Abstract
Although new inputs of acidic anions are decreasing, soil acidification still deserves more academic attention because of the effects of historical stores of SO42− already absorbed into soils. Forest canopy has large, species-specific effects on rainwater chemistry, for which the hydrological mechanism remains unclear. We investigated precipitation, throughfall, stemflow, and litter leachate across three forest types in a severely acid-polluted site located in Southwest China. Precipitation monitored over 4 months, representing summer, fall, winter, and spring, indicated neutral precipitation in Tieshanping with pH ranging from 6.58–7.33. Throughfall and litter leachate in Pinus massoniana Lamb. stands were enriched with greater cation and anion fluxes, as well as more dissolved organic carbon (DOC) flux. Rainwater from pure stands of Cinnamomum camphora (Linn) Presl yielded lower N and DOC inputs to soils with higher base saturation, which would reduce soil acidification and, therefore, improve the sustainability of forest ecosystems.
1. Introduction
Acid deposition is a global environmental problem that has garnered great interest due to its negative impacts on ecosystems [1,2]. Since the early 1980s, the government has made efforts to decrease acidic emissions, especially SO2 linked to acid deposition, yet nitrogen oxide emissions have been increasing with the rapid increase in the use of private cars for nearly two decades, so it still remains a severe issue in southern China [2,3,4]. Soil acidification caused by acid deposition has adverse effects on ecosystems, particularly forest ecosystems [5]. Numerous studies report the damage of acid deposition on forests, including decreased growth and dieback [6], changing biogeochemical cycles [7], altered aboveground and belowground biomes [8,9,10], and impacts on ecosystem function and sustainability [11,12]. Additionally, despite reductions of acidic anion inputs [13,14], the recovery of acidified soils could be delayed due to historic adsorption of SO42− into soils [1]. Soil properties, including soil acidity, are highly affected by forest types [15,16].
Actually, the process of the effects of forest canopies on soil nutrients is very complex. On the one hand, forest covers can alter microclimatic conditions [17], including the surface albedo, temperature, wind and humidity, which may affect decomposition rates [18], soil properties, and further affect plant growth as a feedback loop [17]. The effects of forests on the surface albedo and temperature are also complicated and result in various effects in different latitudes with tropical, temperate or boreal forests [19,20]. Additionally, forest canopies change precipitation partitioning that affects the nutrient cycling in the forest ecosystem: interception, throughfall, and stemflow [21]. Canopy partitioning can impact the dynamics of tree water and nutrient balances [22]. The minerals dissolved in interception can directly enter the plant through foliar and bark uptake, and also can reach forest soil via throughfall or stemflow. Nutrients can also be leached from throughfall and stemflow and input into soils [22]. Interception as well as throughfall and stemflow are influenced by many features of the forest type, such as density, branch angle, the uniformity, bark characteristics, leaf shape and leaf area index [23], which are very complicated in forest ecosystems. Further, the chemical enrichment of throughfall and stemflow and nutrient leaching capacity exhibit considerable species-specific variability [24], which further complicates general conclusions on the effects of forest types on water and soil properties, especially under the background of climate change, because forests interact with climate change via albedo, evaporation, the carbon cycle and other factors [25,26].
Inputs of acidic anions and other nutrients to the forest floor through precipitation are affected by dry deposition on the canopy and the canopy itself, both of which can alter water chemistry and further affect soil quality. Throughfall and stemflow supply available nutrients to the forest floor, which can affect soil acidity [27], although the volume of stemflow is much less than that of throughfall and often ignored [28]. Before nutrient-carrying throughfall and stemflow enter into the soil, they pass through the layer of forest floor litter (except in areas of bare ground), where nutrients from throughfall, stemflow, and litter combine to form litter leachate, which ultimately affects soil quality. A study in British Columbia confirmed that throughfall and stemflow contribute to higher pH, N availability, and total exchangeable bases in forest floor areas associated with bigleaf maple, which have legacy effects on soil fertility [29]. Forest floor leachate chemistry largely determines biogeochemical processes in the soil and soil solution chemistry, affecting soil acidity and the sustainability of the forest ecosystem [30]. It is necessary and important to account for forest floor leachate in addition to throughfall and stemflow when studying the hydrological effects of rainfall on soil in forested ecosystems.
The exchange capacity of basic cations and acid anions in forest canopy and litter have strong species specificity [24,27,31,32]. Individual tree species play an important role in the distribution and cycling of nutrients that influence ecosystem function, biodiversity, resistance to stress, and sustainability [33,34]. Previous studies have reported that different forest types can strongly affect the chemical composition of throughfall, stemflow, and floor leachate, particularly the effect of nitrate and sulfate on anionic charges [24,27,31,32,35]. It was reported that the canopy of conifers intercepted particles and cloud droplets more efficiently than deciduous species, resulting in generally higher dry deposition under conifers [36,37]. Deciduous forests receive less S and N than coniferous stands via throughfall plus stemflow on the forest floor as reviewed by De Schrijver et al. [31]. This is attributable to the higher dry deposition capacity of conifers, which were also indicated to be more vulnerable to acidification with higher seepage of NO3−, SO42−, and cations compared to deciduous stands [38,39]. Previous studies revealed higher DOC and H+ fluxes and lower pH and exchangeable cations in forest floor leachate from conifers compared to broadleaved stands [27,30,40]. Thus, while throughfall, stemflow, and forest floor leachates may play an important role in influencing acidity and nutrients of soils, their importance has been rarely studied [32].
Tieshanping, located in the southwest of China, is one of the most severely acid-polluted sites in China. Stands of Pinus massoniana (Pi) that naturally regenerated between 1958 and 1962 showed decreased growth attributed to acid deposition. A native deciduous species, Cinnamomum camphora (Linn) Presl (Ci), was planted between the 1980s and 1990s to improve forest health. A pure plantation of Ci was established with mixed stands of Pi and Ci (Pi_Ci) and pure Pi stands to test the impacts of different tree species on soil acidification. Numerous studies were performed in this hot spot, analyzing N deposition, forest health, soil quality, and litter decomposition [1,6,41,42]; however, the forest hydrology studies conducted in this location focused only on throughfall in Pi stands [1,43,44,45]. Therefore, this canonically acidic site provides an opportunity to study the effects of precipitation, throughfall, stemflow, and floor leachate on soil acidity in different forest types and fill the knowledge gap on this topic.
In this study, our objectives were to ascertain the impacts of different waterflow types and forest canopy types on nutrient dynamics in southwest China. We hypothesized that (1) compared to decades ago, the pH of precipitation in Tieshanping has increased and transformed from sulfate acid rain to nitrate acid rain; (2) the influence of throughfall on the nutrient cycle is greater than that of stemflow; (3) a plantation of broadleaved species (Ci) is best to improve the quality of acidic soil, followed by mixed deciduous and conifer, and pure conifer stands.
2. Materials and Methods
2.1. Study Area and Plot Design
The experiment was conducted in Tieshanping Forest Park (106°41.24′ E, 29°37.42′ N), which was a heavily polluted “acid rain control zone” located about 25 km northwest of Chongqing in subtropical southwest China. In June 2018, in each of the three different forest stands (Pi, Pi_Ci, Ci) we established four 20 m × 20 m square study plots. The details about the study site can be found in Chen et al. [10]. The soil is classified as Haplic Acrisol in FAO (IUSS Working Group WRB, 2006), with low pH values (3.80–4.54 from the O horizon to the lower A horizon) [10].
2.2. Collection and Analysis of Water Samples
Bulk precipitation, throughfall, stemflow, and litter leachate were sampled as per Wang et al. [46] and measured in July 2018, October 2018, January 2019 and April 2019, which represent summer, autumn, winter, and spring, respectively. Bulk precipitation and throughfall were collected using stainless steel funnels (20 cm diameter) placed about 1.5 m above the forest floor. Three collectors were placed in an open field outside of the forest perimeter to measure bulk precipitation. For throughfall measurement (Figure 1a), five collectors were randomly placed in each plot. The funnels drained into opaque 10 L plastic buckets, which were placed below the ground surface. The stemflow (Figure 1b) was collected by 2 cm o.d. Tygon tubing with a length of about 100 cm. Before installing the tubing, the bark was shaved to ensure the seal without damage to the cambium layer. The tubing was split longitudinally and wrapped spirally around the tree stem, fastened with staples, sealed to the bole with acrylic caulk, and inserted into a 10 L plastic bucket. Litter leachate (Figure 1c) was collected using a topless 30 cm × 30 cm × 8 cm (length × width × height) stainless steel cuboid, which was covered with wire netting (1.5 mm mesh size). Each collector was connected with tubing to a 10 L plastic bucket, which was placed below ground level. The collectors were installed directly under the litter layer with five replicates per plot.

Figure 1.
Water sample collectors: (a) throughfall, (b) stemflow, (c) litter leachate.
All water samples were collected weekly in the four sampling months and lumped into monthly samples for chemical analysis (stored at 4 °C before pooling). In total, there were 12 samples for precipitation and 16 samples, respectively, for throughfall, stemflow, and litter leachate for each forest type spanning July 2018 to April 2019.
All samples were filtered through 0.45 μm member filters before analyses. Water pH and electrical conductivity (EC) were measured by a benchtop pH meter (REX PHS-3C, Shanghai, China) and a benchtop conductivity meter (REX DDS-12A, Shanghai, China), respectively. We used a total organic carbon analyzer (TOC-L, SHIMADZU, Kyoto, Japan) to determine the dissolved organic carbon in all samples. The content of Ca and Mg cations was analyzed by an atomic absorption spectrophotometer (AAS, HITACHI ZA-3300, Tokyo, Japan), and K and Na concentration was measured using a flame photometer (FP640, Shanghai, China). An ion chromatograph (ICP, Dionex ICS-900, USA) was used to determine the anion concentration in all samples. NO3-N and NH4-N were measured using a flow analyzer (Seal AA3, Germany).
2.3. Data Curation and Statistical Analysis
Ion concentration was represented as ratio of cations to anions (RCA), calculated as ∑+ (sum of K+, Na+, Ca2+, Mg2+, NH4+ concentration) to ∑− (sum of F−, Cl−, SO42−, NO3− concentration). The ion fluxes were calculated by multiplying ion concentrations by water fluxes. Base cation and anion fluxes (∑+ flux and ∑− flux) were, respectively, the sum of K+, Na+, Ca2+, Mg2+, NH4+, and F−, Cl−, SO42−, NO3−.
Water pH, EC, DOC, and RCA were analyzed by using one-way ANOVA with Tukey’s HSD to determine the different ion concentration among forest types respectively for stemflow, throughfall, and litter leachate. For each forest type, one-way ANOVA, Tukey’s HSD was also used to detect the seasonal variation across water properties. Independent t-tests were performed to determine significant differences across water properties, between precipitation and stemflow, throughfall, or litter leachate. All figures were created in OriginPro 9.0.
3. Results
3.1. Precipitation
The pH values of precipitation varied from 6.58 to 7.33 in the four sampling months, which indicated the precipitation was not acid rain in these months. The pH, EC and DOC contents had similar seasonal variation with highest values in January (Table 1).

Table 1.
pH, EC (μs·cm−1), DOC concentration (mg·L−1) and ion concentration (μeq·L−1) in precipitation.
Ca2+ was proportionally the most abundant cation, ranging between 24.81–71.62%, and SO42− was the most abundant anion (varying from 55.65–66.05%), followed by NO3− (11.02–22.73%). There was no significant seasonal variation for RCA in precipitation (p = 0.862) (Table 1). Following the peak ion concentration in January, cation and anion flux also peaked in January (Figure 2). Ca2+ flux predominated in July and April (respectively 51.16% and 52.53% of the whole cation flux), while K flux was greatest in October with 7.84 kg·hm−2. In January, Ca and Mg flux combined accounted for about 30% of the whole cation flux. SO42− flux accounted for 59.94–73.34% of all anion flux (Figure 2).
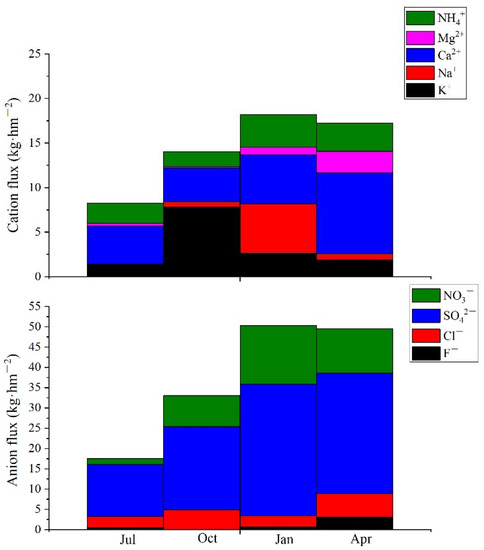
Figure 2.
Cation and anion flux in precipitation.
3.2. Throughfall, Stemflow, and Litter Leachate
3.2.1. pH, EC, and DOC
The pH values (6.56–7.87) of all throughfall samples were slightly higher than that of precipitation, although not statistically significant (p > 0.05), with the exception of the October samples in Pi_Ci and Ci. The pH values in all stemflow samples were significantly lower than that of precipitation (p < 0.05), with the exception of the January samples. Forest types had significant effects on stemflow pH (p < 0.001), but not on throughfall pH (p = 0.491). Stemflow pH was about 5.6 in Pi and Pi_Ci, except for the January samples, while the pH of stemflow in Ci varied from 5.66–6.62. There was significant seasonal variation in pH values for stemflow (p < 0.001) and throughfall (p < 0.001), with significantly higher values in January. There were no observed seasonal effects on the pH of litter leachate (p = 0.061), whereas forest type had a significant effect (p < 0.001). The pH values of litter leachate in the Pi_Ci and Ci plots were significantly (p < 0.05) higher than that in the Pi plot, except for Pi_Ci sampling in January (Figure 3a).
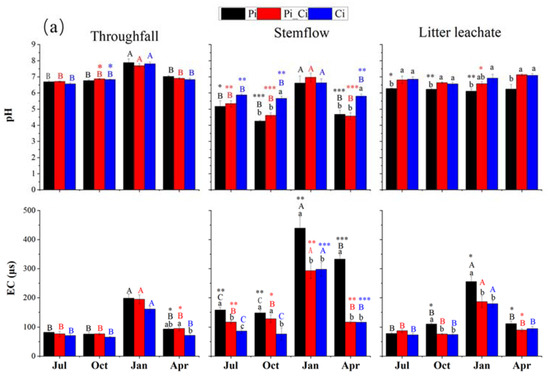
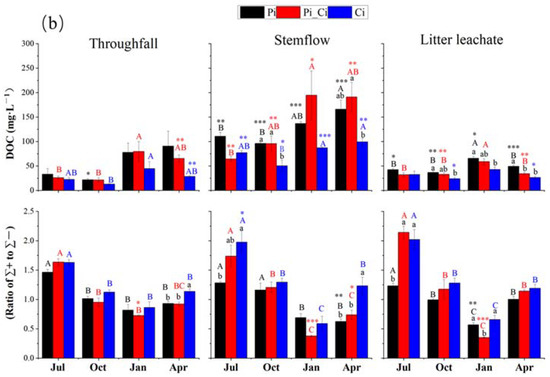
Figure 3.
pH, electrical conductivity (a), dissolved organic carbon, and ratio of ∑+ to ∑− (RCA) (b) of throughfall, stemflow, and litter leachate in different forest types during the sampling months. Capital letters indicate significant differences between sampling times for waterflows of each forest type. Lowercase letters indicate significant effects of forest type at the same sampling time. Asterisks indicate significant differences between precipitation and stemflow, throughfall, and litter leachate at the same sampling time. *, ** and *** respectively indicate significance at p < 0.05, 0.01 and 0.001.
Both EC and DOC of stemflow were significantly greater compared to those of precipitation (p < 0.05), except for EC of Ci in July and October. Forest type had opposite effects on EC and DOC concentration compared to pH of stemflow and litter leachate, with unclear impacts on throughfall. In Pi stands, the EC of stemflow (ranging from 147.7 to 439.25 μs) was significantly higher than that in Pi_Ci (117.50–292.88 μs) and Ci (76.45–297.75 μs) during all the sampling months (p < 0.05), except for the October samples in Pi_Ci. Among all water samples, the DOC concentration in precipitation was the lowest, with an average of 24 mg·L−1, compared to higher DOC levels in throughfall, litter leachate, and especially stemflow. Forest type did not impact DOC of throughfall. DOC concentration in stemflow (mean of 114.18 mg·L−1) was significantly higher than in precipitation in all plantations from all sampling months (p < 0.05). DOC concentration in stemflow from Ci (50.28–99.41 mg·L−1) was lower than that from Pi_Ci (64.44–194.57 mg·L−1) and Pi (96.34–166.16), and the difference was statistically significant in October (p = 0.027) and April (p = 0.032). Similarly, DOC concentration in Ci litter leachate was significantly lower than that in the Pi_Ci and Pi (p < 0.001) litter leachate samples (Figure 3b).
DOC flux in throughfall for Pi and Pi_Ci was triple and double that of precipitation in April, respectively. Due to lower volume, DOC flux in stemflow was reduced compared to that in precipitation, despite the higher concentration of DOC in stemflow. DOC flux in litter leachate was lower than that in precipitation, except in July, and was much lower than that in throughfall. Throughfall DOC in Ci was lower by 46.70%, 34.27%, 42.70% and 55.59% in July, October, January, and April, respectively, and litter leachate DOC flux was significantly lower, by 41.58%, compared to Pi in April (p = 0.027) (Table 2).
Table 2.
The flux of DOC in precipitation, throughfall, stemflow and litter leachate. BP: bulk precipitation; TF: throughfall; SF: stemflow; LL: litter leachate. Different lowercase letters indicate significant difference among forest types.
3.2.2. Ion Concentration and Flux
Most cation and anion contents were higher in concentration in throughfall, litter leachate, and especially stemflow, compared to precipitation. Ca2+ and SO42− were, respectively, the most abundant cation and anion (Figure 4). The mean sum of cation concentration was 605, 535.53, 966.26 and 662.97 μeq·L−1, and the mean sum of anion concentration was 755.94, 872.13, 1812.54 and 1110.88 μeq·L−1 in precipitation, throughfall, stemflow, and litter leachate, respectively (Figure 4).
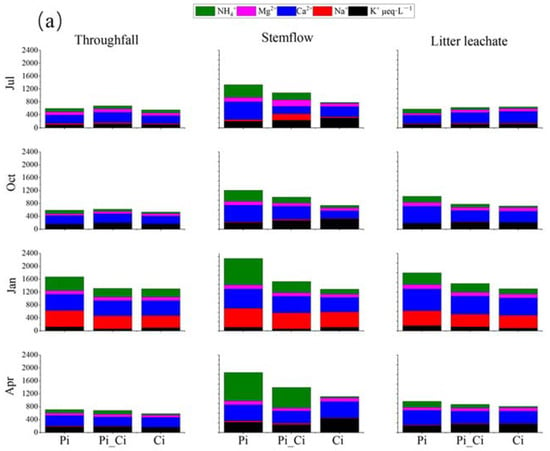
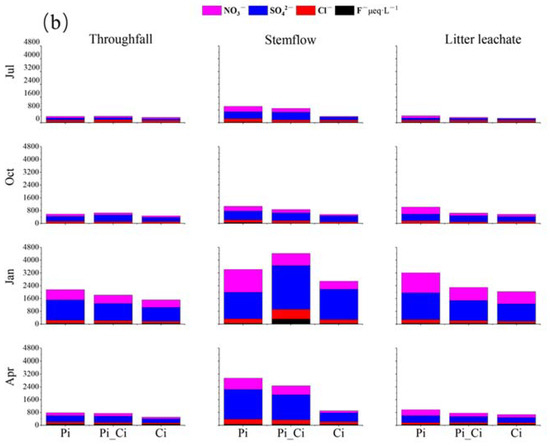
Figure 4.
Cation (a) and anion (b) concentrations in throughfall, stemflow, and litter leachate among different forest types during different sampling months.
We also found that the mean sum of cation or anion concentration could be ranked in the order Pi > Pi_Ci > Ci. In Ci, the greater difference in anions (lower by 32.87% and 39.08% compared to Pi_Ci and Pi) than cations (lower by 15.37% and 27.28% than Pi_Ci and Pi) resulted in higher RCA in Ci (Figure 3b). RCA in July was significantly higher than that in the other sampling months for all forest types (p < 0.05), except for stemflow sampling in October in Pi. Generally, RCA in Ci was larger than that in Pi, and the difference was statistically significant in July for stemflow (p = 0.025) and litter leachate (p = 0.011), and in April for stemflow (p = 0.005) and throughfall (p = 0.014). The mean value of RCA in Ci was greater by 35.60%, 12.52% and 35.53% compared to Pi, and by 25.26%, 12.29% and 6.97% compared to Pi_Ci for stemflow, throughfall, and litter leachate, respectively (Figure 3b).
Due to the low nutrient flux (0.01875 to 0.3695 and 0.03950 to 0.6302 kg·hm−2, respectively, for cation and anion flux) in stemflow, in Figure 5 we show the flux as throughfall plus stemflow (TF+SF) and litter leachate (LL). The litter layer intercepted nutrient flux from forest rainwater, with the exception of NO3−. The leaf litter in Pi stands leached the most NO3− (3.39–9.60 kg·hm−2), which was much greater than that from Ci stand litter (0.34 to 5.24 kg·hm−2). Due to different capacities for leaching NO3− and intercepting SO42−, litter leachate of Ci showed the lowest flux of NO3− plus SO42− (5.23, 10.99, 17.04, and 14.79 kg·hm−2), followed by Pi_Ci (7.84, 10.24, 17.96 and 18.47 kg·hm−2), and then Pi, with the highest values observed (10.1, 15.98, 19.79, and 22.74 kg·hm−2) in July, October, January, and April, respectively.
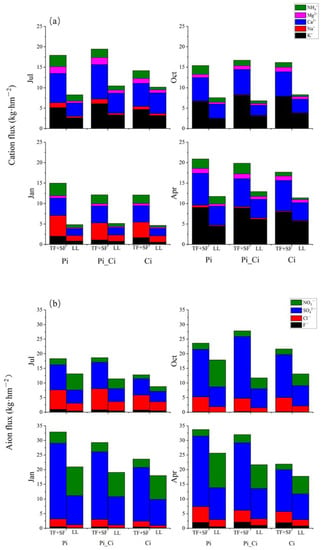
Figure 5.
Cation (a) and anion (b) flux in throughfall plus stemflow and litter leachate. TF: throughfall; SF: stemflow; LL: litter leachate.
The cation and anion fluxes of TF + SF in Ci were lower than those in Pi_Ci and Pi (Figure 5), except in October, while the ratio of cation flux to anion flux of throughfall and stemflow in Ci was significantly higher than that in Pi in July and April (Table 3). Compared to Ci stands, the ratio of cation flux to anion flux in litter leachate of Pi_Ci was greater by 17.39–46.03%. In Ci stands, the ratio of cation flux to anion flux was 80.95%, 39.53% and 36.17% greater than that in Pi stands in July, October, and April, respectively (Table 3). Additionally, for stemflow and litter leachate, base saturation in Ci was significantly higher than that in Pi, except for litter leachate in January, and also significantly higher than that in Pi_Ci, except for stemflow in January and litter leachate in July and January (Table 4).
Table 3.
Ratio of cation flux to anion flux across three forest types during four sampling months. Different lowercase letters indicate significant difference among forest types.
Table 4.
Base saturation ((K+ + Ca2+ + Mg2+): ∑+) in throughfall, stemflow and litter leachate across different forest types. Different lowercase letters indicate significant difference among forest types.
4. Discussion
4.1. The Change of Precipitation
The data in this study indicated that precipitation in Tieshanping was neutral with pH values varying from 6.58–7.33. To evaluate the change in precipitation quality, we compared our data with previous data from 2001–2004 [47] and 2003 [48] in the same study site (Table 1). The mean pH values from 2001–2004 [47] and 2003 [48] were about 4.1, which was more acidic than the mean pH observed in this study, from 2018–2019, by over 2 units, and this could be due to decreased acidic emissions in China [1]. Most of the average ion concentrations substantially increased compared to mean values from 2001–2004, especially for K+ (3.5×), Ca2+ (8.6×), Cl− (6.8×), SO42− and NO3− (6–7×). In 2003 [48], most ion concentrations were much lower than those of 2001–2004, except for Ca2+, SO42− and NO3−, which were still lower than the values observed in this study. The ratio of SO42− to NO3− in our study was an average of 3.16, which was much lower than that of the other two previous studies, at 4.27 and 5.3, respectively, in Xiang et al. [47] and Wang and Xu [48] in the same study site in Tieshanping. This increasing proportion of NO3− in precipitation compared to previous studies [47,48] was also confirmed by Chen [41] in Chongqing. The increased RCA also confirmed the transformation of precipitation from acidic towards neutral. These data partially confirm our first hypothesis that the pH of precipitation and the proportion of NO3− have increased. However, due to the large interannual variability observed over 2001–2004 [47], the 1-year monitoring data in this study are not sufficient to assert that precipitation in Tieshanping is presently neutral, not acidic. In order to do so, more observational data over a longer period would be needed.
4.2. The Effects of Water Flow
In our study, both cation and anion fluxes in litter leachate were lower than that in throughfall plus stemflow, which indicated an interception of nutrients at the litter layer. To further detect how much throughfall and stemflow influence the nutrient flux of litter leachate, we performed variation portioning analysis (VPA) using cation, anion, and DOC flux. As indicated by VPA, a total of 68% of litter leachate nutrient flux could be explained by throughfall and stemflow. Nutrient flux from throughfall and stemflow respectively contributed 20% and 3% to litter leachate nutrient flux, while their common contribution was 45%. The contribution of throughfall and stemflow to the forest floor has also been confirmed by previous data [29,32]. The relatively small contribution of stemflow observed in this study is supported by Bellot et al. [49], which found that nutrient inputs of stemflow only represented about 2–3% of total dissolved inputs into the soils. Despite the low contribution, stemflow can play an important role in forest ecosystems, as it concentrates water in the area immediately proximal to the tree trunk, providing a supply of water and nutrients to the tree and soil far more so than throughfall [49,50].
This present study also indicates that stemflow is enriched with a greater nutrient concentration than throughfall. We found the highest DOC content, cation and anion concentration, and lowest pH value in stemflow, followed by throughfall and litter leachate. Higher concentrations of nutrients in stemflow can be attributed to both nutrients leaching from stems and very low stemflow volume [51]. When we replaced nutrient flux with concentration in VPA analysis, the contribution of stemflow to litter leachates increased from 3% to 9%, which further illustrates the importance of stemflow in influencing nutrient concentrations in the root zone that affect rhizosphere microbial communities and function [52]. Thus, our second hypothesis is partially correct in that throughfall does contribute a greater nutrient flux to the forest floor than stemflow; however, the transient impacts of stemflow are more intensive on the root zone, specifically.
4.3. The Effects of Forest Type
A previous review that analyzed data from 17 stand pairs determined that coniferous forests intercept more pollutants than deciduous stands [31], a conclusion that our findings support as well. In this study, cation and anion flux in Ci were much lower than in Pi and Pi_Ci in throughfall plus stemflow; however, Ci had the highest pH and RCA and the lowest DOC in stemflow across three forest stands. A study in northwest China illustrated that the enrichment capacity of acid anions and exchangeable bases in the canopy of conifers is stronger than that of broadleaved species [27]. While these results were consistent with ours, Han et al. [27] recommended planting coniferous species to improve forest ecosystem function, a measure that may not be suitable in our study site. The study site in Han et al. [27] was located in northwest China, which does not experience acid rain or soil acidification [53]. Our study was conducted in a typical acid-polluted area with severely acidic soil (average pH of 3.98, 4.54, and 4.6, respectively, in O_horizon of Pi, Pi_Ci and Ci, [10]), which would benefit from species that could supply less acidic rain water and greater base saturation. We also calculated the base saturation of throughfall, stemflow, and litter leachate, finding that Ci had the highest base saturation, followed by Pi_Ci (Table 4). We also found that the sum of anion flux in litter leachate from Ci was much lower than that of Pi, which was mainly due to a lower amount of NO3− flux in Ci. The lower flux of NO3− from litter leachate in Ci than in Pi observed in this study is consistent with results from central Japan, which showed that average NO3− flux of litter leachate in evergreen forests was higher than that in deciduous forests [54]. Our data showed that throughfall N input was negatively associated with soil pH, which is supported by a study from an urban-to-rural transect in southern China [55]. In this study, throughfall NO3− flux was the highest in Pi stands, and the lowest in Ci stands, where lower N input of throughfall (plus stemflow) and litter leachate contributed to higher pH values of the soils, an observation supported by our previous data as well [10].
We also found that the DOC concentration and flux in throughfall and litter leachate were minimum in Ci and maximum in Pi. Though DOC flux in stemflow of Ci was relatively higher, it was negligible because of its small volume compared to throughfall and litter leachate. In our study, the DOC concentration in throughfall and DOC fluxes in forest floor leachates were found to be higher in coniferous species stands than in broadleaved species stands, which was supported by some previous studies [29,40,56]. DOC could influence water acidity due to the composition of short-chain acids and large molecules such as humic and fulvic acids [57]. Previous studies indicated that tree species with high DOC in their forest floor leachates stimulate base cation leaching, which would cause soil acidification [58].
In summary, Ci is more beneficial for improving soil acidification than Pi based on the acidic status of soils in Tieshanping. However, our findings are only based on measurements from four 1-month periods, which limited our knowledge of the effects of forest types on soil acidity. Longer-term and more-detailed monitoring data on different forest types in this study area will be needed in the future to further ascertain the effects of different water flow and forest types on soil quality.
5. Conclusions
Our monitoring data indicated that precipitation in Tieshanping was neutral and the proportion of NO3− was increased. Despite very low nutrient fluxes, stemflow also played an important role in affecting the nutrient cycling in the area near the stem by increasing nutrient concentrations. According to hydrological chemical characteristics, pure plantations of Ci could supply lower N and DOC inputs to soils with higher base saturation for improving soil acidity, followed by mixed Ci and Pi stands. In stands of Ci, higher RCA and base saturation accompanied by lower N and DOC inputs in throughfall plus stemflow and litter leachate yielded higher pH in water flow entering the soils, which would support recovery from soil acidification and further ensure sustainable development of forest ecosystems in acid-polluted locations. This study underscores the changing rainwater chemistry of forest hydrological processes and the effects on soil acidification across mixed and pure stands of coniferous species and broadleaved species. While these findings are based on four 1-month periods of measurement, to clearly determine the water chemistry of precipitation and the effects of forest type in Tieshanping, more observational data over a longer period are needed.
Author Contributions
Conceptualization, Z.C.; Data curation, Z.C. and Y.W.; Formal analysis, R.C.; Funding acquisition, Z.C.; Investigation, Z.C. and J.C.; Methodology, Y.W.; Project administration, Z.C.; Software, R.C.; Writing—original draft, Z.C.; Writing—review and editing, Z.C. and X.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Fundamental Research Funds for the Central Non-profit Research Institution of CAF (CAFYBB2018SZ004) and the National Natural Science Foundation of China (31971630).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors appreciate Tieshanping Forest Park of Chongqing for helping with the field work. We are also grateful to Mingquan Zou and Qian Zhang for their contribution to the field sampling and soil analysis. Aishwarya Veerabahu from the University of California, Riverside is highly appreciated for her English editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Duan, L.; Yu, Q.; Zhang, Q.; Wang, Z.; Pan, Y.; Larssen, T.; Tang, J.; Mulder, J. Acid deposition in Asia: Emissions, deposition, and ecosystem effects. Atmos. Environ. 2016, 146, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; He, N.; Wang, Q.; Zhu, J.; Gao, Y.; Zhang, Y.; Jia, Y.; Yu, G. Development of atmospheric acid deposition in China from the 1990s to the 2010s. Environ. Pollut. 2017, 231, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.X.; Zhao, B.; Cai, S.Y.; Klimont, Z.; Nielsen, C.P.; Morikawa, T.; Woo, J.H.; Kim, Y.; Fu, X.; Xu, J.Y.; et al. Emission trends and mitigation options for air pollutants in East Asia. Atmos. Chem. Phys. 2014, 14, 6571–6603. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Liu, M.; Song, Y.; Huang, X.; Yao, H.; Cai, X.; Zhang, H.; Kang, L.; Liu, X.; Yan, X.; et al. High-resolution ammonia emissions inventories in China from 1980 to 2012. Atmos. Chem. Phys. 2016, 16, 2043–2058. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, Z.; He, J.Z.; Wang, Q.; Shen, C.; Ge, Y. Ectomycorrhizal fungi inoculation alleviates simulated acid rain effects on soil ammonia oxidizers and denitrifiers in Masson pine forest. Environ. Microbiol. 2019, 21, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Solberg, S.; Yu, P.; Myking, T.; Vogt, R.D.; Du, S. Assessments of tree crown condition of two Masson pine forests in the acid rain region in south China. For. Ecol. Manag. 2007, 242, 530–540. [Google Scholar] [CrossRef]
- Ouyang, X.J.; Zhou, G.Y.; Huang, Z.L.; Liu, J.X.; Zhang, D.Q.; Li, J. Effect of Simulated Acid Rain on Potential Carbon and Nitrogen Mineralization in Forest Soils. Pedosphere 2008, 18, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Nduka, J.K.C.; Orisakwe, O.E.; Ezenweke, L.O.; Ezenwa, T.E.; Chendo, M.N.; Ezeabasili, N.G. Acid rain phenomenon in Niger Delta region of Nigeria: Economic, biodiversity, and public health concern. Sci. World J. 2008, 8, 811–818. [Google Scholar] [CrossRef] [Green Version]
- Reis, S.; Grennfelt, P.; Klimont, Z.; Amann, M.; ApSimon, H.; Hettelingh, J.P.; Holland, M.; LeGall, A.C.; Maas, R.; Posch, M.; et al. From acid rain to climate change. Science 2012, 338, 1153–1154. [Google Scholar] [CrossRef]
- Chen, Z.; Maltz, M.R.; Zhang, Y.; O’brien, B.J.; Neff, M.; Wang, Y.; Cao, J. Plantations of cinnamomum camphora (Linn) presl with distinct soil bacterial communities mitigate soil acidity within polluted locations in southwest China. Forests 2021, 12, 657. [Google Scholar] [CrossRef]
- Singh, A.; Agrawal, M. Acid rain and its ecological consequences. J. Environ. Biol. 2008, 29, 15–24. [Google Scholar] [PubMed]
- Onu, P.U.; Quan, X.; Xu, L.; Orji, J.; Onu, E. Evaluation of sustainable acid rain control options utilizing a fuzzy TOPSIS multi-criteria decision analysis model framework. J. Clean. Prod. 2017, 141, 612–625. [Google Scholar] [CrossRef] [Green Version]
- Graf Pannatier, E.; Thimonier, A.; Schmitt, M.; Walthert, L.; Waldner, P. A decade of monitoring at Swiss Long-Term Forest Ecosystem Research (LWF) sites: Can we observe trends in atmospheric acid deposition and in soil solution acidity? Environ. Monit. Assess. 2011, 174, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Marx, A.; Hintze, S.; Sanda, M.; Jankovec, J.; Oulehle, F.; Dusek, J.; Vitvar, T.; Vogel, T.; van Geldern, R.; Barth, J.A.C. Acid rain footprint three decades after peak deposition: Long-term recovery from pollutant sulphate in the Uhlirska catchment (Czech Republic). Sci. Total Environ. 2017, 598, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Menyailo, O.V.; Hungate, B.A.; Zech, W. Tree species mediated soil chemical changes in a Siberian artificial afforestation experiment: Tree species and soil chemistry. Plant Soil 2002, 242, 171–182. [Google Scholar] [CrossRef]
- Binkley, D.; Menyailo, O. Gaining insights on the effects of tree species on soils. In Tree Species Effects on Soils: Implications for Global Change; Binkley, D., Menyailo, O., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2005; pp. 1–16. [Google Scholar]
- Dunkerley, D. A review of the effects of throughfall and stemflow on soil properties and soil erosion. In Precipitation Partitioning by Vegetation; Van Stan, J.T., Gutmann, E., Friesen, J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 183–214. [Google Scholar]
- Qualls, R.G. Role of precipitation partitioning in litter biogeochemistry. In Precipitation Partitioning by Vegetation; Van Stan, J.T., Gutmann, E., Friesen, J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 163–182. [Google Scholar]
- Bathiany, S.; Claussen, M.; Brovkin, V.; Raddatz, T.; Gayler, V. Combined biogeophysical and biogeochemical effects of large-scale forest cover changes in the MPI earth system model. Biogeosciences 2010, 7, 1383–1399. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Lan, X.; Li, W.; Zhu, W.; Li, T. Influence of land use change on the surface albedo and climate change in the qinling-daba mountains. Sustainability 2021, 13, 10153. [Google Scholar] [CrossRef]
- Nikodem, A.; Kodešová, R.; Bubeníčková, L. Simulation of the influence of rainfall redistribution in spruce and beech forest on the leaching of Al and SO42- from forest soils. J. Hydrol. Hydromechanics 2013, 61, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Aubrey, D.P. Relevance of precipitation partitioning to the tree water and nutrient balance. In Precipitation Partitioning by Vegetation; Van Stan, J.T., Gutmann, E., Friesen, J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 147–162. [Google Scholar]
- Crockford, R.H.; Richardson, D.P. Partitioning of rainfall into throughfall, stemslow and interception effect of forest type, ground cover and climate. Hydrol. Process. 2000, 14, 2903–2920. [Google Scholar] [CrossRef]
- Legout, A.; van der Heijden, G.; Jaffrain, J.; Boudot, J.P.; Ranger, J. Tree species effects on solution chemistry and major element fluxes: A case study in the Morvan (Breuil, France). For. Ecol. Manag. 2016, 378, 244–258. [Google Scholar] [CrossRef]
- Betts, R.A.; Cox, P.M.; Lee, S.E.; Woodward, F.I. Contrasting physiological and structural vegetation feedbacks in climate change simulations. Nature 1997, 387, 796–799. [Google Scholar] [CrossRef]
- Betts, R.A. Offset of the potential carbon sink from boreal forestation by decreases in surface albedo. Nature 2000, 408, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Zhang, C.; Liu, Y.; Li, Y.; Zhou, T.; Khan, S.; Chen, N.; Zhao, C. The capacity of ion adsorption and purification for coniferous forests is stronger than that of broad-leaved forests. Ecotoxicol. Environ. Saf. 2021, 215, 112137. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.W.; Untersteiner, H.; Schume, H.; Jost, G. Throughfall fluxes in a secondary spruce (Picea abies), a beech (Fagus sylvatica) and a mixed spruce-beech stand. For. Ecol. Manag. 2008, 255, 605–618. [Google Scholar] [CrossRef]
- Hamdan, K.; Schmidt, M. The influence of bigleaf maple on chemical properties of throughfall, stemflow, and forest floor in coniferous forest in the Pacific Northwest. Can. J. For. Res. 2012, 42, 868–878. [Google Scholar] [CrossRef]
- Van Nevel, L.; Mertens, J.; De Schrijver, A.; Baeten, L.; De Neve, S.; Tack, F.M.G.; Meers, E.; Verheyen, K. Forest floor leachate fluxes under six different tree species on a metal contaminated site. Sci. Total Environ. 2013, 447, 99–107. [Google Scholar] [CrossRef]
- De Schrijver, A.; Geudens, G.; Augusto, L.; Staelens, J.; Mertens, J.; Wuyts, K.; Gielis, L.; Verheyen, K. The effect of forest type on throughfall deposition and seepage flux: A review. Oecologia 2007, 153, 663–674. [Google Scholar] [CrossRef]
- Carnol, M.; Bazgir, M. Nutrient return to the forest floor through litter and throughfall under 7 forest species after conversion from Norway spruce. For. Ecol. Manag. 2013, 309, 66–75. [Google Scholar] [CrossRef]
- Chapin, F.S.; Walker, B.H.; Hobbs, R.J.; Hooper, D.U.; Lawton, J.H.; Sala, O.E.; Tilman, D. Biotic control over the functioning of ecosystems. Science 1997, 277, 500–504. [Google Scholar] [CrossRef] [Green Version]
- Fujinuma, R.; Bockheim, J.; Balster, N. Base-cation cycling by individual tree species in old-growth forests of Upper Michigan, USA. Biogeochemistry 2005, 74, 357–376. [Google Scholar] [CrossRef]
- Strobel, B.W.; Borggaard, O.K.; Hansen, H.C.B.; Andersen, M.K.; Raulund-Rasmussen, K. Dissolved organic carbon and decreasing pH mobilize cadmium and copper in soil. Eur. J. Soil Sci. 2005, 56, 189–196. [Google Scholar] [CrossRef]
- Erisman, J.W.; Draaijers, G. Deposition to forests in Europe: Most important factors influencing dry deposition and models used for generalisation. Environ. Pollut. 2003, 124, 379–388. [Google Scholar] [CrossRef]
- De Vries, W.; van der Salm, C.; Reinds, G.J.; Erisman, J.W. Element fluxes through European forest ecosystems and their relationships with stand and site characteristics. Environ. Pollut. 2007, 148, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.; Lohse, K.A.; Hall, S.J. The globalization of nitrogen deposition: Consequences for terrestrial ecosystems. Ambio 2002, 31, 113–119. [Google Scholar] [CrossRef]
- Gundersen, P.; Schmidt, I.K.; Raulund-Rasmussen, K. Leaching of nitrate from temperate forests—Effects of air pollution and forest management. Environ. Rev. 2006, 14, 1–57. [Google Scholar] [CrossRef]
- Lindroos, A.J.; Derome, J.; Derome, K.; Smolander, A. The effect of Scots pine, Norway spruce and silver birch on the chemical composition of stand throughfall and upper soil percolation water in northern Finland. Boreal Environ. Res. 2011, 16, 240–250. [Google Scholar]
- Chen, X.Y.; Mulder, J. Atmospheric deposition of nitrogen at five subtropical forested sites in South China. Sci. Total Environ. 2007, 378, 317–330. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Liu, Y.; Guo, H.; Li, T.; Li, Z.H.; Shi, G. Long-term effects of liming on health and growth of a Masson pine stand damaged by soil acidification in Chongqing, China. PLoS ONE 2014, 9, e94230. [Google Scholar] [CrossRef]
- Lydersen, E.; Angell, V.; Eilertsen, O.; Muniz, I.P.; Larssen, T.; Seip, H.M.; Aagaard, P.; Vogt, R.D.; Mulder, J. Planning of an Integrated Acidification Study and Survey on Acid Rain Impacts in China. Final Report; Norwegian Institute for Water Research: Oslo, Norway, 1997. [Google Scholar]
- Larssen, T.; Duan, L.; Mulder, J. Deposition and leaching of sulfur, nitrogen and calcium in four forested catchments in China: Implications for acidification. Environ. Sci. Technol. 2011, 45, 1192–1198. [Google Scholar] [CrossRef]
- Huang, Y.; Kang, R.; Mulder, J.; Zhang, T.; Duan, L. Nitrogen saturation, soil acidification, and ecological effects in a subtropical pine forest on acid soil in southwest China. J. Geophys. Res. G Biogeosci. 2015, 120, 2457–2472. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Wang, Y.H.; Li, Z.H.; Wang, Y.J. The characteristics of rainfall interception and water storage of natural secondary forests of Masson pine at Tieshanping of Chongqing. Acta Ecol. Sinica 2021, 41, 6542–6551. [Google Scholar]
- Xiang, R.J.; Chai, L.Y.; Zhang, Q.M.; Xiao, J.S.; Zhao, D.W. Zhao. Ions distributional characteristics of wet precipitation in typical acid rain areas of China. J. Cent. South Univ. 2012, 43, 38–45. [Google Scholar]
- Wang, W.X.; Xu, P.J. Research progress in precipitation chemistry in China. Process Chem. 2009, 21, 266–281. [Google Scholar]
- Bellot, J.; Àvila, A.; Rodrigo, A. Throughfall and Stemflow in Ecology of Mediterranean Evergreen Oak Forests; Springer: Berlin/Heidelberg, Germany, 1999; pp. 209–222. [Google Scholar]
- Levia, D.F.; Frost, E.E. A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems. J. Hydrol. 2003, 274, 1–29. [Google Scholar] [CrossRef]
- Wang, M.C.; Liu, C.P.; Sheu, B.H. Characterization of organic matter in rainfall, throughfall, stemflow, and streamwater from three subtropical forest ecosystems. J. Hydrol. 2004, 289, 275–285. [Google Scholar] [CrossRef]
- Levia, D.F.; Germer, S. A review of stemflow generation dynamics and stemflow-environment interactions in forests and shrublands. Rev. Geophys. 2015, 53, 673–714. [Google Scholar] [CrossRef]
- Xu, Z.; Li, Y.; Tang, Y.; Han, G. Chemical and strontium isotope characterization of rainwater at an urban site in Loess Plateau, Northwest China. Atmos. Res. 2009, 94, 481–490. [Google Scholar] [CrossRef]
- Ashik, T.; Onishi, T.; Islam, R.; Cao, R.; Ohtsuka, T. A paired catchment study of nitrogen dynamics in a cool-temperate mixed deciduous broad-leaved and coniferous evergreen forests, central Japan. In Proceedings of the International Symposium on a New Era in Food Science and Technology, Gifu, Japan, 9–10 October 2019; p. 23. [Google Scholar]
- Fang, Y.; Yoh, M.; Koba, K.; Zhu, W.; Takebayashi, Y.; Xiao, Y.; Lei, C.; Mo, J.; Zhang, W.; Lu, X. Nitrogen deposition and forest nitrogen cycling along an urban-rural transect in southern China. Glob. Chang. Biol. 2011, 17, 872–885. [Google Scholar] [CrossRef]
- Fröberg, M.; Hansson, K.; Kleja, D.B.; Alavi, G. Dissolved organic carbon and nitrogen leaching from Scots pine, Norway spruce and silver birch stands in southern Sweden. For. Ecol. Manag. 2011, 262, 1742–1747. [Google Scholar] [CrossRef] [Green Version]
- Dalva, M.; Moore, T.R. Sources and sinks of dissolved organic carbon in a forested swamp catchment. Biogeochemistry 1991, 15, 1–19. [Google Scholar] [CrossRef]
- Finzi, A.C.; Canham, C.D.; Breemen, N. Canopy tree-soil interactions within temperate forests: Species effects on pH and cations. Ecol. Appl. 1998, 8, 447–454. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).