
Antidiabetic, Antiglycation, and Antioxidant Activities of Ethanolic Seed Extract of Passiflora edulis and Piceatannol In Vitro

 , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Antidiabetic Activity
2.2. Inhibition of AGE in the Initial and Intermediate States of Glycation by the Extract
2.3. Effect on the Formation of Amyloid Fibrils In Vitro
2.4. Antioxidant Activity
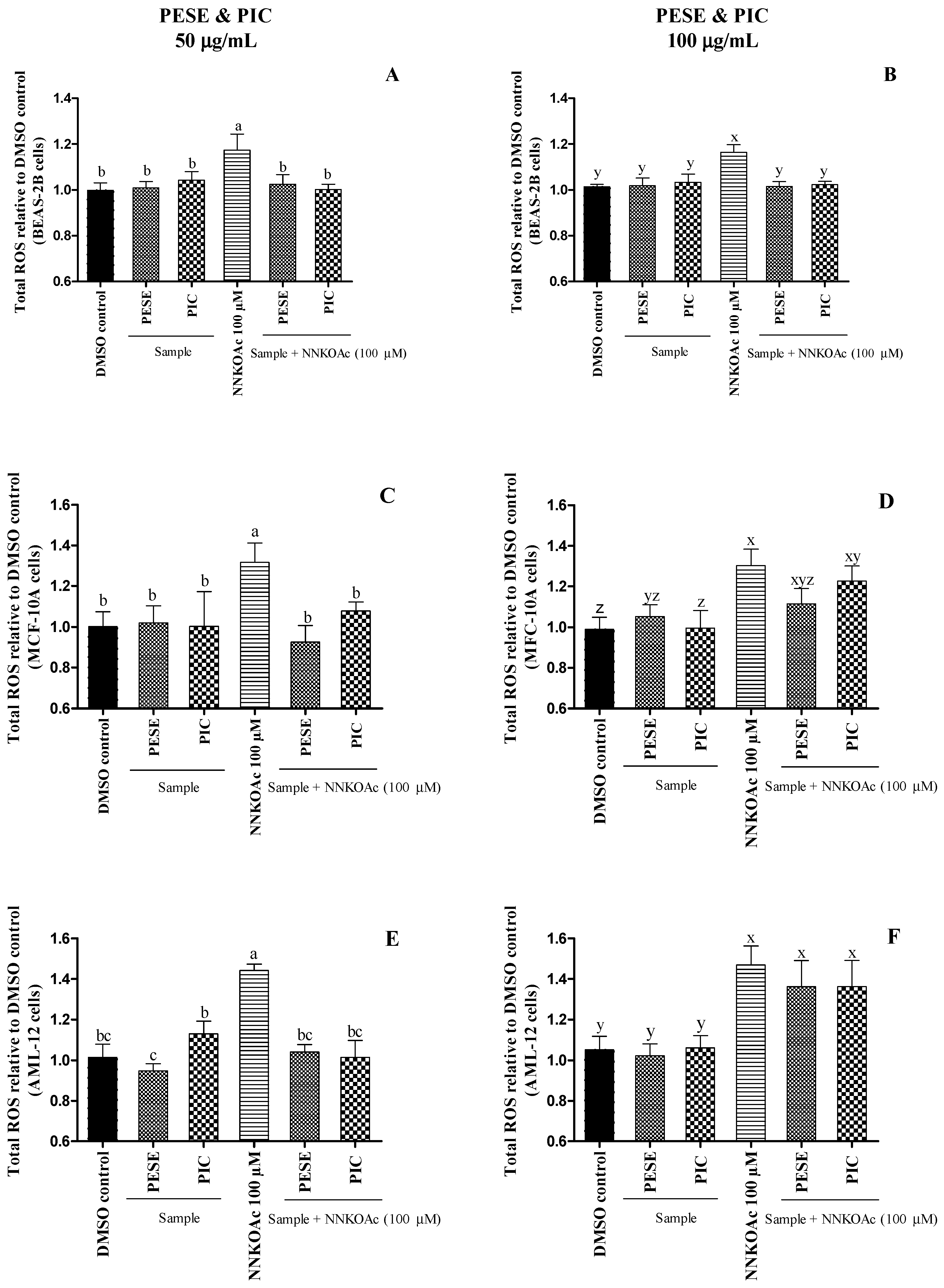
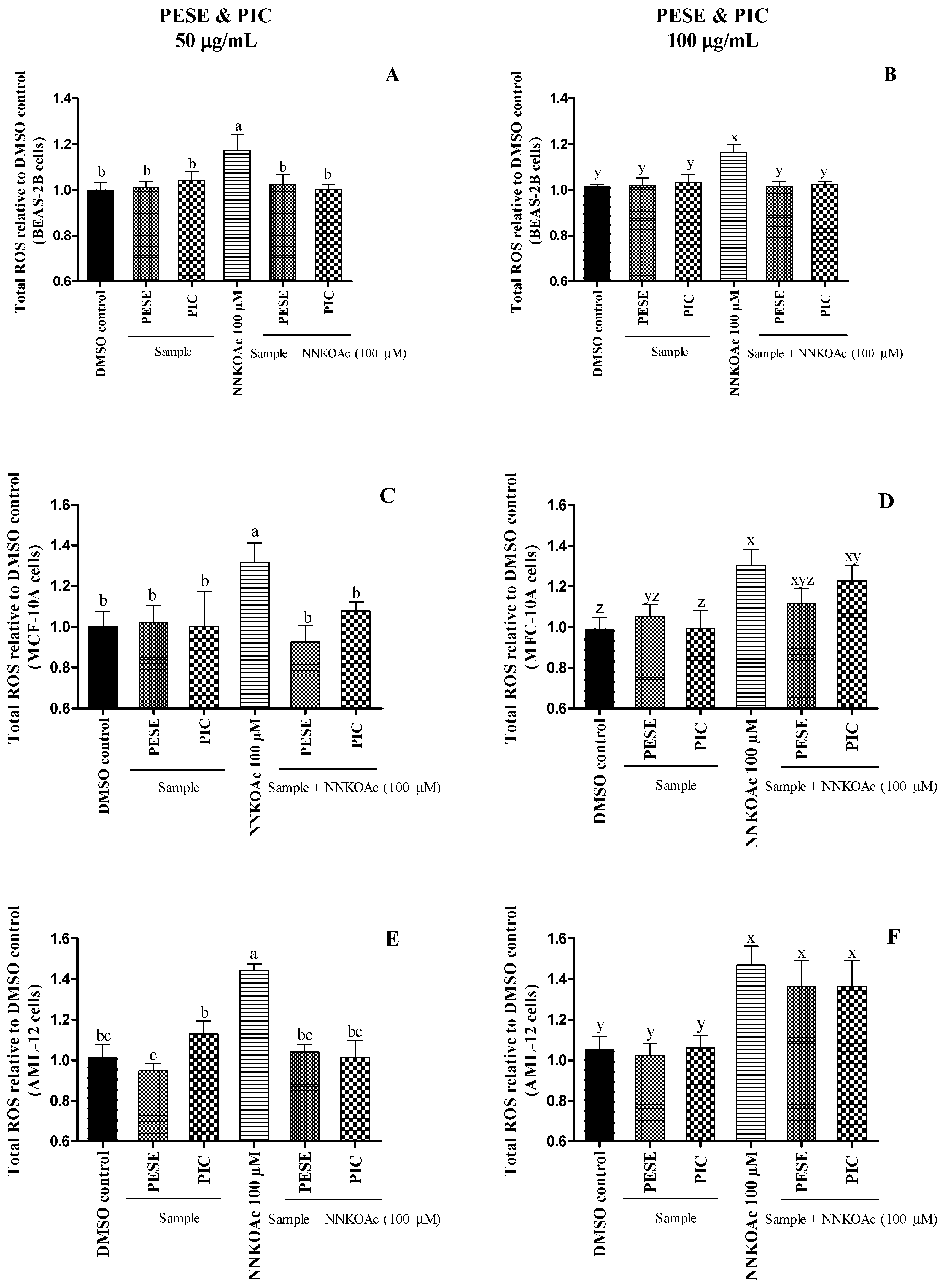
2.5. The Extract Reduces Intracellular ROS-Induced by a Carcinogen
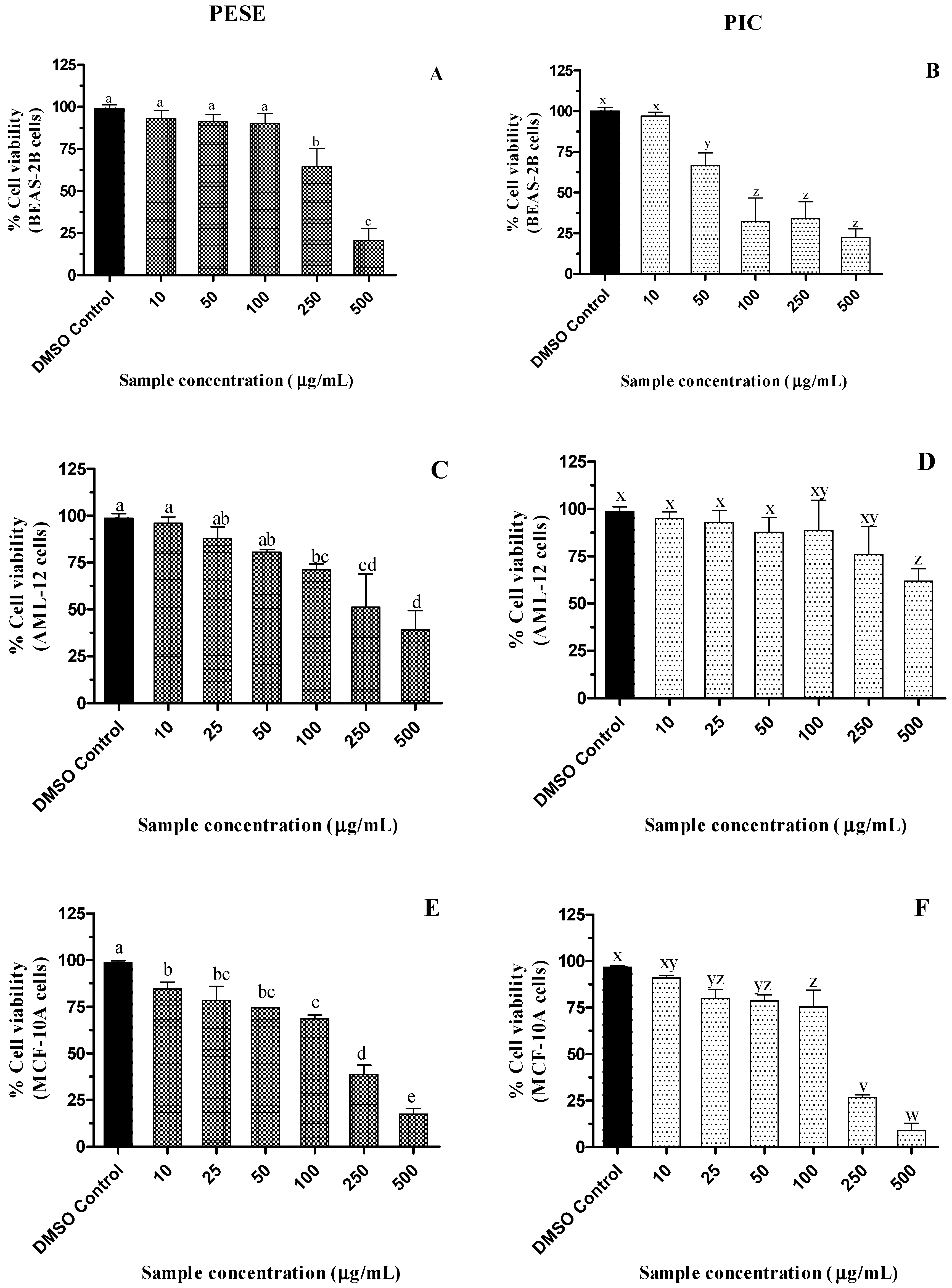
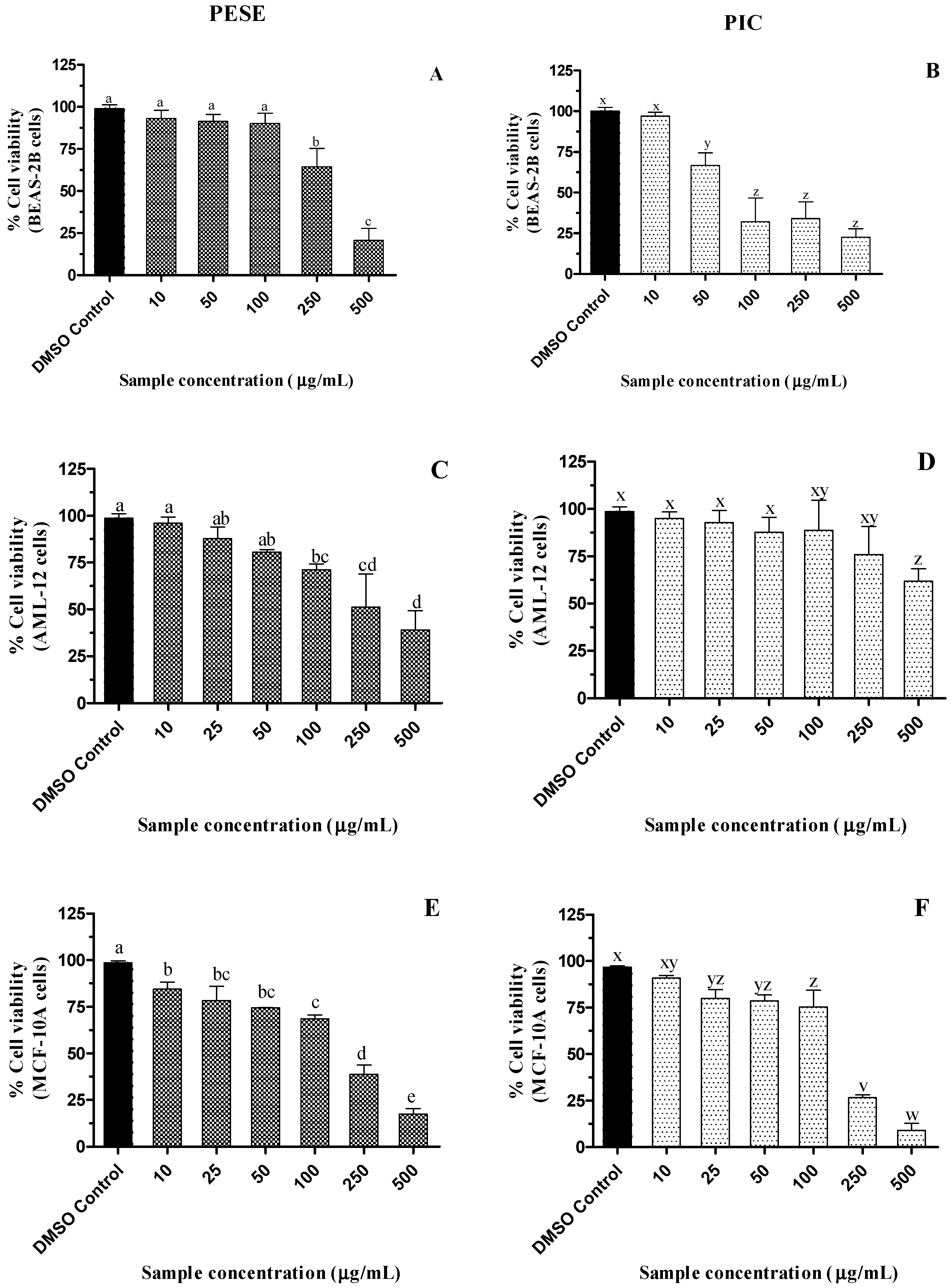
2.6. Cytotoxicity of the Seed Extract of P. edulis (PESE) and PIC
3. Material and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Antidiabetic Activity Assays In Vitro
3.3.1. Alpha-Amylase Inhibition Assay
3.3.2. Alpha-Glucosidase Inhibition Assay
3.3.3. Dipeptidyl Dipeptidase Enzyme (DPP-4) Inhibition Assay
3.4. Antiglycation Activity Assays In Vitro
3.4.1. Inhibition of Advanced Glycation End Products (AGE) Formation in the Initial Stage of Glycation
3.4.2. Inhibition of AGE Formation in the Intermediate Stage of Glycation
3.4.3. Inhibition of the Formation of Amyloid Fibrils Using ThT Assay
3.4.4. Evaluation of MGO Capture Using Derivatization with OPD
3.5. Scavenging of Free Radical and Reactive Oxygen Species (ROS) Assays
3.5.1. Total Phenolic Content (TPC)
3.5.2. Radical Scavenging Assay DPPH•
3.5.3. Hypochlorous Acid (HOCl) Scavenging Activity
3.5.4. Superoxide Anion Radical Scavenging Activity
3.5.5. Measurement of Intracellular ROS Level
3.6. Cell Cultures and Cell Viability Assay
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Da Silva Francischini, D.; Lopes, A.P.; Segatto, M.L.; Stahl, A.M.; Zuin, V.G. Development and Application of Green and Sustainable Analytical Methods for Flavonoid Extraction from Passiflora Waste. BMC Chem. 2020, 14, 56. [Google Scholar] [CrossRef] [PubMed]
- IBGE. Produção de Maracujá. Available online: https://www.ibge.gov.br/explica/producao-agropecuaria/maracuja/br (accessed on 10 December 2020).
- He, X.; Luan, F.; Yang, Y.; Wang, Z.; Zhao, Z.; Fang, J.; Wang, M.; Zuo, M.; Li, Y. Passiflora Edulis: An Insight into Current Researches on Phytochemistry and Pharmacology. Front. Pharmacol. 2020, 11, 617. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Taiwe, G.S. Passiflora edulis. In Medicinal Spices and Vegetables from Africa: Therapeutic Potential Against Metabolic, Inflamatory, Infectious and Systemic Diseases; Elsevier Science & Technology: Amsterdam, The Netherlands, 2017; pp. 513–523. ISBN 978-0-12-809286-6. [Google Scholar]
- Krambeck, K.; Oliveira, A.; Santos, D.; Pintado, M.M.; Silva, J.B.; Lobo, J.M.S.; Amaral, M.H. Identification and Quantification of Stilbenes (Piceatannol and Resveratrol) in Passiflora edulis by-Products. Pharmaceuticals 2020, 13, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Jung, J.I.; Cho, H.J.; Her, S.; Kwon, S.H.; Yu, R.; Kang, Y.H.; Lee, K.W.; Park, J.H.Y. Inhibition of Tumor Progression by Oral Piceatannol in Mouse 4T1 Mammary Cancer Is Associated with Decreased Angiogenesis and Macrophage Infiltration. J. Nutr. Biochem. 2015, 26, 1368–1378. [Google Scholar] [CrossRef]
- Kershaw, J.; Kim, K.H. The Therapeutic Potential of Piceatannol, a Natural Stilbene, in Metabolic Diseases: A Review. J. Med. Food 2017, 20, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Nagapan, T.S.; Ghazali, A.R.; Basri, D.F.; Lim, W.N. Photoprotective Effect of Stilbenes and Its Derivatives against Ultraviolet Radiation-Induced Skin Disorders. Biomed. Pharmacol. J. 2018, 11, 1199–1208. [Google Scholar] [CrossRef]
- Maruki-Uchida, H.; Morita, M.; Yonei, Y.; Sai, M. Effect of Passion Fruit Seed Extract Rich in Piceatannol on the Skin of Women: A Randomized, Placebo-Controlled, Double-Blind Trial. J. Nutr. Sci. Vitaminol. 2018, 64, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Surh, Y.-J.; Na, H.-K. Anti-Inflammatory Nutraceuticals and Chronic Diseases. In Advances in Experimental Medicine and Biology; Gupta, S.C., Prasad, S., Aggarwal, B.B., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; Volume 928, pp. 185–212. ISBN 9783319413327. [Google Scholar]
- Gregg, E.; Buckley, J.; Ali, M.; Davies, J.; Flood, D.; Griffiths, B.; Lim, L.-L.; Manne-Goehler, J.; Pearson-Stuttard, J.; Shaw, J. Improving Health Outcomes of People with Diabetes Mellitus: Target Setting to Reduce the Global Burden of Diabetes Mellitus by 2030; World Health Organization: Geneva, Switzerland, 2021; Volume 8. [Google Scholar]
- International Diabetes Federation. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; Volume 102, ISBN 9782930229980. [Google Scholar]
- Tupas, G.D.; Otero, M.C.B.; Ebhohimen, I.E.; Egbuna, C.; Aslam, M. Antidiabetic Lead Compounds and Targets for Drug Development. In Phytochemicals as Lead Compounds for New Drug Discovery; Egbuna, C., Kumar, S., Ifemeje, J., Ezzat, S., Kaliyaperumal, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 127–141. ISBN 9780128178904. [Google Scholar]
- Marín-Peñalver, J.J.; Martín-Timón, I.; Sevillano-Collantes, C.; del Cañizo-Gómez, F.J. Update on the Treatment of Type 2 Diabetes Mellitus. World J. Diabetes 2016, 7, 354–395. [Google Scholar] [CrossRef]
- Sekhon-Loodu, S.; Rupasinghe, H.P.V. Evaluation of Antioxidant, Antidiabetic and Antiobesity Potential of Selected Traditional Medicinal Plants. Front. Nutr. 2019, 6, 53. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Azimullah, S.; Beiram, R. Diabetes as a Risk Factor for Alzheimer’s Disease in the Middle East and Its Shared Pathological Mediators. Saudi J. Biol. Sci. 2020, 27, 736–750. [Google Scholar] [CrossRef]
- Kong, Y.; Wang, F.; Wang, J.; Liu, C.; Zhou, Y.; Xu, Z.; Zhang, C.; Sun, B.; Guan, Y. Pathological Mechanisms Linking Diabetes Mellitus and Alzheimer’s Disease: The Receptor for Advanced Glycation End Products (RAGE). Front. Aging Neurosci. 2020, 12, 217. [Google Scholar] [CrossRef] [PubMed]
- Torres, N.M.P.O.; Xavier, J.A.; Goulart, M.O.F.; Alves, R.B.; Freitas, R.P. The Chemistry of Advanced Glycation End-Products. Rev. Virtual Quim. 2018, 10, 375–392. [Google Scholar] [CrossRef]
- Zaman, M.; Khan, A.N.; Wahiduzzaman; Zakariya, S.M.; Khan, R.H. Protein Misfolding, Aggregation and Mechanism of Amyloid Cytotoxicity: An Overview and Therapeutic Strategies to Inhibit Aggregation. Int. J. Biol. Macromol. 2019, 134, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Miao, M. Dietary Polyphenols Modulate Starch Digestion and Glycaemic Level: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Uchida-Maruki, H.; Inagaki, H.; Ito, R.; Kurita, I.; Sai, M.; Ito, T. Piceatannol Lowers the Blood Glucose Level in Diabetic Mice. Biol. Pharm. Bull. 2015, 38, 629–633. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Gao, P.; Tao, L.; Wen, J.; Wang, L.; Yi, Y.; Chen, Y.; Wang, J.; Xu, X.; Zhang, J.; et al. Piceatannol Attenuates Streptozotocin-Induced Type 1 Diabetes in Mice. Biocell 2020, 44, 353–361. [Google Scholar] [CrossRef]
- Cao, Q.; Teng, J.; Wei, B.; Huang, L.; Xia, N. Phenolic Compounds, Bioactivity, and Bioaccessibility of Ethanol Extracts from Passion Fruit Peel Based on Simulated Gastrointestinal Digestion. Food Chem. 2021, 356, 129682. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Lucci, P.; Núñez, O.; Tundis, R.; Balzano, M.; Frega, N.G.; Conte, L.; Moret, S.; Filatova, D.; Moyano, E.; et al. Native Colombian Fruits and Their By-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential. Foods 2019, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.H.; Ning, D.S.; Fu, Y.X.; Li, D.P.; Zou, Z.Q.; Xie, Y.C.; Yu, L.L.; Li, L.C. Preparative Isolation of Piceatannol Derivatives from Passion Fruit (Passiflora edulis) Seeds by High-Speed Countercurrent Chromatography Combined with High-Performance Liquid Chromatography and Screening for α-Glucosidase Inhibitory Activities. J. Agric. Food Chem. 2020, 68, 1555–1562. [Google Scholar] [CrossRef]
- Justino, A.B.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; da Silva, N.M.; Espindola, F.S. Annona muricata Linn. Leaf as a Source of Antioxidant Compounds with In Vitro Antidiabetic and Inhibitory Potential against α-Amylase, α-Glucosidase, Lipase, Non-Enzymatic Glycation and Lipid Peroxidation. Biomed. Pharmacother. 2018, 100, 83–92. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Z.; Wang, X.; Wang, S.; Cao, J.; Liu, Y. Exploring the Inhibitory Mechanism of Piceatannol on α-Glucosidase Relevant to Diabetes Mellitus. RSC Adv. 2020, 10, 4529–4537. [Google Scholar] [CrossRef] [PubMed]
- Talakatta, G.; Sarikhani, M.; Muhamed, J.; Dhanya, K.; Somashekar, B.S.; Mahesh, P.A.; Sundaresan, N.; Ravindra, P.V. Diabetes Induces Fibrotic Changes in the Lung through the Activation of TGF-β Signaling Pathways. Sci. Rep. 2018, 8, 11920. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Katakami, N.; Matsuhisa, M.; Matsuoka, T.A. Role of Reactive Oxygen Species in the Progression of Type 2 Diabetes and Atherosclerosis. Mediat. Inflamm. 2010, 2010, 453892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Dicarbonyl Stress, Protein Glycation and the Unfolded Protein Response. Glycoconj. J. 2021, 38, 331–340. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Y.; Liu, H. Corn Silk Extract Inhibit the Formation of Nε-Carboxymethyllysine by Scavenging Glyoxal/Methyl Glyoxal in a Casein Glucose-Fatty Acid Model System. Food Chem. 2020, 309, 125708. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, M., II; Syed, A.S.; Jung, K.; Kim, C.Y. Rapid Identification of Methylglyoxal Trapping Constituents from Onion Peels by Pre-Column Incubation Method. Nat. Prod. Sci. 2017, 23, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yagiz, Y.; Buran, T.J.; Nunes, C.D.N.; Gu, L. Phytochemicals from Berries and Grapes Inhibited the Formation of Advanced Glycation End-Products by Scavenging Reactive Carbonyls. Food Res. Int. 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Taghavi, F.; Habibi-Rezaei, M.; Amani, M.; Saboury, A.A.; Moosavi-Movahedi, A.A. The Status of Glycation in Protein Aggregation. Int. J. Biol. Macromol. 2017, 100, 67–74. [Google Scholar] [CrossRef]
- Xue, C.; Lin, T.Y.; Chang, D.; Guo, Z. Thioflavin T as an Amyloid Dye: Fibril Quantification, Optimal Concentration and Effect on Aggregation. R. Soc. Open Sci. 2017, 4, 160696. [Google Scholar] [CrossRef] [Green Version]
- Xavier, J.A.; Santos, J.C.; Nova, M.A.V.; Gonçalves, C.M.; Borbely, K.S.C.; Pires, K.S.N.; dos Santos, F.A.R.; Valentim, I.B.; Barbosa, J.H.P.; da Silva, F.C.; et al. Anti-Zika Virus Efects, Placenta Protection and Chemical Composition of Passiflora edulis Seeds Ethanolic Extract. J. Braz. Chem. Soc. 2022, 33, 701–714. [Google Scholar] [CrossRef]
- Rotta, E.M.; Giroux, H.J.; Lamothe, S.; Bélanger, D.; Sabik, H.; Visentainer, J.V.; Britten, M. Use of Passion Fruit Seed Extract (Passiflora edulis Sims) to Prevent Lipid Oxidation in Dairy Beverages during Storage and Simulated Digestion. LWT 2020, 123, 109088. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Aryaeian, N.; Sedehi, S.K.; Arablou, T. Polyphenols and Their Effects on Diabetes Management: A Review. Med. J. Islam. Repub. Iran 2017, 31, 886–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlin, J.P.J.; Dellaire, G.; Murphy, K.; Rupasinghe, H.P.V. Vitamin-Containing Antioxidant Formulation Reduces Carcinogen-Induced Dna Damage through Atr/Chk1 Signaling in Bronchial Epithelial Cells in Vitro. Biomedicines 2021, 9, 1665. [Google Scholar] [CrossRef]
- Amararathna, M.; Hoskin, D.W.; Rupasinghe, H.P.V. Anthocyanin-Rich Haskap (Lonicera caerulea L.) Berry Extracts Reduce Nitrosamine-Induced DNA Damage in Human Normal Lung Epithelial Cells in Vitro. Food Chem. Toxicol. 2020, 141, 111404. [Google Scholar] [CrossRef]
- Clague, J.; Shao, L.; Lin, J.; Chang, S.; Zhu, Y.; Wang, W.; Wood, C.G.; Wu, X. Sensitivity to NNKOAc Is Associated with Renal Cancer Risk. Carcinogenesis 2009, 30, 706–710. [Google Scholar] [CrossRef]
- Sjögren, G.; Sletten, G.; Dahl, J.E. Cytotoxicity of Dental Alloys, Metals, and Ceramics Assessed by Millipore Filter, Agar Overlay, and MTT Tests. J. Prosthet. Dent. 2000, 84, 229–236. [Google Scholar] [CrossRef]
- Yepes, A.; Ochoa-Bautista, D.; Murillo-Arango, W.; Quintero-Saumeth, J.; Bravo, K.; Osorio, E. Purple Passion Fruit Seeds (Passiflora Edulis f. Edulis Sims) as a Promising Source of Skin Anti-Aging Agents: Enzymatic, Antioxidant and Multi-Level Computational Studies. Arab. J. Chem. 2021, 14, 102905. [Google Scholar] [CrossRef]
- Kido, L.A.; Hahm, E.-R.; Kim, S.-H.; Baseggio, A.M.; Cagnon, V.H.A.; Singh, S.V.; Maróstica, M.R. Prevention of Prostate Cancer in Transgenic Adenocarcinoma of the Mouse Prostate Mice by Yellow Passion Fruit Extract and Antiproliferative Effects of Its Bioactive Compound Piceatannol. J. Cancer Prev. 2020, 25, 87–99. [Google Scholar] [CrossRef]
- de Silva, A.B.K.H.; Rupasinghe, H.P.V. Polyphenols Composition and Anti-Diabetic Properties in Vitro of Haskap (Lonicera caerulea L.) Berries in Relation to Cultivar and Harvesting Date. J. Food Compos. Anal. 2020, 88, 103402. [Google Scholar] [CrossRef]
- de Melo, I.S.V.; dos Santos, A.F.; de Lemos, T.L.G.; Goulart, M.O.F.; Santana, A.E.G. Oncocalyxone a Functions as an Anti-Glycation Agent In Vitro. PLoS ONE 2015, 10, e0131222. [Google Scholar] [CrossRef] [Green Version]
- Moosavi-Movahedi, A.A.; Ghamari, F.; Ghaffari, S.M.; Salami, M.; Farivar, F.; Moosavi-Movahedi, F.; Johari, A.; Aminin, A.L.N. Natural Peptide Anti-Glycation Effect in the Presence of Aloe vera Phenolic Components on Human Serum Albumin. RSC Adv. 2015, 5, 248–254. [Google Scholar] [CrossRef]
- Cicco, N.; Lanorte, M.T.; Paraggio, M.; Viggiano, M.; Lattanzio, V. A Reproducible, Rapid and Inexpensive Folin-Ciocalteu Micro-Method in Determining Phenolics of Plant Methanol Extracts. Microchem. J. 2009, 91, 107–110. [Google Scholar] [CrossRef]
- de Almeida Xavier, J.; Valentim, I.B.; Camatari, F.O.S.; de Almeida, A.M.M.; Goulart, H.F.; de Souza Ferro, J.N.; de Oliveira Barreto, E.; Cavalcanti, B.C.; Bottoli, C.B.G.; Goulart, M.O.F. Polyphenol Profile by Uhplc-Ms/Ms, Anti-Glycation, Antioxidant and Cytotoxic Activities of Several Samples of Propolis from the Northeastern Semi-Arid Region of Brazil. Pharm. Biol. 2017, 55, 1884–1893. [Google Scholar] [CrossRef]
- Lucas, M.; Freitas, M.; Xavier, J.A.; Moura, F.A.; Goulart, M.O.F.; Ribeiro, D.; Fernandes, E. The Scavenging Effect of Curcumin, Piperine and Their Combination against Physiological Relevant Reactive Pro-Oxidant Species Using In Vitro Non-Cellular and Cellular Models. Chem. Pap. 2021, 75, 5269–5277. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Sample | Alpha-Amylase IC50 (μg/mL) | Alpha-Glucosidase IC50 (μg/mL) | DPP-4 IC50 (μg/mL) |
|---|---|---|---|
| PESE | 32.1 ± 2.7 a | 76.2 ± 1.9 A | 71.1 ± 2.6 x |
| PIC | 85.4 ± 0.7 b | 20.4 ± 7.6 B | 1137.5 ± 120.2 y |
| ACB | 0.4 ± 0.1 c | 251.6 ± 4.5 C | - |
| STG | - | - | 0.005 ± 0.001 z |
| Fructose + Glucose | MGO | |||
|---|---|---|---|---|
| Sample | Antiglycation | Inhibition of Amyloid Fibrils (%) | Antiglycation | Inhibition of Amyloid Fibrils (%) |
| IC50 (μg/mL) | IC50 (μg/mL) | |||
| PESE | 367 ± 1.9 a | 87.4 ± 2.7 b | 360 ± 9.1 a | 71.9 ± 4.5 b |
| PIC | 51.5 ± 1.4 b | 100 ± 5.3 a | 67.4 ± 4.6 b | 100 ± 3.8 a |
| AMG | 25.5 ± 5.0 c | 35.0 ± 5.9 c | 50.4 ± 1.8 c | 30.3 ± 4.4 c |
| Sample | TPC (mg GAE/g DW) | DPPH• IC50 μg/mL | HOCl IC50 μg/mL | O2•− IC50 μg/mL |
|---|---|---|---|---|
| PESE | 227 ± 3.9 | 20.4 ± 2.1 a | 1.7 ± 0.3 a | 38.2 ± 0.5 a |
| PIC | - | 6.3 ± 1.3 b (25.8) | 1.2 ± 0.5 b (7.8) | 7.3 ± 0.02 c (30.1) |
| QCT | - | 4.8 ± 1.0 b (15.9) | 1.9 ± 0.3 a (4.0) | 8.8 ± 0.3 b (29.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, F.A.R.; Xavier, J.A.; da Silva, F.C.; Merlin, J.P.J.; Goulart, M.O.F.; Rupasinghe, H.P.V. Antidiabetic, Antiglycation, and Antioxidant Activities of Ethanolic Seed Extract of Passiflora edulis and Piceatannol In Vitro. Molecules 2022, 27, 4064. https://doi.org/10.3390/molecules27134064
dos Santos FAR, Xavier JA, da Silva FC, Merlin JPJ, Goulart MOF, Rupasinghe HPV. Antidiabetic, Antiglycation, and Antioxidant Activities of Ethanolic Seed Extract of Passiflora edulis and Piceatannol In Vitro. Molecules. 2022; 27(13):4064. https://doi.org/10.3390/molecules27134064
Chicago/Turabian Styledos Santos, Flávia A. R., Jadriane A. Xavier, Felipe C. da Silva, J. P. Jose Merlin, Marília O. F. Goulart, and H. P. Vasantha Rupasinghe. 2022. "Antidiabetic, Antiglycation, and Antioxidant Activities of Ethanolic Seed Extract of Passiflora edulis and Piceatannol In Vitro" Molecules 27, no. 13: 4064. https://doi.org/10.3390/molecules27134064
APA Styledos Santos, F. A. R., Xavier, J. A., da Silva, F. C., Merlin, J. P. J., Goulart, M. O. F., & Rupasinghe, H. P. V. (2022). Antidiabetic, Antiglycation, and Antioxidant Activities of Ethanolic Seed Extract of Passiflora edulis and Piceatannol In Vitro. Molecules, 27(13), 4064. https://doi.org/10.3390/molecules27134064