

Investigation of the In Vitro Immunomodulatory Effects of Extracts from Green-Lipped Mussels (Perna canaliculus)

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction Methods and Characterization of Green-Lipped Mussel Oil
2.2. Green-Lipped Mussel Oil Extract Treatment
2.3. Maintenance of Suspension Cells in Culture
2.4. Monocytes Differentiate into Macrophage Cells
2.5. MTT Cell Viability to Differentiated Macrophage Cell Lines
2.6. Monocytes and PMA-Differentiated Cells Stimulation
2.7. Jurkat Cell Line Stimulation
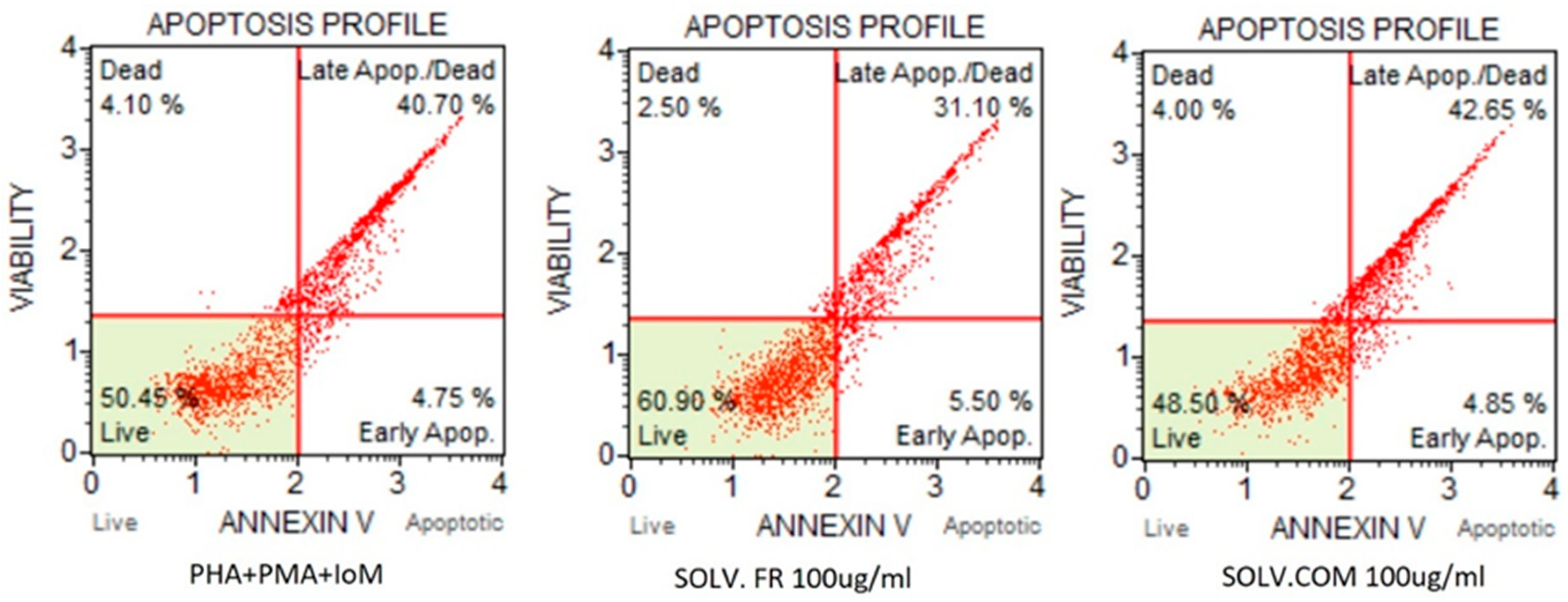
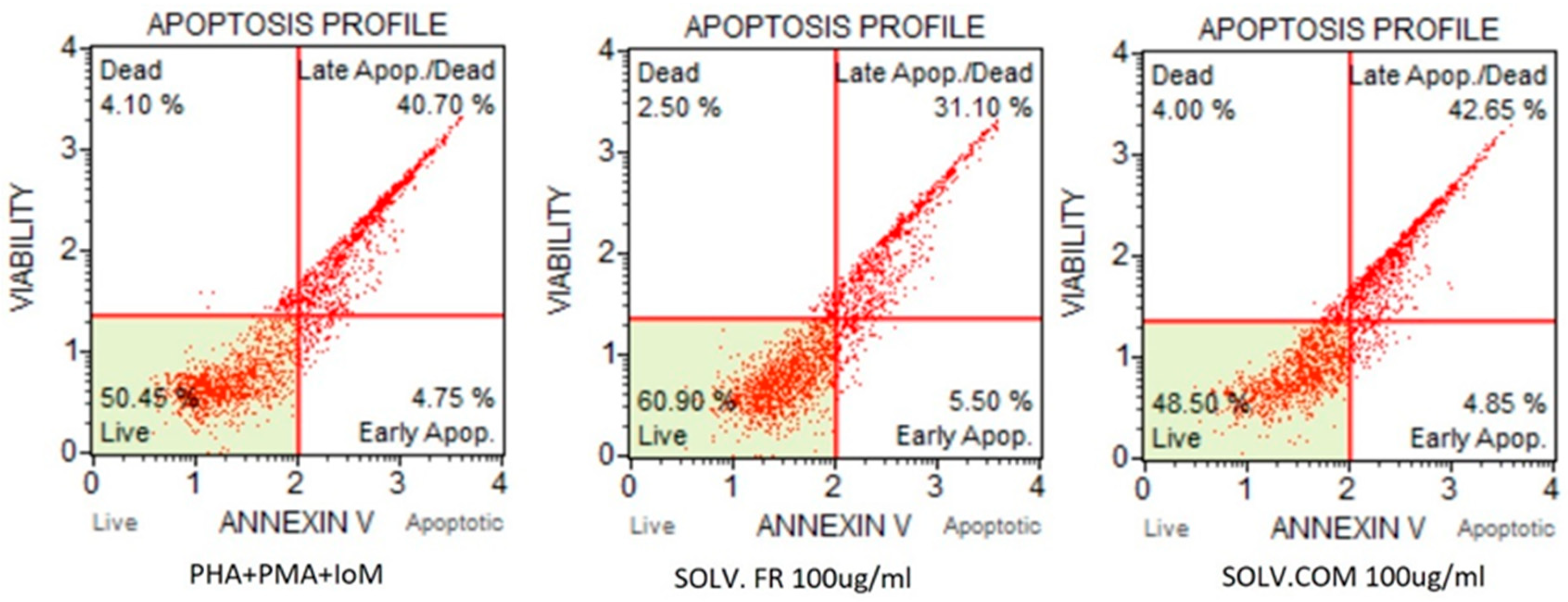
2.8. Apoptosis Assay
2.9. Enzyme-Linked Immunosorbent Assay (ELISA) of Proinflammatory Cytokines
2.10. Statistical Analysis
3. Results
3.1. Analysing the Lipid Content of Green-Lipped Mussel
Effects of Mussel Extracts on Cell Viability
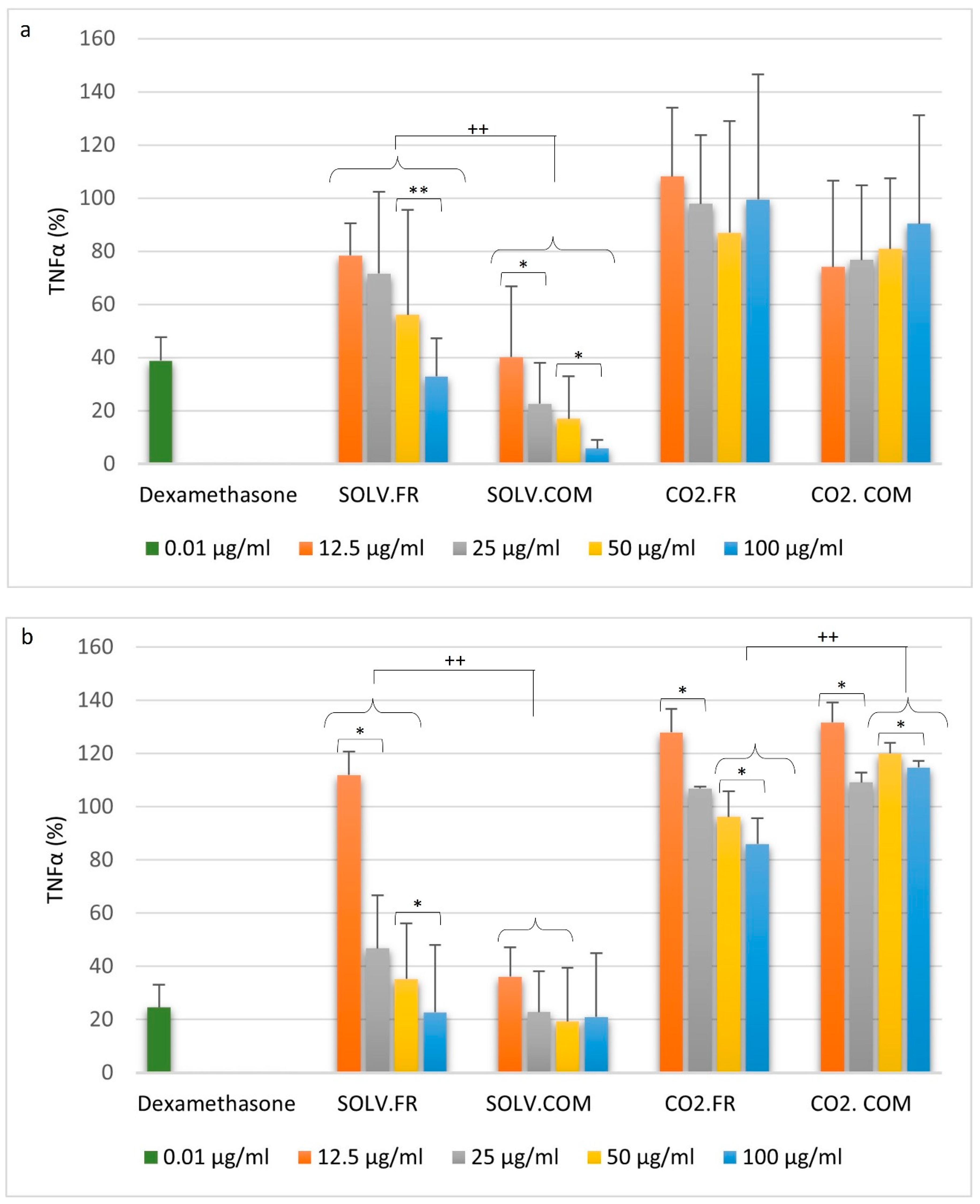
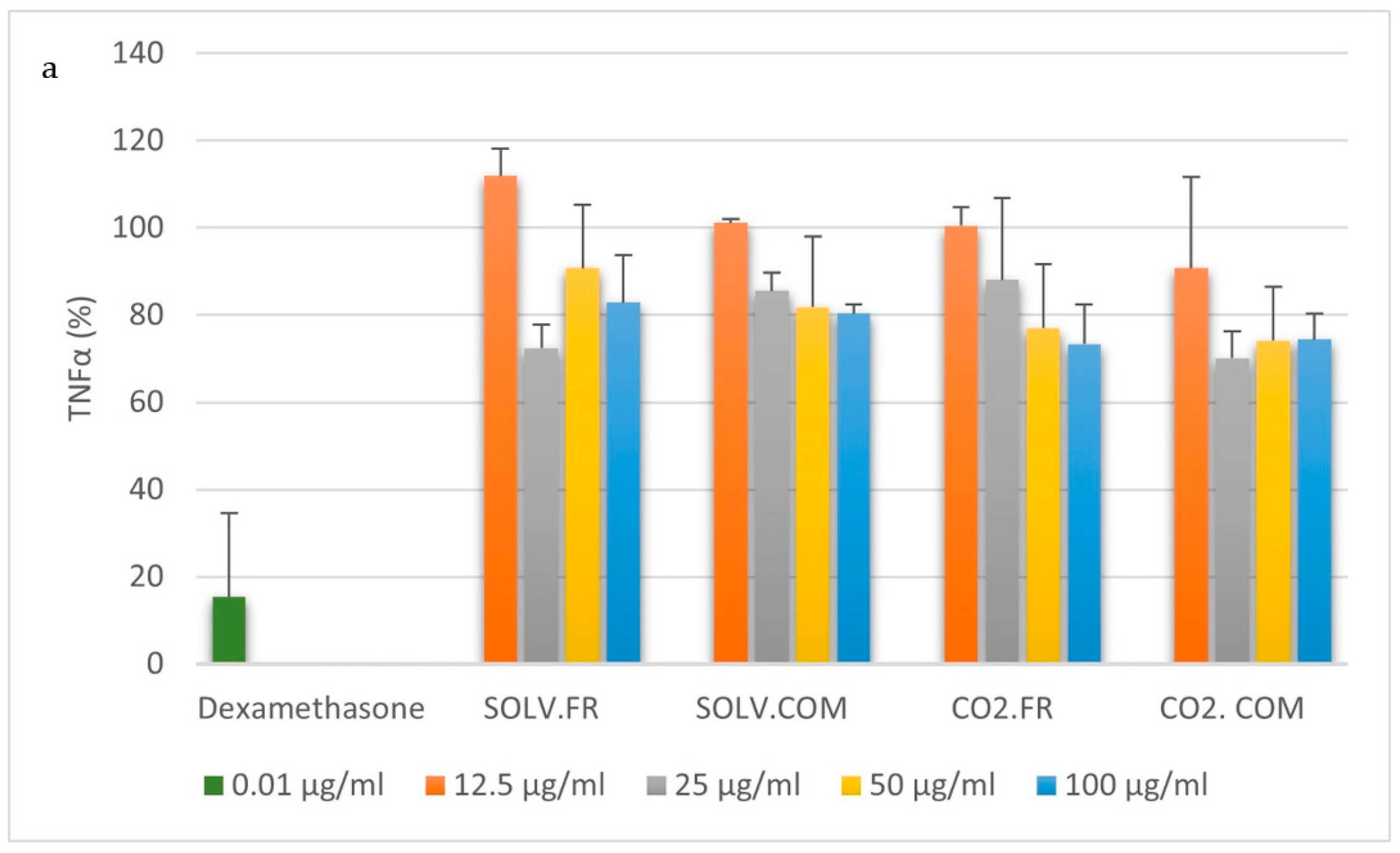
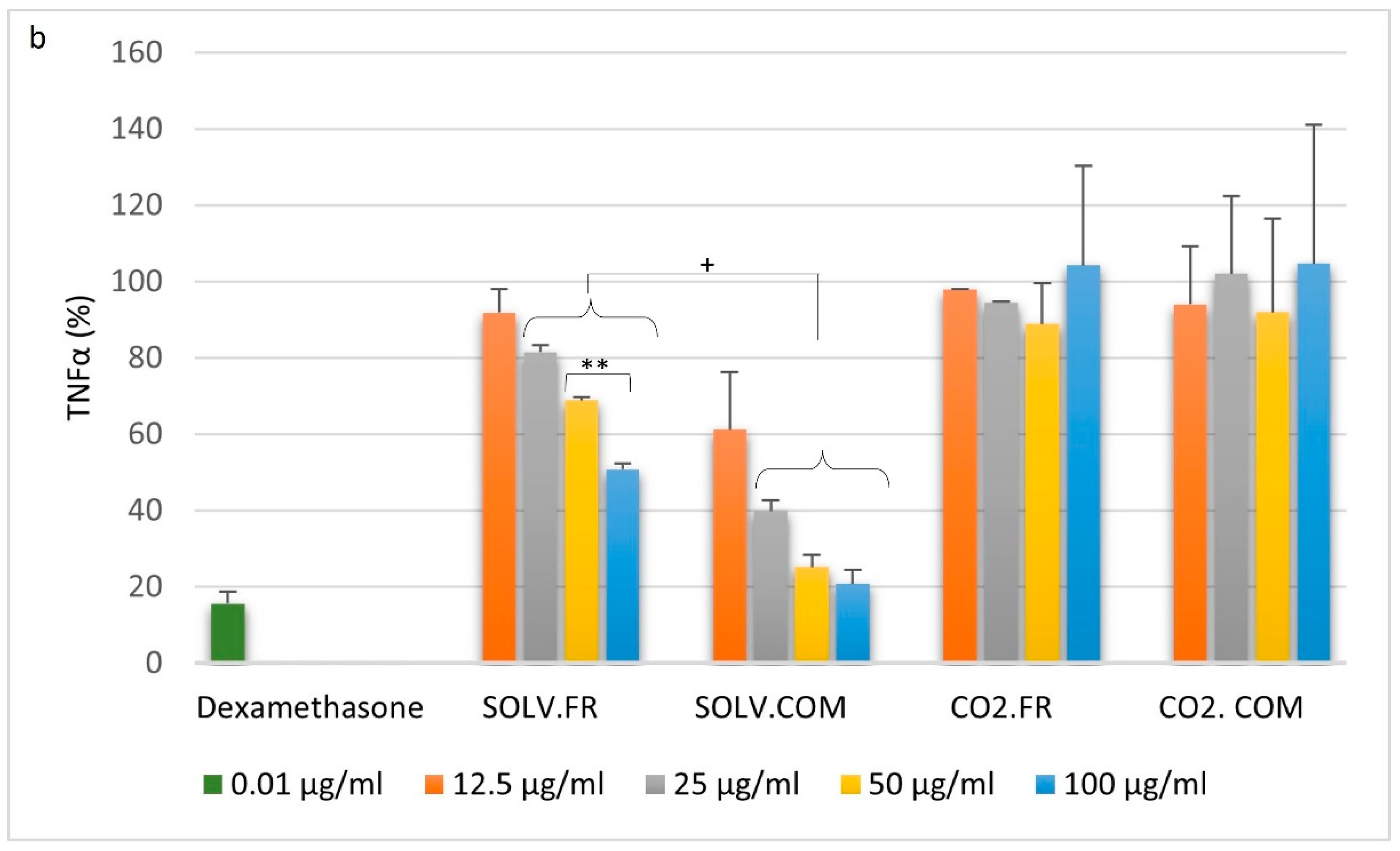
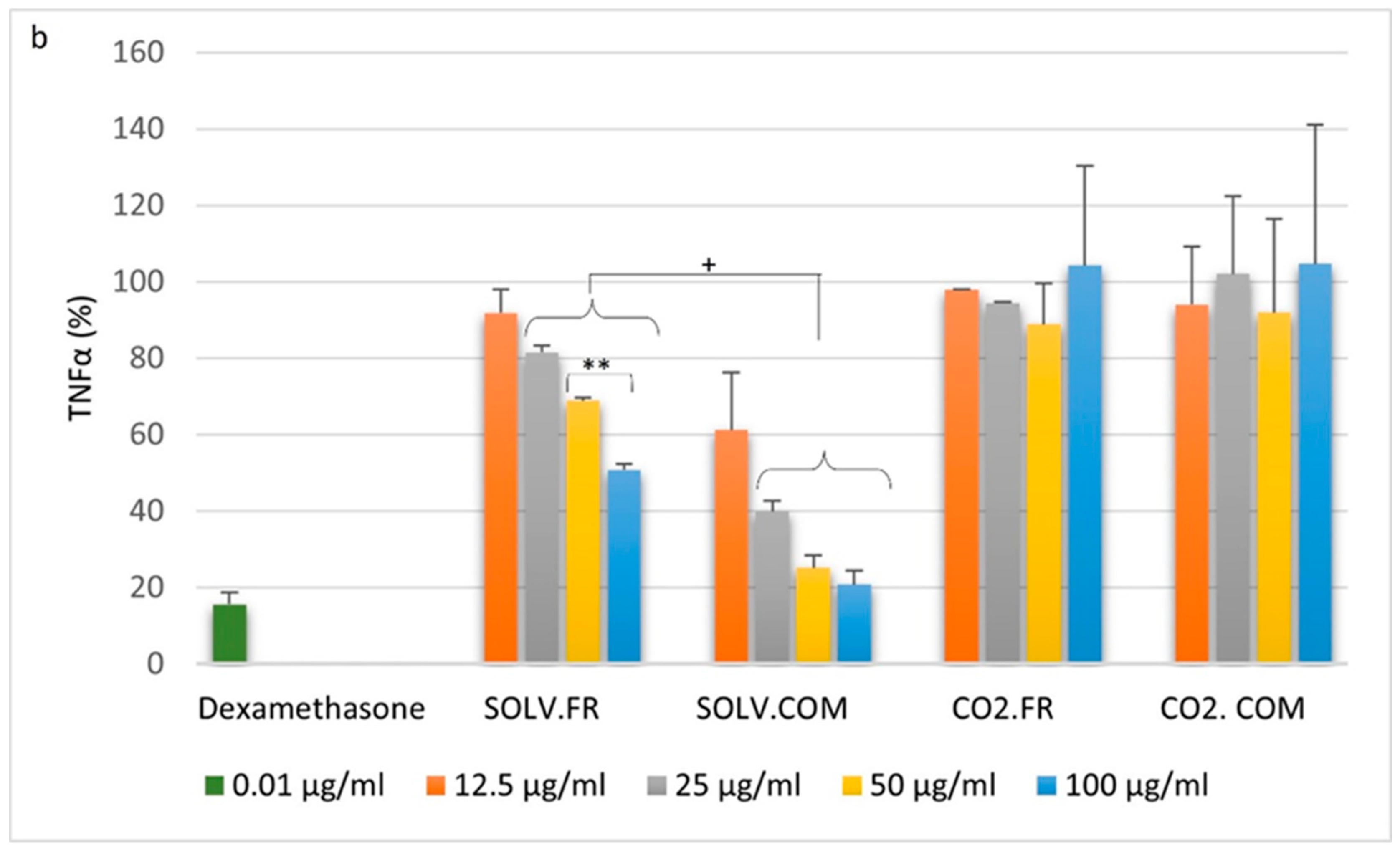
3.2. Immune Cell Response to Mussel Oil
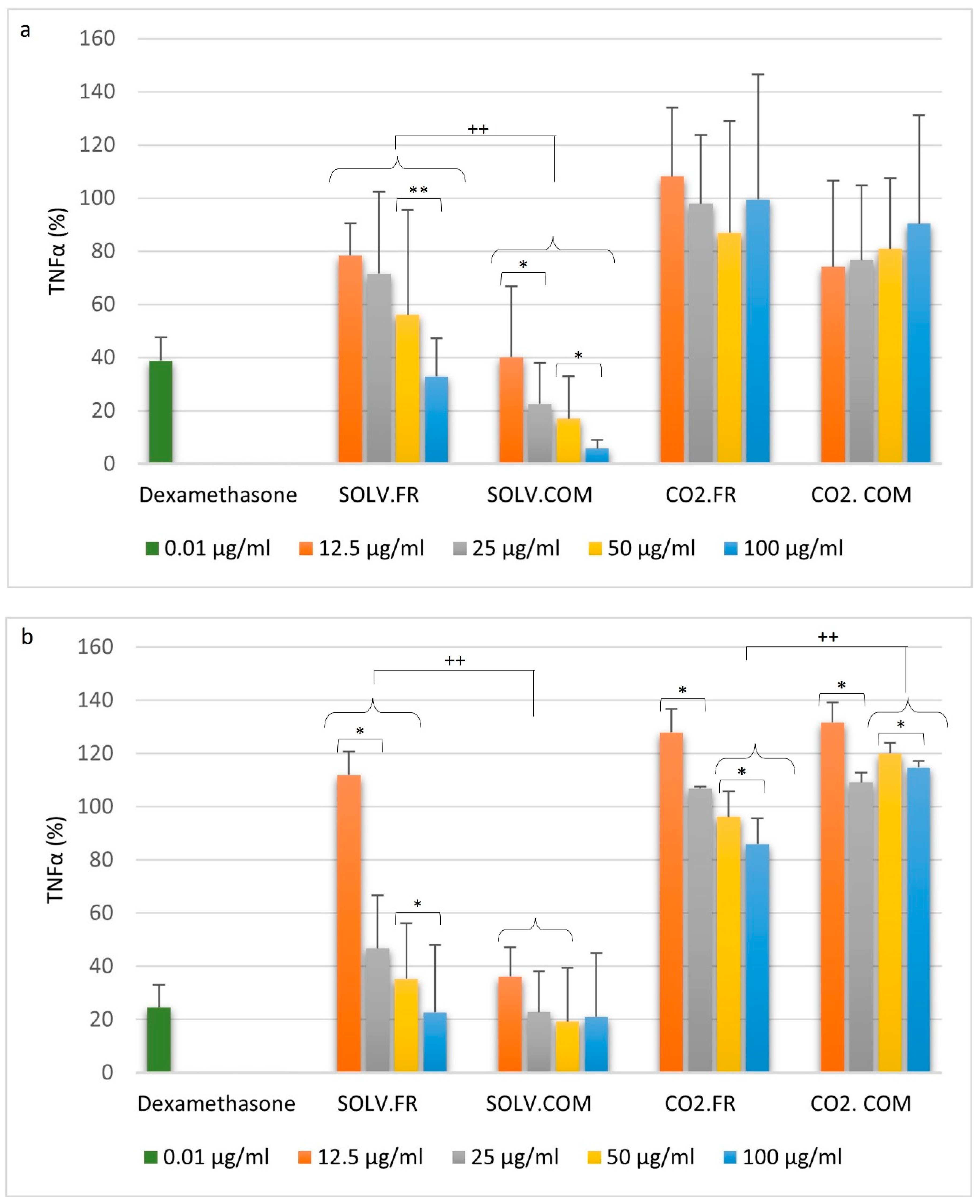
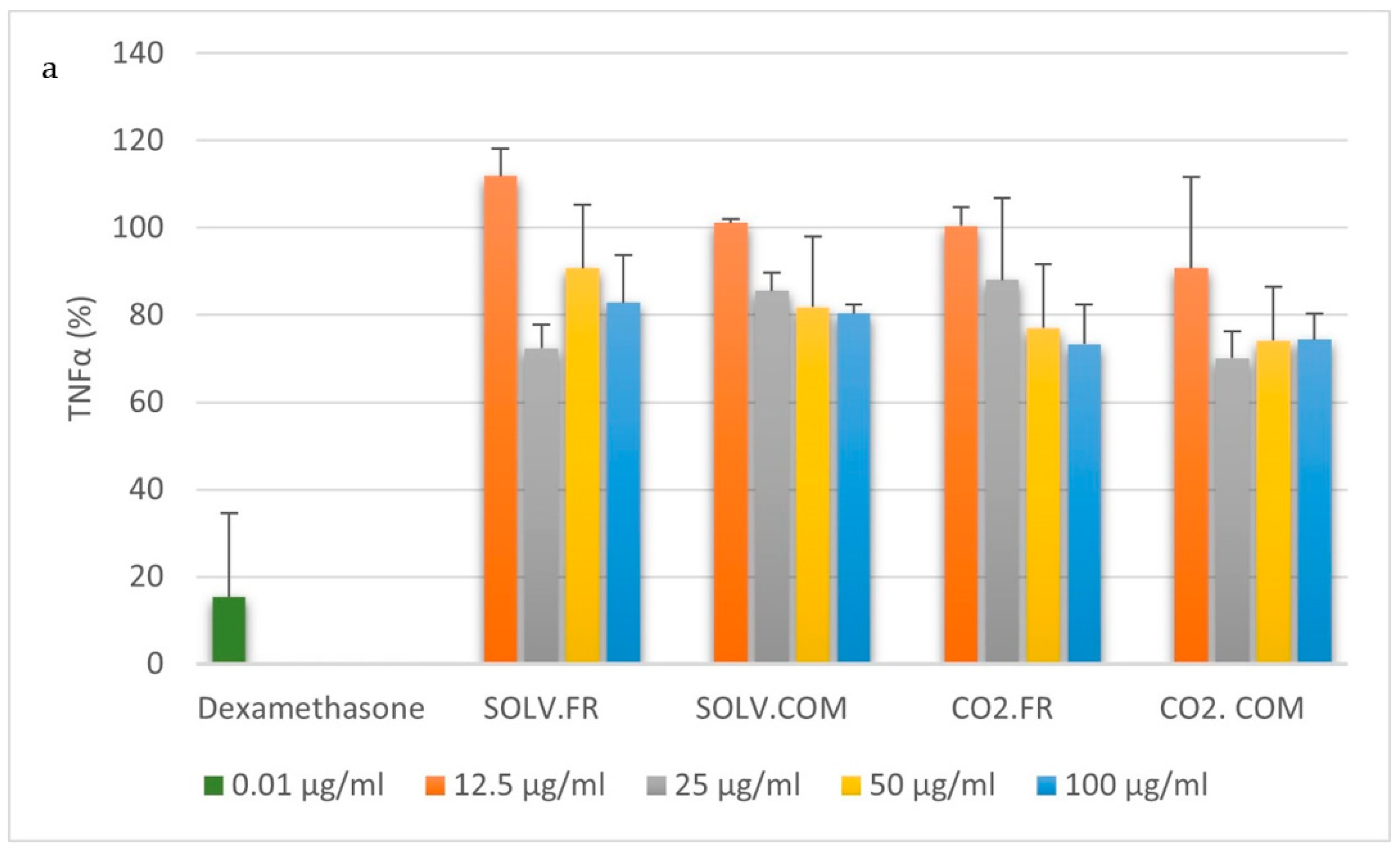
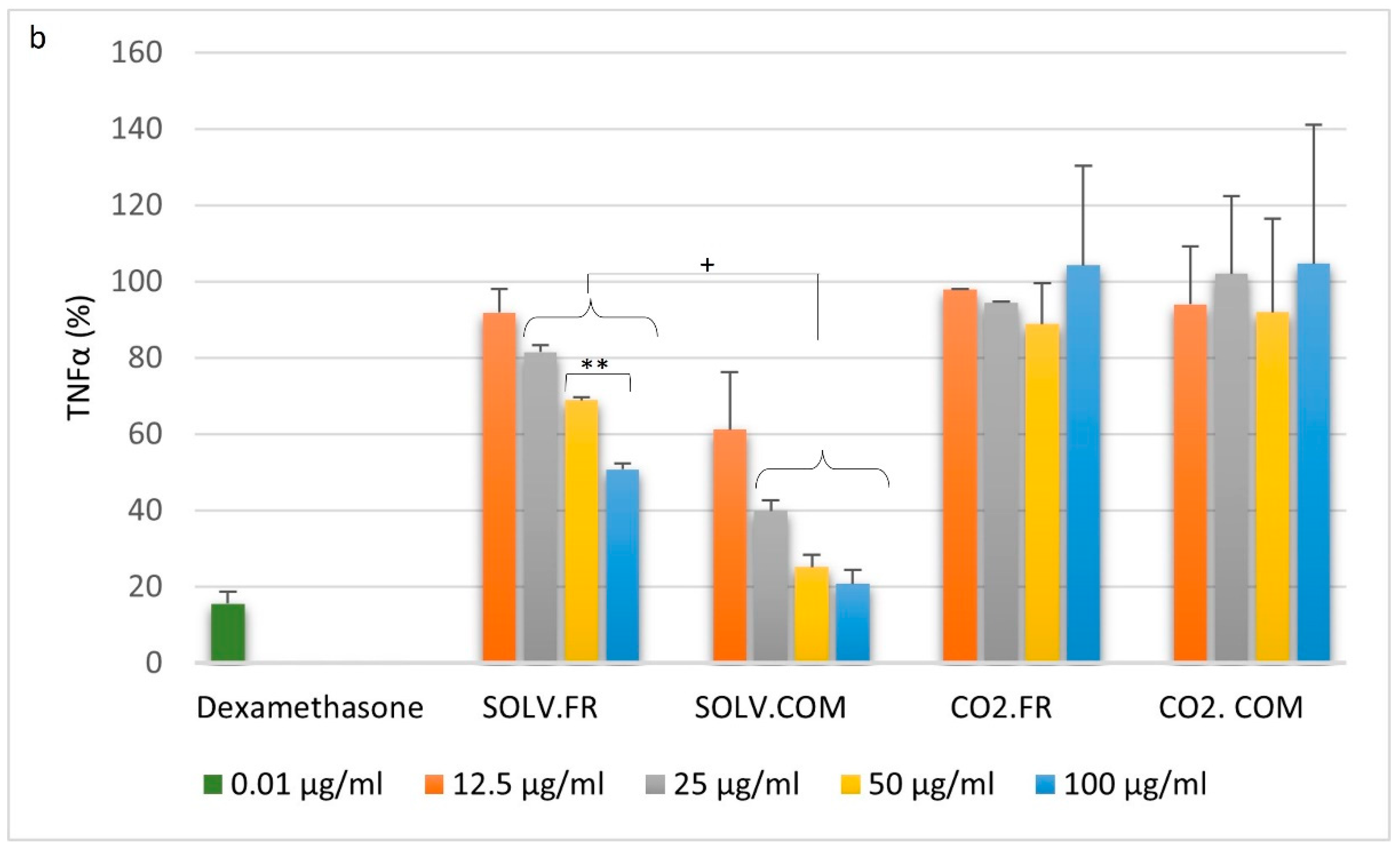
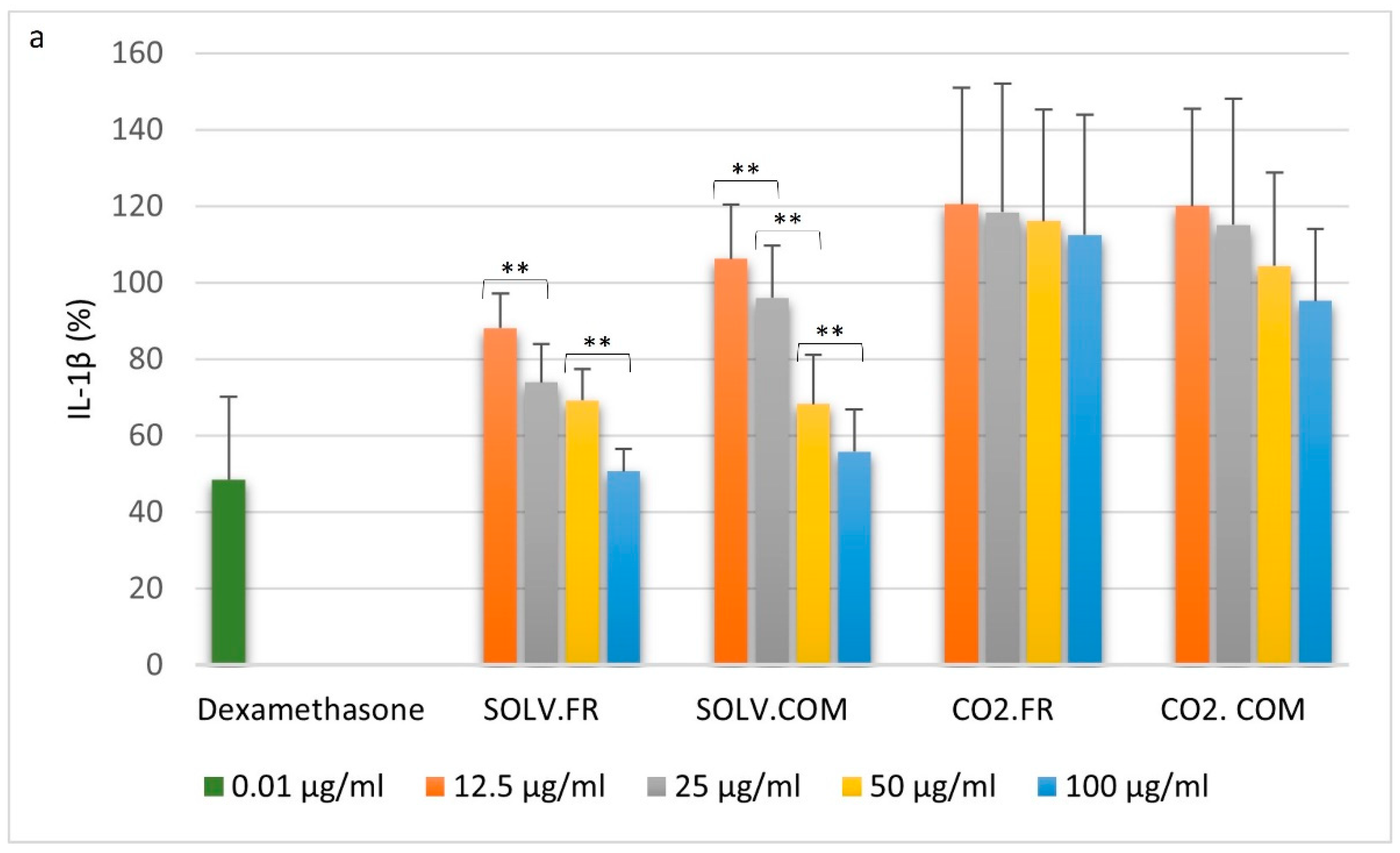
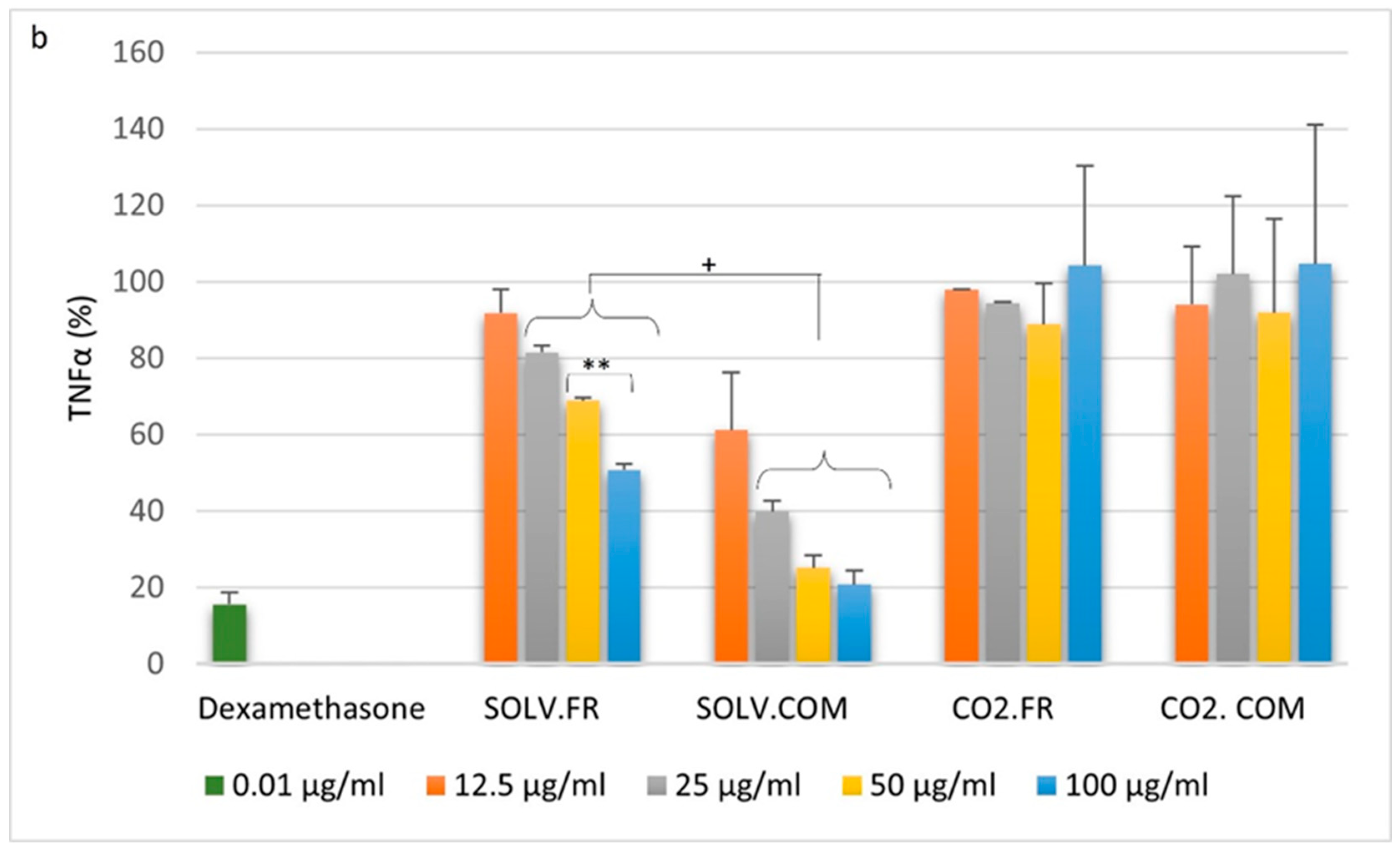
3.2.1. Monocytes
3.2.2. Macrophages
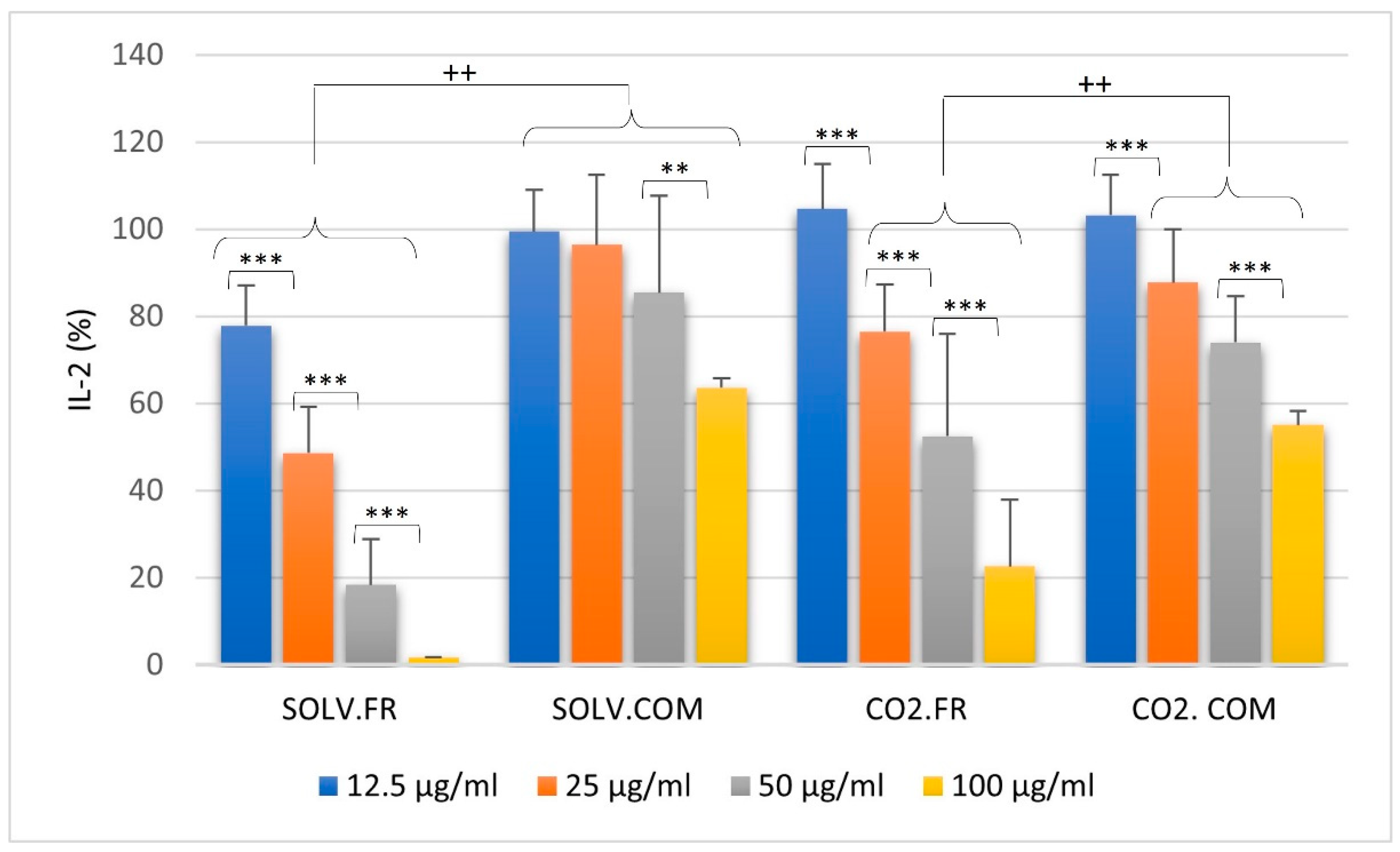
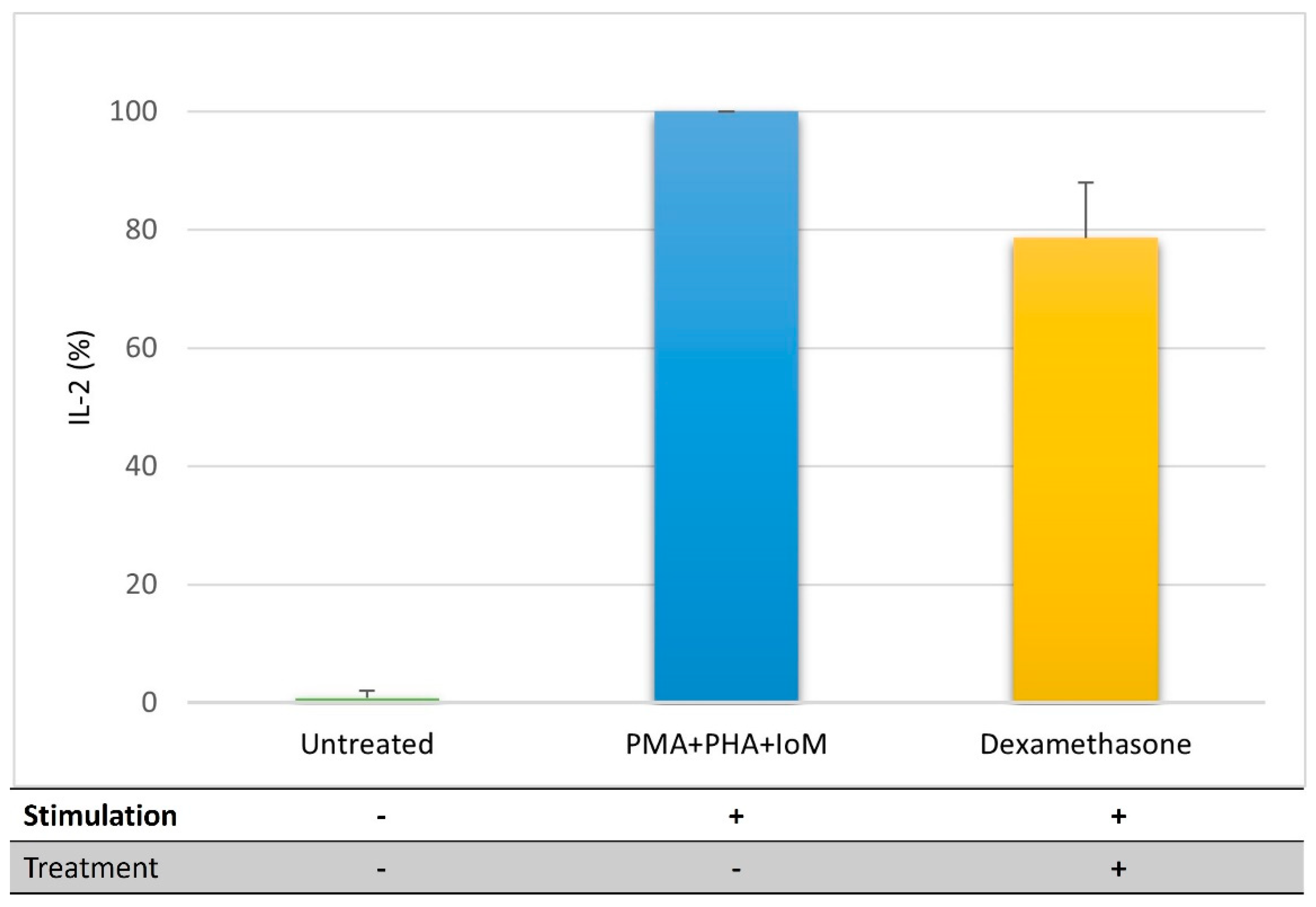
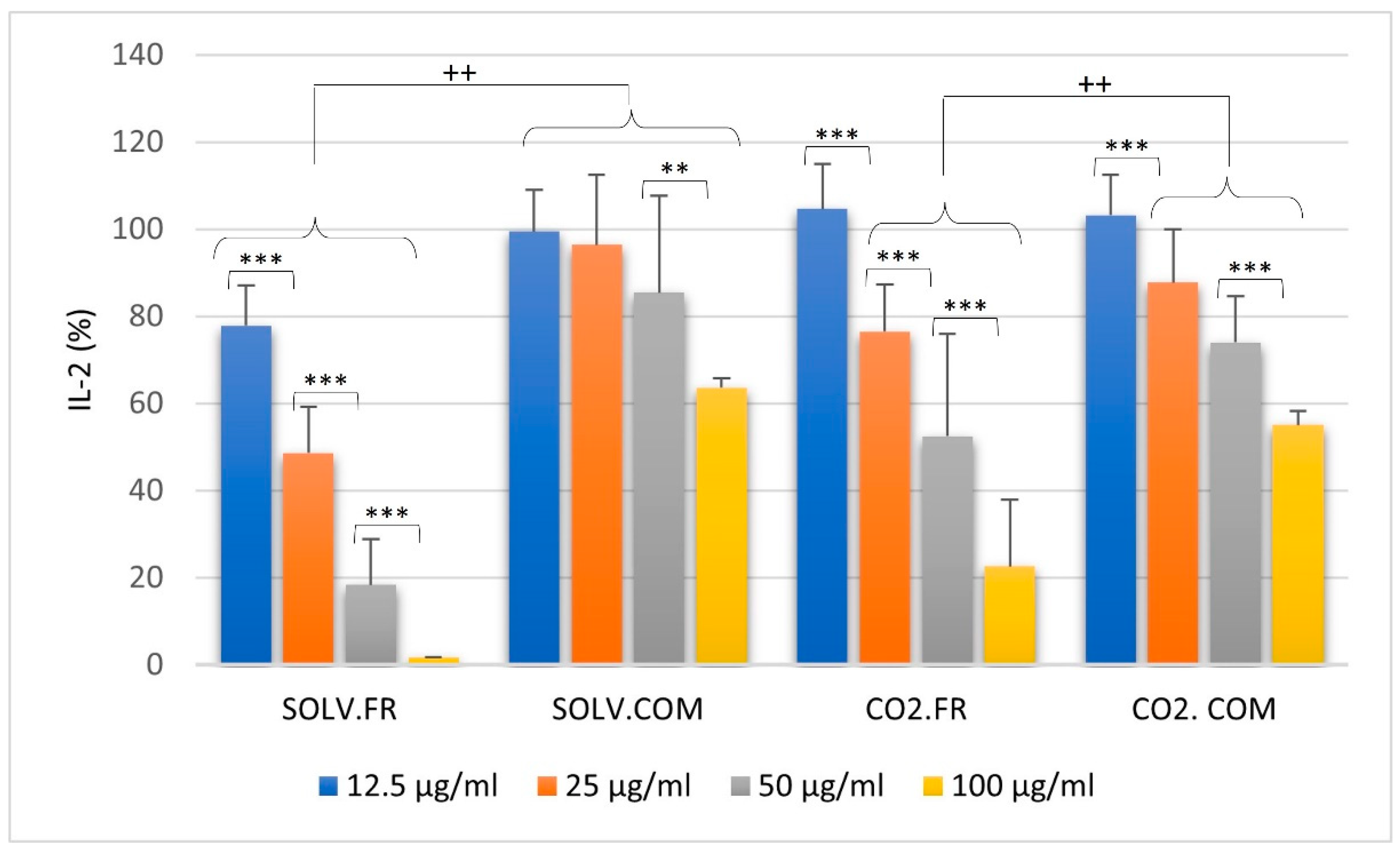
3.2.3. T lymphocytes
4. Discussion
4.1. Impact of GLM Oil on Inflammation and Immune Regulation
4.2. Comparative Analysis of Immune Modulation by GLM Lipid Extracts
4.3. Understanding Immune Modulation and Inflammation through GLM Extract Composition
4.4. Effects of Fatty Acid Ratios on Immune Cells and Cytokine Regulation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, L.; Di Benedetto, S.; Pawelec, G. Human Immune System in Aging. In Encyclopedia of Gerontology and Population Aging; Springer: Cham, Switzerland, 2019; pp. 1–12. [Google Scholar] [CrossRef]
- Margraf, A.; Perretti, M. Immune cell plasticity in inflammation: Insights into description and regulation of immune cell phenotypes. Cells 2022, 11, 1824. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125 (Suppl. S2), S3–S23. [Google Scholar] [CrossRef]
- Calder, P.C. Immunomodulation by omega-3 fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14 (Suppl. S2), 49. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, T.A.; Best, T.M.; Merrick, M.A. The dual roles of neutrophils and macrophages in inflammation. J. Athl. Train. 2006, 41, 457–465. [Google Scholar] [PubMed]
- Luster, A.D.; Alon, R.; von Andrian, U.H. Immune cell migration in inflammation: Present and future therapeutic targets. Nat. Immunol. 2005, 6, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Germolec, D.R.; Shipkowski, K.A.; Frawley, R.P.; Evans, E. Markers of Inflammation. Methods Mol. Biol. 2018, 1803, 57–79. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Lacy, P.; Stow, J.L. Cytokine release from innate immune cells: Association with diverse membrane trafficking pathways. Blood J. Am. Soc. Hematol. 2011, 118, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Hawiger, D.; Inaba, K.; Dorsett, Y.; Guo, M.; Mahnke, K.; Rivera, M.; Ravetch, J.V.; Steinman, R.M.; Nussenzweig, M.C. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 2001, 194, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Haddad, P.S.; Azar, G.A.; Groom, S.; Boivin, M. Natural health products, modulation of immune function and prevention of chronic diseases. Evid.-Based Complement. Altern. Med. 2005, 2, 513–520. [Google Scholar] [CrossRef]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef]
- Afroze, S.; Janakiraman, A.K.; Gunasekaran, B.; Djearamane, S.; Wong, L.S. Potentials of omega-3 fatty acids as therapeutic drugs and its obstacles in the pathway: A critical review. J. Pharm. Pharmacogn. Res. 2024, 12, 120–145. [Google Scholar] [CrossRef]
- Minton, S.T.; Almada, A.L.; Evans, J.L.; Laidlaw, M.; Opheim, J. Comparative membrane incorporation of omega-3 fish oil triglyceride preparations differing by degree of re-esterification: A sixteen-week randomized intervention trial. PLoS ONE 2023, 18, e0265462. [Google Scholar] [CrossRef]
- Barchi, J.J., Jr.; Strain, C.N. The effect of a methyl group on structure and function: Serine vs. threonine glycosylation and phosphorylation. Front. Mol. Biosci. 2023, 10, 1117850. [Google Scholar] [CrossRef]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef]
- Yaqoob, P.; Calder, P.C. Fatty acids and immune function: New insights into mechanisms. Br. J. Nutr. 2007, 98, S41–S45. [Google Scholar] [CrossRef]
- Utami, K.P.; Wasityastuti, W.; Soesatyo, M.H. The Involvement of Lipids in Immune System Regulation: A Mini-Review. JKKI J. Kedokt. Dan Kesehat. Indones. 2021, 12, 68–78. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of omega-3 fatty acids on immune cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef]
- Johnson, W.T.; Dorn, N.C.; Ogbonna, D.A.; Bottini, N.; Shah, N.J. Lipid-based regulators of immunity. Bioeng. Transl. Med. 2022, 7, e10288. [Google Scholar] [CrossRef]
- Radzikowska, U.; Rinaldi, A.O.; Celebi Sozener, Z.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef]
- Lawson, B.R.; Belkowski, S.M.; Whitesides, J.F.; Davis, P.; Lawson, J.W. Immunomodulation of murine collagen-induced arthritis by N, N-dimethylglycine and a preparation of Perna canaliculus. BMC Complement. Altern. Med. 2007, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- McPhee, S.; Hodges, L.D.; Wright, P.F.; Wynne, P.M.; Kalafatis, N.; Harney, D.W.; Macrides, T.A. Anti-cyclooxygenase effects of lipid extracts from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 346–356. [Google Scholar] [CrossRef]
- Mani, S.; Lawson, J.W. In vitro modulation of inflammatory cytokine and IgG levels by extracts of Perna canaliculus. BMC Complement. Altern. Med. 2006, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Treschow, A.P.; Hodges, L.D.; Wright, P.F.; Wynne, P.M.; Kalafatis, N.; Macrides, T.A. Novel anti-inflammatory omega-3 PUFAs from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, M.W.; Macrides, T.A.; Kalafatis, N.; Betts, W.H.; Haynes, D.R.; Broadbent, J. Anti-inflammatory activity of a lipid fraction (lyprinol) from the NZ green-lipped mussel. Inflammopharmacology 1997, 5, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bao, C.; Cho, S.H.; Lee, H.J. Green lipped mussel oil complex suppresses lipopolysaccharide stimulated inflammation via regulating nuclear factor-κB and mitogen activated protein kinases signaling in RAW264. 7 murine macrophages. Food Sci. Biotechnol. 2017, 26, 815–822. [Google Scholar] [CrossRef]
- Bierer, T.L.; Bui, L.M. Improvement of arthritic signs in dogs fed green-lipped mussel (Perna canaliculus). J. Nutr. 2002, 132, 1634S–1636S. [Google Scholar] [CrossRef]
- Bui, L.M.; Bierer, T.L. Influence of green lipped mussels (Perna canaliculus) in alleviating signs of arthritis in dogs. Vet. Ther. 2003, 4, 397–407. [Google Scholar] [PubMed]
- Lee, C.-H.; Lum, J.H.-K.; Ng, C.K.-C.; McKay, J.; Butt, Y.K.-C.; Wong, M.-S.; Lo, S.C.-L. Pain controlling and cytokine-regulating effects of lyprinol, a lipid extract of Perna canaliculus, in a rat adjuvant-induced arthritis model. Evid.-Based Complement. Altern. Med. 2009, 6, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fu, Y.; Zheng, J.; Li, D. Anti-inflammatory activity and mechanism of a lipid extract from hard-shelled mussel (Mytilus coruscus) on chronic arthritis in rats. Mar. Drugs 2014, 12, 568–588. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, H.; Song, X.; Kuang, X.; Liu, S.; Zhu, S.; Li, D. The preventive effect of mussel oil on gestational diabetes mellitus in pregnant mice fed by a high-fat and high-sucrose diet. Food Funct. 2023, 14, 1198–1208. [Google Scholar] [CrossRef]
- Miller, T.E.; Ormrod, D. The anti-inflammatory activity of Perna canaliculus (NZ green lipped mussel). N. Z. Med. J. 1980, 92, 187–193. [Google Scholar]
- Miller, T.; Dodd, J.; Ormrod, D.; Geddes, R. Anti-inflammatory activity of glycogen extracted from Perna canaliculus (NZ green-lipped mussel). Agents Actions 1993, 38, C139–C142. [Google Scholar] [CrossRef]
- Balkrishna, A.; Das, S.K.; Pokhrel, S.; Joshi, A.; Laxmi; Verma, S.; Sharma, V.K.; Sharma, V.; Sharma, N.; Joshi, C.S. Colchicine: Isolation, LC-MS QTof Screening, and Anticancer Activity Study of Gloriosa superba Seeds. Molecules 2019, 24, 2772. [Google Scholar] [CrossRef] [PubMed]
- Baxter, E.W.; Graham, A.E.; Re, N.A.; Carr, I.M.; Robinson, J.I.; Mackie, S.L.; Morgan, A.W. Standardized protocols for differentiation of THP-1 cells to macrophages with distinct M(IFNγ+LPS), M(IL-4) and M(IL-10) phenotypes. J. Immunol. Methods 2020, 478, 112721. [Google Scholar] [CrossRef]
- Lessa, R.C.; Ebrahimi, B.; Guan, X.; Li, Y.; Lu, J. Exploring the In Vitro Protective Effects of Green-Lipped Mussel (GLM) Oil Extract against Biomarkers of Glucose Metabolism and Inflammation in Chondrocyte Cells. Nutraceuticals 2023, 4, 1–22. [Google Scholar] [CrossRef]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef]
- Gomez-Olarte, S.; Bolanos, N.I.; Echeverry, M.; Rodriguez, A.N.; Cuellar, A.; Puerta, C.J.; Marino, A.; Gonzalez, J.M. Intermediate Monocytes and Cytokine Production Associated with Severe Forms of Chagas Disease. Front. Immunol. 2019, 10, 1671. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.F.; Wang, H. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes’ and Macrophages’ Diverse Functions in Inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef] [PubMed]
- Dong, C. Cytokine Regulation and Function in T Cells. Annu. Rev. Immunol. 2021, 39, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Gholijani, N.; Gharagozloo, M.; Kalantar, F.; Ramezani, A.; Amirghofran, Z. Modulation of cytokine production and transcription factors activities in human Jurkat T cells by thymol and carvacrol. Adv. Pharm. Bull. 2015, 5 (Suppl. S1), 653. [Google Scholar] [CrossRef] [PubMed]
- Street, N.E.; Mosmann, T.R. Functional diversity of T lymphocytes due to secretion of different cytokine patterns. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1991, 5, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Riddy, D.M.; Goy, E.; Delerive, P.; Summers, R.J.; Sexton, P.M.; Langmead, C.J. Comparative genotypic and phenotypic analysis of human peripheral blood monocytes and surrogate monocyte-like cell lines commonly used in metabolic disease research. PLoS ONE 2018, 13, e0197177. [Google Scholar] [CrossRef]
- de Bardet, J.C.; Cardentey, C.R.; González, B.L.; Patrone, D.; Mulet, I.L.; Siniscalco, D.; Robinson-Agramonte, M.d.l.A. Cell Immortalization: In Vivo Molecular Bases and In Vitro Techniques for Obtention. BioTech 2023, 12, 14. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Nascimento, C.R.; Fernandes, N.A.R.; Maldonado, L.A.G.; Junior, C.R. Comparison of monocytic cell lines U937 and THP-1 as macrophage models for in vitro studies. Biochem. Biophys. Rep. 2022, 32, 101383. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.-Y.; Rakariyatham, K. Phospholipids-Reference Module in Food Science. In Encyclopedia of Food Chemistry; Fereidoon, S., Melton, L., Varelis, P., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 546–549. [Google Scholar] [CrossRef]
- Ma, Y.; Poole, K.; Goyette, J.; Gaus, K. Introducing Membrane Charge and Membrane Potential to T Cell Signaling. Front. Immunol. 2017, 8, 1513. [Google Scholar] [CrossRef] [PubMed]
- Cauvi, D.M.; Hawisher, D.; Dores-Silva, P.R.; Lizardo, R.E.; De Maio, A. Macrophage reprogramming by negatively charged membrane phospholipids controls infection. FASEB J. 2019, 33, 2995–3009. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, T.; Aveiro, S.; Guedes, S.; Lopes, D.; Melo, T.; Neves, B.M.; Domingues, R.; Domingues, P. Multi-Omic Profiling of Macrophages Treated with Phospholipids Containing Omega-3 and Omega-6 Fatty Acids Reveals Complex Immunomodulatory Adaptations at Protein, Lipid and Metabolic Levels. Int. J. Mol. Sci. 2022, 23, 2139. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Park, K. Omega-3 and omega-6 polyunsaturated fatty acids and metabolic syndrome: A systematic review and meta-analysis. Clin. Nutr. 2020, 39, 765–773. [Google Scholar] [CrossRef]
- Liput, K.P.; Lepczynski, A.; Ogluszka, M.; Nawrocka, A.; Polawska, E.; Grzesiak, A.; Slaska, B.; Pareek, C.S.; Czarnik, U.; Pierzchala, M. Effects of Dietary n-3 and n-6 Polyunsaturated Fatty Acids in Inflammation and Cancerogenesis. Int. J. Mol. Sci. 2021, 22, 6965. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; O’Keefe, J. The Importance of Maintaining a Low Omega-6 Omega-3 Ratio for Reducing the Risk of Inflammatory Cytokine Storms. Mo. Med. 2020, 117, 539–542. [Google Scholar]
- Semenoglou, I.; Eliasson, L.; Uddstål, R.; Tsironi, T.; Taoukis, P.; Xanthakis, E. Supercritical CO2 extraction of oil from Arctic charr side streams from filleting processing. Innov. Food Sci. Emerg. Technol. 2021, 71, 102712. [Google Scholar] [CrossRef]
- Verlengia, R.; Gorjao, R.; Kanunfre, C.C.; Bordin, S.; Martins De Lima, T.; Martins, E.F.; Curi, R. Comparative effects of eicosapentaenoic acid and docosahexaenoic acid on proliferation, cytokine production, and pleiotropic gene expression in Jurkat cells. J. Nutr. Biochem. 2004, 15, 657–665. [Google Scholar] [CrossRef]
- Hidalgo, M.A.; Carretta, M.D.; Burgos, R.A. Long Chain Fatty Acids as Modulators of Immune Cells Function: Contribution of FFA1 and FFA4 Receptors. Front. Physiol. 2021, 12, 668330. [Google Scholar] [CrossRef]
- Ander, B.P.; Dupasquier, C.M.; Prociuk, M.A.; Pierce, G.N. Polyunsaturated fatty acids and their effects on cardiovascular disease. Exp. Clin. Cardiol. 2003, 8, 164. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summary (% TLE) | Organic Solvent | Supercritical CO₂ | |||
|---|---|---|---|---|---|
| Fresh | Commercial | Fresh | Commercial | ||
| Mean ± SD | p-Value | ||||
| Aliphatic Hydrocarbon | 0.68 ± 0.013 | 3.70 ± 0.68 | 7.31 ± 1.706 | 3.65 ± 1.92 | 0.03 |
| Triacylglycerol | 25.79 ± 6.63 | 25.39 ± 3.68 | 37.69 ± 2.70 | 30.07 ± 2.81 | |
| Free Fatty Acid | 27.32 ± 5.82 | 35.84 ± 6.67 | 47.47 ± 0.89 | 54.38 ± 0.0015 | 0.01 |
| Sterol | 11.03 ± 2.01 | 7.50 ± 6.28 | 6.88 ± 0.07 | 6.86 ± 0.39 | |
| Phospholipid | 35.17 ± 2.82 | 27.56 ± 4.75 | 1.96 ± 0.03 | 5.05 ± 0.50 | 0.0006 |
| Class | Free Fatty Acids | Organic Solvent | Supercritical CO₂ | ||||
|---|---|---|---|---|---|---|---|
| Fresh | Commercial | Fresh | Commercial | ||||
| Mean ± SD (mg/g) | p-Value | ||||||
| PUFA | ω-3 | cis-18:3 n-3 (ALA) | 4.15 ± 1.71 ᵃ | 8.3 ± 0.07 ᵃᵇ | 5.13 ± 0.21 | 5.31 ± 0.32 ᵇ | 0.02 |
| cis-20:3 n-3 (ETE) | 0.23 ± 0.006 ᶜ | 1.12 ± 0.08 ᶜᵈ | 0.32 ± 0.0004 | 0.52 ± 0.003 ᵈ | 0.01 | ||
| cis-20:5 n-3 (EPA) | 52.03 ± 0.006 ᵉᵏ | 45.36 ± 2.91 ᶠᵏ | 61.71 ± 6.44 ᵉᵐ | 77.16 ± 1.09 ᶠᵐ | 0.0002 | ||
| cis-22:6 n-3 (DHA) | 78.46 ± 2.19 ᶢᶤ | 24.84 ± 6.49 ᶤ | 89.35 ± 3.12 ᶢᶣ | 26.31 ± 0.07 ᶣ | 0.01 | ||
| ω-5 | cis-14:1n-5 | 0.08 ± 0.001 | 0.04 ± 0.0001 | 0.09 ± 0.0008 | 0.15 ± 0.0007 | ||
| ω-6 | trans-18:2 n-6 | 4.98 ± 2.57 | 4.87 ± 0.006 | 5.94 ± 0.23 | 5.42 ± 0.37 | ||
| cis-18:3 n-6 (GLA) | 0.3 ± 0.01 | 0.15 ± 0.002 | 0.42 ± 0.003 | 0.39 ± 0.004 | |||
| cis-20:2 n-6 | 1.05 ± 0.12 ᵑ | 2.28 ± 0.003 ᵑ | 1.36 ± 0.005 | 1.62 ± 0.009 | 0.01 | ||
| cis-20:3 n-6 (DGLA) | 0.4 ± 0.02 | 0.26 ± 0.0001 ᵀ | 0.49 ± 0.001 | 0.77 ± 0.004 ᵀ | 0.01 | ||
| cis-20:4 n-6 (AA) | 6.1 ± 2.7 | 4.42 ± 0.006 | 5.6 ± 0.08 | 7.01 ± 0.32 | |||
| ω-7 | cis-16:1n-7 | 22.52 ± 50.74 | 22.73 ± 7.09 | 26.46 ± 32.52 | 29.2 ± 5.22 | ||
| ω-9 | trans-18:1 n-9 | 7.1 ± 6.07 | 6.44 ± 0.12 | 7.46 ± 0.32 | 4.85 ± 0.41 | ||
| cis-22:1 n-9 | 0.16 ± 0.004 ᵖ | 0.41 ± 0.005 ᵖᵣ | 0.14 ± 0 ᵂ | 0.7 ± 0.0005 ᵣᵂ | 0.0001 | ||
| Monosaturated | cis-20:1 n-9 | 9.39 ± 8.99 | 4.23 ± 0.004 | 8.45 ± 0.01 | 4.73 ± 0.14 | ||
| cis-24:1 n-9 | 0.16 ± 0.002 | 0.1 ± 0.0003 | 0.12 ± 0.0009 | 0.14 ± 0.0001 | |||
| Saturated | 06:0 | 0.003 ± 0 | 0.003 ± 0 | 0.003 ± 0 | 0.0005 ± 0 | ||
| 08:0 | 0.005 ± 0 | 0.01 ± 0 | 0.0032 ± 0 | 0.0015 ± 0 | |||
| 10:0 | 0.006 ± 0 | 0.01 ± 0 | 0.0064 ± 0 | 0.0065 ± 0 | |||
| 11:0 | 0.004 ± 0 ᶵ | 0.04 ± 0 ᵤᶵ | 0.0098 ± 0 | 0.01 ± 0 ᵤ | 0.005 | ||
| 12:0 | 0.07 ± 0.002 | 0.14 ± 0.0004 | 0.08 ± 0.0018 ᵥ | 0.37 ± 0.01 ᵥ | 0.03 | ||
| 13:0 | 0.25 ± 0.02 | 0.2 ± 0.003 | 0.34 ± 0.04 | 0.23 ± 0.0001 | |||
| 14:0 | 15.51 ± 35.82 | 16.12 ± 0.79 | 17.74 ± 19.36 | 18.14 ± 0.19 | |||
| 15:0 | 2.75 ± 0.97 | 1.07 ± 0.05 | 2.8 ± 0.14 | 1.71 ± 0.06 | |||
| 16:00 | 67.4 ± 3.95 ᶻ | 35.5 ± 55.55 ᶻ | 72.15 ± 103.63 | 48.1 ± 14.96 | 0.01 | ||
| 17:0 | 3.38 ± 1.14 | 1.51 ± 0.08 | 3.03 ± 0.001 | 1.56 ± 0.02 | |||
| 18:0 | 14.26 ± 17.7 | 8.55 ± 2.26 | 12.83 ± 0.02 | 10.11 ± 0.3 | |||
| 20:0 | 0.53 ± 0.03 ᴬ | 0.12 ± 0.0004 ᴬ | 0.37 ± 0 | 0.16 ± 0.002 | 0.03 | ||
| 21:0 | 0.04 ± 0.0002 | 0.01 ± 0 | 0.03 ± 0 | 0.03 ± 0.0001 | |||
| 22:00 | 0.35 ± 0.01 ᴮ | 0.11 ± 0.0003 ᴮ | 0.21 ± 0 | 0.13 ± 0 | 0.03 | ||
| 23:00 | 0.08 ± 0.0005 ᴰ | 0.02 ± 0 ᴰ | 0.05 ± 0 | 0.02 ± 0 | 0.01 | ||
| 24:00 | 0.41 ± 0.01 ᴱ | 0.11 ± 0.0003 ᴱ | 0.22 ± 0.0001 | 0.15 ± 0.0003 | 0.03 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lessa, R.C.; Ebrahimi, B.; Li, H.; Guan, X.; Li, Y.; Lu, J. Investigation of the In Vitro Immunomodulatory Effects of Extracts from Green-Lipped Mussels (Perna canaliculus). Nutraceuticals 2024, 4, 127-146. https://doi.org/10.3390/nutraceuticals4010009
Lessa RC, Ebrahimi B, Li H, Guan X, Li Y, Lu J. Investigation of the In Vitro Immunomodulatory Effects of Extracts from Green-Lipped Mussels (Perna canaliculus). Nutraceuticals. 2024; 4(1):127-146. https://doi.org/10.3390/nutraceuticals4010009
Chicago/Turabian StyleLessa, Roberta Cardim, Belgheis Ebrahimi, Hui Li, Xiao Guan, Yan Li, and Jun Lu. 2024. "Investigation of the In Vitro Immunomodulatory Effects of Extracts from Green-Lipped Mussels (Perna canaliculus)" Nutraceuticals 4, no. 1: 127-146. https://doi.org/10.3390/nutraceuticals4010009
APA StyleLessa, R. C., Ebrahimi, B., Li, H., Guan, X., Li, Y., & Lu, J. (2024). Investigation of the In Vitro Immunomodulatory Effects of Extracts from Green-Lipped Mussels (Perna canaliculus). Nutraceuticals, 4(1), 127-146. https://doi.org/10.3390/nutraceuticals4010009