
Efficient Removal of Methylene Blue Using Living Biomass of the Microalga Chlamydomonas moewusii: Kinetics and Equilibrium Studies



Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining the Microalgal Biomass
2.2. Stock Solution of the Dye
2.3. Characterization of the Biosorbent
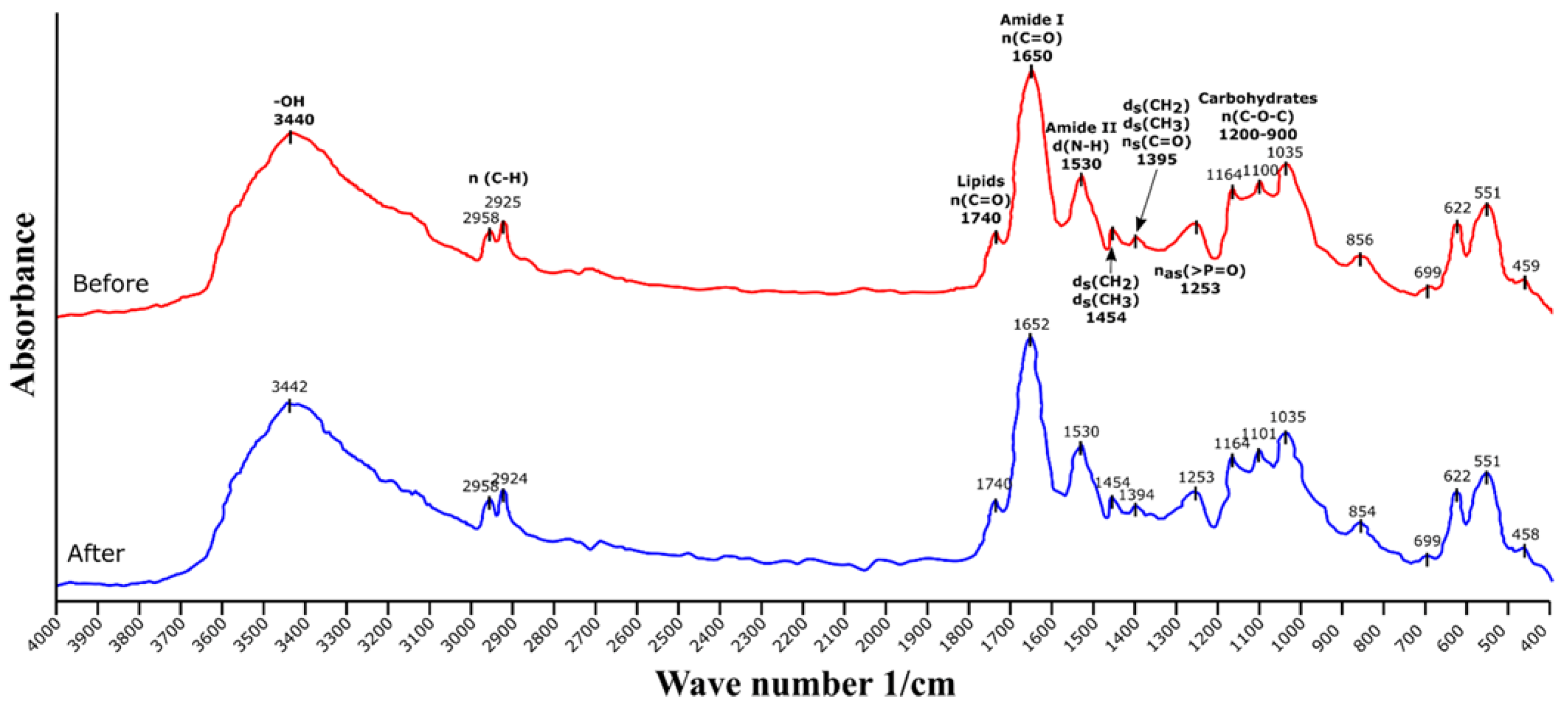
2.3.1. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
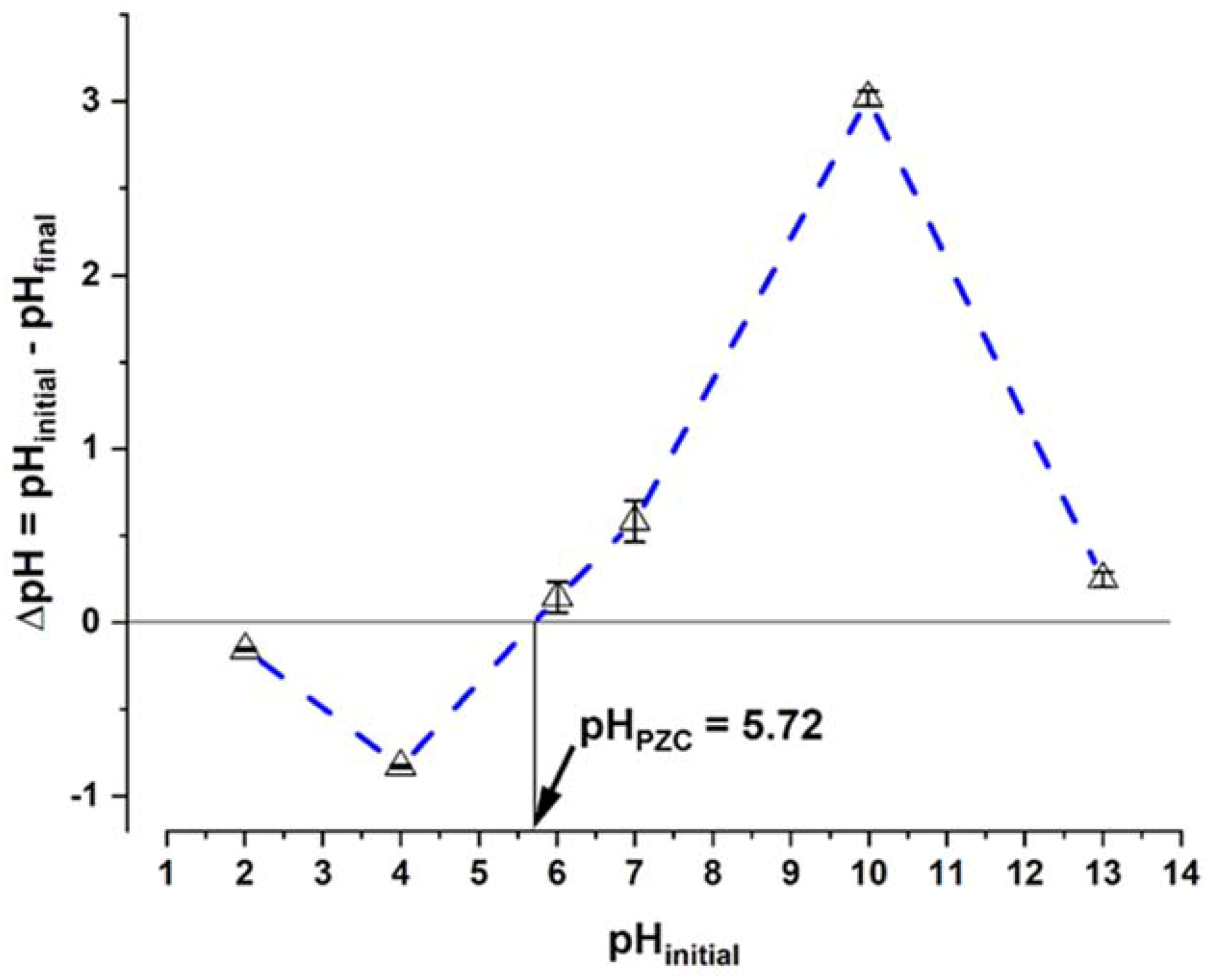
2.3.2. Point of Zero Charge (pHPZC) Determination
2.4. Batch Biosorption Studies
- A determined volume of the stock culture of the microalga equivalent to an amount of dry biomass of 40 mg (0.8 g/L) (this volume was also obtained after determining the cell density of the culture by means of a Neubauer chamber);
- An adequate volume of any methylene blue stock solution according to the concentration tested;
- Sterile deionized water until reaching a final volume of 50 mL.
2.5. Determination of the Effect of pH
2.6. Analytical Method
2.7. Removal Kinetics Analyses
2.8. Isotherm Studies
2.9. Statistical Analysis
3. Results and Discussion
3.1. Stability of the Dye: Photodegradation
3.2. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
3.3. Point of Zero Charge (pHPZC)
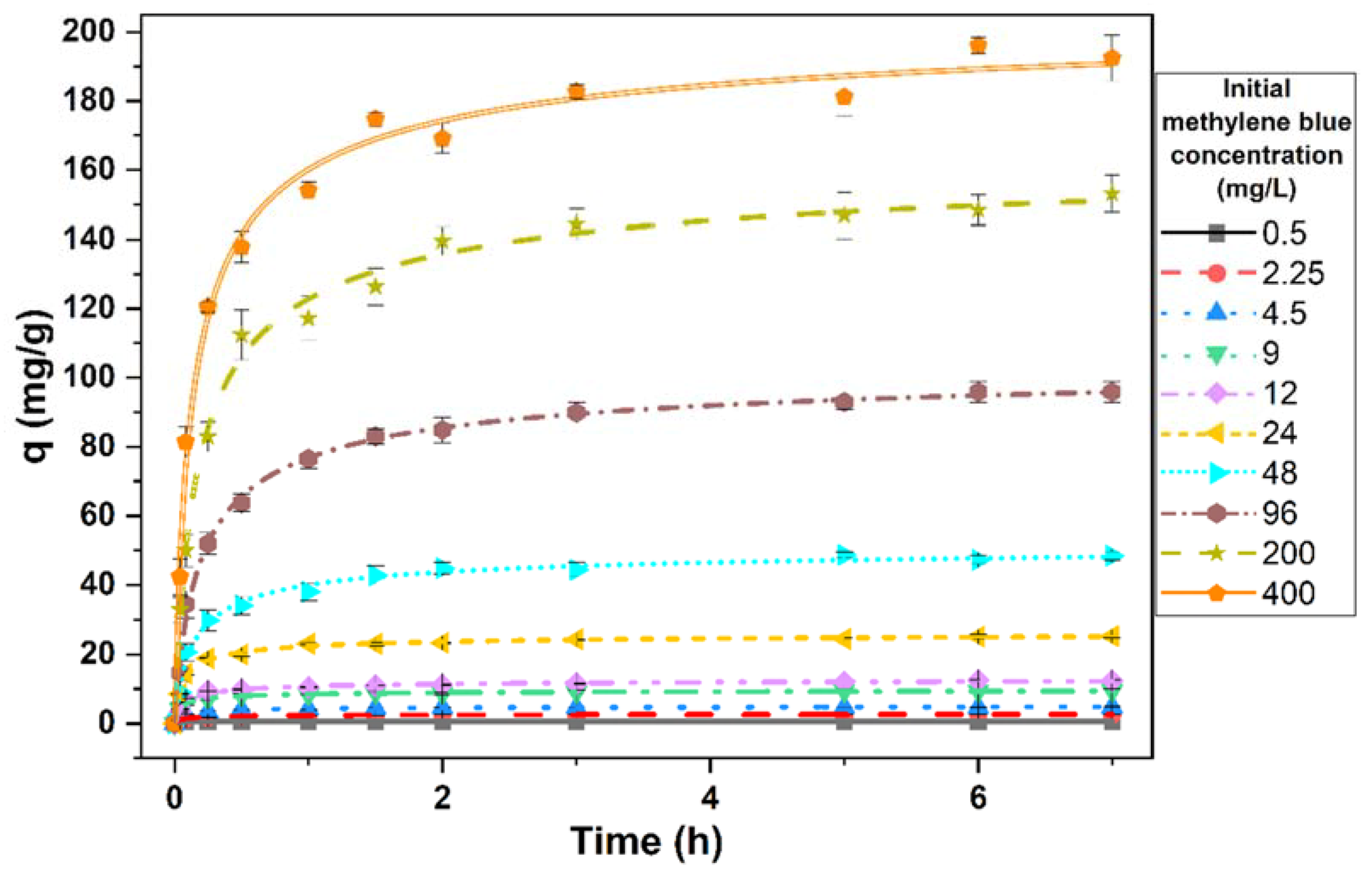
3.4. Effect of Contact Time on the Removal of Methylene Blue by Living Cells of the Microalga
3.5. Effect of the Initial Dye Concentration
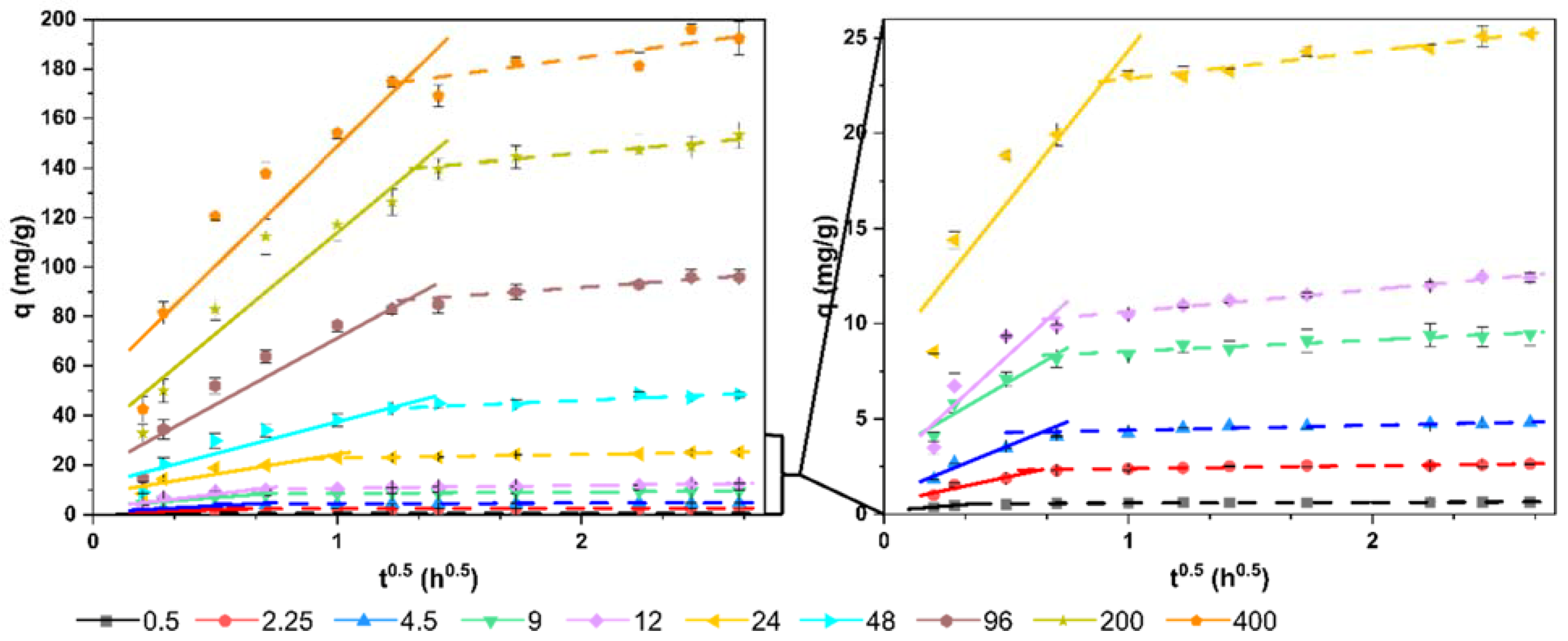
3.6. Removal Kinetics
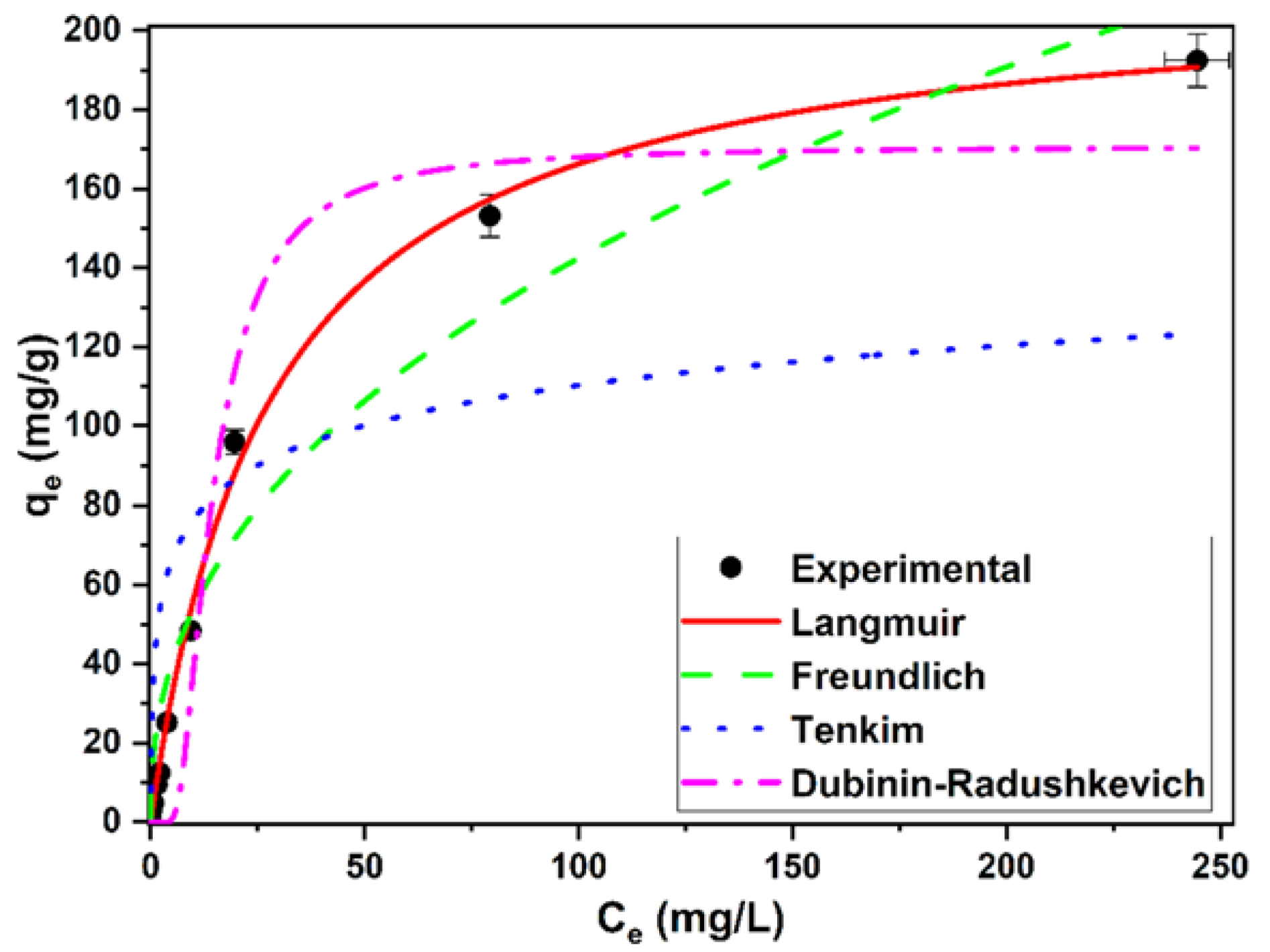
3.7. Isotherm Studies
3.8. Effect of pH
3.9. Comparison with Other Sorbents
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sabnis, R.W. Handbook of Biological Dyes and Stains: Synthesis and Industrial Applications; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Gita, S.; Shukla, S.P.; Deshmukhe, G.; Choudhury, T.G.; Saharan, N.; Singh, A.K. Toxicity Evaluation of Six Textile Dyes on Growth, Metabolism and Elemental Composition (C, H, N, S) of Microalgae Spirulina platensis: The Environmental Consequences. Bull. Environ. Contam. Toxicol. 2021, 106, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Croce, R.; Cina, F.; Lombardo, A.; Crispeyn, G.; Cappelli, C.I.; Vian, M.; Maiorana, S.; Benfenati, E.; Baderna, D. Aquatic toxicity of several textile dye formulations: Acute and chronic assays with Daphnia magna and Raphidocelis subcapitata. Ecotoxicol. Environ. Saf. 2017, 144, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Lellis, B.; Fávaro-Polonio, C.Z.; Pamphile, J.A.; Polonio, J.C. Effects of textile dyes on health and the environment and bioremediation potential of living organisms. Biotechnol. Res. Innov. 2019, 3, 275–290. [Google Scholar] [CrossRef]
- Oz, M.; Lorke, D.E.; Hasan, M.; Petroianu, G.A. Cellular and molecular actions of Methylene Blue in the nervous system. Med. Res. Rev. 2011, 31, 93–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgin, J.; Franco, D.S.P.; Netto, M.S.; Allasia, D.; Oliveira, M.L.S.; Dotto, G.L. Treatment of water containing methylene by biosorption using Brazilian berry seeds (Eugenia uniflora). Environ. Sci. Pollut. Res. Int. 2020, 27, 20831–20843. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Nascimento, L.; Brito, M.; da Silva, K.; Paschoal, W., Jr.; Fujiyama, R. Biosorption of Methylene Blue Dye Using Natural Biosorbents Made from Weeds. Materials 2019, 12, 2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebron, Y.A.R.; Moreira, V.R.; de Souza Santos, L.V. Biosorption of methylene blue and eriochrome black T onto the brown macroalgae Fucus vesiculosus: Equilibrium, kinetics, thermodynamics and optimization. Environ. Technol. 2021, 42, 279–297. [Google Scholar] [CrossRef]
- Santaeufemia, S.; Abalde, J.; Torres, E. Eco-friendly rapid removal of triclosan from seawater using biomass of a microalgal species: Kinetic and equilibrium studies. J. Hazard. Mater. 2019, 369, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yan, X.; Yao, J.; Zheng, S.; Wei, Q. Feasibility and comparative analysis of cadmium biosorption by living Scenedesmus obliquus FACHB-12 biofilms. Chemosphere 2021, 275, 130125. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Bhat, A.H.; Buang, A. Enhanced biosorption of transition metals by living Chlorella vulgaris immobilized in Ca-alginate beads. Environ. Technol. 2019, 40, 1793–1809. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Chang, H.W.; Kao, P.C.; Pan, J.L.; Chang, J.S. Biosorption of cadmium by CO2-fixing microalga Scenedesmus obliquus CNW-N. Bioresour. Technol. 2012, 105, 74–80. [Google Scholar] [CrossRef]
- Bohutskyi, P.; Chow, S.; Ketter, B.; Fung Shek, C.; Yacar, D.; Tang, Y.; Zivojnovich, M.; Betenbaugh, M.J.; Bouwer, E.J. Phytoremediation of agriculture runoff by filamentous algae poly-culture for biomethane production, and nutrient recovery for secondary cultivation of lipid generating microalgae. Bioresour. Technol. 2016, 222, 294–308. [Google Scholar] [CrossRef]
- Santaeufemia, S.; Torres, E.; Mera, R.; Abalde, J. Bioremediation of oxytetracycline in seawater by living and dead biomass of the microalga Phaeodactylum tricornutum. J. Hazard. Mater. 2016, 320, 315–325. [Google Scholar] [CrossRef]
- Dil, E.A.; Ghaedi, M.; Ghezelbash, G.R.; Asfaram, A. Multi-responses optimization of simultaneous biosorption of cationic dyes by live yeast Yarrowia lipolytica 70562 from binary solution: Application of first order derivative spectrophotometry. Ecotoxicol. Environ. Saf. 2017, 139, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, R.K.; Parhi, P.K.; Pandey, S.; Bindhani, B.K.; Thatoi, H.; Panda, C.R. Active and passive biosorption of Pb(II) using live and dead biomass of marine bacterium Bacillus xiamenensis PbRPSD202: Kinetics and isotherm studies. J. Environ. Manag. 2019, 247, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Hifney, A.F.; Zien-Elabdeen, A.; Adam, M.S.; Gomaa, M. Biosorption of ketoprofen and diclofenac by living cells of the green microalgae Chlorella sp. Environ. Sci. Pollut. Res. Int. 2021, 28, 69242–69252. [Google Scholar] [CrossRef] [PubMed]
- Santaeufemia, S.; Abalde, J.; Torres, E. Efficient removal of dyes from seawater using as biosorbent the dead and living biomass of the microalga Phaeodactylum tricornutum: Equilibrium and kinetics studies. J. Appl. Phycol. 2021, 33, 3071–3090. [Google Scholar] [CrossRef]
- Lyubimenko, R.; Busko, D.; Richards, B.S.; Schafer, A.I.; Turshatov, A. Efficient Photocatalytic Removal of Methylene Blue Using a Metalloporphyrin-Poly(vinylidene fluoride) Hybrid Membrane in a Flow-Through Reactor. ACS Appl. Mater. Interfaces 2019, 11, 31763–31776. [Google Scholar] [CrossRef]
- Zhang, D.; Dai, F.; Zhang, P.; An, Z.; Zhao, Y.; Chen, L. The photodegradation of methylene blue in water with PVDF/GO/ZnO composite membrane. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 96, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Moghazy, M. Activated biomass of the green microalga Chlamydomonas variabilis as an efficient biosorbent to remove methylene blue dye from aqueous solutions. Water SA 2019, 45, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.; Coimbra, R.N.; Escapa, C.; Figueiredo, S.A.; Freitas, O.M.; Otero, M. Green Microalgae Scenedesmus obliquus Utilization for the Adsorptive Removal of Nonsteroidal Anti-Inflammatory Drugs (NSAIDs) from Water Samples. Int. J. Environ. Res. Public Health 2020, 17, 3707. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, M.; Rahman, S.S.A.; Vedachalam, R.; Kumar, P.S.; Sabareesh, G.; Karuppiah, S. Ipomoea carnea: A novel biosorbent for the removal of methylene blue (MB) from aqueous dye solution: Kinetic, equilibrium and statistical approach. Int. J. Phytoremediation 2021, 23, 982–1000. [Google Scholar] [CrossRef] [PubMed]
- Moghazy, R.M.; Labena, A.; Husien, S. Eco-friendly complementary biosorption process of methylene blue using micro-sized dried biosorbents of two macro-algal species (Ulva fasciata and Sargassum dentifolium): Full factorial design, equilibrium, and kinetic studies. Int. J. Biol. Macromol. 2019, 134, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Xia, J.; Long, J. Biosorption of methylene blue by nonliving biomass of the brown macroalga Sargassum hemiphyllum. Water Sci. Technol. 2017, 76, 1574–1583. [Google Scholar] [CrossRef]
- Ozkan, A.; Berberoglu, H. Physico-chemical surface properties of microalgae. Colloids Surf. B Biointerfaces 2013, 112, 287–293. [Google Scholar] [CrossRef]
- Yagub, M.T.; Sen, T.K.; Afroze, S.; Ang, H.M. Dye and its removal from aqueous solution by adsorption: A review. Adv. Colloid Interface Sci. 2014, 209, 172–184. [Google Scholar] [CrossRef]
- Fathollahi, A.; Coupe, S.J.; El-Sheikh, A.H.; Sanudo-Fontaneda, L.A. The biosorption of mercury by permeable pavement biofilms in stormwater attenuation. Sci. Total Environ. 2020, 741, 140411. [Google Scholar] [CrossRef]
- Kocaman, S. Synthesis and cationic dye biosorption properties of a novel low-cost adsorbent: Coconut waste modified with acrylic and polyacrylic acids. Int. J. Phytoremediation 2020, 22, 551–566. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Witek-Krowiak, A. State of the art for the biosorption process—A review. Appl. Biochem. Biotechnol. 2013, 170, 1389–1416. [Google Scholar] [CrossRef] [Green Version]
- Hall, K.R.; Eagleton, L.C.; Acrivos, A.; Vermeulen, T. Pore- and Solid-Diffusion Kinetics in Fixed-Bed Adsorption under Constant-Pattern Conditions. Ind. Eng. Chem. Fundam. 1966, 5, 212–223. [Google Scholar] [CrossRef]
- Goldman, J.C.; Azov, Y.; Riley, C.B.; Dennett, M.R. The effect of pH in intensive microalgal cultures. I. Biomass regulation. J. Exp. Mar. Biol. Ecol. 1982, 57, 1–13. [Google Scholar] [CrossRef]
- Vadlamani, A.; Viamajala, S.; Pendyala, B.; Varanasi, S. Cultivation of Microalgae at Extreme Alkaline pH Conditions: A Novel Approach for Biofuel Production. ACS Sustain. Chem. Eng. 2017, 5, 7284–7294. [Google Scholar] [CrossRef]
- Raja, R.; Shanmugam, H.; Ganesan, V.; Carvalho, I.S. Biomass from Microalgae: An Overview. J. Oceanogr. Mar. Res. 2014, 2, 1–3. [Google Scholar] [CrossRef]
- Geada, P.; Moreira, C.; Silva, M.; Nunes, R.; Madureira, L.; Rocha, C.M.R.; Pereira, R.N.; Vicente, A.A.; Teixeira, J.A. Algal proteins: Production strategies and nutritional and functional properties. Bioresour. Technol. 2021, 332, 125125. [Google Scholar] [CrossRef]
- Molinuevo-Salces, B.; Riaño, B.; Hernández, D.; García-González, M.C. Microalgae and Wastewater Treatment: Advantages and Disadvantages. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Alam, M., Wang, Z., Eds.; Springer: Singapore, 2019; pp. 505–533. [Google Scholar]
- Torres, E. Biosorption: A Review of the Latest Advances. Processes 2020, 8, 1584. [Google Scholar] [CrossRef]
- Xiong, J.Q.; Kurade, M.B.; Jeon, B.H. Can Microalgae Remove Pharmaceutical Contaminants from Water? Trends Biotechnol. 2018, 36, 30–44. [Google Scholar] [CrossRef]
- Bouzikri, S.; Ouasfi, N.; Benzidia, N.; Salhi, A.; Bakkas, S.; Khamliche, L. Marine alga “Bifurcaria bifurcata”: Biosorption of Reactive Blue 19 and methylene blue from aqueous solutions. Environ. Sci. Pollut. Res. Int. 2020, 27, 33636–33648. [Google Scholar] [CrossRef]
- Tan, I.A.W.; Hameed, B.; Ahmad, A.L. Equilibrium and Kinetic Studies on Basic Dye Adsorption by Oil Palm Fibre Activated Carbon. Chem. Eng. J. 2007, 127, 111–119. [Google Scholar] [CrossRef]
- De Araujo, T.P.; Tavares, F.O.; Vareschini, D.T.; Barros, M.A.S.D. Biosorption mechanisms of cationic and anionic dyes in a low-cost residue from brewer’s spent grain. Environ. Technol. 2021, 42, 2925–2940. [Google Scholar] [CrossRef]
- Parlayici, S.; Pehlivan, E. Biosorption of methylene blue and malachite green on biodegradable magnetic Cortaderia selloana flower spikes: Modeling and equilibrium study. Int. J. Phytoremediation 2021, 23, 26–40. [Google Scholar] [CrossRef]
- Georgin, J.; Marques, B.S.; Peres, E.C.; Allasia, D.; Dotto, G.L. Biosorption of cationic dyes by Para chestnut husk (Bertholletia excelsa). Water Sci. Technol. 2018, 77, 1612–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabaraki, R.; Sadeghinejad, N. Biosorption of six basic and acidic dyes on brown alga Sargassum ilicifolium: Optimization, kinetic and isotherm studies. Water Sci. Technol. 2017, 75, 2631–2638. [Google Scholar] [CrossRef]
- Gurses, A.; Doğar, Ç.; Yalçin, M.; Açıkyıldız, M.; Bayrak, R.; Karaca, S. The Adsorption Kinetics of the Cationic Dye, Methylene Blue, onto Clay. J. Hazard. Mater. 2006, 131, 217–228. [Google Scholar] [CrossRef]
- Malarvizhi, R.; Ho, Y.-S. The influence of pH and the structure of the dye molecules on adsorption isotherm modeling using activated carbon. Desalination 2010, 264, 97–101. [Google Scholar] [CrossRef]
- Işikver, Y. Removal of some cationic dyes from aqueous solution by acrylamide- or 2-hydroxyethyl methacrylate-based copolymeric hydrogels. Fibers Polym. 2017, 18, 2070–2078. [Google Scholar] [CrossRef]
- Caparkaya, D.; Cavas, L. Biosorption of Methylene Blue by a Brown Alga Cystoseira barbatula Kutzing. Acta Chim. Slov. 2008, 55, 547–553. [Google Scholar]
- Gurusamy, A.; Juang, R.-S.; Lee, D.-J. Use of Cellulose-Based Wastes for Adsorption of Dyes from Aqueous Solutions. J. Hazard. Mater. 2002, 92, 263–274. [Google Scholar] [CrossRef]
- Bhattacharyya, K.; Sharma, A. Kinetics and Thermodynamics of Methylene Blue Adsorption on Neem (Azadirachta indica) Leaf Powder. Dye. Pigment. 2005, 65, 51–59. [Google Scholar] [CrossRef]
- Sharma, Y.C.; Upadhyay, S. Removal of a Cationic Dye from Wastewaters by Adsorption on Activated Carbon Developed from Coconut Coir. Energy Fuels 2009, 23, 2983–2988. [Google Scholar] [CrossRef]
- El-Sikaily, A.; Khaled, A.; El Nemr, A.; Abdelwahab, O. Removal of Methylene Blue from aqueous solution by marine green alga Ulva lactuca. J. Chem. Ecol. 2006, 22, 149–157. [Google Scholar] [CrossRef]
- Rehman, M.S.U.; Kim, I.; Han, J.-I. Adsorption of methylene blue dye from aqueous solution by sugar extracted spent rice biomass. Carbohydr. Polym. 2012, 90, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Ramamurthi, V.; Sivanesan, S. Modeling the Mechanism Involved during the Sorption of Methylene Blue onto Fly Ash. J. Colloid Interface Sci. 2005, 284, 14–21. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Dutta, B. On the adsorption and diffusion of Methylene Blue in glass fibers. J. Colloid Interface Sci. 2005, 286, 807–811. [Google Scholar] [CrossRef]
- Danouche, M.; El Ghachtouli, N.; El Arroussi, H. Phycoremediation mechanisms of heavy metals using living green microalgae: Physicochemical and molecular approaches for enhancing selectivity and removal capacity. Heliyon 2021, 7, e07609. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Chai, H.; Yu, Z.; Wu, X.; Liu, Y.; Shen, L.; Li, J.; Ye, J.; Liu, D.; Ma, T.; et al. Behavior and Mechanism of Cesium Biosorption from Aqueous Solution by Living Synechococcus PCC7002. Microorganisms 2020, 8, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetic Model | Differential Equation | Equation |
|---|---|---|
| Pseudo-first-order model | ) | |
| Pseudo-second-order model | ||
| Pseudo-third-order model | ||
| Pseudo-fourth-order model | ||
| Intraparticle diffusion model (Weber–Morris) | - |
| Isotherm Model | Equations |
|---|---|
| Langmuir | ) |
| Freundlich | |
| Temkin | |
| Dubinin–Radushkevich |
| Initial Dye Concentration (mg/L) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0.5 | 2.25 | 4.5 | 9 | 12 | 24 | 48 | 96 | 200 | 400 | |
| p (%) | 99.9 ± 0.2 | 92.4 ± 0.3 | 85.4 ± 0.6 | 83.8 ± 2.1 | 82.7 ± 0.9 | 84.2 ± 0.7 | 80.5 ± 1.2 | 79.6 ± 0.2 | 60.7 ± 0.7 | 38.7 ± 1.3 |
| Kinetic Model | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Initial Methylene Blue Concentration (mg/L) | Pseudo-First-Order | Pseudo-Second-Order | Pseudo-Third-Order | Pseudo-Fourth-Order | Intraparticle Diffusion | |||||
| r2 | AIC | r2 | AIC | r2 | AIC | r2 | AIC | r2 | AIC | |
| 0.5 | 0.965 | −74.48 | 0.992 | −93.29 | 0.996 | −101.48 | 0.994 | −95.63 | 0.421 | −40.55 |
| 2.25 | 0.957 | −35.97 | 0.992 | −57.29 | 0.994 | −60.14 | 0.989 | −53.19 | 0.564 | −8.16 |
| 4.5 | 0.955 | −20.85 | 0.993 | −44.91 | 0.997 | −55.11 | 0.993 | −44.86 | 0.590 | 5.88 |
| 9 | 0.953 | −4.96 | 0.990 | −24.39 | 0.996 | −36.99 | 0.994 | −31.58 | 0.557 | 22.03 |
| 12 | 0.954 | 2.419 | 0.983 | −9.73 | 0.986 | −12.11 | 0.983 | −9.58 | 0.627 | 27.58 |
| 24 | 0.954 | 19.22 | 0.989 | 1.12 | 0.993 | −3.30 | 0.989 | 1.613 | 0.600 | 45.36 |
| 48 | 0.940 | 39.94 | 0.983 | 24.42 | 0.991 | 17.60 | 0.989 | 18.28 | 0.727 | 58.11 |
| 96 | 0.955 | 53.51 | 0.991 | 34.20 | 0.996 | 23.69 | 0.995 | 25.66 | 0.767 | 73.26 |
| 200 | 0.953 | 64.47 | 0.989 | 46.41 | 0.995 | 37.60 | 0.994 | 39.19 | 0.755 | 84.48 |
| 400 | 0.941 | 72.34 | 0.985 | 55.98 | 0.993 | 47.55 | 0.992 | 48.16 | 0.721 | 91.15 |
| Kinetic Parameters (Pseudo-Third-Order) | |||
|---|---|---|---|
| Initial Methylene Blue Concentration (mg/L) | qe (mg/g) | k3 (g2/(mg2 h1)) | t1/2 (h) |
| 0.5 | 0.66 ± 0.02 | 130.12 ± 12.85 | 0.009 |
| 2.25 | 2.80 ± 0.03 | 2.54 ± 0.27 | 0.025 |
| 4.5 | 5.14 ± 0.04 | 0.70 ± 0.05 | 0.027 |
| 9 | 9.90 ± 0.08 | 0.26 ± 0.02 | 0.020 |
| 12 | 13.11 ± 0.25 | 0.09 ± 0.01 | 0.033 |
| 24 | 26.81 ± 0.35 | 0.02 ± 0.003 | 0.028 |
| 48 | 53.31 ± 0.99 | 0.003 ± 4 × 10−4 | 0.066 |
| 96 | 108.07 ± 1.41 | 4.7 × 10−4 ± 4 × 10−4 | 0.092 |
| 200 | 169.35 ± 2.42 | 2.1 × 10−4 ± 2 × 10−5 | 0.082 |
| 400 | 210.10 ± 3.37 | 1.9 × 10−4 ± 2 × 10−5 | 0.060 |
| Parameters | ||
|---|---|---|
| Initial Methylene Blue Concentration (mg/L) | ki (mg/(g h0.5)) | I (mg/g) |
| 0.5 | 0.14 ±0.05 | 0.36 ± 0.07 |
| 2.25 | 0.69± 0.18 | 1.20 ± 0.26 |
| 4.5 | 1.30 ± 0.32 | 2.14 ± 0.47 |
| 9 | 2.41 ± 0.63 | 4.47 ± 0.93 |
| 12 | 3.48 ± 0.80 | 5.03 ± 1.17 |
| 24 | 6.86 ± 1.65 | 10.93 ± 2.45 |
| 48 | 15.48 ± 2.81 | 15.41 ± 4.17 |
| 96 | 32.28 ± 5.29 | 26.56 ± 7.84 |
| 200 | 49.88 ± 8.44 | 44.60 ± 12.51 |
| 400 | 60.37 ± 11.14 | 63.59 ± 16.51 |
| Isotherm Model | Constants and Error Functions | Value |
|---|---|---|
| Langmuir | qmax (mg/g) | 212.41 ± 4.55 |
| KL (L/mg) | 0.04 ± 0.002 | |
| RL | 0.06 − 0.98 | |
| r2 | 0.997 | |
| AIC | 35.60 | |
| Freundlich | 1/n | 0.42 ± 0.05 |
| KF (mg1−(1/n) L1/n /g) | 20.41 ± 4.66 | |
| r2 | 0.950 | |
| AIC | 67.56 | |
| Temkin | AT (L/mg) | 18.84 ± 28.61 |
| bT (g J/(mg mol)) | 165.53 ± 49.72 | |
| r2 | 0.531 | |
| AIC | 90.84 | |
| D–R | qmax (mg/g) | 170.75 ± 11.64 |
| BD (mol2/J2) | 2.79 × 10−5 ± 7 × 10−6 | |
| ED (kJ/mol) | 0.13 | |
| r2 | 0.943 | |
| AIC | 67.61 |
| Materials | qmax† (mg/g) | KF†† (mg1−(1/n) L1/n/g) | Contact Time (h) | [Dye] (mg/L) | References |
|---|---|---|---|---|---|
| Bifurcaria bifurcata | 2744.5 | 189.8 | 0.25 | 10–1000 | [39] |
| Fucus vesiculosus | 698.48 | 225.3 | 24 | 100–2500 | [8] |
| Oil palm shell carbon | 384.62 | 132.28 | 30 | 50–500 | [40] |
| Brewer’s spent grain | 298.35 | 69.51 | 7 | 5–250 | [41] |
| Brazilian berry seeds (Eugenia uniflora) | 189.6 | 34.4 | 3 | 25–200 | [6] |
| Magnetic Cortaderia selloana flower spikes | 119.05 | 1.41 | 0.5 | 25–350 | [42] |
| Chestnut husk | 117.2 | 19.4 | 0.67 | 50–500 | [43] |
| Chlamydomonas variabilis activated by H2SO4 | 115 | 68.5 | 0.5 | 20–80 | [21] |
| Sargassum ilicifolium | 99.7 | - | 0.67 | 1.28–38 | [44] |
| Cyanthilium cinereum | 76.34 | 10.07 | 0.83 | 10–50 | [7] |
| Clay | 58.20 | - | 2 | 10–100 | [45] |
| Paspalum maritimum | 56.18 | 13.08 | 0.83 | 10–50 | [7] |
| Wood apple rind carbon | 40.1 | 21.3 | 2 | 10–100 | [46] |
| Hydrogel P(AAm-co-AcA) | 39.59 | - | 24 | 5–50 | [47] |
| Ipomoea carnea | 39.38 | 3.96 | 2.7 | 10–50 | [23] |
| Cystoseira barbatula | 38.61 | 81.8 | 6 | 5–100 | [48] |
| Banana peel | 20.80 | 1.34 | 24 | 10–120 | [49] |
| Neem leaf powder | 19.61 | 9.47 | 5 | 25–70 | [50] |
| Orange peel | 18.60 | 1.75 | 24 | 10–120 | [49] |
| Chlamydomonas variabilis (dead biomass) | 18.3 | 1.26 | 0.5 | 20–80 | [21] |
| Coconut coir | 15.59 | 0.98 | 2.33 | 60–100 | [51] |
| Ulva lactuca | 10.99 | 1.45 | 2 | 5–25 | [52] |
| Spent rice biomass | 8.3 | - | 2 | 25–50 | [53] |
| Fly ash | 5.57 | 4.38 | 2 | 20–60 | [54] |
| Glass fibres | 2.24 | 2.12 | 6 | 25–50 | [55] |
| C. moewussi (living, unmodified) | 212.41 | 20.41 | 7 | 0.5–400 | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seoane, R.; Santaeufemia, S.; Abalde, J.; Torres, E. Efficient Removal of Methylene Blue Using Living Biomass of the Microalga Chlamydomonas moewusii: Kinetics and Equilibrium Studies. Int. J. Environ. Res. Public Health 2022, 19, 2653. https://doi.org/10.3390/ijerph19052653
Seoane R, Santaeufemia S, Abalde J, Torres E. Efficient Removal of Methylene Blue Using Living Biomass of the Microalga Chlamydomonas moewusii: Kinetics and Equilibrium Studies. International Journal of Environmental Research and Public Health. 2022; 19(5):2653. https://doi.org/10.3390/ijerph19052653
Chicago/Turabian StyleSeoane, Raquel, Sergio Santaeufemia, Julio Abalde, and Enrique Torres. 2022. "Efficient Removal of Methylene Blue Using Living Biomass of the Microalga Chlamydomonas moewusii: Kinetics and Equilibrium Studies" International Journal of Environmental Research and Public Health 19, no. 5: 2653. https://doi.org/10.3390/ijerph19052653
APA StyleSeoane, R., Santaeufemia, S., Abalde, J., & Torres, E. (2022). Efficient Removal of Methylene Blue Using Living Biomass of the Microalga Chlamydomonas moewusii: Kinetics and Equilibrium Studies. International Journal of Environmental Research and Public Health, 19(5), 2653. https://doi.org/10.3390/ijerph19052653