Patients. Med. Sci. 2021, 9, 46. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Zheng, X.; Liu, D.; Huang, R.; Ge, H.; Cheng, L.; Zhang, M.; Cheng, H. Physical Activity and Depression in Breast Cancer Patients: Mechanisms and Therapeutic Potential. Curr. Oncol. 2025, 32, 77. https://doi.org/10.3390/curroncol32020077
Li A, Zheng X, Liu D, Huang R, Ge H, Cheng L, Zhang M, Cheng H. Physical Activity and Depression in Breast Cancer Patients: Mechanisms and Therapeutic Potential. Current Oncology. 2025; 32(2):77. https://doi.org/10.3390/curroncol32020077
Chicago/Turabian StyleLi, Anlong, Xinyi Zheng, Dajie Liu, Runze Huang, Han Ge, Ling Cheng, Mingjun Zhang, and Huaidong Cheng. 2025. "Physical Activity and Depression in Breast Cancer Patients: Mechanisms and Therapeutic Potential" Current Oncology 32, no. 2: 77. https://doi.org/10.3390/curroncol32020077
APA StyleLi, A., Zheng, X., Liu, D., Huang, R., Ge, H., Cheng, L., Zhang, M., & Cheng, H. (2025). Physical Activity and Depression in Breast Cancer Patients: Mechanisms and Therapeutic Potential. Current Oncology, 32(2), 77. https://doi.org/10.3390/curroncol32020077