
Sex Differences in VO2max and the Impact on Endurance-Exercise Performance



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
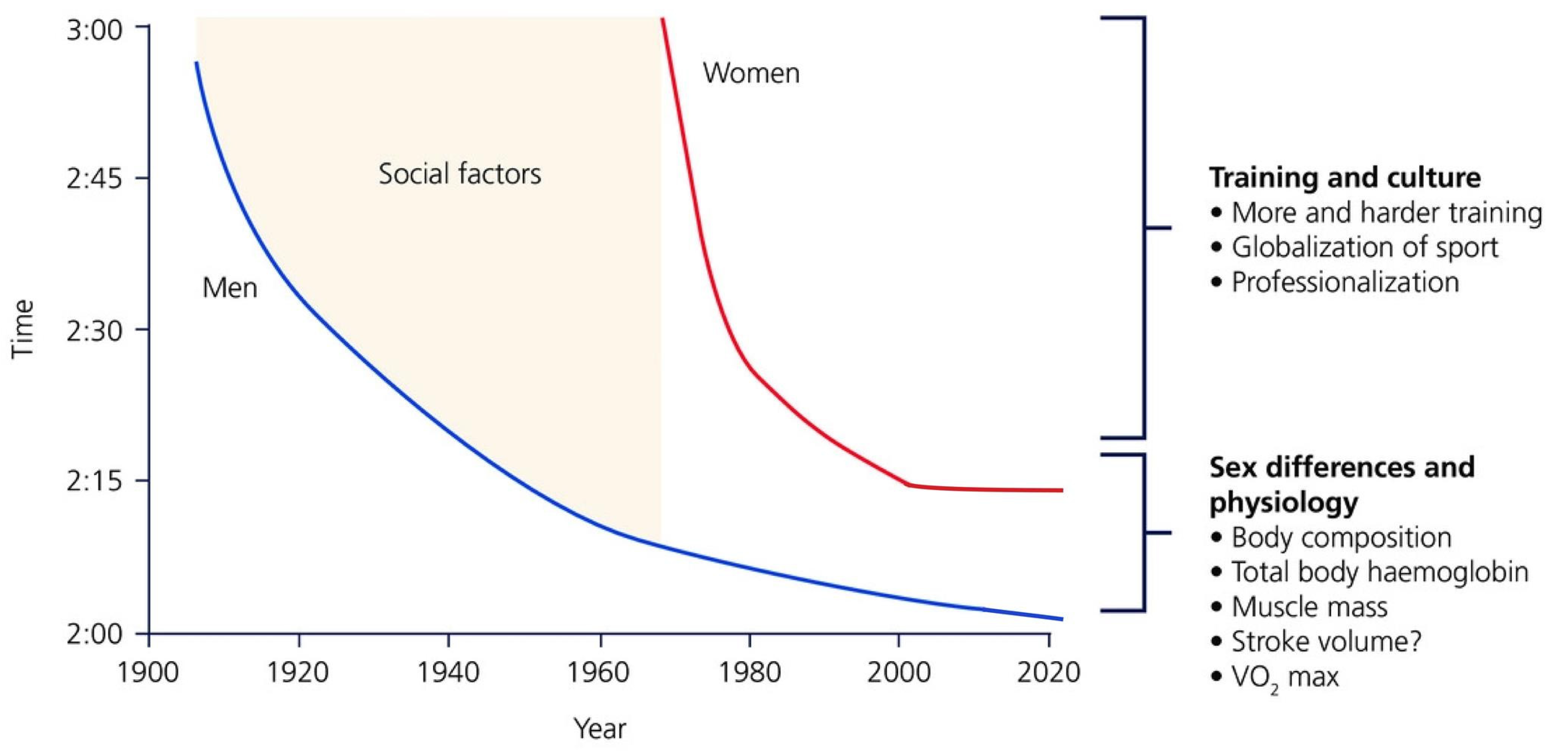
1. Introduction
1.1. Maximal Oxygen Consumption (VO2max)
1.2. Other Determinants of Endurance-Exercise Performance
1.2.1. Running Economy
1.2.2. Lactate Threshold
1.2.3. Critical Power
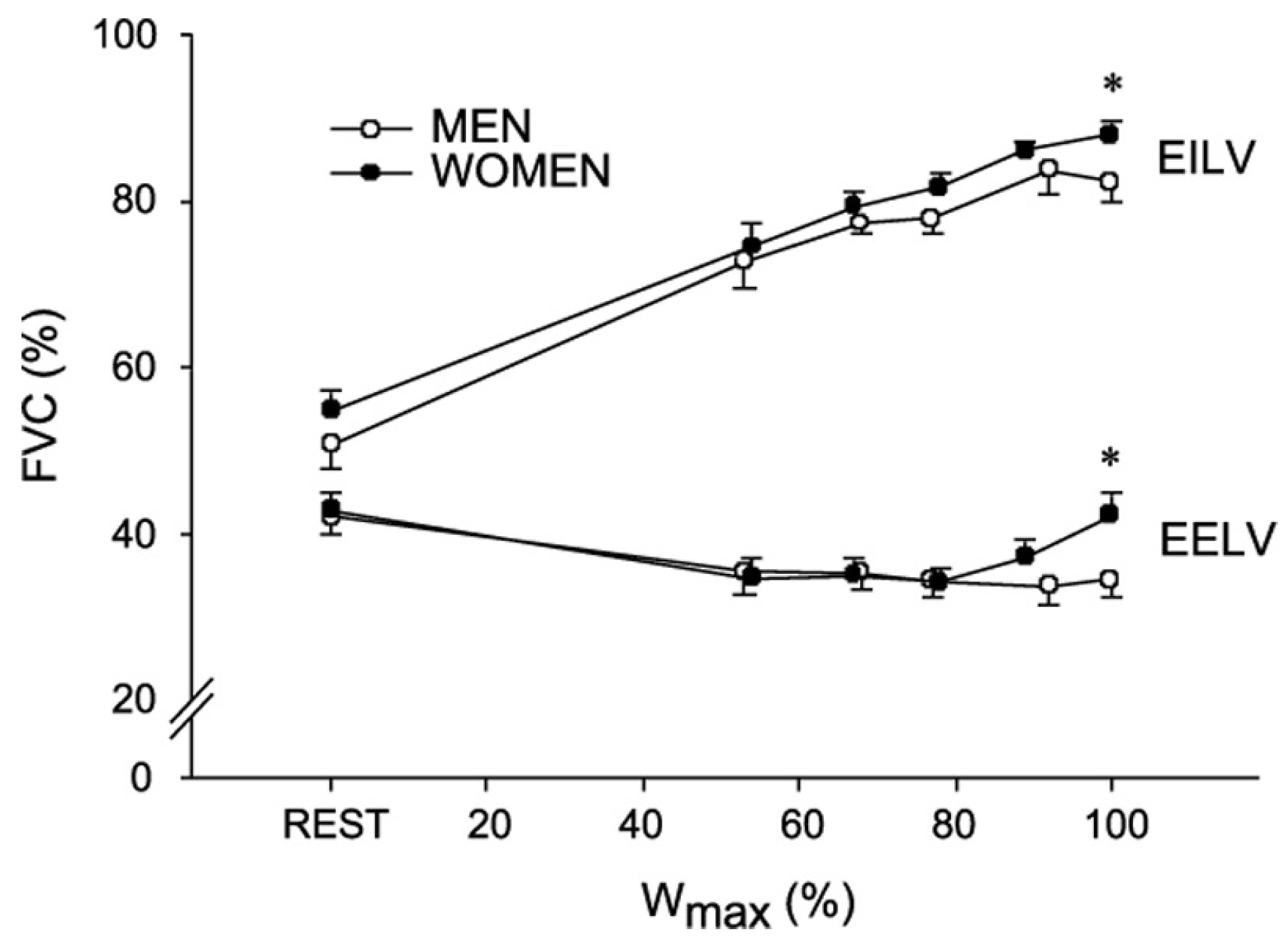
2. Pulmonary and Respiratory Considerations
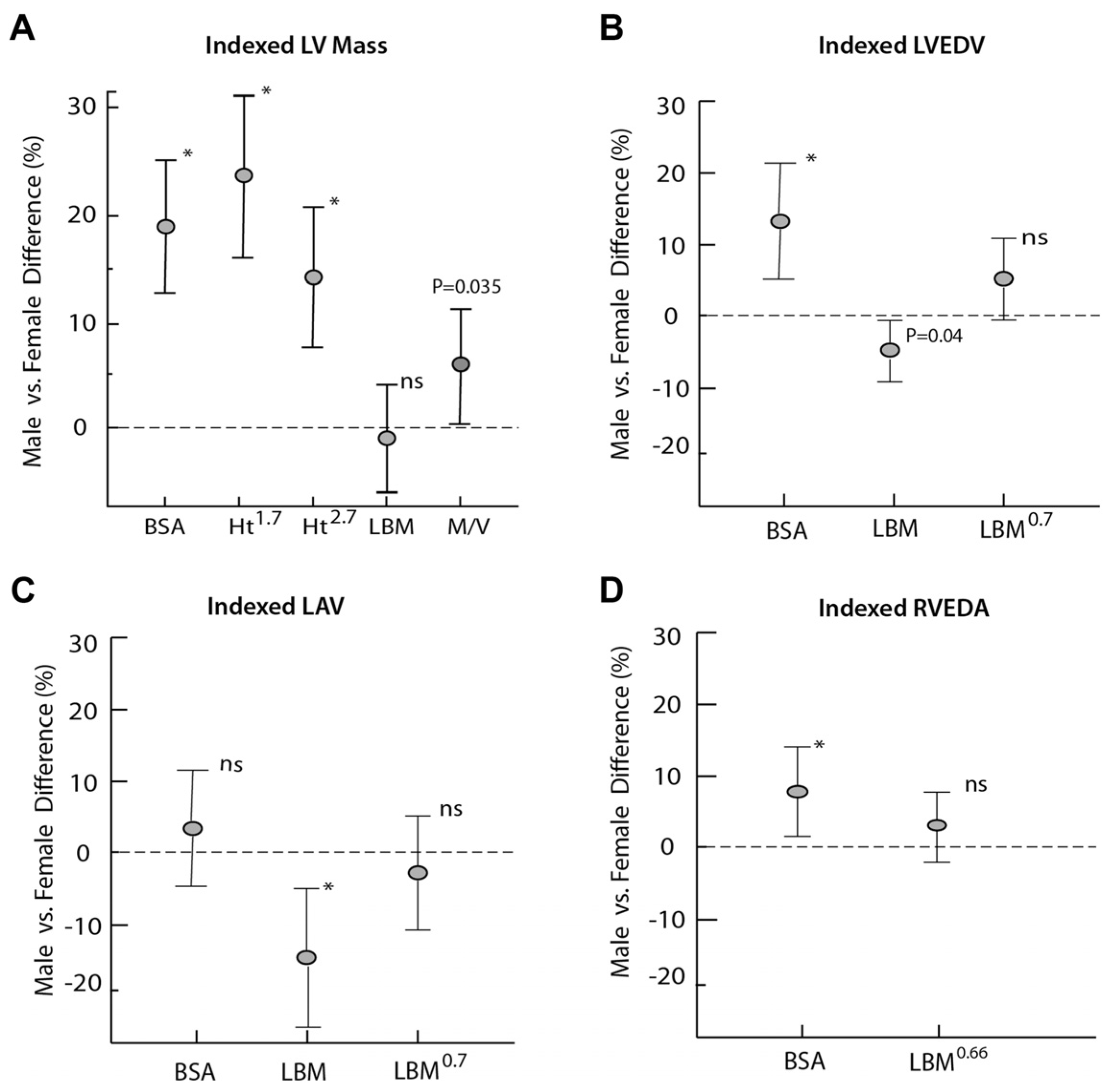
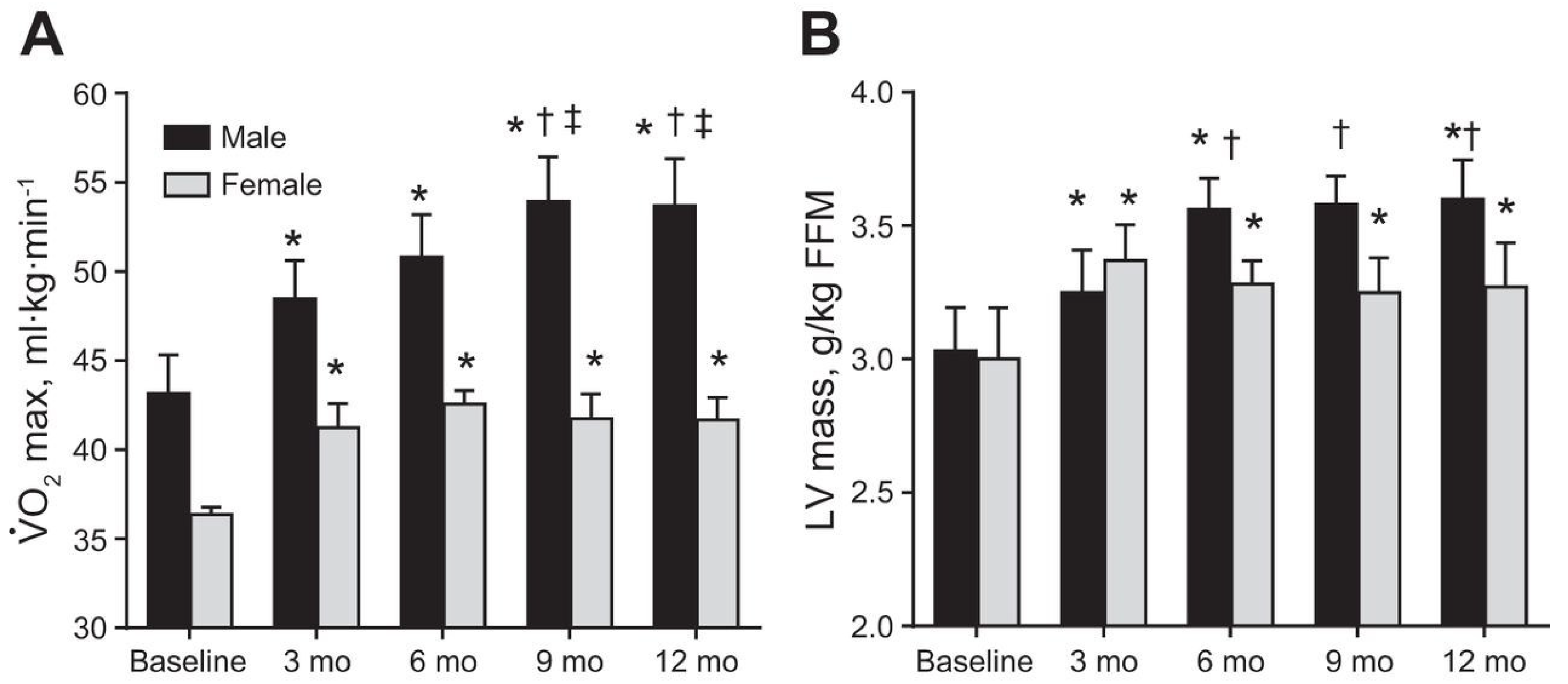
3. Cardiac Considerations
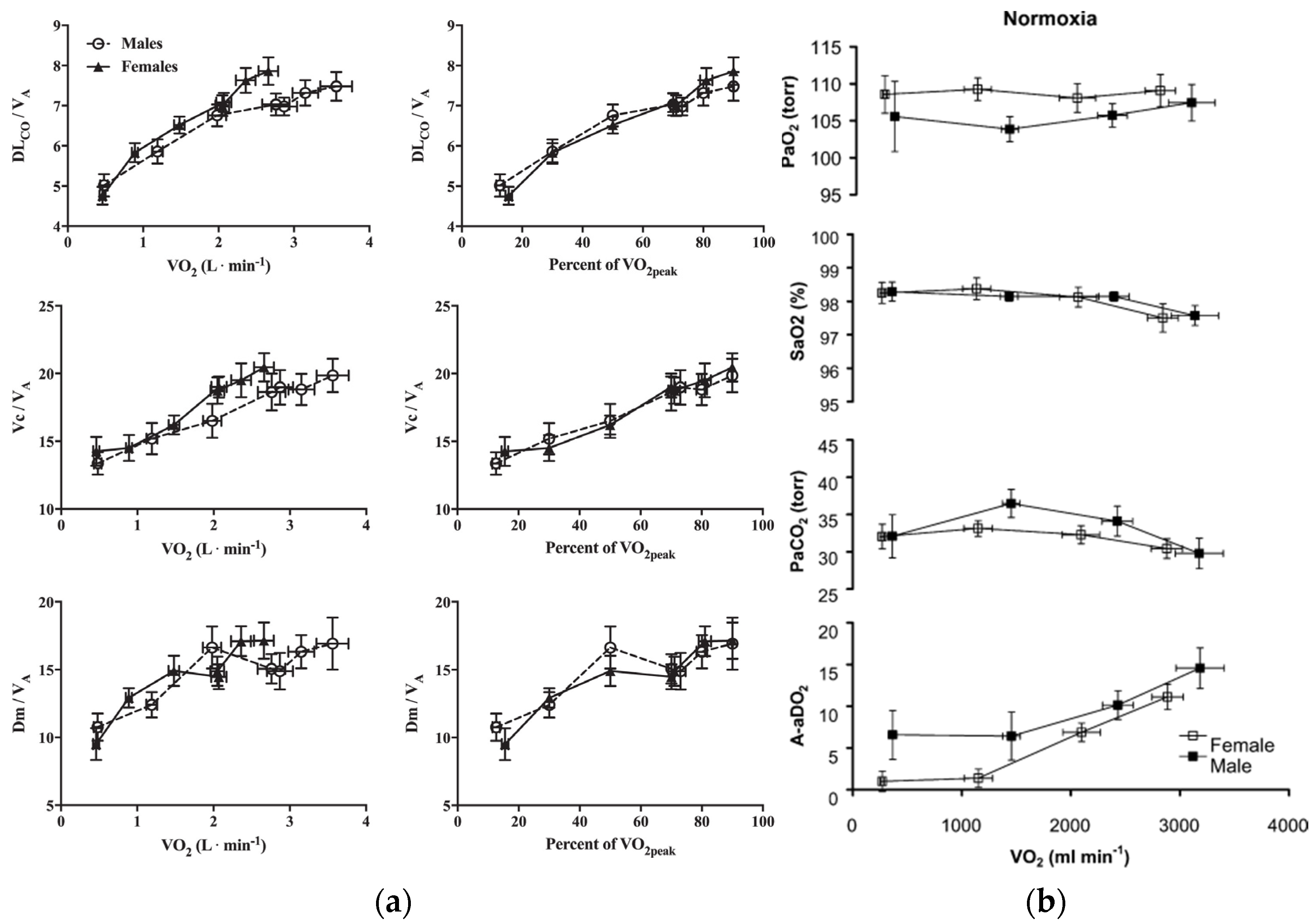
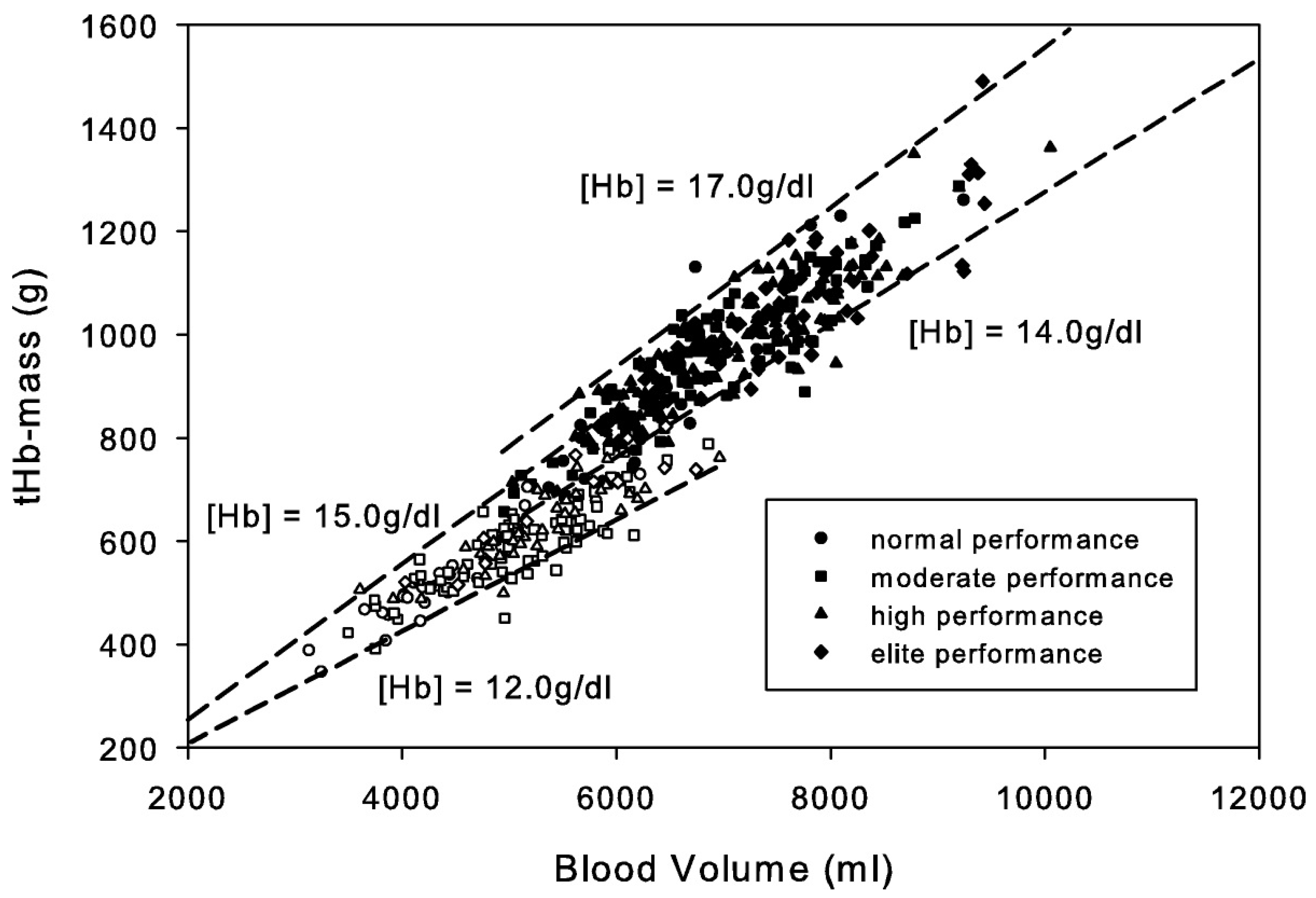
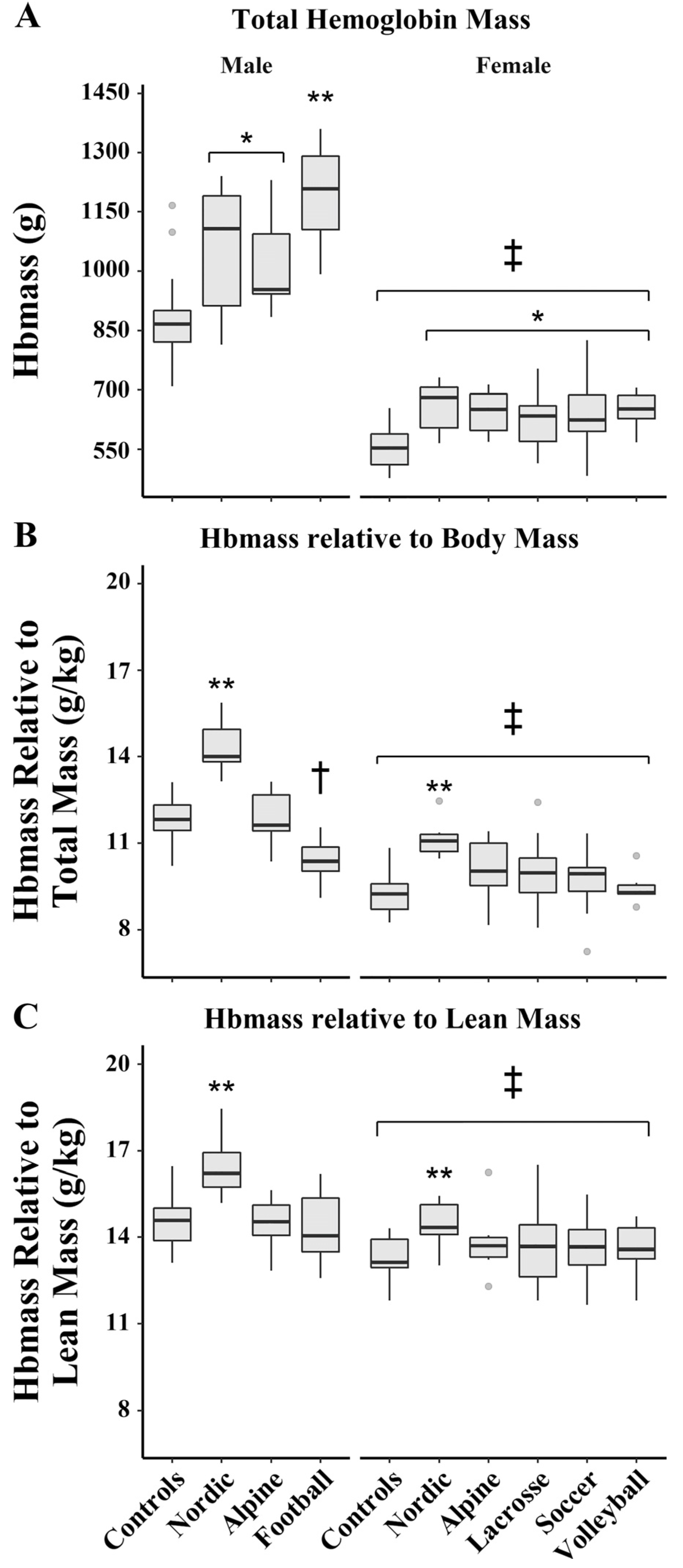
4. Hematological Considerations
5. Training Strategies—Heat and Hydration
6. Authors Closing Commentary
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Running USA Trends Report. Available online: https://www.wpr.org/sites/default/files/running_usa_trends_report_2019-r4.pdf (accessed on 31 July 2021).
- Cheuvront, S.N.; Carter, R.; Deruisseau, K.C.; Moffatt, R.J. Running performance differences between men and women: An update. Sports Med. 2005, 35, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Joyner, M.J. Physiological limits to endurance exercise performance: Influence of sex. J. Physiol. 2017, 595, 2949–2954. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.R., Jr. Scientific contributions of A. V. Hill: Exercise physiology pioneer. J. Appl. Physiol. 2002, 93, 1567–1582. [Google Scholar] [CrossRef] [PubMed]
- Costello, J.; Bieuzen, F.; Bleakley, C. Where are all the female participants in Sports and Exercise Medicine research? Eur. J. Sport Sci. 2014, 14, 847–851. [Google Scholar] [CrossRef]
- Sims, S.T.; Heather, A.K. Myths and Methodologies: Reducing scientific design ambiguity in studies comparing sexes and/or menstrual cycle phases. Exp. Physiol. 2017, 103, 1309–1317. [Google Scholar] [CrossRef]
- Bruinvels, G.; Burden, R.J.; McGregor, A.J.; Ackerman, K.E.; Dooley, M.; Richards, T.; Pedlar, C. Sport, exercise and the menstrual cycle: Where is the research? Br. J. Sports Med. 2017, 51, 487–488. [Google Scholar] [CrossRef]
- Liu, K.A.; Mager, N.A. Women’s involvement in clinical trials: Historical perspective and future implications. Pharm. Pract. 2016, 14, 708. [Google Scholar] [CrossRef]
- Coyle, E.F. Physiological determinants of endurance exercise performance. J. Sci. Med. Sport 1999, 2, 181–189. [Google Scholar] [CrossRef]
- Joyner, M.J.; Dominelli, P.B. Central cardiovascular system limits to aerobic capacity. Exp. Physiol. 2020, 106, 2299–2303. [Google Scholar] [CrossRef]
- Jones, A. Sports Science & Coaching The Physiology of the World Record Holder for the Women’s Marathon. Int. J. Sports Sci. Coach. 2006, 1, 101–116. [Google Scholar]
- Bassett, D.R.; Howley, E.T. Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med. Sci. Sports Exerc. 2000, 32, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Farrell, P.A.; Wilmore, J.H.; Coyle, E.F.; Billing, J.E.; Costill, D.L. Plasma lactate accumulation and distance running performance. Med. Sci. Sports 1979, 11, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Lundby, C.; Robach, P. Performance enhancement: What are the physiological limits? Physiology 2015, 30, 282–292. [Google Scholar] [CrossRef]
- VMendonca, G.; Matos, P.; Correia, J.M. Running economy in recreational male and female runners with similar levels of cardiovascular fitness. J. Appl. Physiol. 2020, 29, 508–515. [Google Scholar] [CrossRef]
- Støa, E.M.; Helgerud, J.; Rønnestad, B.R.; Hansen, J.; Ellefsen, S.; Støren, Ø. Factors Influencing Running Velocity at Lactate Threshold in Male and Female Runners at Different Levels of Performance. Front. Physiol. 2020, 11, 585267. [Google Scholar] [CrossRef]
- Pilegaard, H.; Bangsbo, J.; Richter, E.A.; Juel, C. Lactate transport studied in sarcolemmal giant vesicles from human muscle biopsies: Relation to training status. J. Appl. Physiol. 1994, 77, 1858–1862. [Google Scholar] [CrossRef]
- Poole, D.C.; Burnley, M.; Vanhatalo, A.; Rossiter, H.B.; Jones, A.M. Critical Power: An Important Fatigue Threshold in Exercise Physiology. Med. Sci. Sports Exerc. 2016, 48, 2320–2334. [Google Scholar] [CrossRef]
- Clark, I.E.; Vanhatalo, A.; Bailey, S.J.; Wylie, L.J.; Kirby, B.S.; Wilkins, B.W.; Jones, A.M. Effects of Two Hours of Heavy-Intensity Exercise on the Power-Duration Relationship. Med. Sci. Sports Exerc. 2018, 50, 1658–1668. [Google Scholar] [CrossRef]
- Black, M.I.; Jones, A.M.; Blackwell, J.R.; Bailey, S.J.; Wylie, L.J.; McDonagh, S.T.; Thompson, C.; Kelly, J.; Sumners, P.; Mileva, K.N. Muscle metabolic and neuromuscular determinants of fatigue during cycling in different exercise intensity domains. J. Appl. Physiol. 2017, 122, 446–459. [Google Scholar] [CrossRef]
- Burnley, M.; Vanhatalo, A.; Jones, A.M. Distinct profiles of neuromuscular fatigue during muscle contractions below and above the critical torque in humans. J. Appl. Physiol. 2012, 113, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Wilkerson, D.P.; DiMenna, F.; Fulford, J.; Poole, D.C. Musclemetabolic responses to exercise above and below the ‘‘critical power’’ assessed using 31P-MRS. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R585–R593. [Google Scholar] [CrossRef] [PubMed]
- Vanhatalo, A.; Black, M.I.; DiMenna, F.J.; Blackwell, J.R.; Schmidt, J.F.; Thompson, C.; Wylie, L.J.; Mohr, M.; Bangsbo, J.; Krustrup, P.; et al. The mechanistic bases of the power–time relationship: Muscle metabolic responses and relationships to muscle fibre type. J. Physiol. 2016, 594, 4407–4423. [Google Scholar] [CrossRef] [PubMed]
- Ansdell, P.; Škarabot, J.; Atkinson, E.; Corden, S.; Tygart, A.; Hicks, K.M.; Thomas, K.; Hunter, S.K.; Howatson, G.; Goodall, S. Sex differences in fatigability following exercise normalised to the power-duration relationship. J. Physiol. 2020, 598, 5717–5737. [Google Scholar] [CrossRef]
- Ansdell, P.; Thomas, K.; Hicks, K.M.; Hunter, S.K.; Howatson, G.; Goodall, S. Physiological sex differences affect the integrative response exercise: Acute and chronic implications. Exp. Physiol. 2020, 105, 2007–2021. [Google Scholar] [CrossRef]
- Sheel, A.W.; Richards, J.C.; Foster, G.E.; Guenette, J.A. Sex Differences in Respiratory Exercise Physiology. Sports Med. 2004, 34, 567–579. [Google Scholar] [CrossRef]
- Amann, M. Pulmonary system limitations to endurance exercise performance in humans. Exp. Physiol. 2009, 97, 311–318. [Google Scholar] [CrossRef]
- McKenzie, D.C. Respiratory physiology: Adaptations to high-level exercise. Br. J. Sports Med. 2012, 46, 381–384. [Google Scholar] [CrossRef]
- Dempsey, J.A.; Hanson, P.G.; Henderson, K.S. Exercise-induced arterial hypoxaemia in healthy human subjects at sea level. J. Physiol. 1984, 355, 161–175. [Google Scholar] [CrossRef]
- Harms, C.A.; Babcock, M.A.; McClaran, S.R.; Pegelow, D.F.; Nickele, G.A.; Nelson, W.B.; Dempsey, J.A. Respiratory muscle work compromises leg blood flow during maximal exercise. J. Appl. Physiol. 1997, 82, 1573–1583. [Google Scholar] [CrossRef]
- Powers, S.K.; Dodd, S.; Lawler, J.; Landry, G.; Kirtley, M.; McKnight, T.; Grinton, S. Incidence of exercise induced hypoxemia in elite endurance athletes at sea level. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 58, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.; Powers, S.K.; Stuart, M.K. Hemoglobin desaturation in highly trained athletes during heavy exercise. Med. Sci. Sports Exerc. 1986, 18, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Harms, C.S.; McClaran, S.; Nickele, G.A.; Pegelow, D.F.; Nelson, W.B.; Dempsey, J.A. Effect of exercise induced arterial O2 desaturation on VO2 max in women. Exerc. Sci. Med. Sport 2000, 32, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Lawler, J.; Dempsey, J.A.; Dodd, S.; Landry, G. Effects of incomplete pulmonary gas exchange on VO2 max. J. Appl. Physiol. 1989, 66, 2491–2495. [Google Scholar] [CrossRef] [PubMed]
- Squires, R.W.; Buskirk, E.R. Aerobic capacity during acute exposure to simulated altitude, 914 to 2286 meters. Med. Sci. Sports Exerc. 1982, 14, 36–40. [Google Scholar] [CrossRef]
- Ceridon, M.L.; Beck, K.C.; Olson, T.P.; Bilezikian, J.A.; Johnson, B.D. Calculating alveolar capillary conductance and pulmonary capillary blood volume: Comparing the multiple-and single-inspired oxygen tension methods. J. Appl. Physiol. 2010, 109, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Dominelli, P.B.; Ripoll, J.G.; Cross, T.J.; Baker, S.E.; Wiggins, C.C.; Welch, B.T.; Joyner, M.J. Sex differences in large conducting airway anatomy. J. Appl. Physiol. 2018, 125, 960–965. [Google Scholar] [CrossRef]
- Sheel, A.W.; Dominelli, P.B.; Molgat-Seon, Y. Revisiting dysanapsis: Sex-based differences in airways and the mechanics of breathing during exercise. Exp. Physiol. 2016, 101, 213–218. [Google Scholar] [CrossRef]
- Hopkins, S.R.; Harms, C.A. Gender and pulmonary gas exchange during exercise. Exerc. Sport Sci. Rev. 2004, 32, 50–56. [Google Scholar] [CrossRef]
- Guenette, J.A.; Witt, J.D.; McKenzie, D.C.; Road, J.D.; Sheel, A.W. Respiratory mechanics during exercise in endurance-trained men and women. J. Physiol. 2007, 581, 1309–1322. [Google Scholar] [CrossRef]
- Guenette, J.A.; Romer, L.M.; Querido, J.S.; Chua, R.; Eves, N.D.; Road, J.D.; Mckenzie, D.C.; Sheel, A.W. Sex differences in exercise-induced diaphragmatic fatigue in endurance-trained athletes. J. Appl. Physiol. 2010, 109, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Wanke, T.; Formanek, D.; Schenz, G.; Popp, W.; Gatol, H.; Zwick, H. Mechanical load on the ventilatory muscles during an incremental cycle ergometer test. Eur. Respir. J. 1991, 4, 385–392. [Google Scholar] [PubMed]
- Sheel, W.A.; Guenette, J.A. Mechanics of Breathing during Exercise in Men and Women. Exerc. Sport Sci. Rev. 2008, 36, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, S.S.; Dominelli, P.B.; Sporer, B.C.; Koehle, M.S.; Sheel, A.W. Heliox breathing equally influences respiratory mechanics and cycling performance in trained males and females. J. Appl. Physiol. 2015, 118, 255–264. [Google Scholar] [CrossRef]
- Hicks, A.L.; Kent-Braun, J.; Ditor, D.S. Sex Differences in Human Skeletal Muscle Fatigue. Exerc. Sport Sci. Rev. 2001, 29, 109–112. [Google Scholar] [CrossRef]
- Smith, J.R.; Broxterman, R.M.; Hammer, S.M.; Alexander, A.M.; Didier, K.D.; Kurti, S.P.; Barstow, T.J.; Harms, C.A. Sex differences in the cardiovascular consequences of the inspiratory muscle metaboreflex. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, 574–581. [Google Scholar] [CrossRef]
- Welch, J.F.; Archiza, B.; Guenette, J.A.; West, C.R.; Sheel, A.W. Sex differences in diaphragmatic fatigue: The cardiovascular response to inspiratory resistance. J. Physiol. 2018, 596, 4017–4032. [Google Scholar] [CrossRef]
- Bouwsema, M.M.; Tedjasaputra, V.; Stickland, M.K. Are there sex differences in the capillary blood volume and diffusing capacity response to exercise? J. Appl. Physiol. 2017, 122, 460–469. [Google Scholar] [CrossRef]
- Olfert, I.M.; Balouch, J.; Kleinsasser, A.; Knapp, A.; Wagner, H.; Wagner, P.D.; Hopkins, S.R. Does gender affect human pulmonary gas exchange during exercise? J. Physiol. 2004, 557, 529–541. [Google Scholar] [CrossRef]
- Di Bello, V.; Santoro, G.; Talarico, L.; Di Muro, C.; Caputo, M.T.; Giorgi, D.; Bertini, A.; Bianchi, M.; Giusti, C. Left ventricular function during exercise in athletes and in sedentary men. Med. Sci. Sports Exerc. 1996, 28, 190–196. [Google Scholar] [CrossRef]
- Gledhill, N.; Cox, D.; Jamnik, R. Endurance athletes’ stroke volume does not plateau: Major advantage is diastolic function. Med. Sci. Sports Exerc. 1994, 26, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Conlee, R.K.; Jensen, R.; Fellingham, G.W.; George, J.D.; Fisher, A.G. Stroke volume does not plateau during graded exercise in elite male distance runners. Med. Sci. Sports Exerc. 2001, 33, 1849–1854. [Google Scholar] [CrossRef] [PubMed]
- Stickland, M.K.; Welsh, R.C.; Petersen, S.R.; Tyberg, J.V.; Anderson, W.D.; Jones, R.L.; Taylor, D.A.; Bouffard, M.; Haykowsky, M.J.; Stickland, M.K.; et al. Does fitness level modulate the cardiovascular hemodynamic response to exercise? J. Appl. Physiol. 2006, 100, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Foryst-Ludwig, A.; Kintscher, U. Sex differences in exercise-induced cardiac hypertrophy. Pflug. Arch. Eur. J. Physiol. 2013, 465, 731–737. [Google Scholar] [CrossRef]
- Bernardo, B.C.; Weeks, K.L.; Pretorius, L.; McMullen, J.R. Molecular distinction between physiological and pathological cardiac hypertrophy: Experimental findings and therapeutic strategies. Pharmacol. Ther. 2010, 128, 191–227. [Google Scholar] [CrossRef]
- Heineke, J.; Molkentin, J.D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nat. Rev. Mol. Cell Biol. 2006, 7, 589–600. [Google Scholar] [CrossRef]
- Bassareo, P.P.; Crisafulli, A. Gender Differences in Hemodynamic Regulation and Cardiovascular Adaptations to Dynamic Exercise. Curr. Cardiol. Rev. 2019, 16, 65–72. [Google Scholar] [CrossRef]
- Levine, B.D.; Lane, L.D.; Buckey, J.C.; Friedman, D.B.; Blomqvist, C.G. Left ventricular pressure-volume and Frank-Starling relations in endurance athletes. Implications for orthostatic tolerance and exercise performance. Circulation 2019, 84, 1016–1023. [Google Scholar] [CrossRef]
- Pelliccia, A.; Maron, B.J.; Spataro, A.; Proschan, M.A.; Spirito, P. The upper limit of physiologic cardiac hypertrophy inhighly trained elite athletes. N. Engl. J. Med. 1992, 324, 295–301. [Google Scholar] [CrossRef]
- Whyte, G.P.; George, K.; Sharma, S.; Firoozi, S.; Stephens, N.; Senior, R.; McKenna, W.J. The upper limit of physiological cardiac hypertrophy in elite male and female athletes: The British experience. Eur. J. Appl. Physiol. 2004, 92, 592–597. [Google Scholar] [CrossRef]
- Giraldeau, G.; Kobayashi, Y.; Finocchiaro, G.; Wheeler, M.; Perez, M.; Kuznetsova, T.; Lord, R.; George, K.P.; Oxborough, D.; Schnittger, I.; et al. Gender Differences in Ventricular Remodeling and Function in College Athletes, Insights from Lean Body Mass Scaling and Deformation Imaging. Am. J. Cardiol. 2015, 116, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Pressler, A.; Haller, B.; Scherr, J.; Heitkamp, D.; Esefeld, K.; Boscheri, A.; Wolfarth, B.; Halle, M. Association of body composition and left ventricular dimensions in elite athletes. Eur. J. Prev. Cardiol. 2012, 19, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Howden, E.J.; Perhonen, M.; Peshock, R.M.; Zhang, R.; Arbab-Zadeh, A.; Adams-Huet, B.; Levine, B.D. Females have a blunted cardiovascular response to one year of intensive supervised endurance training. J. Appl. Physiol. 2015, 119, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Loucks, A.B. Low Energy Availability in the Marathon and Other Endurance Sports. Sports Med. 2007, 37, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Canestro, C.; Montero, D. Sex Dimorphism of VO2max Trainability: A Systematic Review and Meta-analysis. Sports Med. 2019, 49, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Green, H.J.; Carter, S.; Grant, S.; Tupling, R.; Coates, G.; Ali, M. Vascular volumes and hematology in male and female runners and cyclists. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 79, 244–250. [Google Scholar] [CrossRef]
- Kanstrup, I.L.; Ekblom, B. Blood volume and hemoglobin concentrations as determinants of maximal aerobic power. Med. Sci. Sports Exerc. 1984, 16, 256–262. [Google Scholar] [CrossRef]
- Schmidt, W.; Prommer, N. Impact of alterations in total hemoglobin mass on VO2 max. Exerc. Sport Sci. Rev. 2010, 38, 68–75. [Google Scholar] [CrossRef]
- Schmidt, W.; Prommer, N. Effects of various training modalities on blood volume. Scand. J. Med. Sci. Sports 2008, 18, 57–69. [Google Scholar] [CrossRef]
- Ulrich, G.; Bärtsch, P.; Friedmann-Bette, B. Total haemoglobin mass and red blood cell profile in endurance-trained and non endurance-trained adolescent athletes. Eur. J. Appl. Physiol. 2011, 111, 2855–2864. [Google Scholar] [CrossRef]
- Heinicke, K.; Wolfarth, B.; Winchenbach, P.; Biermann, B.; Schmid, A.; Huber, G.; Friedmann, B.; Schmidt, W. Blood volume and hemoglobin mass in elite athletes of different disciplines. Int. J. Sports Med. 2001, 22, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Prommer, N.; Sottas, P.E.; Schoch, C.; Schumacher, Y.O.; Schmidt, W. Total hemoglobin mass--a new parameter to detect blood doping? Med. Sci. Sports Exerc. 2008, 40, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Murphy, W.G. The sex difference in haemoglobin levels in adults—Mechanisms, causes, and consequences. Blood Rev. 2014, 28, 41–47. [Google Scholar] [CrossRef]
- Jelkmann, W. Regulation of erythropoietin production. J. Physiol. 2011, 589, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Shahani, S.; Braga-Basaria, M.; Maggio, M.; Basaria, S. Androgens and erythropoiesis:past and present. J. Endocrinol. Invest. 2009, 32, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Handelsman, D.J.; Hirschberg, A.L.; Bermon, S. Circulating testosterone as the hormonal basis of sex differences in athletic performance. Endocr. Rev. 2018, 39, 803–829. [Google Scholar] [CrossRef]
- Rickenlund, A.; Carlström, K.; Ekblom, B.; Brismar, T.B.; von Schoultz, B.; Hirschberg, A.L. Hyperandrogenicity is an alternative mechanism underlying oligomenorrhea or amenorrhea in female athletes and may improve physical performance. Fertil. Steril. 2003, 79, 947–955. [Google Scholar] [CrossRef]
- Hirschberg, A.L. Female hyperandrogenism and elite sport. Endocr. Connect. 2020, 9, R81–R92. [Google Scholar] [CrossRef]
- Javed, A.; Kashyap, R.; Lteif, A.N. Hyperandrogenism in female athletes with functional hypothalamic amenorrhea: A distinct phenotype. Int. J. Women’s Health 2015, 7, 103–111. [Google Scholar] [CrossRef][Green Version]
- Hagmar, M.; Berglund, B.; Brismar, K.; Hirschberg, A.L. Hyperandrogenism may explain reproductive dysfunction in female Olympic athletes. Med. Sci. Sports Exerc. 2009, 41, 1241–1248. [Google Scholar] [CrossRef]
- Karunasena, N.; Han, T.S.; Mallappa, A.; Elman, M.; Merke, D.P.; Ross RJ, M.; Daniel, E. Androgens correlate with increased erythropoiesis in women with congenital adrenal hyperplasia. Clin. Endocrinol. 2017, 86, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Bock, A.V. The constancy of the volume of the blood plasma. Arch. Intern. Med 1921, 27, 83–101. [Google Scholar] [CrossRef]
- Gersh, I.; Still, M.A. Blood vessels in fat tissue. Relation to problems of gas exchange. J. Exp. Med. 1945, 81, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.G.; Evans, W.A. Clinical studies of the blood volume. II. The relation of plasma and total blood volume to venous pressure, blood velocity rate, physical measurements, age and sex in ninety normal humans. J. Clin. Investig. 1937, 16, 317–328. [Google Scholar] [CrossRef]
- Goodrich, X.A.; Frisco, D.J.; Kim, S.; Holliday, M.; Rueda, M.; Poddar, S.; Byrnes, W.C. The importance of lean mass and iron deficiency when comparing hemoglobin mass in male and female athletic groups. J. Appl. Physiol. 2020, 129, 855–863. [Google Scholar] [CrossRef]
- Rushton, D.H.; Barth, J.H. What is the evidence for gender differences in ferritin and haemoglobin? Crit. Rev. Oncol./Hematol. 2010, 73, 1–9. [Google Scholar] [CrossRef]
- Valberg, L.S.; Sorbie, J.; Ludwig, J.; Pelletier, O. Serum ferritin and the iron status of canadians. Can. Med. Acc. J. 1976, 114, 417–421. [Google Scholar]
- Hallberg, L. Iron absorption and iron deficiency. Human nutrition. Clin. Nutr. 1982, 36C, 250–278. [Google Scholar]
- Ledingham, I.M. Factors influencing oxygen availability. J. Clin. Pathol. Suppl. 1977, 11, 1–6. [Google Scholar] [CrossRef]
- Mairbäurl, H. Red blood cells in sports: Effects of exercise and training on oxygen supply by red blood cells. Front. Physiol. 2013, 4, 332. [Google Scholar] [CrossRef]
- Hinton, P.S. Iron and the endurance athlete. Appl. Physiol. Nutr. Metab. 2014, 39, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Yanovich, R.; Ketko, I.; Charkoudian, N. Sex Differences in Human Thermoregulation: Relevance for 2020 and Beyond. Physiology 2020, 35, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Giersch, G.; Morrissey, M.C.; Katch, R.K.; Colburn, A.T.; Sims, S.T.; Stachenfeld, N.S.; Casa, D.J. Menstrual cycle and thermoregulation during exercise in the heat: A systematic review and meta-analysis. J. Sci. Med. Sport 2020, 23, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Giersch, G.; Charkoudian, N.; Stearns, R.L.; Casa, D.J. Fluid Balance and Hydration Considerations for Women: Review and Future Directions. Sports Med. 2020, 50, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, D.; Kenny, G.P. Does sex have an independent effect on thermoeffector responses during exercise in the heat? J. Physiol. 2012, 590, 5963–5973. [Google Scholar] [CrossRef]
- Gagnon, D.; Crandall, C.G.; Kenny, G.P. Sex differences in postsynaptic sweating and cutaneous vasodilation. J. Appl. Physiol. 1985, 114, 394–401. [Google Scholar] [CrossRef]
- Ali, Y.S.; Daamen, N.; Jacob, G.; Jordan, J.; Shannon, J.R.; Biaggioni, I.; Robertson, D. Orthostatic intolerance: A disorder of young women. Obstet. Gynecol. Surv. 2000, 55, 251–259. [Google Scholar] [CrossRef]
- Hardy, J.D.; Du Bois, E.F. Differences between men and women in their response to heat and cold. Proc. Natl. Acad. Sci. USA 1940, 26, 389–398. [Google Scholar] [CrossRef]
- Hart, E.C.; Charkoudian, N. Sympathetic neural mechanisms in human blood pressure regulation. Curr. Hypertens. Rep. 2011, 13, 237–243. [Google Scholar] [CrossRef]
- Hart, E.C.; Charkoudian, N. Sympathetic neural regulation of blood pressure: Influences of sex and aging. Physiology 2014, 2, 8–15. [Google Scholar] [CrossRef]
- Hart, E.C.; Charkoudian, N.; Wallin, B.G.; Curry, T.B.; Eisenach, J.; Joyner, M.J. Sex and ageing differences in resting arterial pressure regulation:the role of the -adrenergic receptors. J. Physiol. 2011, 589, 5285–5297. [Google Scholar] [CrossRef] [PubMed]
- Kirby, N.V.; Lucas, S.J.E.; Lucas, R.A.I. Nine-, but not four-days heat acclimation improves self-paced endurance performance in females. Front. Physiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Mee, J.A.; Gibson, O.R.; Doust, J.; Maxwell, N.S. A comparison of males and females’ temporal patterning to short- and long-term heat acclimation. Scand. J. Med. Sci. Sports 2015, 25, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.N.; Jay, O. Biophysical aspects of human thermoregulation during heat stress. Auton. Neurosci. 2016, 196, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Gonzalez-Alonso, J. Cardiovascular drift during prolonged exercise: New perspectives. Exerc. Sport Sci. Rev. 2001, 29, 88–92. [Google Scholar]
- Rowell, L.B. Human cardiovascular adjustments to exercise and thermal stress. Physiol. Rev. 1974, 54, 75–159. [Google Scholar] [CrossRef]
- Wickham, K.A.; McCarthy, D.G.; Spriet, L.L.; Cheung, S.S. Sex differences in the physiological responses to exercise-induced dehydration: Consequences and mechanisms. J. Appl. Physiol. 2021, 131, 504–510. [Google Scholar] [CrossRef]
- Logan-Sprenger, H.M.; Heigenhauser, G.J.F.; Killian, K.J.; Spriet, L.L. Effects of dehydration during cycling on skeletal muscle metabolism in females. Med. Sci. Sports Exerc. 2012, 44, 1949–1957. [Google Scholar] [CrossRef]
- Ali, A.; Gardiner, R.; Foskett, A.; Gant, N. Fluid balance, thermoregulation and sprint and passing skill performance in female soccer players. Scand. J. Med. Sci. Sports 2011, 21, 437–445. [Google Scholar] [CrossRef]
- Logan-Sprenger, H.M.; Heigenhauser, G.J.F.; Jones, G.L.; Spriet, L.L. Increase in skeletal muscle glycogenolysis and perceived exertion with progressive dehydration during cycling in hydrated men. Int. J. Sport. Nutr. Exerc. Metab. 2013, 23, 220–229. [Google Scholar] [CrossRef]
- Ali, A.; Williams, C. Isokinetic and isometric muscle function of the knee extensors and flexors during simulated soccer activity: Effect of exercise and dehydration. J. Sports Sci. 2013, 31, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Ritz, P.; Vol, S.; Berrut, G.; Tack, I.; Arnaud, M.J.; Tichet, J. Influence of gender and body composition on hydration and body water spaces. Clin. Nutr. 2008, 27, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Sims, S.; Yeager, S. Roar: How to Match Your Food and Fitness to Your Unique Female Physiology for Optimum Performance, Great Health, and a Strong, Lean Body for Life; Rodale, Inc.: New York, NY, USA, 2016. [Google Scholar]
- Emmonds, S.; Heyward, O.; Jones, B. The Challenge of Applying and Undertaking Research in Female Sport. Sports Med. 2019, 5, 51. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santisteban, K.J.; Lovering, A.T.; Halliwill, J.R.; Minson, C.T. Sex Differences in VO2max and the Impact on Endurance-Exercise Performance. Int. J. Environ. Res. Public Health 2022, 19, 4946. https://doi.org/10.3390/ijerph19094946
Santisteban KJ, Lovering AT, Halliwill JR, Minson CT. Sex Differences in VO2max and the Impact on Endurance-Exercise Performance. International Journal of Environmental Research and Public Health. 2022; 19(9):4946. https://doi.org/10.3390/ijerph19094946
Chicago/Turabian StyleSantisteban, Kelsey J., Andrew T. Lovering, John R. Halliwill, and Christopher T. Minson. 2022. "Sex Differences in VO2max and the Impact on Endurance-Exercise Performance" International Journal of Environmental Research and Public Health 19, no. 9: 4946. https://doi.org/10.3390/ijerph19094946
APA StyleSantisteban, K. J., Lovering, A. T., Halliwill, J. R., & Minson, C. T. (2022). Sex Differences in VO2max and the Impact on Endurance-Exercise Performance. International Journal of Environmental Research and Public Health, 19(9), 4946. https://doi.org/10.3390/ijerph19094946