
The Response of a Leaky Gut Cell Culture Model (Caco-2/THP-1 Co-Culture) to Administration of Alternative Protein Sources

Abstract
1. Introduction
2. Materials and Methods
2.1. Products and Aqueous Extracts
2.2. Caco-2/THP1-Blue™ Co-Culture
2.3. Statistics
3. Results
3.1. TEER
3.2. Immune Markers
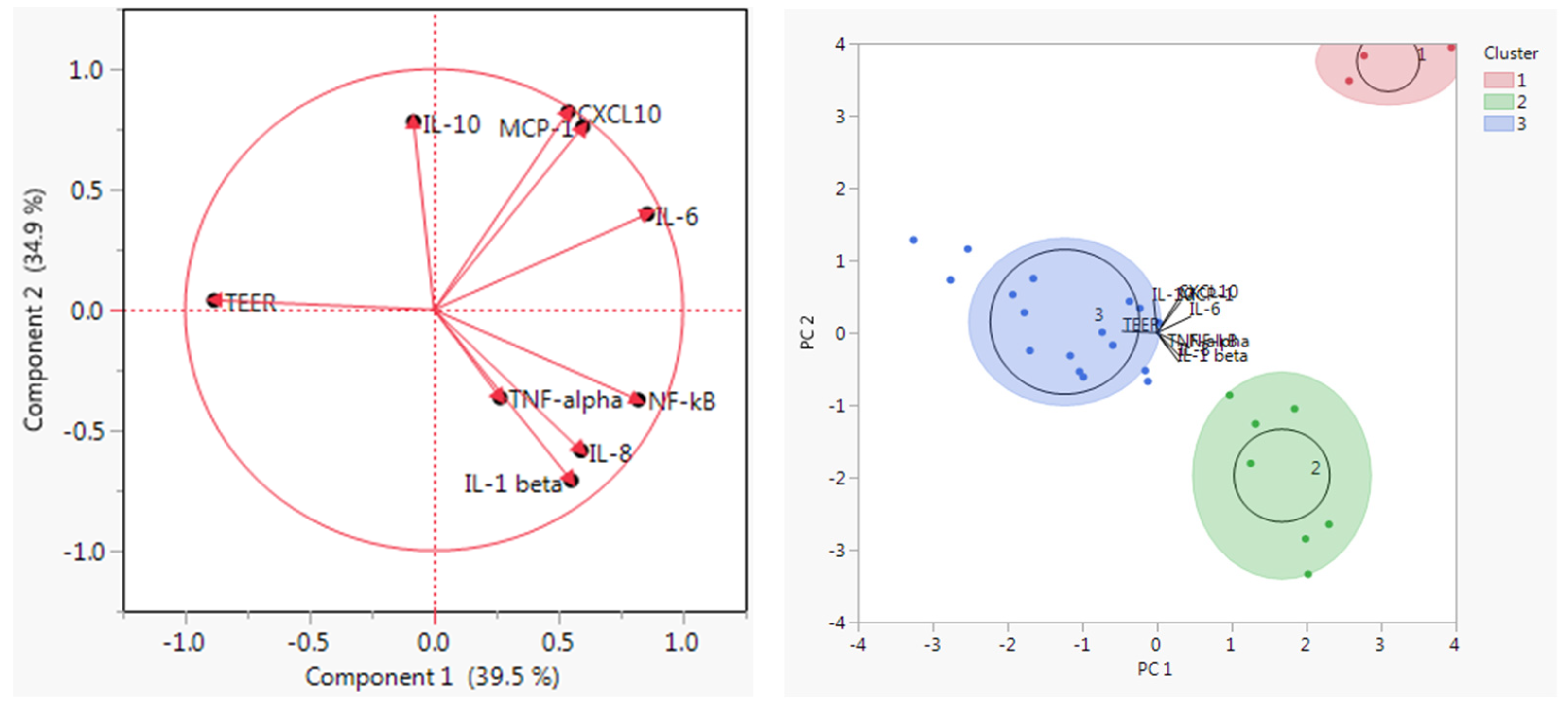
3.3. Principal Components Analysis and K Means Clustering
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hassan, S.M. Soybean, Nutrition and Health. In Soybean—Bio-Active Compounds; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2013. [Google Scholar]
- Li, D.F.; Nelssen, J.L.; Reddy, P.G.; Blecha, F.; Hancock, J.D.; Allee, G.L.; Goodband, R.D.; Klemm, R.D. Transient hypersensitivity to soybean meal in the early-weaned pig. J. Anim. Sci. 1990, 68, 1790–1799. [Google Scholar] [CrossRef]
- Engle, M.J. The role of soybean meal hypersensitivity in postweaning lag and diarrhea in piglets. J. Swine Health Prod. 1994, 2, 7–10. [Google Scholar]
- Merrifield, D.L.; Dimitroglou, A.; Bradley, G.; Baker, R.T.; Davies, S.J. Soybean meal alters autochthonous microbial populations, microvilli morphology and compromises intestinal enterocyte integrity of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2009, 32, 755–766. [Google Scholar] [CrossRef]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean meal induces intestinal inflammation in zebrafish larvae. PLoS ONE 2013, 8, e69983. [Google Scholar] [CrossRef]
- Zheng, L.; Duarte, M.E.; Loftus, A.S.; Kim, S.W. Intestinal Health of Pigs Upon Weaning: Challenges and Nutritional Intervention. Front. Vet. Sci. 2021, 8, 628258. [Google Scholar] [CrossRef]
- Romarheim, O.H.; Øverland, M.; Mydland, L.T.; Skrede, A.; Landsverk, T. Bacteria Grown on Natural Gas Prevent Soybean Meal-Induced Enteritis in Atlantic Salmon. J. Nutr. 2011, 141, 124–130. [Google Scholar] [CrossRef]
- Almeida, F.N.; Sulabo, R.C.; Stein, H.H. Amino acid digestibility and concentration of digestible and metabolizable energy in a threonine biomass product fed to weanling pigs. J. Anim. Sci. 2014, 92, 4540–4546. [Google Scholar] [CrossRef]
- Ornan, E.M.; Reifen, R. Revisiting Protein Quality Assessment to Include Alternative Proteins. Foods 2022, 11, 3740. [Google Scholar] [CrossRef]
- Aleman, R.S.; Moncada, M.; Aryana, K.J. Leaky Gut and the Ingredients That Help Treat It: A Review. Molecules 2003, 28, 619. [Google Scholar] [CrossRef]
- Cochran, K.E.; Lamson, N.G.; Whitehead, K.A. Expanding the utility of the dextran sulfate sdium (DSS) mouse model to induce a clinically relevant loss of intestinal barrier function. Peer J. 2020, 8, e8681. [Google Scholar] [CrossRef]
- Satsu, H.; Ishimoto, Y.; Nakano, T.; Mochizuki, T.; Iwanaga, T.; Shimizu, M. Induction by activated macrophage-like THP-1 cells of apoptotic and necrotic cell death in intestinal epithelial Caco-2 monolayers via tumor necrosis factor-alpha. Exp. Cell Res. 2006, 312, 3909–3919. [Google Scholar] [CrossRef] [PubMed]
- Daguet, D.; Pinheiro, I.; Verhelst, A.; Possemiers, S.; Marzorati, M. Arabinogalactan and fructooligosachharides improve the gut barrier function in distinct areas of the colon in the Simulator of the Human Intestinal Microbial Ecosystem. J. Funct. Foods 2016, 20, 369–379. [Google Scholar] [CrossRef]
- Possemiers, S.; Pinheiro, I.; Verhelst, A.; Abbeele, P.V.D.; Maignien, L.; Laukens, D.; Reeves, S.G.; Robinson, L.E.; Raas, T.; Schneider, Y.-J.; et al. A Dried Yeast Fermentate Selectively Modulates both the Luminal and Mucosal Gut Microbiota and Protects against Inflammation, As Studied in an Integrated In Vitro Approach. J. Agric. Food Chem. 2013, 61, 9380–9392. [Google Scholar]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of Butyrate on Intestinal Barrier Function in a Caco-2 Cell Monolayer Model of Intestinal Barrier. Pediatr. Res. 2007, 61, 37–41. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, G.; Sun, Z.; Che, D.; Bao, N.; Zhang, X. Effects of soybean agglutinin on intestinal barrier permeability and tight junction protein expression in weaned piglets. Int. J. Mol. Sci. 2011, 12, 8502–8512. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Kumar, V.; Hossain, M.S.; Ragaza, J.A.; Rubio Benito, M. The Potential Impacts of Soy Protein on Fish Gut Health, Soybean for Human Consumption and Animal Feed. In Soybean for Human Consumption and Animal Feed; Sudarić, A., Ed.; IntechOpen: London, UK, 2020. [Google Scholar]
- Sealey, W.M.; Barrows, F.T.; Smith, C.E.; Overturf, K.; LaPatra, S.E. Soybean meal level and probiotics in first feeding fry diets alter the ability of rainbow trout Oncorhynchus mykiss to utilize high levels of soybean meal during grow-out. Aquaculture 2009, 293, 195–203. [Google Scholar] [CrossRef]
- Lv, Z.; Fan, H.; Zhang, B.; Xing, K.; Guo, Y. Dietary genistein supplementation for breeders and their offspring improves the growth performance and immune function of broilers. Sci. Rep. 2018, 8, 5161. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Jiang, X.R.; Wei, Z.X.; Cai, L.; Yin, J.D.; Li, X.L. Effects of soybean isoflavones on the growth performance, intestinal morphology and antioxidative properties in pigs. Animal 2020, 14, 2262–2270. [Google Scholar] [CrossRef]
- Kar, S.K.; van der Hee, B.; Loonen, L.M.P.; Taverne, N.; Taverne-Thiele, J.J.; Schokker, D.; Smits, M.A.; Jansman, A.J.M.; Wells, J.M. Effects of undigested protein-rich ingredients on polarised small intestinal organoid monolayers. J. Anim. Sci. Biotechnol. 2020, 11, 51. [Google Scholar] [CrossRef]
- Bitzer, Z.T.; Wopperer, A.L.; Chrisfield, B.J.; Tao, L.; Cooper, T.K.; Vanamala, J.; Elias, R.J.; Hayes, J.E.; Lambert, J.D. Soy protein concentrate mitigates markers of colonic inflammation and loss of gut barrier function in vitro and in vivo. J. Nutr. Biochem. 2017, 40, 201–208. [Google Scholar] [CrossRef]
- Leser, T.D.; Amenuvor, J.Z.; Jensen, T.K.; Lindecrona, R.H.; Boye, M.; Møller, K. Culture-Independent Analysis of Gut Bacteria: The Pig Gastrointestinal Tract Microbiota Revisited. Appl. Environ. Microbiol. 2002, 68, 673–690. [Google Scholar] [CrossRef]
- Knecht, D.; Cholewińska, P.; Jankowska-Mąkosa, A.; Katarzyna, C. Development of Swine’s Digestive Tract Microbiota and Its Relation to Production Indices—A Review. Animals 2020, 10, 527. [Google Scholar] [CrossRef]
- Guo, W.; Zhu, S.; Feng, G.; Wu, L.; Feng, Y.; Guo, T.; Yang, Y.; Wu, H.; Zeng, M. Microalgae aqueous extracts exert intestinal protective effects in Caco-2 cells and dextran sodium sulphate-induced mouse colitis. Food Funct. 2020, 11, 1098–1109. [Google Scholar] [CrossRef]
- Lindenberg, F.C.; Ellekilde, M.; Thörn, A.C.; Kihl, P.; Larsen, C.S.; Hansen, C.H.; Metzdorff, S.B.; Aalbæk, B.; Hansen, A.K. Dietary LPS traces influences disease expression of the diet-induced obese mouse. Res. Vet. Sci. 2019, 123, 195–203. [Google Scholar] [CrossRef]
- Stephens, M.; von der Weid, P.Y. Lipopolysaccharides modulate intestinal epithelial permeability and inflammation in a species-specific manner. Gut Microbes 2020, 11, 421–432. [Google Scholar] [CrossRef]
- Shurson, G. Yeast and yeast derivatives in feed additives and ingredients: Sources, characteristics, animal responses, and quantification methods. Anim. Feed. Sci. Technol. 2018, 235, 60–76. [Google Scholar] [CrossRef]
- Kogan, G.; Kocher, A. Role of yeast cell wall polysaccharides in pig nutrition and health protection. Livest. Sci. 2007, 109, 161–165. [Google Scholar] [CrossRef]
- Lee, J.; Kyoung, H.; Cho, J.; Choe, J.; Kim, Y.; Liu, Y.; Kang, J.; Lee, H.; Kim, H.; Song, M. Dietary Yeast Cell Wall Improves Growth Performance and Prevents of Diarrhea of Weaned Pigs by Enhancing Gut Health and Anti-Inflammatory Immune Responses. Animals 2021, 11, 2269. [Google Scholar] [CrossRef]
- Grammes, F.; Reveco, F.E.; Romarheim, O.H.; Landsverk, T.; Mydland, L.T.; Øverland, M. Candida utilis and Chlorella vulgaris counteract intestinal inflammation in Atlantic salmon (Salmo salar L.). PLoS ONE 2013, 8, e83213. [Google Scholar] [CrossRef]
- Reveco-Urzua, F.E.; Hofossæter, M.; Rao Kovi, M.; Mydland, L.T.; Ånestad, R.; Sørby, R.; Press, C.M.; Lagos, L.; Øverland, M. Candida utilis yeast as a functional protein source for Atlantic salmon (Salmo salar L.): Local intestinal tissue and plasma proteome responses. PLoS ONE 2019, 14, e0218360. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Furbeyre, H.; van Milgen, J.; Mener, T.; Gloaguen, M.; Labussière, E. Effects of oral supplementation with Spirulina and Chlorella on growth and digestive health in piglets around weaning. Animal 2018, 12, 2264–2273. [Google Scholar] [CrossRef] [PubMed]
- Fields, F.J.; Lejzerowicz, F.; Schroeder, D.; Ngoi, S.M.; Tran, M.; McDonald, D.; Jiang, L.; Chang, J.T.; Knight, R.; Mayfield, S. Effects of the microalgae Chlamydomonas on gastrointestinal health. J. Funct. Foods 2020, 65, 103738. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and Anti-Inflammatory Activities of Hydrolysates and Peptide Fractions Obtained by Enzymatic Hydrolysis of Selected Heat-Treated Edible Insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef]
- Borrelli, L.; Varriale, L.; Dipineto, L.; Pace, A.; Menna, L.F.; Fioretti, A. Insect Derived Lauric Acid as Promising Alternative Strategy to Antibiotics in the Antimicrobial Resistance Scenario. Front. Microbiol. 2021, 12, 620798. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient | Extract (Pre-Diluted) | ||||||
|---|---|---|---|---|---|---|---|
| Dry Matter (%) | Crude Protein 1 (%) | Crude Fat (%) | Carbohydrates (%) | Ash (%) | Protein 2 (mg/mL) | Total Solids (mg/mL) | |
| Soybean meal 3 | 88.0 | 45.4 | 2.50 | 33.9 | 6.2 | 1.39 | 23.8 |
| Fish meal 4 | 93.6 | 60.5 | 9.7 | 3.0 | 20.4 | 1.06 | 25 |
| Cyberlindnera jadinii biomass 5 | 95.3 | 57.0 | 6.1 | 24.7 | 7.6 | 0.53 | 4.3 |
| Saccharomyces sp. biomass (dried yeast) 6 | 94.0 | 50.2 | 7.2 | 32.8 | 3.8 | 0.85 | 7.3 |
| Bio-Mos 7 | 91.9 | 31.3 | 7.2 | 48.2 | 5.2 | 1.72 | 11.3 |
| Chlorella pyrenoidosa biomass 8 | 94.5 | 58.1 | 10.5 | 19.4 | 6.5 | 0.61 | 20 |
| Methylobacterium extorquens biomass 9 | 96.2 | 82.0 | 6.6 | 5.5 | 2.1 | 1.15 | 19.2 |
| Escherichia coli biomass 10 | 93.0 | 74.1 | 8.1 | 5.9 | 4.9 | 0.93 | 18.6 |
| Hermetia illucens larvae (Black Soldier Fly) 11 | 95.5 | 40.6 | 33.8 | 15.1 | 6.0 | 0.72 | 5.2 |
| TEER (% of Initial Value) | NF-kB (OD630nm) | IL-6 (pg/mL) | IL-10 (pg/mL) | IL-1β (pg/mL) | TNF-α (pg/mL) | IL-8 (ng/mL) | CXCL10 (pg/mL) | MCP-1 (pg/mL) | |
|---|---|---|---|---|---|---|---|---|---|
| LPS+ | 86.9 | 0.65 | 163.29 | 6.22 | 283.20 | 5996.63 | 725.28 | 1646.50 | 524,244.23 |
| HC + LPS | N/A | 0.55 | 21.87 | 1.27 | 297.13 | 1242.43 | 366.62 | 195.09 | 2250.61 |
| NaB | 115.07 | 1.24 | 795.73 | 18.14 | 234.31 | 2899.94 | 727.70 | 64.12 | 4142.36 |
| LPS- | N/A | 0.41 | 3.46 | 0.00 | 32.01 | 22.00 | 186.62 | 16.30 | 5220.30 |
| TEER (% of Initial Value) | NF-kB (OD630nm) | IL-6 (pg/mL) | IL-10 (pg/mL) | IL-1β (pg/mL) | TNF-α (pg/mL) | IL-8 (ng/mL) | CXCL10 (pg/mL) | MCP-1 (pg/mL) | |
|---|---|---|---|---|---|---|---|---|---|
| Torula yeast | 135.37 A | 0.45 E | 44.54 C | 12.86 AB | 180.83 D | 5916.93 B | 543.89 BC | 383.68 BC | 6566.08 BC |
| Dried yeast | 121.41 AB | 0.48 DE | 64.14 C | 10.74 AB | 366.19 BC | 2946.69 B | 399.91 C | 346.66 BC | 4494.80 C |
| Bio-Mos | 120.94 B | 0.57 BCD | 53.45 C | 12.66 AB | 498.53 B | 5317.92 B | 897.80 ABC | 250.32 BC | 4397.87 C |
| Chlorella | 117.33 BC | 0.63 AB | 95.04 C | 8.17 BC | 445.16 BC | 4236.6 B | 659.45 BC | 559.11 B | 11,866.53 B |
| Black Soldier Fly larvae | 115.36 BC | 0.52 CDE | 79.52 C | 8.74 BC | 400.88 BC | 5167.12 B | 582.14 BC | 396.23 BC | 8477.62 BC |
| Fish meal | 110.40 BCD | 0.57 BCD | 69.19 C | 9.15 AB | 355.99 BC | 7633.55 AB | 442.35 C | 382.22 BC | 6991.48 BC |
| Methylobacterium | 103.62 CDE | 0.70 A | 100.25 BC | 2.81 C | 690.33 A | 6455.12 B | 1520.03 A | 157.34 C | 8306.42 BC |
| Soybean meal | 96.82 DEF | 0.62 ABC | 165.94 A | 15.22 A | 335.87 BCD | 3896.47 B | 734.59 BC | 1825.65 A | 524,244.23 A |
| E. coli | 95.41 EF | 0.62 ABC | 87.07 C | 9.58 AB | 479.30 B | 14,621.90 A | 1105.73 AB | 393.43 BC | 8212.45 BC |
| LPS (+) control | 86.90 F | 0.65 AB | 163.29 AB | 6.22 BC | 283.20 CD | 5996.63 AB | 724.28 BC | 1646.50 A | 524,244.23 A |
| Pooled SEM | 2.91 | 0.02 | 11.23 | 1.21 | 33.13 | 2376.97 | 126.87 | 74.18 | 1285.84 |
| p > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0052 | 0.0002 | <0.0001 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzorati, M.; Van den Abbeele, P.; Verstrepen, L.; De Medts, J.; Ekmay, R.D. The Response of a Leaky Gut Cell Culture Model (Caco-2/THP-1 Co-Culture) to Administration of Alternative Protein Sources. Nutraceuticals 2023, 3, 175-184. https://doi.org/10.3390/nutraceuticals3010013
Marzorati M, Van den Abbeele P, Verstrepen L, De Medts J, Ekmay RD. The Response of a Leaky Gut Cell Culture Model (Caco-2/THP-1 Co-Culture) to Administration of Alternative Protein Sources. Nutraceuticals. 2023; 3(1):175-184. https://doi.org/10.3390/nutraceuticals3010013
Chicago/Turabian StyleMarzorati, Massimo, Pieter Van den Abbeele, Lynn Verstrepen, Jelle De Medts, and Ricardo D. Ekmay. 2023. "The Response of a Leaky Gut Cell Culture Model (Caco-2/THP-1 Co-Culture) to Administration of Alternative Protein Sources" Nutraceuticals 3, no. 1: 175-184. https://doi.org/10.3390/nutraceuticals3010013
APA StyleMarzorati, M., Van den Abbeele, P., Verstrepen, L., De Medts, J., & Ekmay, R. D. (2023). The Response of a Leaky Gut Cell Culture Model (Caco-2/THP-1 Co-Culture) to Administration of Alternative Protein Sources. Nutraceuticals, 3(1), 175-184. https://doi.org/10.3390/nutraceuticals3010013