tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer

 and
and
Abstract
1. Introduction
2. Methods
2.1. Data Collection
2.2. Identification and Quantification of 5′-tRFs in PRAD
2.3. Quantification of mRNA Expression Levels
2.4. Identification of 5′-tRFs Dysregulated in PRAD
2.5. Inference of Potential Functions of 5′-tRFs
2.6. Construction of Prognostic Predictor of 5′-tRFs
2.7. Identification of Tumor Subtypes Based on tRF Expression
2.8. Mutational Data Analysis
2.9. Statistical Analysis
3. Results
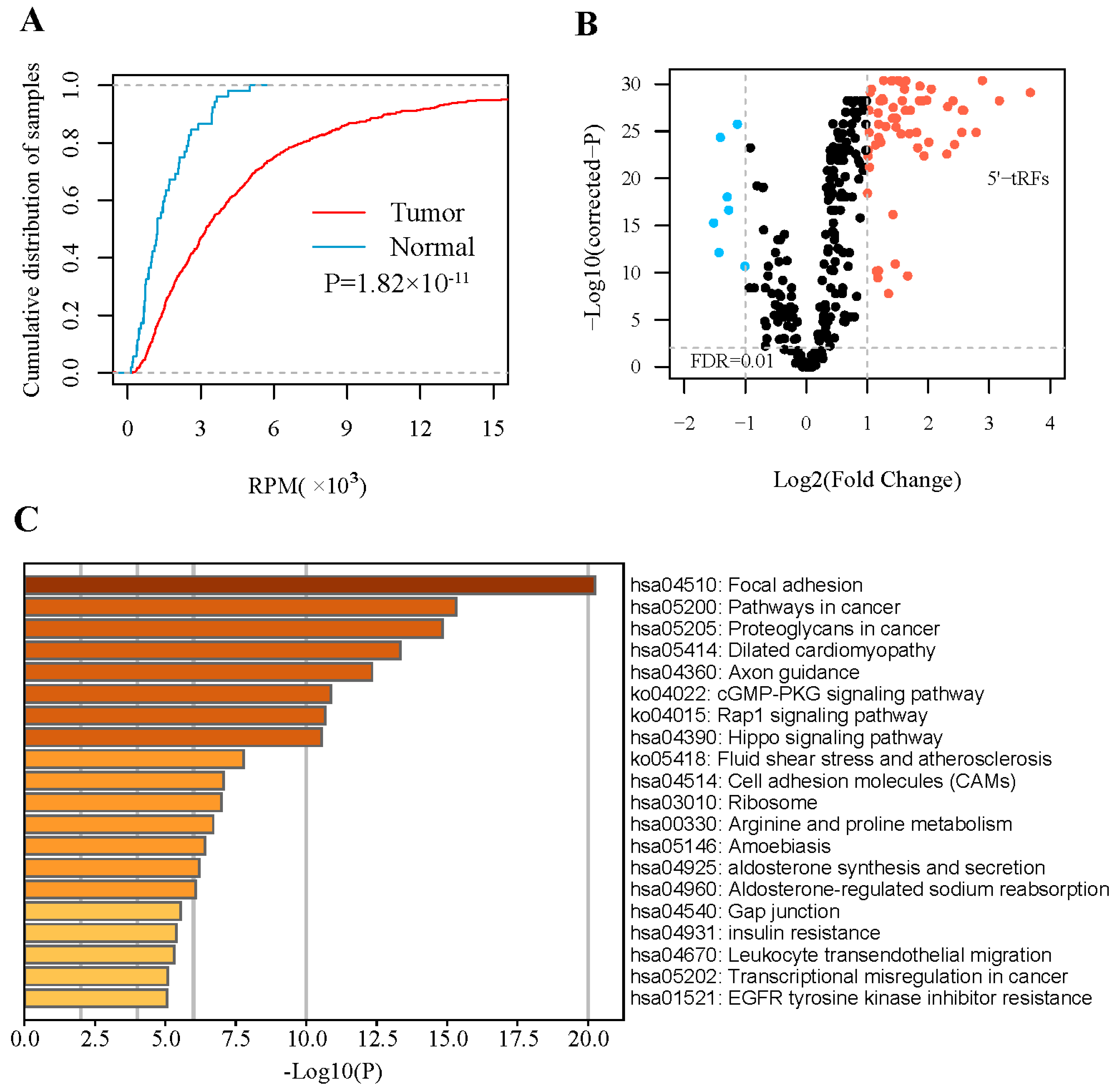
3.1. 5′- tRFs Are Dysregulated in PRAD
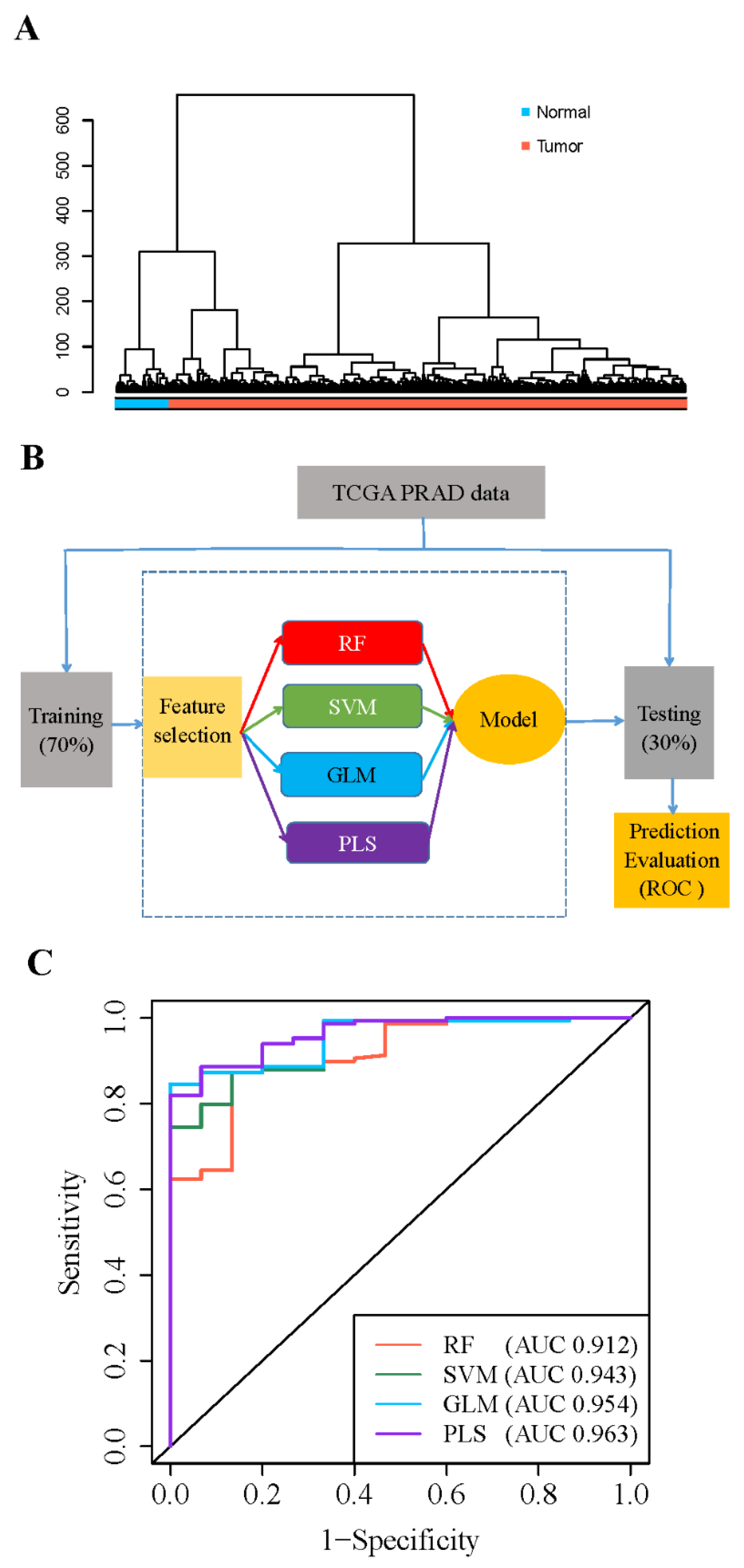
3.2. 5′- tRFs Are Novel Biomarkers for Diagnosis of PRAD
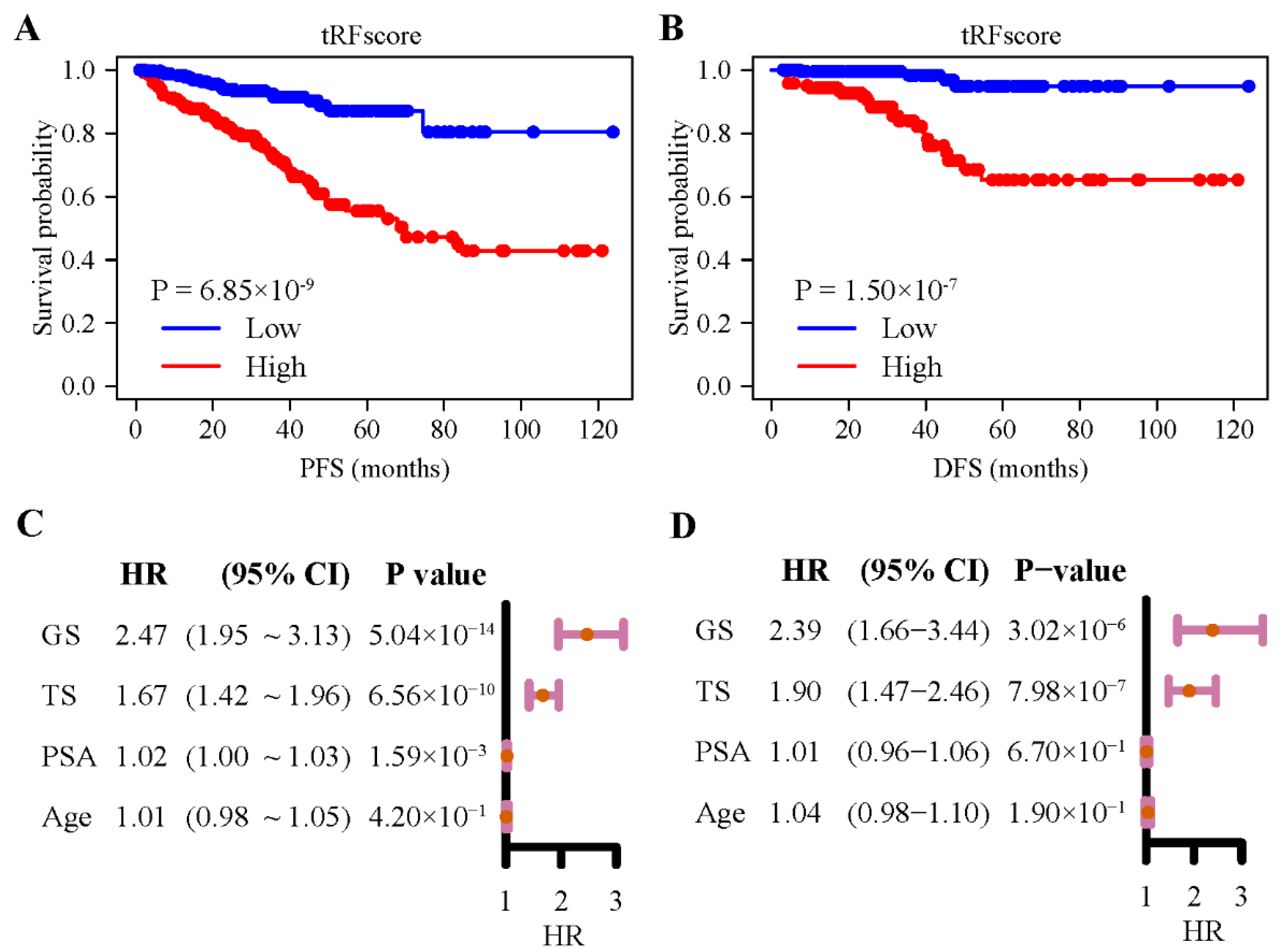
3.3. 5′- tRFs Are Novel Biomarkers for Prognosis of PRAD
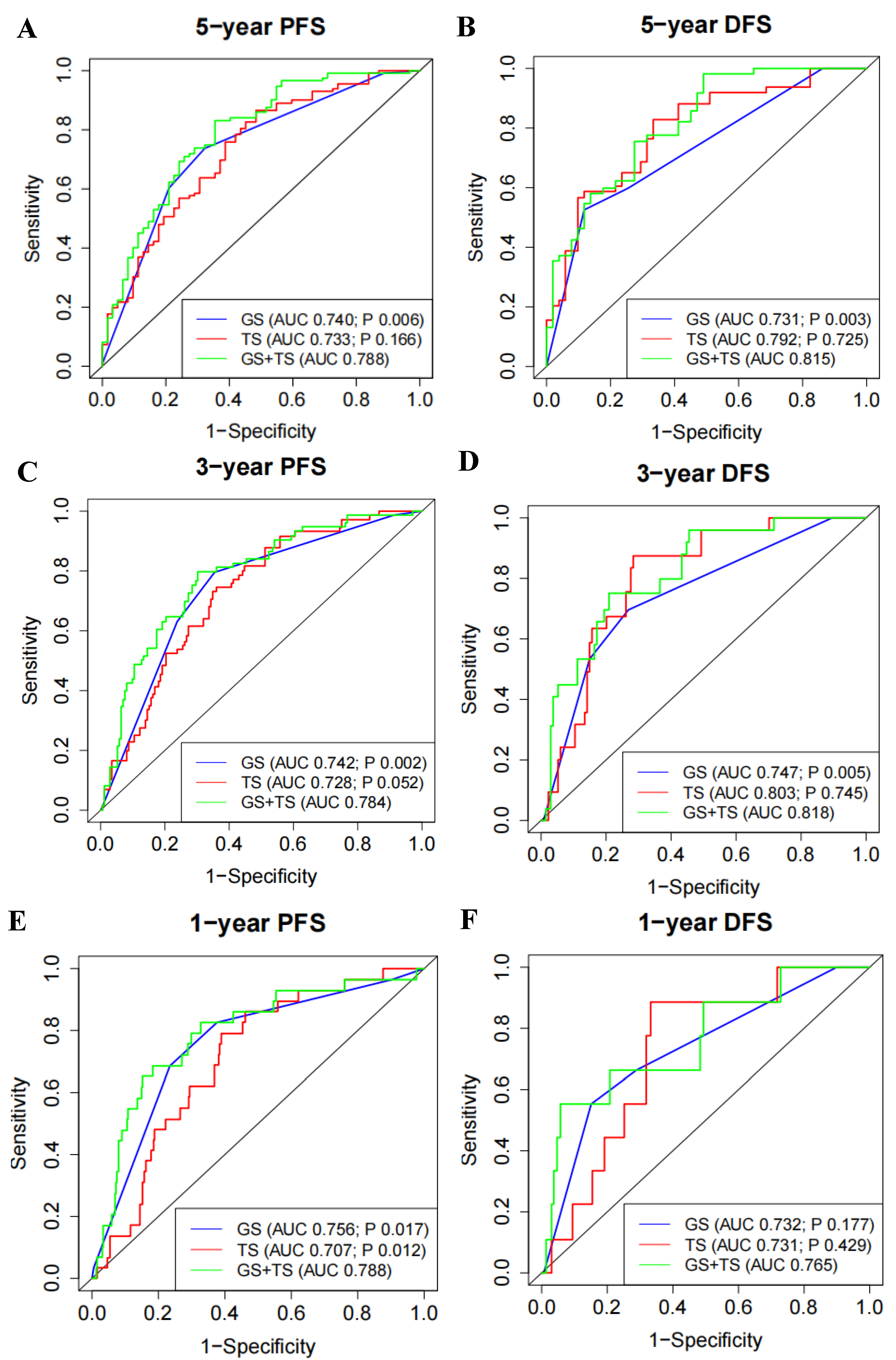
3.4. 5′- tRFs Provide Independent Prognostic Information for PRAD
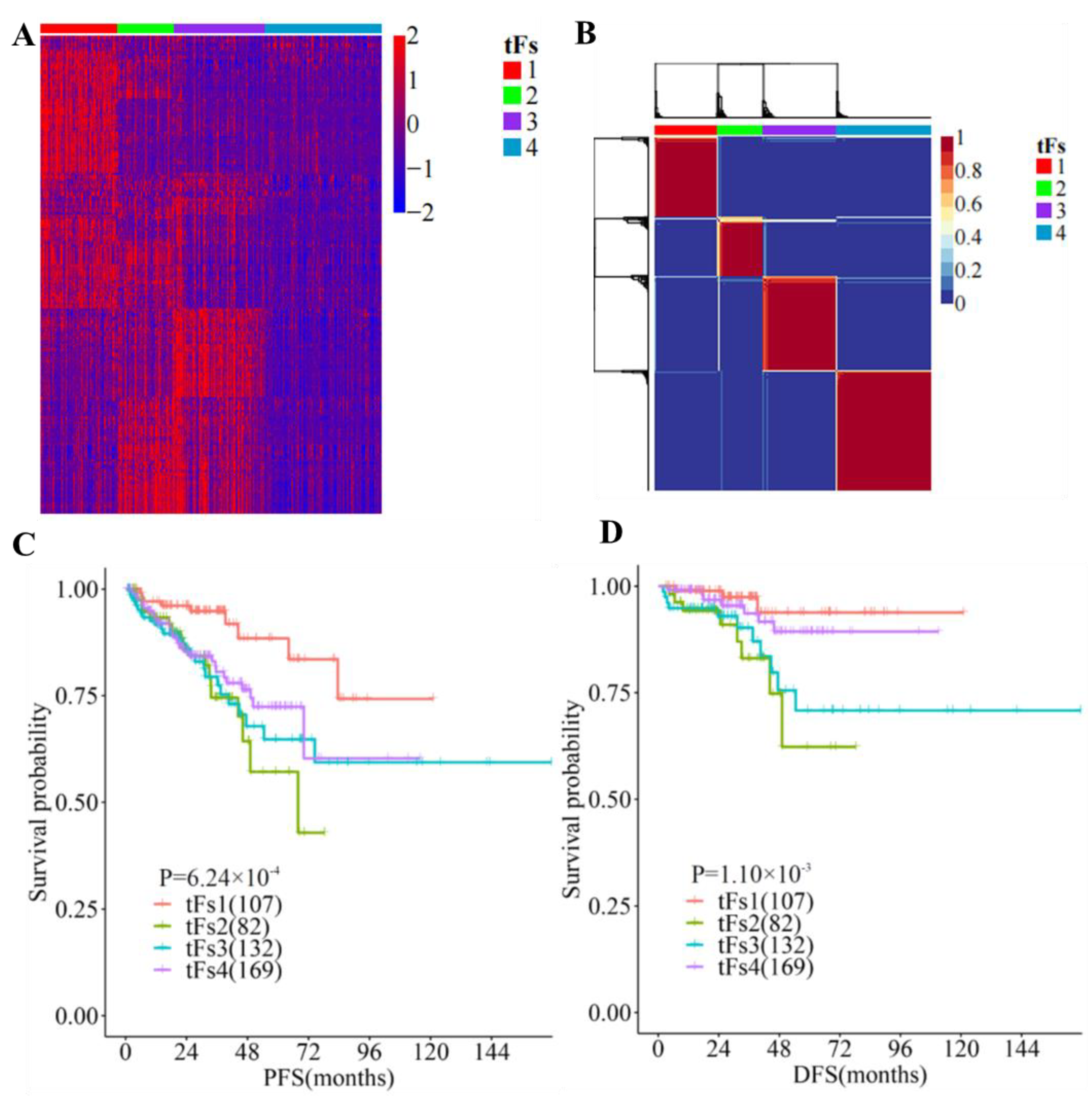
3.5. 5′- tRFs Are Novel Biomarkers for the Tumor Classification of PRAD
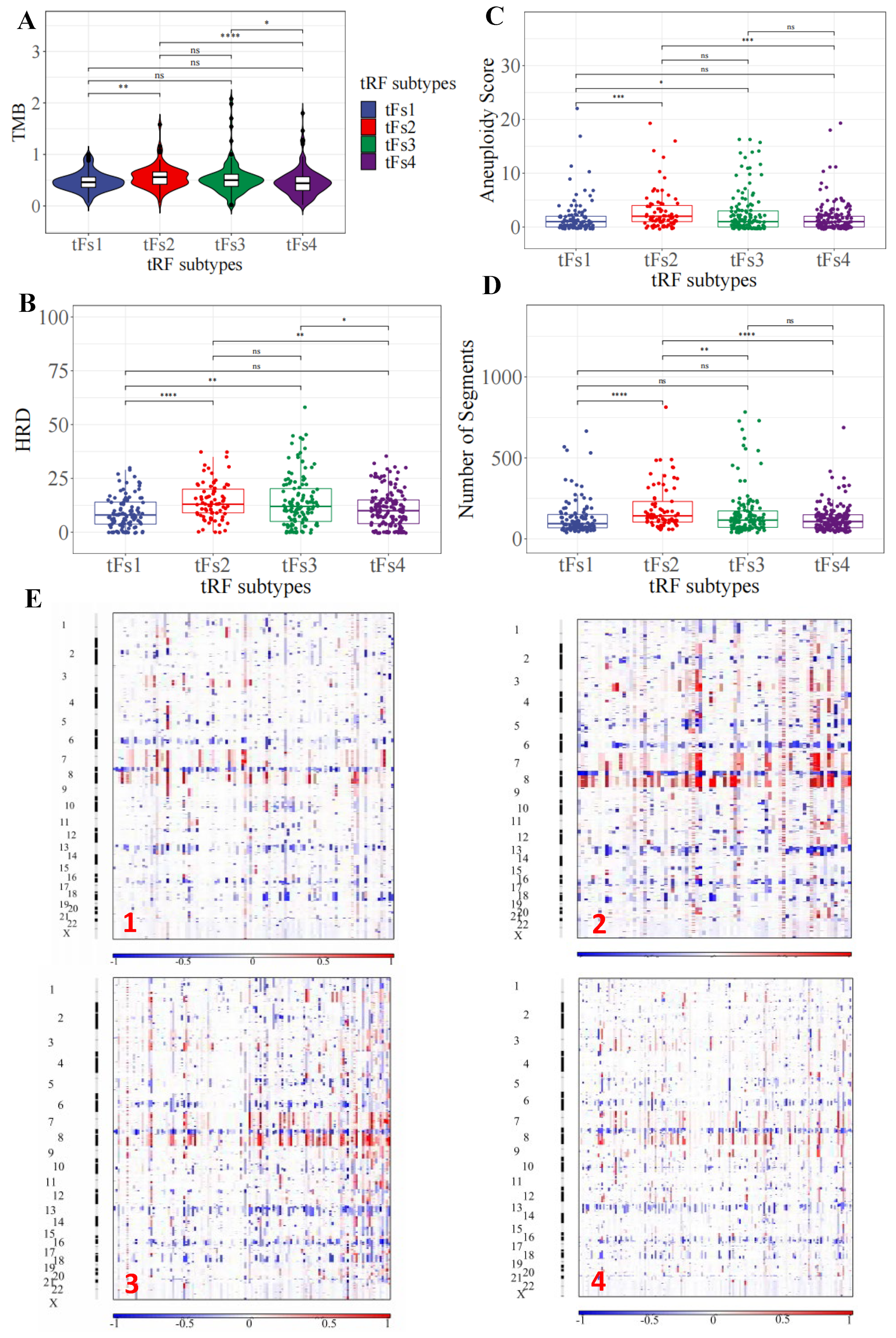
3.6. Genomic Landscapes of 5′-tRFs Tumor Subtypes of PRAD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| tRFs | tRNA-derived RNA fragments |
| PRAD | prostate adenocarcinoma |
| PSA | serum prostate-specific antigen |
| ncRNAs | small noncoding RNAs |
| ANG | angiogenin |
| TCGA | The Cancer Genome Atlas |
| ICGC | International Cancer Genome Consortium |
| BWA | burrows-wheeler transform |
| RPM | reads per million |
| RPKM | read per kilobase per million |
| FDR | false discovery rate |
| RF | random forest |
| SVM | support vector machine |
| GLM | generalized linear model |
| PLS | partial least squares |
| ROC | receiver operation characteristic |
| GBA | guilt by association |
| GO | gene ontology |
| KEGG | Kyoto encyclopedia of genes and genomes |
| PFS | progression-free survival |
| HR | hazard ratio |
| CI | confidence interval |
| LASSOl | east absolute shrinkage and selection operator |
| NMF | non-negative matrix factorization |
| IQRs | interquartile ranges |
| SNP | single nucleotide polymorphism |
| INS | small insertions |
| DEL | deletions |
| TMB | tumor mutational burden |
| HRD | homologous recombination deficiency |
| AS | aneuploidy score |
| SD | standard deviation |
| TS | tRF score |
| GS | Gleason score |
| DFS | disease-free survival |
References
- Xia, C.; Dong, X.; Li, H.; Cao, M.; Sun, D.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Li, N.; et al. Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. 2022, 135, 584–590. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, R.; Sant, M.; Coleman, M.P.; Francisci, S.; Baili, P.; Pierannunzio, D.; Trama, A.; Visser, O.; Brenner, H.; Ardanaz, E.; et al. Cancer survival in Europe 1999–2007 by country and age: Results of EUROCARE-5-a population-based study. Lancet Oncol. 2014, 15, 23–34. [Google Scholar] [CrossRef]
- Virgo, K.S.; Basch, E.; Loblaw, D.A.; Oliver, T.K.; Rumble, R.B.; Carducci, M.A.; Nordquist, L.; Taplin, M.-E.; Winquist, E.; Singer, E.A. Second-Line Hormonal Therapy for Men with Chemotherapy-Naïve, Castration-Resistant Prostate Cancer: American Society of Clinical Oncology Provisional Clinical Opinion. J. Clin. Oncol. 2017, 35, 1952–1964. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.S.; Li, J.; Lavery, H.J.; Yadav, K.K.; Tewari, A.K. Next-generation sequencing technology in prostate cancer diagnosis, prognosis, and personalized treatment. Urol. Oncol. 2015, 33, 267.e1–267.e13. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Nonomura, N. Urinary biomarkers of prostate cancer. Int. J. Urol. 2018, 25, 770–779. [Google Scholar] [CrossRef]
- Di Minno, A.; Aveta, A.; Gelzo, M.; Tripodi, L.; Pandolfo, S.D.; Crocetto, F.; Imbimbo, C.; Castaldo, G. 8-Hydroxy-2-Deoxyguanosine and 8-Iso-Prostaglandin F2alpha: Putative Biomarkers to assess Oxidative Stress Damage Following Robot-Assisted Radical Prostatectomy (RARP). J. Clin. Med. 2022, 11, 6102. [Google Scholar] [CrossRef]
- Kristiansen, G. Diagnostic and prognostic molecular biomarkers for prostate cancer. Histopathology 2012, 60, 125–141. [Google Scholar] [CrossRef]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef]
- Yu, M.; Lu, B.; Zhang, J.; Ding, J.; Liu, P.; Lu, Y. tRNA-derived RNA fragments in cancer: Current status and future perspectives. J. Hematol. Oncol. 2020, 13, 121. [Google Scholar] [CrossRef]
- Phizicky, E.M.; Hopper, A.K. tRNA biology charges to the front. Genes Dev. 2010, 24, 1832–1860. [Google Scholar] [CrossRef]
- Sun, C.; Fu, Z.; Wang, S.; Li, J.; Li, Y.; Zhang, Y.; Yang, F.; Chu, J.; Wu, H.; Huang, X.; et al. Roles of tRNA-derived fragments in human cancers. Cancer Lett. 2018, 414, 16–25. [Google Scholar] [CrossRef]
- Yamasaki, S.; Ivanov, P.; Hu, G.-F.; Anderson, P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef]
- Sun, X.; Yang, J.; Yu, M.; Yao, D.; Zhou, L.; Li, X.; Qiu, Q.; Lin, W.; Lu, B.; Chen, E.; et al. Global identification and characterization of tRNA-derived RNA fragment landscapes across human cancers. NAR Cancer 2020, 2, zcaa031. [Google Scholar] [CrossRef]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef]
- Goodarzi, H.; Liu, X.H.; Nguyen, H.C.B.; Zhang, S.; Fish, L.; Tavazoie, S.F. Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell 2015, 161, 790–802. [Google Scholar] [CrossRef]
- Kim, H.K.; Fuchs, G.; Wang, S.; Wei, W.; Zhang, Y.; Park, H.; Roy-Chaudhuri, B.; Li, P.; Xu, J.; Chu, K.; et al. A transfer-RNA-derived small RNA regulates ribosome biogenesis. Nature 2017, 552, 57–62. [Google Scholar] [CrossRef]
- Chu, X.; He, C.; Sang, B.; Yang, C.; Yin, C.; Ji, M.; Qian, A.; Tian, Y. Transfer RNAs-derived small RNAs and their application potential in multiple diseases. Front. Cell Dev. Biol. 2022, 10, 954431. [Google Scholar] [CrossRef]
- Honda, S.; Loher, P.; Shigematsu, M.; Palazzo, J.P.; Suzuki, R.; Imoto, I.; Rigoutsos, I.; Kirino, Y. Sex hormone-dependent tRNA halves enhance cell proliferation in breast and prostate cancers. Proc. Natl. Acad. Sci. USA 2015, 112, E3816–E3825. [Google Scholar] [CrossRef]
- Yang, C.; Lee, M.; Song, G.; Lim, W. tRNA(Lys)-Derived Fragment Alleviates Cisplatin-Induced Apoptosis in Prostate Cancer Cells. Pharmaceutics 2021, 13, 55. [Google Scholar] [CrossRef]
- Balatti, V.; Nigita, G.; Veneziano, D.; Drusco, A.; Stein, G.S.; Messier, T.L.; Farina, N.H.; Lian, J.B.; Tomasello, L.; Liu, C.-G.; et al. tsRNA signatures in cancer. Proc. Natl. Acad. Sci. USA 2017, 114, 8071–8076. [Google Scholar] [CrossRef]
- Pekarsky, Y.; Balatti, V.; Palamarchuk, A.; Rizzotto, L.; Veneziano, D.; Nigita, G.; Rassenti, L.Z.; Pass, H.I.; Kipps, T.J.; Liu, C.-G.; et al. Dysregulation of a family of short noncoding RNAs, tsRNAs, in human cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 5071–5076. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Sun, X.; Zhou, L.; Amanullah, M.; Pan, X.; Liu, Y.; Liang, M.; Liu, P.; Lu, Y. OncotRF: An online resource for exploration of tRNA-derived fragments in human cancers. RNA Biol. 2020, 17, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, S.; Vosa, U.; van der Graaf, A.; Franke, L.; de Magalhaes, J.P. Faculty Opinions recommendation of Gene co-expression analysis for functional classification and gene-disease predictions. Brief Bioinform. 2018, 19, 575–592. [Google Scholar] [CrossRef] [PubMed]
- Knijnenburg, T.A.; Wang, L.; Zimmermann, M.T.; Chambwe, N.; Gao, G.F.; Cherniack, A.D.; Fan, H.; Shen, H.; Way, G.P.; Greene, C.S.; et al. Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas. Cell Rep. 2018, 23, 239–254.e6. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689.e3. [Google Scholar] [CrossRef]
- Li, X.; Heyer, W.-D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008, 18, 99–113. [Google Scholar] [CrossRef]
- Ok Atilgan, A.; Ozdemir, B.H.; Yilmaz Akcay, E.; Tepeoglu, M.; Borcek, P.; Dirim, A. Association between focal adhesion kinase and matrix metalloproteinase-9 expression in prostate adenocarcinoma and their influence on the progression of prostatic adenocarcinoma. Ann. Diagn. Pathol. 2020, 45, 151480. [Google Scholar] [CrossRef]
- Marx, A.; Schumann, A.; Höflmayer, D.; Bady, E.; Hube-Magg, C.; Möller, K.; Tsourlakis, M.C.; Steurer, S.; Büscheck, F.; Eichenauer, T.; et al. Up regulation of the Hippo signalling effector YAP1 is linked to early biochemical recurrence in prostate cancers. Sci. Rep. 2020, 10, 8916. [Google Scholar] [CrossRef]
- Munkley, J.; Mills, I.G.; Elliott, D.J. The role of glycans in the development and progression of prostate cancer. Nat. Rev. Urol. 2016, 13, 324–333. [Google Scholar] [CrossRef]
- Fay, E.K.; Graff, J.N. Immunotherapy in Prostate Cancer. Cancers 2020, 12, 1752. [Google Scholar] [CrossRef]
- Karnes, R.J.; Cheville, J.C.; Ida, C.M.; Sebo, T.J.; Nair, A.A.; Tang, H.; Munz, J.-M.; Kosari, F.; Vasmatzis, G. The Ability of Biomarkers to Predict Systemic Progression in Men with High-Risk Prostate Cancer Treated Surgically Is Dependent on ERG Status. Cancer Res. 2010, 70, 8994–9002. [Google Scholar] [CrossRef]
- Feng, F.Y.; Brenner, J.C.; Hussain, M.; Chinnaiyan, A.M. Molecular Pathways: Targeting ETS Gene Fusions in Cancer. Clin. Cancer Res. 2014, 20, 4442–4448. [Google Scholar] [CrossRef]
- Ray-Coquard, I.; Pautier, P.; Pignata, S.; Pérol, D.; González-Martín, A.; Berger, R.; Fujiwara, K.; Vergote, I.; Colombo, N.; Mäenpää, J.; et al. Olaparib plus Bevacizumab as First-Line Maintenance in Ovarian Cancer. New Engl. J. Med. 2019, 381, 2416–2428. [Google Scholar] [CrossRef]
- Zhao, E.Y.; Shen, Y.; Pleasance, E.; Kasaian, K.; Leelakumari, S.; Jones, M.; Bose, P.; Ch’Ng, C.; Reisle, C.; Eirew, P.; et al. Homologous Recombination Deficiency and Platinum-Based Therapy Outcomes in Advanced Breast Cancer. Clin. Cancer Res. 2017, 23, 7521–7530. [Google Scholar] [CrossRef]
- Sun, C.; Yang, F.; Zhang, Y.; Chu, J.; Wang, J.; Wang, Y.; Zhang, Y.; Li, J.; Li, Y.; Fan, R.; et al. tRNA-Derived Fragments as Novel Predictive Biomarkers for Trastuzumab-Resistant Breast Cancer. Cell. Physiol. Biochem. 2018, 49, 419–431. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tF-1 (n = 107) | tF-2 (n = 82) | tF-3 (n = 132) | tF-4 (n = 169) | p Value | |
|---|---|---|---|---|---|
| Age | 60.60 ± 6.72 | 61.41 ± 6.37 | 61.60 ± 6.78 | 60.54 ± 7.01 | 0.744 |
| PSA | 9.22 ± 9.42 | 11.68 ± 8.83 | 12.74 ± 15.95 | 10.53 ± 11.8 | 0.020 |
| Gleason score | 2.07 × 10−6 | ||||
| 6 | 17 (15.89%) | 2 (2.44%) | 13 (9.85%) | 13 (7.69%) | |
| 7 | 69 (64.49%) | 36 (43.90%) | 51 (38.64%) | 87 (51.48%) | |
| 8 | 13 (12.15%) | 9 (10.98%) | 17 (12.88%) | 25 (14.79%) | |
| 9/10 | 8 (7.47%) | 35 (42.68%) | 51 (38.63%) | 44 (26.04%) | |
| Grading group | 3.93 × 10−7 | ||||
| 1 | 17 (15.89%) | 2 (2.44%) | 13 (9.85%) | 13 (7.69%) | |
| 2 | 47 (43.93%) | 15 (18.29%) | 31 (23.48%) | 51 (30.18%) | |
| 3 | 22 (20.56%) | 21 (25.61%) | 20 (15.15%) | 36 (21.30%) | |
| 4 | 13 (12.15%) | 9 (10.98%) | 17 (12.88%) | 25 (14.79%) | |
| 5 | 8 (7.47%) | 35 (42.68%) | 51 (38.64%) | 44 (26.04%) |
| tF-1 (n = 89) | tF-2 (n = 52) | tF-3 (n = 38) | tF-4 (n = 147) | p Value | |
|---|---|---|---|---|---|
| ERG (fusion) | 1.24 × 10−4 | ||||
| Yes | 34(38.20%) | 12(23.08%) | 16(42.10%) | 84(57.14%) | |
| No | 55(61.80%) | 40(76.92%) | 22(57.90%) | 63(42.86%) | |
| FOXA1 (mutation) | |||||
| Yes | 5(5.62%) | 5(9.62%) | 1(2.63%) | 2(1.36%) | 0.038 |
| No | 84(94.38%) | 47(90.38%) | 37(97.37%) | 145(98.64%) | |
| KMT2D (mutation) | 0.0018 | ||||
| Yes | 1(1.12%) | 7(13.46%) | 1(2.63%) | 2(1.36%) | |
| No | 88(98.88%) | 45(86.54%) | 37(97.37%) | 145(98.64%) | |
| ZMYM3 (mutation) | 0.063 | ||||
| Yes | 0(0.00%) | 3(5.77%) | 1(2.63%) | 2(1.36%) | |
| No | 89(100%) | 49(94.23%) | 37(97.37%) | 145(98.64%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Yu, M.; Cheng, S.; Zhou, X.; Li, J.; Lu, Y.; Liu, P.; Ding, S. tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer. Curr. Oncol. 2023, 30, 981-999. https://doi.org/10.3390/curroncol30010075
Liu W, Yu M, Cheng S, Zhou X, Li J, Lu Y, Liu P, Ding S. tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer. Current Oncology. 2023; 30(1):981-999. https://doi.org/10.3390/curroncol30010075
Chicago/Turabian StyleLiu, Weigang, Mengqian Yu, Sheng Cheng, Xiaoxu Zhou, Jia Li, Yan Lu, Pengyuan Liu, and Shiping Ding. 2023. "tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer" Current Oncology 30, no. 1: 981-999. https://doi.org/10.3390/curroncol30010075
APA StyleLiu, W., Yu, M., Cheng, S., Zhou, X., Li, J., Lu, Y., Liu, P., & Ding, S. (2023). tRNA-Derived RNA Fragments Are Novel Biomarkers for Diagnosis, Prognosis, and Tumor Subtypes in Prostate Cancer. Current Oncology, 30(1), 981-999. https://doi.org/10.3390/curroncol30010075