
Effectiveness Evaluation of a UV-C-Photoinactivator against Selected ESKAPE-E Pathogens


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. UV-C System
2.3. Exposure to the UV-C System in a Liquid Medium
2.3.1. Cell Viability
2.3.2. Quantification of Colonies
2.4. Exposure to the UV-C System in Solid Medium
2.5. Statistical Analysis
2.6. Scanning Electron Microscopy (SEM)
3. Results
3.1. Exposure to the UV-C System in a Liquid Medium
3.1.1. Cell Viability
3.1.2. Quantification of Colonies
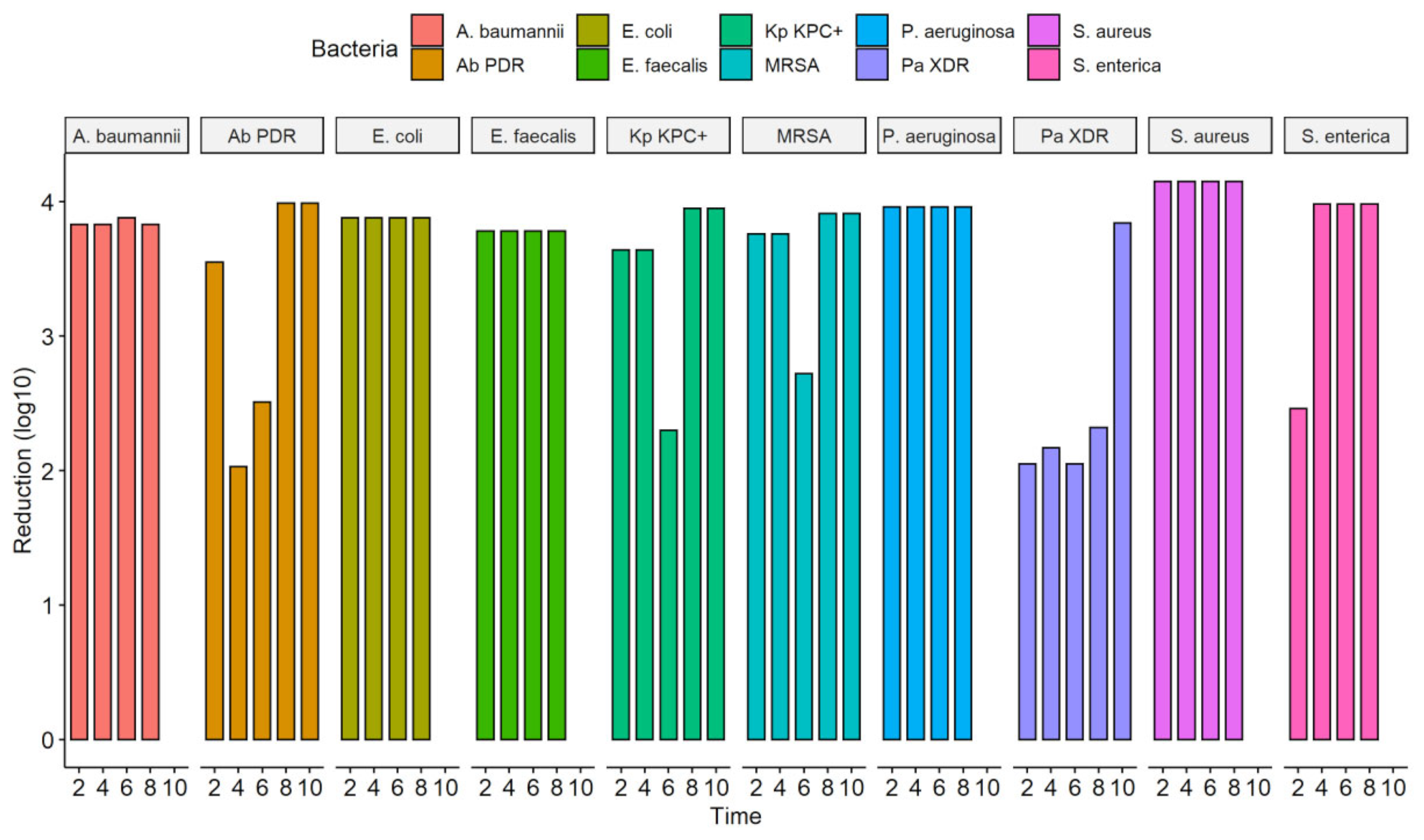
3.2. Exposure to the UV-C System in Solid Medium
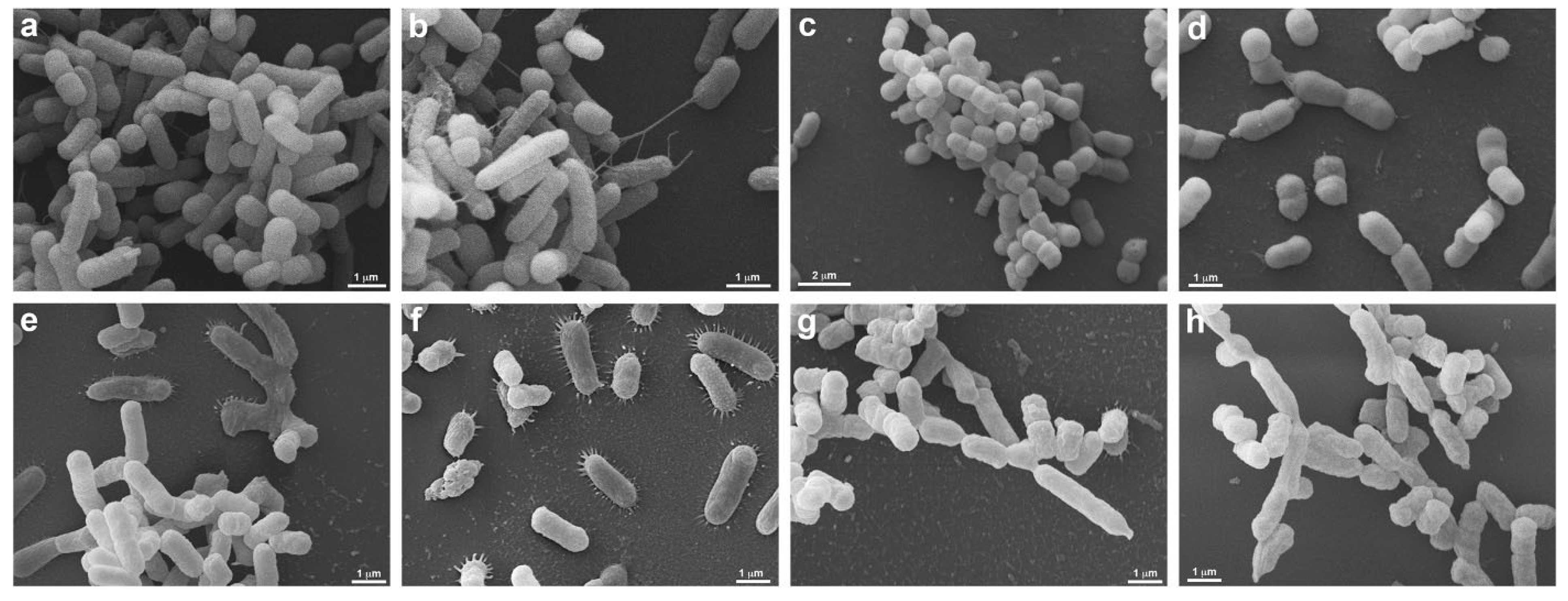
3.3. Scanning Electron Microscopy (SEM)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tabah, A.; Koulenti, D.; Laupland, K.; Misset, B.; Valles, J.; de Carvalho, F.B.; Paiva, J.A.; Çakar, N.; Ma, X.; Eggimann, P.; et al. Characteristics and determinants of outcome of hospital-acquired bloodstream infections in intensive care units: The EUROBACT International Cohort Study. Intensiv. Care Med. 2012, 38, 1930–1945. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Eisenberg, J.N.S.; Spicknall, I.H.; Koopman, J.S. Dynamics and Control of Infections Transmitted from Person to Person Through the Environment. Am. J. Epidemiol. 2009, 170, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Rashid, T.; VonVille, H.M.; Hasan, I.; Garey, K.W. Shoe soles as a potential vector for pathogen transmission: A systematic review. J. Appl. Microbiol. 2016, 121, 1223–1231. [Google Scholar] [CrossRef]
- Agarwal, M.; Hamilton-Stewart, P.; Dixon, R.A. Contaminated operating room boots: The potential for infection. Am. J. Infect. Control 2002, 30, 179–183. [Google Scholar] [CrossRef][Green Version]
- Alam, M.J.; Anu, A.; Walk, S.T.; Garey, K.W. Investigation of potentially pathogenic Clostridium difficile contamination in household environs. Anaerobe 2014, 27, 31–33. [Google Scholar] [CrossRef]
- Amirfeyz, R.; Tasker, A.; Ali, S.; Bowker, K.; Blom, A. Theatre Shoes—A Link in the Common Pathway of Postoperative Wound Infection? Ann. R. Coll. Surg. Engl. 2007, 89, 605–608. [Google Scholar] [CrossRef]
- Paduszyńska, K.; Rucińska, L.G.M.; Pomorski, L. Physician as an Infective Vector at a Department of Surgery. Pol. J. Surg. 2015, 86, 511–517. [Google Scholar] [CrossRef]
- Kanwar, A.; Thakur, M.; Wazzan, M.; Satyavada, S.; Cadnum, J.L.; Jencson, A.L.; Donskey, C.J. Clothing and shoes of personnel as potential vectors for transfer of health care–associated pathogens to the community. Am. J. Infect. Control 2019, 47, 577–579. [Google Scholar] [CrossRef]
- Hambraeus, A.; Bengtsson, S.; Laurell, G. Bacterial contamination in a modern operating suite. 3. Importance of floor contamination as a source of airborne bacteria. Epidemiol. Infect. 1978, 80, 169–174. [Google Scholar] [CrossRef]
- Alexander, J.W.; Van Sweringen, H.; VanOss, K.; Hooker, E.A.; Edwards, M.J. Surveillance of Bacterial Colonization in Operating Rooms. Surg. Infect. 2013, 14, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Feldman, O.; Meir, M.; Shavit, D.; Idelman, R.; Shavit, I. Exposure to a Surrogate Measure of Contamination from Simulated Patients by Emergency Department Personnel Wearing Personal Protective Equipment. JAMA 2020, 323, 2091–2093. [Google Scholar] [CrossRef] [PubMed]
- Lipner, S. Head to Toe: Recommendations for Physician Head and Shoe Coverings to Limit COVID-19 Transmission. Cutis 2021, 107, E27–E29. [Google Scholar] [CrossRef]
- Guo, Z.-D.; Wang, Z.-Y.; Zhang, S.-F.; Li, X.; Li, L.; Li, C.; Cui, Y.; Fu, R.-B.; Dong, Y.-Z.; Chi, X.-Y.; et al. Aerosol and Surface Distribution of Severe Acute Respiratory Syndrome Coronavirus 2 in Hospital Wards, Wuhan, China, 2020. Emerg. Infect. Dis. 2020, 26, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Redmond, S.N.; Dousa, K.M.; Jones, L.D.; Li, D.F.; Cadnum, J.L.; Navas, M.E.; Kachaluba, N.M.; Silva, S.Y.; Zabarsky, T.F.; Eckstein, E.C.; et al. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) nucleic acid contamination of surfaces on a coronavirus disease 2019 (COVID-19) ward and intensive care unit. Infect. Control Hosp. Epidemiol. 2020, 42, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Laube, H.; Friese, A.; von Salviati, C.; Guerra, B.; Rösler, U. Transmission of ESBL/AmpC-producing Escherichia coli from broiler chicken farms to surrounding areas. Veter- Microbiol. 2014, 172, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.E.; Bergh, I.; Karlsson, J.; Eriksson, B.I.; Nilsson, K. Traffic flow in the operating room, an explorative and de-scriptive study on air quality during orthopedic trauma implant surgery. Am. J. Infect Control. 2012, 40, 750e5. [Google Scholar] [CrossRef]
- Limmathurotsakul, D.; Kanoksil, M.; Wuthiekanun, V.; Kitphati, R.; DeStavola, B.; Day, N.P.J.; Peacock, S.J. Activities of Daily Living Associated with Acquisition of Melioidosis in Northeast Thailand: A Matched Case-Control Study. PLOS Neglected Trop. Dis. 2013, 7, e2072. [Google Scholar] [CrossRef]
- Gwaltney, J.J.M.; Hendley, J.O. Transmission of experimental rhinovirus infection by contaminated surfaces. Am. J. Epidemiol. 1982, 116, 828–833. [Google Scholar] [CrossRef]
- Büchler, A.C.; Wicki, M.; Frei, R.; Hinic, V.; Seth-Smith, H.M.B.; Egli, A.; Widmer, A.F. Matching Clostridioides difficile strains obtained from shoe soles of healthcare workers epidemiologically linked to patients and confirmed by whole-genome sequencing. J. Hosp. Infect. 2022, 126, 10–15. [Google Scholar] [CrossRef]
- Janezic, S.; Mlakar, S.; Rupnik, M. Dissemination of Clostridium difficiles pores between environment and households: Dog paws and shoes. Zoonoses Public Health 2018, 65, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Janezic, S.; Blazevic, I.; Eyre, D.W.; Kevorkijan, B.K.; Remec, T.; Rupnik, M. Possible contribution of shoes to Clostridioides difficile transmission within hospitals. Clin. Microbiol. Infect. 2020, 27, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Amass, S.F.; Arighi, M.; Kinyon, J.M.; Hoffman, L.J.; Schneider, J.L.; Draper, D.K. Effectiveness of using a mat filled with a peroxygen disinfectant to minimize shoe sole contamination in a veterinary hospital. J. Am. Veter- Med. Assoc. 2006, 228, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Ayliffe, G.A.; Collins, B.J.; Lowbury, E.J. Cleaning and disinfection of hospital floors. BMJ 1966, 2, 442–445. [Google Scholar] [CrossRef][Green Version]
- Curry, C.; McCarthy, J.; Darragh, H.; Wake, R.; Todhunter, R.; Terris, J. Could tourist boots act as vectors for disease transmission in Antarctica? J. Travel Med. 2006, 9, 190–193. [Google Scholar] [CrossRef]
- Eisenberg, T.; Wolter, W.; Lenz, M.; Schlez, K.; Zschöck, M. Boot swabs to collect environmental samples from common locations in dairy herds for Mycobacterium avium ssp. Paratuberculosis (MAP) detection. J. Dairy Res. 2013, 80, 485–489. [Google Scholar] [CrossRef]
- Chambers, M.K.; Ford, M.R.; White, D.M.; Barnes, D.; Schiewer, S. Transport of fecal bacteria by boots and vehicle tires in a rural Alaskan community. J. Environ. Manag. 2009, 90, 961–966. [Google Scholar] [CrossRef]
- Mangram, A.J.; Horan, T.C.; Pearson, M.L.; Silver, L.C.; Jarvis, W.R. Guideline for Prevention of Surgical Site Infection, 1999. Centers for Disease Control and Prevention (CDC) Hospital Infection Control Practices Advisory Committee. Am. J. Infect. Control. 1999, 27, 97–132; quiz 133–134; discussion 196. [Google Scholar] [CrossRef]
- Galvin, J.; Almatroudi, A.; Vickery, K.; Deva, A.; Lopes, L.K.O.; Costa, D.D.M.; Hu, H. Patient shoe covers: Transferring bacteria from the floor onto surgical bedsheets. Am. J. Infect. Control 2016, 44, 1417–1419. [Google Scholar] [CrossRef]
- Rashid, T.; Poblete, K.; Amadio, J.; Hasan, I.; Begum, K.; Alam, M.J.; Garey, K.W. Evaluation of a shoe sole UVC device to reduce pathogen colonization on floors, surfaces and patients. J. Hosp. Infect. 2018, 98, 96–101. [Google Scholar] [CrossRef]
- Hijnen, W.A.M.; Beerendonk, E.F.; Medema, G.J. Inactivation credit of UV radiation for viruses, bacteria and protozoan (oo)cysts in water: A review. Water Res. 2006, 40, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Mohseni, M.; Taghipour, F. Application of ultraviolet light-emitting diodes (UV-LEDs) for water disinfection: A review. Water Res. 2016, 94, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Oppenländer, T. Photochemical Purification of Water and Air; Wiley-VCH: Weinheim, Germany, 2007; 368p. [Google Scholar]
- Phillips, R. Sources and Applications of Ultraviolet Radiation; Academic Press Inc.: New York, NY, USA, 1983. [Google Scholar]
- Morita, S.; Namikoshi, A.; Hirata, T.; Oguma, K.; Katayama, H.; Ohgaki, S.; Motoyama, N.; Fujiwara, M. Efficacy of UV Irradiation in Inactivating Cryptosporidium parvum Oocysts. Appl. Environ. Microbiol. 2002, 68, 5387–5393. [Google Scholar] [CrossRef] [PubMed]
- Kalisvaart, B. Re-use of wastewater: Preventing the recovery of pathogens by using medium-pressure UV lamp technology. Water Sci. Technol. 2004, 50, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Ploydaeng, M.; Rajatanavin, N.; Rattanakaemakorn, P. UV-C light: A powerful technique for inactivating microorganisms and the related side effects to the skin. Photodermatol. Photoimmunol. Photomed. 2020, 37, 12–19. [Google Scholar] [CrossRef]
- Smolle, C.; Huss, F.; Lindblad, M.; Reischies, F.; Tano, E. Effectiveness of automated ultraviolet-C light for decontamination of textiles inoculated with Enterococcus faecium. J. Hosp. Infect. 2018, 98, 102–104. [Google Scholar] [CrossRef]
- Havill, N.L.; Moore, B.; Boyce, J.M. Comparison of the Microbiological Efficacy of Hydrogen Peroxide Vapor and Ultraviolet Light Processes for Room Decontamination. Infect. Control Hosp. Epidemiol. 2012, 33, 507–512. [Google Scholar] [CrossRef]
- Lall, N.; Henley-Smith, C.J.; De Canha, M.N.; Oosthuizen, C.B.; Berrington, D. Viability Reagent, PrestoBlue, in Comparison with Other Available Reagents, Utilized in Cytotoxicity and Antimicrobial Assays. Int. J. Microbiol. 2013, 2013, 420601. [Google Scholar] [CrossRef]
- Rangel, K.; Cabral, F.O.; Lechuga, G.C.; Carvalho, J.P.R.S.; Villas-Bôas, M.H.S.; Midlej, V.; De-Simone, S.G. Potent Activity of a High Concentration of Chemical Ozone against Antibiotic-Resistant Bacteria. Molecules 2022, 27, 3998. [Google Scholar] [CrossRef]
- Otter, J.A.; Yezli, S.; Salkeld, J.A.; French, G.L. Evidence that contaminated surfaces contribute to the transmission of hospital pathogens and an overview of strategies to address contaminated surfaces in hospital settings. Am. J. Infect. Control 2013, 41, S6–S11. [Google Scholar] [CrossRef]
- Nyangaresi, P.O.; Qin, Y.; Chen, G.; Zhang, B.; Lu, Y.; Shen, L. Effects of single and combined UV-LEDs on inactivation and subsequent reactivation of E. coli in water disinfection. Water Res. 2018, 147, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Taghipour, F.; Mohseni, M. Microorganisms inactivation by wavelength combinations of ultraviolet light-emitting diodes (UV-LEDs). Sci. Total. Environ. 2019, 665, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Jelden, K.C.; Gibbs, S.; Smith, P.W.; Hewlett, A.L.; Iwen, P.C.; Schmid, K.K.; Lowe, J.J. Ultraviolet (UV)-reflective paint with ultraviolet germicidal irradiation (UVGI) improves decontamination of nosocomial bacteria on hospital room surfaces. J. Occup. Environ. Hyg. 2017, 14, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Fattorini, M.; Nante, N.; Rosadini, D.; Serafini, A.; Tani, M.; Cevenini, G. Time Effectiveness of Ultraviolet C Light (UVC) Emitted by Light Emitting Diodes (LEDs) in Reducing Stethoscope Contamination. Int. J. Environ. Res. Public Health 2016, 13, 940. [Google Scholar] [CrossRef]
- Messina, G.; Rosadini, D.; Burgassi, S.; Messina, D.; Nante, N.; Tani, M.; Cevenini, G. Tanning the bugs—A pilot study of an innovative approach to stethoscope disinfection. J. Hosp. Infect. 2016, 95, 228–230. [Google Scholar] [CrossRef]
- Betzalel, Y.; Gerchman, Y.; Cohen-Yaniv, V.; Mamane, H. Multiwell plates for obtaining a rapid microbial dose-response curve in UV-LED systems. J. Photochem. Photobiol. B Biol. 2020, 207, 111865. [Google Scholar] [CrossRef]
- Bowker, C.; Sain, A.; Shatalov, M.; Ducoste, J. Microbial UV fluence-response assessment using a novel UV-LED collimated beam system. Water Res. 2011, 45, 2011–2019. [Google Scholar] [CrossRef]
- Sheikh, J.; Swee, T.T.; Saidin, S.; Bin Yahya, A.; Malik, S.A.; Yin, J.S.S.; Thye, M.T.F. Bacterial disinfection and cell assessment post ultraviolet-C LED exposure for wound treatment. Med. Biol. Eng. Comput. 2021, 59, 1055–1063. [Google Scholar] [CrossRef]
- McMahon, M.A.S.; Xu, J.; Moore, J.E.; Blair, I.S.; McDowell, D.A. Environmental Stress and Antibiotic Resistance in Food-Related Pathogens. Appl. Environ. Microbiol. 2007, 73, 211–217. [Google Scholar] [CrossRef]
- Alekshun, M.N.; Levy, S.B. Alteration of the Repressor Activity of MarR, the Negative Regulator of the Escherichia coli marRAB Locus, by Multiple Chemicals In Vitro. J. Bacteriol. 1999, 181, 4669–4672. [Google Scholar] [CrossRef]
- Rickard, A.H.; Lindsay, S.; Lockwood, G.B.; Gilbert, P. Induction of the mar operon by miscellaneous groceries. J. Appl. Microbiol. 2004, 97, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Marles-Wright, J.; Grant, T.; Delumeau, O.; van Duinen, G.; Firbank, S.J.; Lewis, P.J.; Murray, J.W.; Newman, J.A.; Quin, M.B.; Race, P.R.; et al. Molecular Architecture of the “Stressosome,” a Signal Integration and Transduction Hub. Science 2008, 322, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Sirsat, S.; Burkholder, K.M.; Muthaiyan, A.; Dowd, S.; Bhunia, A.; Ricke, S.C. Effect of sublethal heat stress on Salmonella Typhimurium virulence. J. Appl. Microbiol. 2011, 110, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.S.; Saxena, G.K.; Kataria, J.M. Study on Heat Stress Response in Salmonella typhimurium and Salmonella enteritidis and its Impact on their Attachment to Dressed Broiler Skin Surface. Asian J. Anim. Veter- Adv. 2016, 11, 114–121. [Google Scholar] [CrossRef][Green Version]
- Nerandzic, M.M.; Cadnum, J.L.; Pultz, M.J.; Donskey, C.J. Evaluation of an automated ultraviolet radiation device for decontamination of Clostridium difficile and other healthcare-associated pathogens in hospital rooms. BMC Infect. Dis. 2010, 10, 197. [Google Scholar] [CrossRef]
- Nerandzic, M.M.; Fisher, C.W.; Donskey, C.J. Sorting through the Wealth of Options: Comparative Evaluation of Two Ultraviolet Disinfection Systems. PLoS ONE 2014, 9, e107444. [Google Scholar] [CrossRef]
- Rutala, W.A.; Gergen, M.F.; Weber, D.J. Room Decontamination with UV Radiation. Infect. Control Hosp. Epidemiol. 2010, 31, 1025–1029. [Google Scholar] [CrossRef]
- Mahida, N.; Vaughan, N.; Boswell, T. First UK evaluation of an automated ultraviolet-C room decontamination device (Tru-D™). J. Hosp. Infect. 2013, 84, 332–335. [Google Scholar] [CrossRef]
- Toté, K.; Berghe, D.V.; Levecque, S.; Bénéré, E.; Maes, L.; Cos, P. Evaluation of hydrogen peroxide-based disinfectants in a new resazurin microplate method for rapid efficacy testing of biocides. J. Appl. Microbiol. 2009, 107, 606–615. [Google Scholar] [CrossRef]
- Gayán, E.; Monfort, S.; Álvarez, I.; Condón, S. UV-C inactivation of Escherichia coli at different temperatures. Innov. Food Sci. Emerg. Technol. 2011, 12, 531–541. [Google Scholar] [CrossRef]
- Giese, N.; Darby, J. Sensitivity of microorganisms to different wavelengths of UV light: Implications on modeling of medium pressure UV systems. Water Res. 2000, 34, 4007–4013. [Google Scholar] [CrossRef]
- Phillips, M.; Von Reyn, C.F. Nosocomial Infections Due to Nontuberculous Mycobacteria. Clin. Infect. Dis. 2001, 33, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Kheyrandish, A.; Mohseni, M.; Taghipour, F. Protocol for Determining Ultraviolet Light Emitting Diode (UV-LED) Fluence for Microbial Inactivation Studies. Environ. Sci. Technol. 2018, 52, 7390–7398. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, I.; Guettari, M.; Chroudi, A.; Touati, H.; Hamza, S. Disinfection Technology in Hospitals: Harmful effects of UVC. Tunis Med. 2020, 98, 434–441. [Google Scholar] [PubMed]
- Setlow, J.K.; Duggan, D.E. The resistance of Micrococcus radiodurans to ultraviolet radiation: I. Ultraviolet-induced lesions in the cell’s DNA. Biochim. et Biophys. Acta (BBA) Spéc. Sect. Nucleic Acids Relat. Subj. 1964, 87, 664–668. [Google Scholar] [CrossRef]
- Proft, T.; Baker, E. Pili in Gram-negative and Gram-positive bacteria—Structure, assembly and their role in disease. Cell. Mol. Life Sci. 2008, 66, 613–635. [Google Scholar] [CrossRef]
- Cennamo, P.; Ebbreo, M.; Quarta, G.; Trojsi, G.; De Rosa, A.; Carfagna, S.; Caputo, P.; Castaldi, M.M. UV-C Irradiation as a Tool to Reduce Biofilm Growth on Pompeii Wall Paintings. Int. J. Environ. Res. Public Health 2020, 17, 8392. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Times of Exposure to UV-C * | |||||
|---|---|---|---|---|---|---|
| UV-C (10″) | C (10″) | UV-C (12″) | C (12″) | UV-C (20″) | C (20″) | |
| RFU/% Reduction | Average | RFU/% Reduction | Average | RFU/% Reduction | Average | |
| S. aureus | 730.6/50.2 * | 1465.8 | 1852.5/8.8 | 2030.6 | 334.1/71.9 * | 1190.4 |
| P. aeruginosa | 647.1/42.9 * | 1132.8 | 104.8/93.4 * | 1584.3 | 96.5 /90.9 * | 1059.7 |
| S. enterica | 482.3/11.6 | 545.3 | 630.1/3.3 | 651.3 | 93.2 /82.9 | 547.2 |
| E. coli | 654.8/41.2 * | 1112.7 | 379.2/50.5 * | 766.2 | 94.5 /79.2 * | 453.2 |
| A. baumannii | 595.1/60.1 * | 1489.7 | 1273.2/18.7 | 1565.6 | 132.1 /85.3 * | 900.52 |
| E. faecalis | 867.3/44.9 * | 1573.6 | 175.3/83.2 * | 1040.9 | 94.9/85.3 * | 646.7 |
| Bacterial (Strains) | UV-C System Exposure Times | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2″ | C | 4″ | C | 6″ | C | 8″ | C | 10″ | C | 12″ | C | 20″ | C | |
| CFU/mL Count/% Red/CFU/mL Count | CFU/mL Count/% Red/CFU/mL Count | CFU/mL Count/% Red/CFU/mL Count | CFU/mL Count/% Red/CFU/mL Count | CFU/mL Count/% Red/CFU/mL Count | CFU/mL Count/% Red/CFU/mL Count | CFU/mL Count/% Red/CFU/mL Count | ||||||||
| S. aureus | 2.710/30.3 | 3.888 | 1.044/73.4 | 3.932 | 332/9.9 | 4.088 | 154/96,3 | 4.176 | 110/97.7 | 4.844 | 0/100 | 3.954 | 0/100 | 4.022 |
| P. eruginosa | 1.044/67.1 | 3.176 | 310/91.8 | 3.776 | 44/98.4 | 2.688 | 88/96.6 | 2600 | 22/99.4 | 3.732 | 0/100 | 2.000 | 0/100 | 2.710 |
| S. enterica | 1.400/63.6 | 3.844 | 1.244/74.7 | 4.910 | 488/86.1 | 3.510 | 288/90.6 | 3.066 | 88/97.6 | 3.688 | 0/100 | 3.222 | 0/100 | 3.710 |
| E. coli | 1.666/26.5 | 2.266 | 732/85 | 4.888 | 332/87.1 | 2.576 | 666/79.2 | 3.200 | 288/92.6 | 3.910 | 0/100 | 3.400 | 0/100 | 3.532 |
| A. baumannii | 2.422/27.3 | 3.332 | 1.376/52.7 | 2.910 | 774/71.9 | 2.754 | 1132/69.1 | 3.666 | 466/89.6 | 4.488 | 88/97.7 | 3.866 | 0/100 | 3.800 |
| E. faecalis | 1.622/39.7 | 2.688 | 776/69.9 | 2.576 | 400/86.0 | 2.866 | 1132/69.7 | 3.732 | 754/76.9 | 3.266 | 0/100 | 4.354 | 0/100 | 3.900 |
| Bacterial Strains | UV-C System Exposure Times | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2″ | C | 4″ | C | 6″ | C | 8″ | C | 10″ | C | |
| CFU/mL Count/% Reduction/ CFU/mL Count | CFU/mL Count/% Reduction/ CFU/mL Count | CFU/mL Count/% Reduction/ CFU/mL Count | CFU/mL Count/% Reduction/ CFU/mL Count | CFU/mL Count/% Reduction/ CFU/mL Count | ||||||
| S. aureus | 0/100 | 14.200 | 0/100 | 14.200 | 0/100 | 14.200 | 0/100 | 14.200 | NP | NP |
| P. aeruginosa | 0/100 | 9.033 | 0/100 | 9.033 | 0/100 | 9.033 | 0/100 | 9.033 | NP | NP |
| S. enterica | 33/99.7 | 9.567 | 0/100 | 9.567 | 0/100 | 9.567 | 0/100 | 9.567 | NP | NP |
| E. coli | 0/100 | 7600 | 0/100 | 7.600 | 0/100 | 7.600 | 0/100 | 7.600 | NP | NP |
| A. baumannii | 0/100 | 6.833 | 0/100 | 6.833 | 0/100 | 6.833 | 0/100 | 6.833 | NP | NP |
| E. faecalis | 0/100 | 5967 | 0/100 | 5.967 | 0/100 | 5.967 | 0/100 | 5.967 | NP | NP |
| S. aureus (MRSA) | 0/100 | 5.711 | 0/100 | 5.711 | 11/99.8 | 5.711 | 0/100 | 8.183 | 0/100 | 8.183 |
| P. aeruginosa (XDR) | 44/99.1 | 4.911 | 33/99.3 | 4.911 | 44/ 99.1 | 4.911 | 33/99.5 | 6.983 | 0/100 | 6.983 |
| A. baumannii (PDR) | 0/100 | 3.556 | 33/99.1 | 3.556 | 11/ 99.7 | 3.556 | 0/100 | 9.817 | 0/100 | 9.817 |
| K. pneumoniae KPC+ | 0/100 | 4.356 | 0/100 | 4.356 | 22/ 99.5 | 4.356 | 0/100 | 8.917 | 0/100 | 8.917 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangel, K.; Cabral, F.O.; Lechuga, G.C.; Villas-Bôas, M.H.S.; Midlej, V.; De-Simone, S.G. Effectiveness Evaluation of a UV-C-Photoinactivator against Selected ESKAPE-E Pathogens. Int. J. Environ. Res. Public Health 2022, 19, 16559. https://doi.org/10.3390/ijerph192416559
Rangel K, Cabral FO, Lechuga GC, Villas-Bôas MHS, Midlej V, De-Simone SG. Effectiveness Evaluation of a UV-C-Photoinactivator against Selected ESKAPE-E Pathogens. International Journal of Environmental Research and Public Health. 2022; 19(24):16559. https://doi.org/10.3390/ijerph192416559
Chicago/Turabian StyleRangel, Karyne, Fellipe O. Cabral, Guilherme C. Lechuga, Maria H. S. Villas-Bôas, Victor Midlej, and Salvatore G. De-Simone. 2022. "Effectiveness Evaluation of a UV-C-Photoinactivator against Selected ESKAPE-E Pathogens" International Journal of Environmental Research and Public Health 19, no. 24: 16559. https://doi.org/10.3390/ijerph192416559
APA StyleRangel, K., Cabral, F. O., Lechuga, G. C., Villas-Bôas, M. H. S., Midlej, V., & De-Simone, S. G. (2022). Effectiveness Evaluation of a UV-C-Photoinactivator against Selected ESKAPE-E Pathogens. International Journal of Environmental Research and Public Health, 19(24), 16559. https://doi.org/10.3390/ijerph192416559