
The Effect of Spring Barley Fertilization on the Content of Polycyclic Aromatic Hydrocarbons, Microbial Counts and Enzymatic Activity in Soil





Abstract
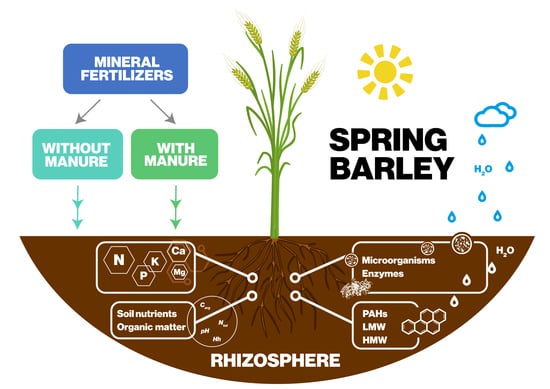
1. Introduction
2. Materials and Methods
2.1. Research Location and Experimental Design
2.2. Chemical Analyses of Soil
2.3. Microbiological and Biochemical Analyses of Soil
2.4. Statistical Analysis
2.5. Weather Conditions
3. Results and Discussion
3.1. Selected Chemical Parameters of Soil
3.1.1. Soil pH and Hydrolytic Acidity
3.1.2. Organic Carbon and Total Nitrogen Content
3.2. Microbiological and Biochemical Properties of Soil
3.2.1. Microbial Abundance
Organotrophic Bacteria
Ammonifying Bacteria
Nitrogen-Fixing Bacteria
Actinobacteria
Fungi
3.2.2. Enzymatic Activity
Dehydrogenases
Catalase
Urease
Acid Phosphatase
Alkaline Phosphatase
3.3. Content of PAHs in Soil
3.3.1. Content of LMW PAHs in Soil
3.3.2. Content of HMW PAHs in Soil
3.3.3. Total Content of 16 PAHs
3.4. Principal Component Analysis: Correlations between Selected Soil Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allamin, I.A.; Shukor, M.Y. Phytoremediation of PAHs in Contaminated Soils: A Review. Bioremediat. Sci. Technol. Res. 2021, 9, 1–6. [Google Scholar] [CrossRef]
- Lipińska, A.; Wyszkowska, J.; Kucharski, J. Microbiological and Biochemical Activity in Soil Contaminated with Pyrene Subjected to Bioaugmentation. Water Air Soil Pollut. 2021, 232, 45. [Google Scholar] [CrossRef]
- Krzebietke, S.; Mackiewicz-Walec, E.; Sienkiewicz, S.; Załuski, D. Effect of Manure and Mineral Fertilisers on the Content of Light and Heavy Polycyclic Aromatic Hydrocarbons in Soil. Sci. Rep. 2020, 10, 4573. [Google Scholar] [CrossRef] [PubMed]
- Mackiewicz-Walec, E.; Krzebietke, S.J.; Sienkiewicz, S. The Influence of Crops on the Content of Polycyclic Aromatic Hydrocarbons in Soil Fertilized with Manure and Mineral Fertilizers. Int. J. Environ. Res. Public Health 2022, 19, 13627. [Google Scholar] [CrossRef] [PubMed]
- Krzebietke, S.J.; Mackiewicz-Walec, E.; Sienkiewicz, S.; Wierzbowska, J.; Załuski, D.; Borowik, A. Polycyclic Aromatic Hydrocarbons in Soil at Different Depths under a Long-Term Experiment Depending on Fertilization. Int. J. Environ. Res. Public Health 2022, 19, 10460. [Google Scholar] [CrossRef]
- Stańczyk-Mazanek, E.; Stępniak, L.; Kępa, U. Analysis of Migration of Polycyclic Aromatic Hydrocarbons from Sewage Sludge Used for Fertilization to Soils, Surface Waters, and Plants. Water 2019, 11, 1270. [Google Scholar] [CrossRef]
- Patel, A.B.; Shaikh, S.; Jain, K.R.; Desai, C.; Madamwar, D. Polycyclic Aromatic Hydrocarbons: Sources, Toxicity, and Remediation Approaches. Front. Microbiol. 2020, 11, 562813. [Google Scholar] [CrossRef]
- Li, X.; Hou, L.; Liu, M.; Zheng, Y.; Li, Y.; Lin, X. Abundance and Diversity of Polycyclic Aromatic Hydrocarbon Degradation Bacteria in Urban Roadside Soils in Shanghai. Appl. Microbiol. Biotechnol. 2015, 99, 3639–3649. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A Review on Polycyclic Aromatic Hydrocarbons: Source, Environmental Impact, Effect on Human Health and Remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef]
- Neina, D. The Role of Soil PH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef]
- Kot-Wasik, A.; Dąbrowska, D.; Namieśnik, J. Degradacja Związków Organicznych w Środowisku. In Nowe Horyzonty i Wyzwania w Analityce i Monitoringu Środowiskowym; Politechnika Gdańska: Gdańsk, Poland, 2003; Volume 33, pp. 699–721. [Google Scholar]
- Covino, S.; Svobodová, K.; Čvančarová, M.; D’Annibale, A.; Petruccioli, M.; Federici, F.; Křesinová, Z.; Galli, E.; Cajthaml, T. Inoculum Carrier and Contaminant Bioavailability Affect Fungal Degradation Performances of PAH-Contaminated Solid Matrices from a Wood Preservation Plant. Chemosphere 2010, 79, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Sayara, T.; Sarrà, M.; Sánchez, A. Effects of Compost Stability and Contaminant Concentration on the Bioremediation of PAHs-Contaminated Soil through Composting. J. Hazard. Mater. 2010, 179, 999–1006. [Google Scholar] [CrossRef]
- Ravanipour, M.; Kalantary, R.R.; Mohseni-Bandpi, A.; Esrafili, A.; Farzadkia, M.; Hashemi-Najafabadi, S. Experimental Design Approach to the Optimization of PAHs Bioremediation from Artificially Contaminated Soil: Application of Variables Screening Development. J. Environ. Health Sci. Eng. 2015, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Wilcke, W. SYNOPSIS Polycyclic Aromatic Hydrocarbons (PAHs) in Soil—A Review. J. Plant Nutr. Soil Sci. 2000, 163, 229–248. [Google Scholar] [CrossRef]
- Johnsen, A.R.; Karlson, U. Diffuse PAH Contamination of Surface Soils: Environmental Occurrence, Bioavailability, and Microbial Degradation. Appl. Microbiol. Biotechnol. 2007, 76, 533–543. [Google Scholar] [CrossRef]
- Oleszczuk, P. Phytotoxicity of Municipal Sewage Sludge Composts Related to Physico-Chemical Properties, PAHs and Heavy Metals. Ecotoxicol. Environ. Saf. 2008, 69, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.G.; Reja, S.I.; Akhtar, N.; Sultana, M.; Deore, P.S.; Ali, F.I. Bioremediation of Polycyclic Aromatic Hydrocarbons (PAHs): Current Practices and Outlook. In Microbial Metabolism of Xenobiotic Compounds; Microorganisms for Sustainability; Arora, P.K., Ed.; Springer Singapore: Singapore, 2019; Volume 10, pp. 189–216. ISBN 9789811374616. [Google Scholar]
- Maliszewska-Kordybach, B. Dissipation of Polycyclic Aromatic Hydrocarbons in Freshly Contaminated Soils—The Effect of Soil Physicochemical Properties and Aging. Water Air Soil Pollut. 2005, 168, 113–128. [Google Scholar] [CrossRef]
- Maliszewska-Kordybach, B. Wpływ Wapnowania Na Trwałość Wielopierścieniowych Węglowodorów Aromatycznych w Glebach. Arch. Ochr. Śr. 1991, 3–4, 69–78. [Google Scholar]
- Krzebietke, S.J.; Wierzbowska, J.; Żarczyński, P.J.; Sienkiewicz, S.; Bosiacki, M.; Markuszewski, B.; Nogalska, A.; Mackiewicz-Walec, E. Content of PAHs in Soil of a Hazel Orchard Depending on the Method of Weed Control. Environ. Monit. Assess. 2018, 190, 422. [Google Scholar] [CrossRef]
- Maliszewska-Kordybach, B. Mikrobiologiczne Przemiany Wielopierścieniowych Węglowodorów Aromatycznych w Środowisku Glebowym. Post. Mikrobiol. 1987, 26, 233–247. [Google Scholar]
- Bala, S.; Garg, D.; Thirumalesh, B.V.; Sharma, M.; Sridhar, K.; Inbaraj, B.S.; Tripathi, M. Recent Strategies for Bioremediation of Emerging Pollutants: A Review for a Green and Sustainable Environment. Toxics 2022, 10, 484. [Google Scholar] [CrossRef] [PubMed]
- Nzila, A.; Musa, M.M. Current Status of and Future Perspectives in Bacterial Degradation of Benzo[a]Pyrene. Int. J. Environ. Res. Public Health 2020, 18, 262. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. International Soil Classifcation System for Naming Soils and Creating Legends for Soil Maps. World Reference Base for Soil Resources 2014, Update 2015; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015. [Google Scholar]
- Mackiewicz-Walec, E.; Krzebietke, S.J. Content of Polycyclic Aromatic Hydrocarbons in Soil in a Multi-Annual Fertilisation Regime. Environ. Monit. Assess. 2020, 192, 314. [Google Scholar] [CrossRef]
- Bojes, H.K.; Pope, P.G. Characterization of EPA’s 16 Priority Pollutant Polycyclic Aromatic Hydrocarbons (PAHs) in Tank Bottom Solids and Associated Contaminated Soils at Oil Exploration and Production Sites in Texas. Regul. Toxicol. Pharmacol. 2007, 47, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Smreczak, B.; Maliszewska-Kordybach, B. Seeds Germination and Root Growth of Selected Plants in Pah Contaminated Soil. Fresenius Environ. Bull. 2003, 12, 946–949. [Google Scholar]
- Alexander, M. Microorganismus and Chemical Pollution. Bioscience 1973, 23, 509–515. [Google Scholar] [CrossRef]
- Wyszkowska, J. Biologiczne Właściwości Gleby Zanieczyszczonej Chromem Sześciowartościowym. Wyd. UWM Rozpr. Monogr. 2002, 65, 1–134. [Google Scholar]
- Parkinson, D.; Gray, F.R.G.; Williams, S.T. Methods for Studying the Ecology of Soil Microorganism; Blackweel: Oxford, UK, 1971; p. 19. [Google Scholar]
- Martin, J. Use of Acid Rose Bengal and Streptomycin in the Plate Method for Estimating Soil Fungi. Soil Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Schinner, F.; Öhlinger, R.; Margiesin, R. Dehydrogenase Activity Aith the Substrate TTC. In Methods Soil Biol; Springer: Berlin/Heidelberg, Germany, 1996; p. 426. [Google Scholar]
- Alef, K.; Nanniperi, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA; Harcourt Brace & Company: London, UK, 1998; 576p. [Google Scholar]
- Johnson, J.I.; Temple, K.L. Some Variables Affecting the Measurement of Catalase Activity in Soil. Soil Sci. Soc. Am. Proc. 1964, 28, 207–216. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Zaborowska, M.; Kucharski, J. Mitigation of the Adverse Impact of Copper, Nickel, and Zinc on Soil Microorganisms and Enzymes by Mineral Sorbents. Materials 2022, 15, 5198. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Wyszkowski, M. Resistance of Aerobic Microorganisms and Soil Enzyme Response to Soil Contamination with Ekodiesel Ultra Fuel. Environ. Sci. Pollut. Res. 2017, 24, 24346–24363. [Google Scholar] [CrossRef] [PubMed]
- Borowik, A.; Wyszkowska, J.; Kucharski, M.; Kucharski, J. Implications of Soil Pollution with Diesel Oil and BP Petroleum with ACTIVE Technology for Soil Health. Int. J. Environ. Res. Public Health 2019, 16, 2474. [Google Scholar] [CrossRef] [PubMed]
- StatSoft, Inc. STATISTICA Data Analysis Software System, version 13.1; StatSoft, Inc.: Tulsa, OK, USA, 2014. [Google Scholar]
- Lemanowicz, J. Mineral Fertilisation as a Factor Determining Selected Sorption Properties of Soil against the Activityof Phosphatases. Plant Soil Environ. 2013, 59, 439–445. [Google Scholar] [CrossRef]
- Carney, K.M.; Hungate, B.A.; Drake, B.G.; Megonigal, J.P. Altered Soil Microbial Community at Elevated CO 2 Leads to Loss of Soil Carbon. Proc. Natl. Acad. Sci. USA 2007, 104, 4990–4995. [Google Scholar] [CrossRef] [PubMed]
- Amezcua-Allieri, M.A.; Ávila-Chávez, M.A.; Trejo, A.; Meléndez-Estrada, J. Removal of Polycyclic Aromatic Hydrocarbons from Soil: A Comparison between Bioremoval and Supercritical Fluids Extraction. Chemosphere 2012, 86, 985–993. [Google Scholar] [CrossRef]
- Farahani, M.; Mirbagheri, S.A.; Javid, A.H.; Karbassi, A.R.; Khorasani, N.J. Biodegradation and Leaching of Polycyclic Aromatic Hydrocarbons in Soil Column. Food Agric. Environ. 2010, 8, 870–875. [Google Scholar]
- Sienkiewicz, S. Oddziaływanie Obornika i Nawozów Mineralnych Na Kształtowanie Żyzności i Produktywności Gleby. UWM Olszt. Rozpr. Monogr. 2003, 74, 1–120. [Google Scholar]
- Czekała, J. Impact of Long-Term Plant Cultivation without Participation of Cereals on Concentrations of Nitrogen Forms in Soil Humus Horizons. J. Res. Appl. Agric. Eng. 2010, 55, 49–53. [Google Scholar]
- Siwik-Ziomek, A.; Lemanowicz, J. The Influence of Fertilization with Phosphorus, Sulphate, Carbon and Nitrogen Content on Hydrolases Activities in Soil. Pol. J. Soil Sci. 2017, 49, 49. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Kucharski, M. Wpływ Temperatury Gleby Na Drobnoustroje. Zesz. Probl. Post. Nauk Rol. 2011, 567, 55–67. [Google Scholar]
- Dąbek-Szreniawska, M.; Kozak, M.A.; Pudło, A.A. Liczebność Bakterii i Aktywność Biochemiczna Gleby Torfowej i Murszowej. Ann. UMCS Ser. E 2004, 59, 2023–2032. [Google Scholar]
- Vetanovetz, R.; Peterson, J. Effect of Carbon Source and Nitrogen on Urease Activity in a Sphagnum Peat Medium. Comm. Soil Sci. Plant Anal. 1992, 23, 379–388. [Google Scholar] [CrossRef]
- Barabasz, W.; Vořišek, K. Bioróżnorodność Mikroorganizmów w Środowiskach Glebowych. In Aktywność Drobnoustrojów w Różnych Środowiskach; AR: Kraków, Poland, 2002; pp. 26–27. [Google Scholar]
- Natywa, M.; Sawicka, A.; Wolna-Maruwka, A. Aktywność Mikrobiologiczna i Enzymatyczna Gleby Pod Uprawą Kukurydzy w Zależności Od Zróżnicowanego Nawożenia Azotem. Woda Sr. Obsz. Wiej. 2010, 10, 111–120. [Google Scholar]
- Sosnowski, J.; Jankowski, K.; Ciepiela, G.A.; Wiśniewska-Kadżajan, B.; Deska, J. Wpływ Fitohormonów i Zróżnicowanych Dawek Azotu Stosowanych w Uprawie Festulolium Braunii z Koniczyną Łąkową Na Liczebność Mikroorganizmów Glebowych. Ochr. Środow. Zas. Nat. 2012, 3, 19–30. [Google Scholar]
- Sosnowski, J.; Jankowski, K. Ocena Liczebności Mikroorganizmów Glebowych Spod Uprawy Mieszanek Festulolium Braunii z Roślinami Motylkowatymi Nawożonych Zróżnicowanymi Dawkami Azotu. Fragm. Agron. 2013, 30, 129–137. [Google Scholar]
- Niewiadomoska, A.; Kleiber, T.; Klama, J.; Swędrzyńska, D. Wpływ Zróżnicowanego Nawożenia Azotowego Na Dynamikę Składu Mikrobiologicznego Gleby i Aktywność Enzymatyczną Dehydrogenaz Pod Trawnikiem. Nauka Przyr. Technol. 2010, 4, 90. [Google Scholar]
- Brzezińska, M. Aktywność Biologiczna Oraz Procesy Jej Towarzyszące w Glebach Organicznych Nawadnianych Oczyszczonymi Ściekami Miejskimi. Acta Agrophys. Rozpr. Monogr. 2006, 131, 1–164. [Google Scholar]
- Wyszkowska, J.; Kucharski, J. Biologiczne Właściwości Gleby Zanieczyszczonej Chwastoxem Trio 540 SL. Rocz. Glebozn. 2004, 55, 311–319. [Google Scholar]
- Ciarkowska, K.; Gambuś, F. Aktywność Dehydrogenaz w Glebach Zanieczyszczonych Metalami Ciężkimi w Rejonie Olkusza. Zesz. Probl. Post. Nauk Rol. 2004, 501, 79–85. [Google Scholar]
- Koper, J.; Siwik-Ziomek, A. Wpływ Wieloletniego Nawożenia Na Aktywność Dehydrogenaz w Glebie Oraz Zawartość Węgla Organicznego i Azotu Ogółem. Zesz. Probl. Post. Nauk Rol. 2006, 515, 197–202. [Google Scholar]
- Saha, S.; Prakash, V.; Kundu, S.; Kumar, N.; Mina, B.L. Soil Enzymatic Activity as Affected by Long Term Application of Farm Yard Manure and Mineral Fertilizer under a Rainfed Soybean–Wheat System in N-W Himalaya. Eur. J. Soil Biol. 2008, 44, 309–315. [Google Scholar] [CrossRef]
- Piotrowska, A.; Koper, J. Wpływ Wieloletniego Nawożenia Organicznego i Mineralnego Na Aktywność Enzymów Oksydoredukcyjnych i Hydrolitycznych w Glebie Spod Pszenicy Ozimej (Triticum aestivum L.). Zesz. Probl. Post. Nauk Rol. 2007, 520, 669–673. [Google Scholar]
- Kucharski, J.; Wałdowska, E. Biological Results of Long-Term Fertilization with Slurry. Zesz. Probl. Post. Nauk Rol. 2001, 476, 197–204. [Google Scholar]
- Kucharski, J. Relacje Między Aktywnością a Żyznością Gleby. In Drobnoustroje w Środowisku. Występowanie, Aktywność i Znaczenie; Red. W. Barabasz: Kraków, Poland, 1997; pp. 327–347. [Google Scholar]
- Lemanowicz, J.; Koper, J. Zawartość Organicznych Związków Węgla i Fosforu Oraz Aktywność Enzymatyczna Ryzosfery Kukurydzy. Rocz. Glebozn 2009, 60, 55–59. [Google Scholar]
- Zaborowska, M.; Wyszkowska, J.; Kucharski, J. Aktywność Mikrobiologiczna Zanieczyszczonej Cynkiem Gleby o Zróżnicowanym PH. Pierwiastki Śladowe—Kryteria Jakości Środowiska Przyrodniczego. In Proceedings of the IX Sympozjum Pierwiastki Śladowe w Środowisku, Sarnówek, Poland, 11–12 May 2006; pp. 250–251. [Google Scholar]
- Kalembasa, S.; Kuzimierska, B. Wpływ Zanieczyszczenia Gleby Niklem Na Tle Nawożenia Organicznego Na Aktywność Dehydrogenaz. Ochr. Sr. Zasobów Nat. 2009, 20, 470–478. [Google Scholar]
- Lemanowicz, J.; Bartkowiak, A.; Breza-Boruta, B. Changes in Phosphorus Content, Phosphatase Activity and Some Physicochemical and Microbiological Parameters of Soil within the Range of Impact of Illegal Dumping Sites in Bydgoszcz (Poland). Environ. Earth Sci. 2016, 75, 510. [Google Scholar] [CrossRef]
- Lemanowicz, J.; Koper, J. Aktywność Enzymów Oksydoredukcyjnych i Zawartość Węgla i Azotu w Ryzosferze Jęczmienia Jarego Nawożonego Obornikiem i Azotem. Zesz. Probl. Post. Nauk Rol. 2010, 556, 437–442. [Google Scholar]
- Kucharski, J.; Wyszkowska, J.; Borowik, A. Aktywnośc Ureazy w Glebie Nawożonej Obornkiem i Kompostem. Zesz. Post. Nauk Roln. 2009, 537, 217–225. [Google Scholar]
- Lemanowicz, J.; Koper, J. Zawartość Wybranych form Fosforu w Glebie i Koniczynie Łąkowej Oraz Aktywność Fosfataz Glebowych Na Tle Zróżnicowanego Nawożenia Mineralnego i Organicznego. Woda Sr. Obsz. Wiej. 2009, 9, 119–139. [Google Scholar]
- Wittmann, C.; Kähkönen, M.A.; Ilvesniemi, H.; Kurola, J.; Salkinoja-Salonen, M.S. Areal Activities and Stratification of Hydrolytic Enzymes Involved in the Biochemical Cycles of Carbon, Nitrogen, Sulphur and Phosphorus in Podsolized Boreal Forest Soils. Soil Biol. Biochem. 2004, 36, 425–433. [Google Scholar] [CrossRef]
- Kuziemska, B.; Kalembasa, S.; Kalembasa, D. Wpływ Wapnowania i Materii Organicznej Na Aktywność Fosfataz w Glebie Zanieczyszczonej Niklem. Inż. Ekol. 2014, 37, 117–127. [Google Scholar]
- Lemanowicz, J.; Krzyżaniak, M. Vertical Distribution of Phosphorus Concentrations, Phosphatase Activity and Further Soil Chemical Properties in Salt-Affected Mollic Gleysols in Poland. Environ. Earth Sci. 2015, 74, 2719–2728. [Google Scholar] [CrossRef]
- Szara, E.; Mercik, S.; Sosulski, T. Formy Fosforu w Doświadczeniach Wieloletnich. Fragm. Agron. 2005, 1, 298–309. [Google Scholar]
- Sienkiewicz, S.; Krzebietke, S.; Wojnowska, T.; Żarczyński, P.; Omilian, M. Effect of Long-Term Differentiated Fertilization with Farmyard Manure and Mineral Fertilizers on the Content of Available Forms of P, K and Mg in Soil. J. Elem. 2009, 14, 779–786. [Google Scholar] [CrossRef]
- Lemanowicz, J.; Bartkowiak, A. Diagnosis of the Content of Selected Heavy Metals in the Soils of the Pałuki Region Against Their Enzymatic Activity. Arch. Environ. Prot. 2013, 39, 23–32. [Google Scholar] [CrossRef]
- Eriksson, M.; Sodersten, E.; Yu, Z.; Dalhammar, G.; Mohn, W.W. Degradation of Polycyclic Aromatic Hydrocarbons at Low Temperature under Aerobic and Nitrate-Reducing Conditions in Enrichment Cultures from Northern Soils. Appl. Environ. Microbiol. 2003, 69, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ni, H.-G.; Sun, J.-L.; Jing, X.; He, J.-S.; Zeng, H. Polycyclic Aromatic Hydrocarbons in Soils from the Tibetan Plateau, China: Distribution and Influence of Environmental Factors. Environ. Sci. Process. Impacts 2013, 15, 661. [Google Scholar] [CrossRef]
- Maliszewska-Kordybach, B.; Smreczak, B.; Klimkowicz-Pawlas, A.; Terelak, H. Monitoring of the Total Content of Polycyclic Aromatic Hydrocarbons (PAHs) in Arable Soils in Poland. Chemosphere 2008, 73, 1284–1291. [Google Scholar] [CrossRef]
- Gałązka, A.; Król, M.; Perzyński, A. Zmiany Aktywności Biologicznej Gleb Skażonych Wielopierścieniowymi Węglowodorami Aromatycznymi (WWA) i Olejem Napędowym (ON) w Ryzosferze Kukurydzy. Zesz. Probl. Post. Nauk Roln. 2009, 535, 111–120. [Google Scholar]
- Samanta, S.K.; Singh, O.V.; Jain, R.K. Polycyclic Aromatic Hydrocarbons: Environmental Pollution and Bioremediation. Trends Biotechnol. 2002, 20, 243–248. [Google Scholar] [CrossRef]
- Wielgosz, E.; Szember, A. Wpływ Wybranych Roślin Na Liczebności i Aktywność Drobnoustrojów Glebowych. Ann. UMCS Sec. E 2006, 61, 107–119. [Google Scholar]
- Maliszewska-Kordybach, B.; Smreczak, B.; Klimkowicz-Pawlas, A. Concentrations, Sources, and Spatial Distribution of Individual Polycyclic Aromatic Hydrocarbons (PAHs) in Agricultural Soils in the Eastern Part of the EU: Poland as a Case Study. Sci. Total Environ. 2009, 407, 3746–3753. [Google Scholar] [CrossRef]
- Lipińska, A.; Wyszkowska, J.; Kucharski, J. Urease Activity in Soil Contaminated with Polycyclic Aromatic Hydrocarbons. Environ. Sci. Pollut. Res. 2015, 22, 18519–18530. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Wyszkowski, M. Activity of Soil Dehydrogenases, Urease, and Acid and Alkaline Phosphatases in Soil Polluted with Petroleum. J. Toxicol. Environ. Health A 2010, 73, 1202–1210. [Google Scholar] [CrossRef]
- Klimkowicz-Pawlas, A.; Maliszewska-Kordybach, B. Effect of Anthracene and Pyrene on Dehydrogenases Activity in Soils Exposed and Unexposed to PAHs. Water. Air. Soil Pollut. 2003, 145, 169–186. [Google Scholar] [CrossRef]
- Smreczak, B.; Klimkowicz-Pawlas, A.; Maliszewska-Kordybach, B. Biodostępność Trwałych Zanieczyszczeń Organicznych (TZO) w Glebach. Stud. Rap. IUNG-PI 2013, 35, 137–153. [Google Scholar]
- Ukalska-Jaruga, A.; Smreczak, B. The Impact of Organic Matter on Polycyclic Aromatic Hydrocarbon (PAH) Availability and Persistence in Soils. Molecules 2020, 25, 2470. [Google Scholar] [CrossRef] [PubMed]
- Maliszewska-Kordybach, B.; Smreczak, B.; Klimkowicz-Pawlas, A. Effects of Anthropopressure and Soil Properties on the Accumulation of Polycyclic Aromatic Hydrocarbons in the Upper Layer of Soils in Selected Regions of Poland. Appl. Geochem. 2009, 24, 1918–1926. [Google Scholar] [CrossRef]
- Wyszkowski, M.; Ziółkowska, A. Zawartość Węgla Organicznego i Niektórych Makropierwiastków w Glebie Zanieczyszczonej Benzyną i Olejem Napędowym. Ochr. Sr. Zasobów Nat. 2009, 40, 112–122. [Google Scholar]
- Jin, A.; He, J.; Chen, S.; Huang, G. Distribution and Transport of PAHs in Soil Profiles of Different Water Irrigation Areas in Beijing, China. Environ. Sci. Process. Impacts 2014, 16, 1526. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Luo, W.; Gao, J.; Xu, L.; Guo, J.; Zhang, Q.; Romesh, K.Y.; Giesy, J.P.; Kang, S.; de Boer, J. Polycyclic Aromatic Hydrocarbons in Soils from the Central-Himalaya Region: Distribution, Sources, and Risks to Humans and Wildlife. Sci. Total Environ. 2016, 556, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar Effects on Soil Biota—A Review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | N0P0K0 | N1P1K1 | N2P1K1 | N3P1K1 | N2P1K2 | N2P1K3 | N2P1K2Mg | N2P1K2MgCa |
|---|---|---|---|---|---|---|---|---|
| pH (1 mol KCl dm−3) | ||||||||
| Manure | 5.7 | 5.8 | 5.3 | 5.1 | 5.2 | 5.3 | 5.2 | 6.3 |
| Without manure | 5.0 | 4.9 | 4.6 | 4.5 | 4.6 | 4.6 | 4.6 | 5.5 |
| Hh (mmol(+) kg−1) | ||||||||
| Manure | 17.7 | 16.7 | 25.6 | 26.9 | 25.9 | 24.2 | 25.0 | 13.2 |
| Without manure | 25.3 | 26.3 | 31.6 | 34.0 | 29.6 | 29.2 | 27.3 | 14.0 |
| Source of Variation | df | Organic Carbon | Total Nitrogen |
|---|---|---|---|
| Manure (M) | 1 | ** | * |
| Block (B) | 2 | ns | ns |
| Mineral fertilization (Min) | 7 | ** | ** |
| M × Min | 7 | ** | ** |
| Error 1 | 30 | - | - |
| Date (D) | 3 | ** | ** |
| D × M | 3 | ** | ** |
| D × B | 6 | ns | ns |
| D × Min | 21 | ** | ** |
| D × M × Min | 21 | ** | ** |
| Error 2 | 90 | - | - |
| Source of Variation | df | PAHs | ||
|---|---|---|---|---|
| LMW (9) | HMW (7) | Total (16) | ||
| Manure (M) | 1 | ** | ** | ** |
| Block (B) | 2 | ns | ns | ns |
| Mineral fertilization (Min) | 7 | ** | ** | ** |
| M × Min | 7 | ** | ** | ** |
| Error 1 | 30 | - | - | - |
| Date (D) | 3 | ** | ** | ** |
| D × M | 3 | ** | ** | ** |
| D × B | 6 | ns | ns | ns |
| D × Min | 21 | ** | ** | ** |
| D × M × Min | 21 | ** | ** | ** |
| Error 2 | 90 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mackiewicz-Walec, E.; Krzebietke, S.J.; Borowik, A.; Klasa, A. The Effect of Spring Barley Fertilization on the Content of Polycyclic Aromatic Hydrocarbons, Microbial Counts and Enzymatic Activity in Soil. Int. J. Environ. Res. Public Health 2023, 20, 3796. https://doi.org/10.3390/ijerph20053796
Mackiewicz-Walec E, Krzebietke SJ, Borowik A, Klasa A. The Effect of Spring Barley Fertilization on the Content of Polycyclic Aromatic Hydrocarbons, Microbial Counts and Enzymatic Activity in Soil. International Journal of Environmental Research and Public Health. 2023; 20(5):3796. https://doi.org/10.3390/ijerph20053796
Chicago/Turabian StyleMackiewicz-Walec, Ewa, Sławomir Józef Krzebietke, Agata Borowik, and Andrzej Klasa. 2023. "The Effect of Spring Barley Fertilization on the Content of Polycyclic Aromatic Hydrocarbons, Microbial Counts and Enzymatic Activity in Soil" International Journal of Environmental Research and Public Health 20, no. 5: 3796. https://doi.org/10.3390/ijerph20053796
APA StyleMackiewicz-Walec, E., Krzebietke, S. J., Borowik, A., & Klasa, A. (2023). The Effect of Spring Barley Fertilization on the Content of Polycyclic Aromatic Hydrocarbons, Microbial Counts and Enzymatic Activity in Soil. International Journal of Environmental Research and Public Health, 20(5), 3796. https://doi.org/10.3390/ijerph20053796