High Expression of POGK Predicts Poor Prognosis in Patients with Hepatocellular Carcinoma

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. POGK Expression Validation
2.2. Patient Data Source and Pprocessing
2.3. Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
2.4. Gene Set Enrichment Analysis (GSEA)
2.5. Statistical Analysis
3. Results
3.1. POGK Expression Analysis
3.2. Baseline Characteristics of Patients
3.3. Correlation between POGK Expression and Clinical Characteristics
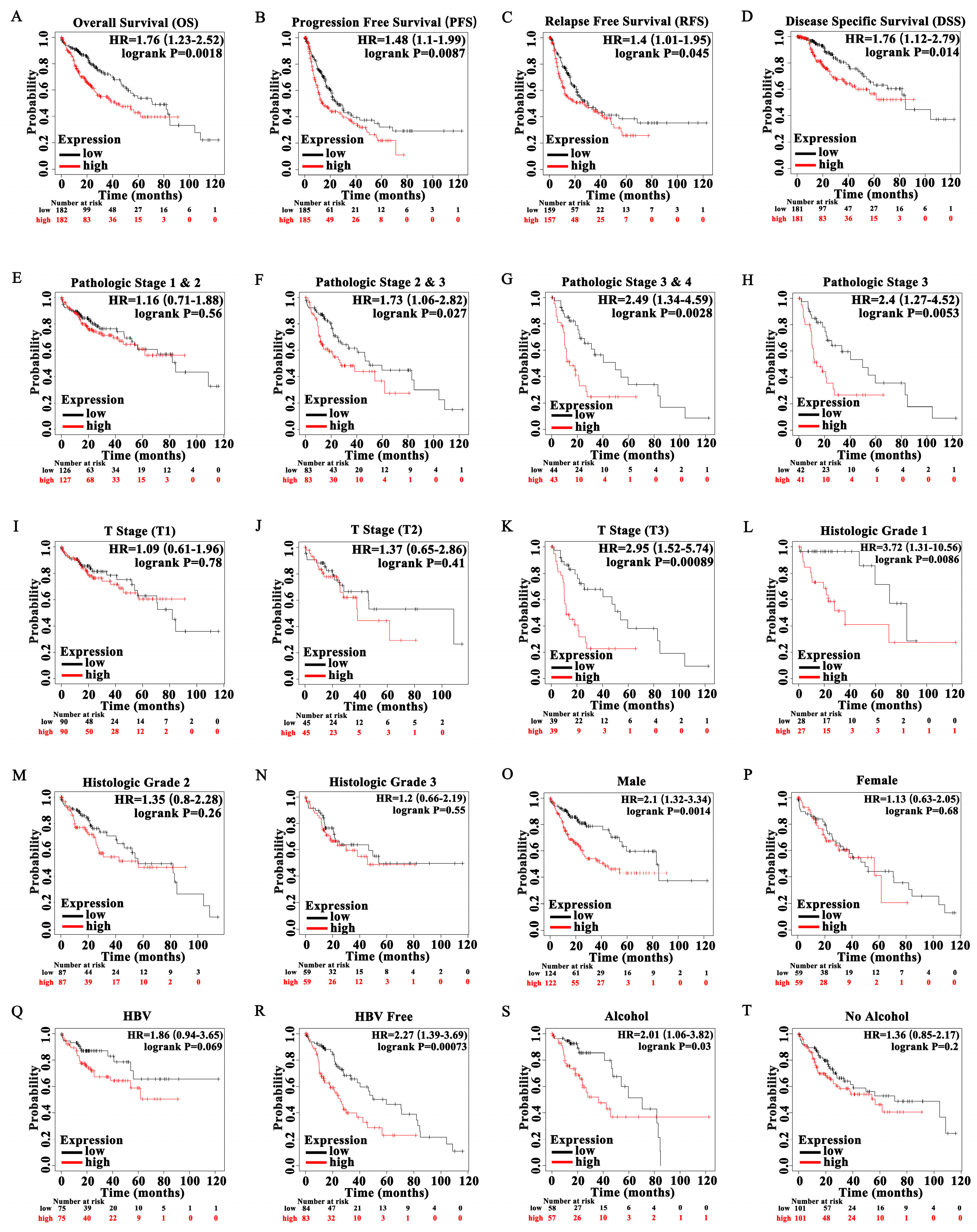
3.4. High Expression of POGK Is a Risk Factor for Survival in HCC
3.5. Diagnostic Value of POGK Gene Expression in HCC
3.6. Functional Enrichment and Analyses of POGK Gene in HCC by GO Analysis
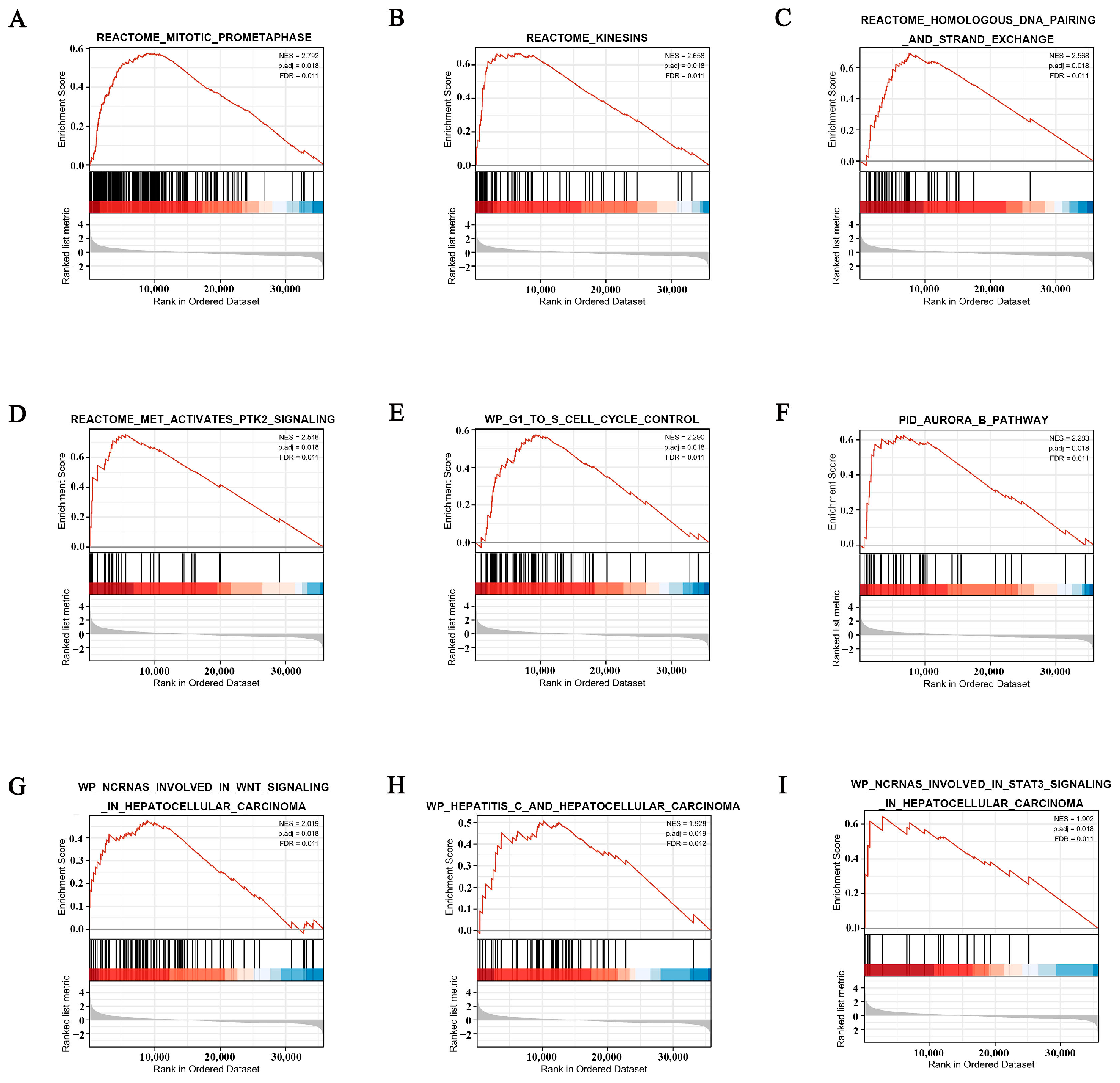
3.7. POGK-Related Signaling Pathways Identified by GSEA
3.8. Correlation between POGK Expression and Immune Infiltration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- De Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef]
- Liu, J.; Liang, W.; Jing, W.; Liu, M. Countdown to 2030: Eliminating hepatitis B disease, China. Bull. World Health Organ. 2019, 97, 230–238. [Google Scholar] [CrossRef]
- Feng, R.-M.; Zong, Y.-N.; Cao, S.-M.; Xu, R.-H. Current cancer situation in China: Good or bad news from the 2018 Global Cancer Statistics? Cancer Commun. 2019, 39, 1–12. [Google Scholar] [CrossRef]
- Aravalli, R.N.; Cressman, E.N.K.; Steer, C.J. Cellular and molecular mechanisms of hepatocellular carcinoma: An update. Arch. Toxicol. 2012, 87, 227–247. [Google Scholar] [CrossRef]
- Jiang, Y.; Han, Q.-J.; Zhang, J. Hepatocellular carcinoma: Mechanisms of progression and immunotherapy. World J. Gastroenterol. 2019, 25, 3151–3167. [Google Scholar] [CrossRef]
- De Sio, F.R.S. Kruppel-associated box (KRAB) proteins in the adaptive immune system. Nucleus 2014, 5, 138–148. [Google Scholar] [CrossRef]
- Ziv, Y.; Bielopolski, D.; Galanty, Y.; Lukas, C.; Taya, Y.; Schultz, D.C.; Lukas, J.; Bekker-Jensen, S.; Bartek, J.; Shiloh, Y. Chromatin relaxation in response to DNA double-strand breaks is modulated by a novel ATM- and KAP-1 dependent pathway. Nat. Cell Biol. 2006, 8, 870–876. [Google Scholar] [CrossRef]
- Noon, A.T.; Shibata, A.; Rief, N.; Löbrich, M.; Stewart, G.; Jeggo, P.A.; Goodarzi, A.A. 53BP1-dependent robust localized KAP-1 phosphorylation is essential for heterochromatic DNA double-strand break repair. Nat. Cell Biol. 2010, 12, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-C.; Tsai, C.Y.; Chang, W.-C.; Lee, W.-H.; Wang, J.-M. RB·E2F1 Complex Mediates DNA Damage Responses through Transcriptional Regulation of ZBRK1. J. Biol. Chem. 2010, 285, 33134–33143. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-F.; Chuang, C.-H.; Li, C.-F.; Liao, C.-C.; Cheng, C.-P.; Cheng, T.-L.; Shen, M.-R.; Tseng, J.T.; Chang, W.-C.; Lee, W.-H.; et al. ZBRK1 Acts as a Metastatic Suppressor by Directly Regulating MMP9 in Cervical Cancer. Cancer Res. 2010, 70, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; O’Herrin, S.M.; Wu, J.; Reagan-Shaw, S.; Ma, Y.; Bhat, K.M.; Gravekamp, C.; Setaluri, V.; Peters, N.; Hoffmann, F.M.; et al. MAGE-A, mMage-b, and MAGE-C Proteins Form Complexes with KAP1 and Suppress p53-Dependent Apoptosis in MAGE-Positive Cell Lines. Cancer Res. 2007, 67, 9954–9962. [Google Scholar] [CrossRef] [PubMed]
- Yokoe, T.; Toiyama, Y.; Okugawa, Y.; Tanaka, K.; Ohi, M.; Inoue, Y.; Mohri, Y.; Miki, C.; Kusunoki, M. KAP1 Is Associated With Peritoneal Carcinomatosis in Gastric Cancer. Ann. Surg. Oncol. 2009, 17, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Machnik, M.; Cylwa, R.; Kiełczewski, K.; Biecek, P.; Liloglou, T.; Mackiewicz, A.; Oleksiewicz, U. The expression signature of cancer-associated KRAB—ZNF factors identified in TCGA pan-cancer transcriptomic data. Mol. Oncol. 2018, 13, 701–724. [Google Scholar] [CrossRef] [PubMed]
- Sobocińska, J.; Molenda, S.; Machnik, M.; Oleksiewicz, U. KRAB-ZFP Transcriptional Regulators Acting as Oncogenes and Tumor Suppressors: An Overview. Int. J. Mol. Sci. 2021, 22, 2212. [Google Scholar] [CrossRef]
- Puzakov, M.V.; Puzakova, L.V.; Cheresiz, S.V.; Sang, Y. The IS630/Tc1/mariner transposons in three ctenophore genomes. Mol. Phylogenetics Evol. 2021, 163, 107231. [Google Scholar] [CrossRef]
- Dupeyron, M.; Baril, T.; Bass, C.; Hayward, A. Phylogenetic analysis of the Tc1/mariner superfamily reveals the unexplored diversity of pogo-like elements. Mob. DNA 2020, 11, 21. [Google Scholar] [CrossRef]
- Liu, Y.; Zong, W.; Diaby, M.; Lin, Z.; Wang, S.; Gao, B.; Ji, T.; Song, C. Diversity and Evolution of pogo and Tc1/mariner Transposons in the Apoidea Genomes. Biology 2021, 10, 940. [Google Scholar] [CrossRef]
- Gao, B.; Sang, Y.; Zong, W.; Diaby, M.; Shen, D.; Wang, S.; Wang, Y.; Chen, C.; Song, C. Evolution and domestication of Tc1/mariner transposons in the genome of African coelacanth (Latimeria chalumnae). Genome 2020, 63, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Wang, Y.; Diaby, M.; Zong, W.; Shen, D.; Wang, S.; Chen, C.; Wang, X.; Song, C. Evolution of pogo, a separate superfamily of IS630-Tc1-mariner transposons, revealing recurrent domestication events in vertebrates. Mob. DNA 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Liu, Y.; Sun, H.; Jia, J.; Wu, T.; Ding, R.; Cheng, X. Identification of abnormally high expression of POGZ as a new biomarker associated with a poor prognosis in osteosarcoma. Eur. J. Histochem. 2021, 65, 3264. [Google Scholar] [CrossRef]
- Pangon, L.; Ng, I.; Giry-Laterriere, M.; Currey, N.; Morgan, A.D.; Benthani, F.; Tran, P.N.; Alsohaily, S.; Segelov, E.; Parker, B.L.; et al. JRK is a positive regulator of β-catenin transcriptional activity commonly overexpressed in colon, breast and ovarian cancer. Oncogene 2015, 35, 2834–2841. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Gong, S.; Osterhoff, G.; Schopow, N. A Novel Four-Gene Prognostic Signature for Prediction of Survival in Patients with Soft Tissue Sarcoma. Cancers 2021, 13, 5837. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, J.; Wei, H.; Liu, Y.; Dai, X.; Xue, J.; Shen, T.; Liu, X. Serum Biomarker Panel for Rapid Early Diagnosis of Lung Cancer. Curr. Cancer Drug Targets 2022. online ahead of print. [Google Scholar] [CrossRef]
- Wintergerst, L.; Selmansberger, M.; Maihöfer, C.; Schüttrumpf, L.; Walch, A.; Wilke, C.; Pitea, A.; Woischke, C.; Baumeister, P.; Kirchner, T.; et al. A prognostic mRNA expression signature of four 16q24.3 genes in radio(chemo)therapy-treated head and neck squamous cell carcinoma (HNSCC). Mol. Oncol. 2018, 12, 2085–2101. [Google Scholar] [CrossRef]
- Wang, H.; Hartswood, E.; Finnegan, D.J. Pogo transposase contains a putative helix-turn-helix DNA binding domain that recognises a 12 bp sequence within the terminal inverted repeats. Nucleic Acids Res. 1999, 27, 455–461. [Google Scholar] [CrossRef]
- Ecco, G.; Imbeault, M.; Trono, D. KRAB zinc finger proteins. Development 2017, 144, 2719–2729. [Google Scholar] [CrossRef]
- Wen, N.; Cai, Y.; Li, F.; Ye, H.; Tang, W.; Song, P.; Cheng, N. The clinical management of hepatocellular carcinoma worldwide: A concise review and comparison of current guidelines: 2022 update. Biosci. Trends 2022, 16, 20–30. [Google Scholar] [CrossRef]
- Rizzo, A.; Ricci, A.D.; Gadaleta-Caldarola, G.; Brandi, G. First-line immune checkpoint inhibitor-based combinations in unresectable hepatocellular carcinoma: Current management and future challenges. Expert Rev. Gastroenterol. Hepatol. 2021, 15, 1245–1251. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Low POGK Expression | High POGK Expression | p Value |
|---|---|---|---|
| n | 187 | 187 | |
| T stage, n (%) | 0.341 | ||
| T1 | 99 (26.7%) | 84 (22.6%) | |
| T2 | 45 (12.1%) | 50 (13.5%) | |
| T3 | 34 (9.2%) | 46 (12.4%) | |
| T4 | 6 (1.6%) | 7 (1.9%) | |
| N stage, n (%) | 0.625 | ||
| N0 | 120 (46.5%) | 134 (51.9%) | |
| N1 | 1 (0.4%) | 3 (1.2%) | |
| M stage, n (%) | 0.361 | ||
| M0 | 130 (47.8%) | 138 (50.7%) | |
| M1 | 3 (1.1%) | 1 (0.4%) | |
| Pathologic stage, n (%) | 0.132 | ||
| Stage I | 94 (26.9%) | 79 (22.6%) | |
| Stage II | 44 (12.6%) | 43 (12.3%) | |
| Stage III | 35 (10%) | 50 (14.3%) | |
| Stage IV | 4 (1.1%) | 1 (0.3%) | |
| Tumor status, n (%) | 0.036 * | ||
| Tumor free | 111 (31.3%) | 91 (25.6%) | |
| With tumor | 66 (18.6%) | 87 (24.5%) | |
| Gender, n (%) | 0.377 | ||
| Female | 56 (15%) | 65 (17.4%) | |
| Male | 131 (35%) | 122 (32.6%) | |
| Race, n (%) | 0.025 * | ||
| Asian | 65 (18%) | 95 (26.2%) | |
| Black or African American | 9 (2.5%) | 8 (2.2%) | |
| White | 102 (28.2%) | 83 (22.9%) | |
| Age, n (%) | 0.133 | ||
| ≤60 | 81 (21.7%) | 96 (25.7%) | |
| >60 | 106 (28.4%) | 90 (24.1%) | |
| Weight, n (%) | 0.002 ** | ||
| ≤70 | 78 (22.5%) | 106 (30.6%) | |
| >70 | 97 (28%) | 65 (18.8%) | |
| Height, n (%) | 0.678 | ||
| <170 | 99 (29%) | 102 (29.9%) | |
| ≥170 | 73 (21.4%) | 67 (19.6%) | |
| BMI, n (%) | 0.033 * | ||
| ≤25 | 79 (23.4%) | 98 (29.1%) | |
| >25 | 91 (27%) | 69 (20.5%) | |
| Residual tumor, n (%) | 0.083 | ||
| R0 | 170 (49.3%) | 157 (45.5%) | |
| R1 | 5 (1.4%) | 12 (3.5%) | |
| R2 | 1 (0.3%) | 0 (0%) | |
| Histologic grade, n (%) | <0.001 *** | ||
| G1 | 39 (10.6%) | 16 (4.3%) | |
| G2 | 102 (27.6%) | 76 (20.6%) | |
| G3 | 39 (10.6%) | 85 (23%) | |
| G4 | 5 (1.4%) | 7 (1.9%) | |
| Adjacent hepatic tissue inflammation, n (%) | 0.092 | ||
| None | 69 (29.1%) | 49 (20.7%) | |
| Mild | 45 (19%) | 56 (23.6%) | |
| Severe | 11 (4.6%) | 7 (3%) | |
| AFP (ng/mL), n (%) | <0.001 *** | ||
| ≤400 | 123 (43.9%) | 92 (32.9%) | |
| >400 | 21 (7.5%) | 44 (15.7%) | |
| Albumin (g/dL), n (%) | 0.947 | ||
| <3.5 | 38 (12.7%) | 31 (10.3%) | |
| ≥3.5 | 124 (41.3%) | 107 (35.7%) | |
| Prothrombin time, n (%) | 0.145 | ||
| ≤4 | 103 (34.7%) | 105 (35.4%) | |
| >4 | 53 (17.8%) | 36 (12.1%) | |
| Child-Pugh grade, n (%) | 0.902 | ||
| A | 121 (50.2%) | 98 (40.7%) | |
| B | 11 (4.6%) | 10 (4.1%) | |
| C | 1 (0.4%) | 0 (0%) | |
| Fibrosis ishak score, n (%) | 0.329 | ||
| 0 | 46 (21.4%) | 29 (13.5%) | |
| 1/2 | 15 (7%) | 16 (7.4%) | |
| 3/4 | 12 (5.6%) | 16 (7.4%) | |
| 5/6 | 45 (20.9%) | 36 (16.7%) | |
| Vascular invasion, n (%) | 0.245 | ||
| No | 114 (35.8%) | 94 (29.6%) | |
| Yes | 52 (16.4%) | 58 (18.2%) | |
| OS event, n (%) | 0.158 | ||
| Alive | 129 (34.5%) | 115 (30.7%) | |
| Dead | 58 (15.5%) | 72 (19.3%) | |
| DSS event, n (%) | 0.414 | ||
| Alive | 148 (40.4%) | 139 (38%) | |
| Dead | 36 (9.8%) | 43 (11.7%) | |
| PFI event, n (%) | 0.148 | ||
| Alive | 103 (27.5%) | 88 (23.5%) | |
| Dead | 84 (22.5%) | 99 (26.5%) | |
| Age, median (IQR) | 63 (53.5, 69) | 60 (51, 68) | 0.091 |
| Characteristics | Total (N) | Odds Ratio (OR) | p Value |
|---|---|---|---|
| T stage (T2 and T3 and T4 vs. T1) | 371 | 1.428 (0.950–2.153) | 0.087 |
| N stage (N1 vs. N0) | 258 | 2.687 (0.339–54.709) | 0.395 |
| M stage (M1 vs. M0) | 272 | 0.314 (0.015–2.487) | 0.318 |
| Pathologic stage (Stage II, Stage III, and Stage IV vs. Stage I) | 350 | 1.348 (0.886–2.055) | 0.164 |
| Tumor status (With tumor vs. Tumor free) | 355 | 1.608 (1.055–2.461) | 0.028 * |
| Gender (Female vs. Male) | 374 | 1.246 (0.808–1.927) | 0.320 |
| Age (>60 vs. ≤60) | 373 | 0.716 (0.476–1.076) | 0.109 |
| Race (White vs. Asian and Black or African American) | 362 | 0.585 (0.385–0.885) | 0.011 * |
| Weight (>70 vs. ≤70) | 346 | 0.493 (0.320–0.756) | 0.001 ** |
| Height (≥170 vs. <170) | 341 | 0.891 (0.578–1.372) | 0.600 |
| BMI (>25 vs. ≤25) | 337 | 0.611 (0.396–0.939) | 0.025 * |
| Residual tumor (R1 and R2 vs. R0) | 345 | 2.166 (0.820–6.348) | 0.131 |
| Histologic grade (G3 and G4 vs. G1 and G2) | 369 | 3.205 (2.064–5.033) | <0.001 *** |
| Adjacent hepatic tissue inflammation (Severe and Mild vs. None) | 237 | 1.584 (0.950–2.656) | 0.079 |
| AFP (ng/mL) (>400 vs. ≤400) | 280 | 2.801 (1.576–5.110) | <0.001 *** |
| Albumin (g/dL) (≥3.5 vs. <3.5) | 300 | 1.058 (0.617–1.824) | 0.839 |
| Prothrombin time (>4 vs. ≤4) | 297 | 0.666 (0.401–1.099) | 0.114 |
| Child-Pugh grade (B and C vs. A) | 241 | 1.029 (0.418–2.484) | 0.949 |
| Fibrosis ishak score (3/4 and 5/6 vs. 0 and 1/2) | 215 | 1.237 (0.722–2.123) | 0.439 |
| Vascular invasion (Yes vs. No) | 318 | 1.353 (0.852–2.154) | 0.201 |
| Characteristics | Total (N) | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|---|
| Hazard Ratio (95% CI) | p Value | Hazard Ratio (95% CI) | p Value | ||
| Age | 373 | ||||
| ≤60 | 177 | Reference | |||
| >60 | 196 | 1.205 (0.850–1.708) | 0.295 | ||
| Gender | 373 | ||||
| Male | 253 | Reference | |||
| Female | 121 | 1.261 (0.885–1.796) | 0.200 | ||
| BMI | 336 | ||||
| ≤25 | 177 | Reference | |||
| >25 | 160 | 0.798 (0.550–1.158) | 0.235 | ||
| T stage | 370 | ||||
| T1 | 183 | Reference | |||
| T2 and T3 and T4 | 188 | 2.126 (1.481–3.052) | <0.001 *** | 0.865 (0.118–6.362) | 0.886 |
| N stage | 258 | ||||
| N0 | 254 | Reference | |||
| N1 | 4 | 2.029 (0.497–8.281) | 0.324 | ||
| M stage | 272 | ||||
| M0 | 268 | Reference | |||
| M1 | 4 | 4.077 (1.281–12.973) | 0.017 * | 2.176 (0.508–9.328) | 0.295 |
| Pathologic stage | 349 | ||||
| Stage I | 173 | Reference | |||
| Stage II and Stage III and Stage IV | 177 | 2.090 (1.429–3.055) | <0.001 *** | 2.690 (0.355–20.400) | 0.338 |
| Tumor status | 354 | ||||
| Tumor free | 202 | Reference | |||
| With tumor | 153 | 2.317 (1.590–3.376) | <0.001 *** | 1.921 (1.203–3.066) | 0.006 ** |
| Histologic grade | 368 | ||||
| G1 and G2 | 233 | Reference | |||
| G3 and G4 | 136 | 1.091 (0.761–1.564) | 0.636 | ||
| Adjacent hepatic tissue inflammation | 236 | ||||
| None | 118 | Reference | |||
| Mild and Severe | 119 | 1.194 (0.734–1.942) | 0.475 | ||
| AFP (ng/mL) | 279 | ||||
| ≤400 | 215 | Reference | |||
| >400 | 65 | 1.075 (0.658–1.759) | 0.772 | ||
| Albumin (g/dL) | 299 | ||||
| <3 5 | 69 | Reference | |||
| ≥3 5 | 231 | 0.897 (0.549–1.464) | 0.662 | ||
| Prothrombin time | 296 | ||||
| ≤4 | 208 | Reference | |||
| >4 | 89 | 1.335 (0.881–2.023) | 0.174 | ||
| Child-Pugh grade | 240 | ||||
| A | 219 | Reference | |||
| B and C | 22 | 1.643 (0.811–3.330) | 0.168 | ||
| Fibrosis ishak score | 214 | ||||
| 0 and 1/2 | 106 | Reference | |||
| 3/4 and 5/6 | 109 | 0.740 (0.445–1.232) | 0.247 | ||
| Vascular invasion | 317 | ||||
| No | 208 | Reference | |||
| Yes | 110 | 1.344 (0.887–2.035) | 0.163 | ||
| POGK | 373 | ||||
| Low | 187 | Reference | |||
| High | 187 | 1.582 (1.112–2.249) | 0.011 * | 1.550 (0.973–2.471) | 0.065 |
| Gene Set Name | Size | ES | NES | p.adjust | q Values |
|---|---|---|---|---|---|
| Mitotic Prometaphase | 202 | 0.576702 | 2.79224 | 0.017522659 | 0.011278 |
| Kinesins | 61 | 0.671025 | 2.657603 | 0.017522659 | 0.011278 |
| Homologous DNA Pairing and Strand Exchange | 42 | 0.692287 | 2.568165 | 0.017522659 | 0.011278 |
| MET Activates PTK2 Signaling | 30 | 0.752825 | 2.545744 | 0.017522659 | 0.011278 |
| G1 to S Cell Cycle Control | 64 | 0.573636 | 2.289791 | 0.017522659 | 0.011278 |
| Aurora B Pathway | 39 | 0.623436 | 2.282505 | 0.017522659 | 0.011278 |
| ncRNAs Involved in WNT Signaling in Hepatocellular Carcinoma | 86 | 0.474281 | 2.018823 | 0.017522659 | 0.011278 |
| Hepatitis C and Hepatocellular Carcinoma | 50 | 0.506931 | 1.928244 | 0.019218103 | 0.012369 |
| ncRNAs Involved in STAT3 Signaling in Hepatocellular Carcinoma | 17 | 0.644878 | 1.902398 | 0.017522659 | 0.011278 |
| Cells | Coefficient of Correlation (Pearson) | p Value (Pearson) | Coefficient of Correlation (Spearman) | p Value (Spearman) |
|---|---|---|---|---|
| aDC | 0.080 | 0.122 | 0.056 | 0.279 |
| B cells | −0.060 | 0.247 | −0.081 | 0.119 |
| CD8 T cells | −0.167 | 0.001 ** | −0.180 | <0.001 *** |
| Cytotoxic cells | −0.399 | <0.001 *** | −0.427 | <0.001 *** |
| DC | −0.319 | <0.001 *** | −0.356 | <0.001 *** |
| Eosinophils | 0.071 | 0.169 | 0.070 | 0.176 |
| iDC | −0.050 | 0.334 | −0.085 | 0.100 |
| Macrophages | 0.088 | 0.090 | 0.059 | 0.251 |
| Mast cells | −0.087 | 0.094 | −0.057 | 0.269 |
| Neutrophils | −0.202 | <0.001 *** | −0.221 | <0.001 *** |
| NK CD56 bright cells | 0.134 | 0.010 * | 0.154 | 0.003 ** |
| NK CD56 dim cells | −0.118 | 0.022 * | −0.158 | 0.002 ** |
| NK cells | 0.041 | 0.425 | −0.026 | 0.617 |
| pDC | −0.309 | <0.001 *** | −0.310 | <0.001 *** |
| T cells | −0.121 | 0.019 * | −0.144 | 0.005 ** |
| T helper cells | 0.283 | <0.001 *** | 0.281 | <0.001 *** |
| Tcm | 0.161 | 0.002 ** | 0.148 | 0.004 ** |
| Tem | 0.082 | 0.112 | 0.053 | 0.311 |
| TFH | 0.134 | 0.010 * | 0.123 | 0.017 * |
| Tgd | −0.058 | 0.262 | −0.143 | 0.006 ** |
| Th1 cells | −0.042 | 0.419 | −0.048 | 0.351 |
| Th17 cells | −0.027 | 0.599 | −0.022 | 0.668 |
| Th2 cells | 0.386 | <0.001 *** | 0.369 | <0.001 *** |
| TReg | −0.227 | <0.001 *** | −0.235 | <0.001 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Huang, Y.; Mei, Y.; Zhang, Y.; Luo, Q.; Zhu, S.; Peng, L.; Gao, Z.; Liu, Y.; Li, J. High Expression of POGK Predicts Poor Prognosis in Patients with Hepatocellular Carcinoma. Curr. Oncol. 2022, 29, 8650-8667. https://doi.org/10.3390/curroncol29110682
Xu W, Huang Y, Mei Y, Zhang Y, Luo Q, Zhu S, Peng L, Gao Z, Liu Y, Li J. High Expression of POGK Predicts Poor Prognosis in Patients with Hepatocellular Carcinoma. Current Oncology. 2022; 29(11):8650-8667. https://doi.org/10.3390/curroncol29110682
Chicago/Turabian StyleXu, Wenxiong, Yanlin Huang, Yongyu Mei, Yeqiong Zhang, Qiumin Luo, Shu Zhu, Liang Peng, Zhiliang Gao, Ying Liu, and Jianguo Li. 2022. "High Expression of POGK Predicts Poor Prognosis in Patients with Hepatocellular Carcinoma" Current Oncology 29, no. 11: 8650-8667. https://doi.org/10.3390/curroncol29110682
APA StyleXu, W., Huang, Y., Mei, Y., Zhang, Y., Luo, Q., Zhu, S., Peng, L., Gao, Z., Liu, Y., & Li, J. (2022). High Expression of POGK Predicts Poor Prognosis in Patients with Hepatocellular Carcinoma. Current Oncology, 29(11), 8650-8667. https://doi.org/10.3390/curroncol29110682