
Deeply in Plasticenta: Presence of Microplastics in the Intracellular Compartment of Human Placentas

 , , , , ,
, , , , ,  ,
,  ,
,  ,
, 
Abstract
:Highlights
- For the first time, microplastics were detected and localized by electron microscopy in human placentas.
- The presence of microplastics was correlated with ultrastructural alterations of some cell organelles in placental tissue, mainly in the syncytiotrophoblast.
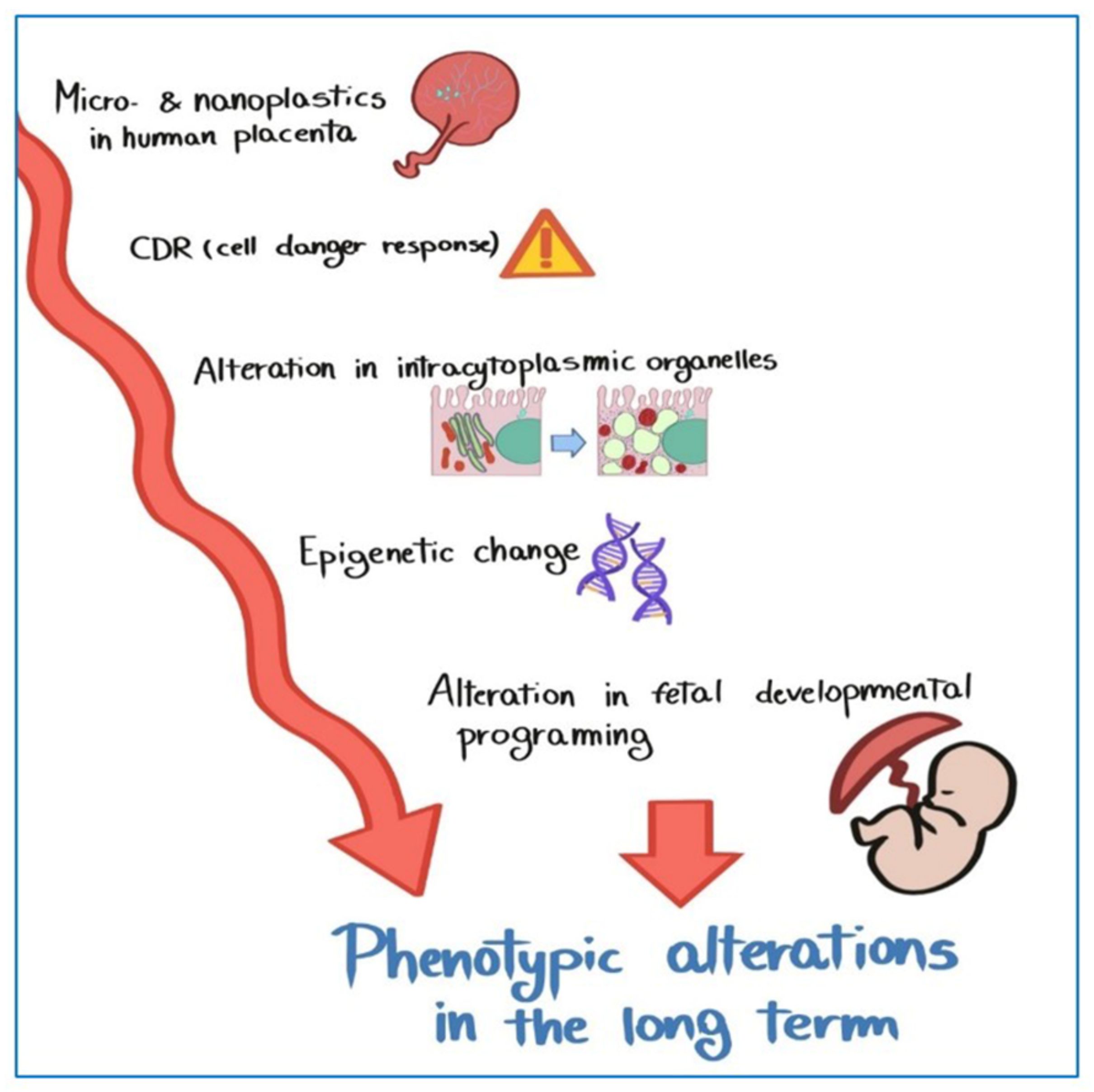
- Microplastics in human placentas could contribute to the activation of pathological traits, such as oxidat ive stress, apoptosis, and inflammation.
- Microplastics in human placentas may cause long-term effects on human health.
Abstract
1. Introduction
- To locate MPs within the intra/extracellular compartment in human placenta by variable pressure scanning electron microscopy (VP-SEM) and transmission electron microscopy (TEM).
- To understand whether their presence and location are associated with possible structural changes of cell organelles.
2. Materials and Methods
2.1. Experimental Design, Enrolment of Patients and Placentas Collection
- -
- Particular diets prescribed by a physician in the four weeks prior childbirth.
- -
- Diarrhea or constipation in the two weeks prior to childbirth.
- -
- Antibiotic use in the two weeks prior childbirth.
- -
- Drug use interfering with intestinal absorption (e.g., active carbon or cholestyramine) in the two weeks prior to childbirth.
- -
- Diagnosis of a gastrointestinal disease (e.g., ulcerative colitis or Crohn disease, cancer, organ transplant, HIV or any other disease that requires medical treatment).
- -
- Invasive or abrasive odontoiatric treatments in the two weeks prior to childbirth.
- -
- Current or recent participation (within the four weeks prior childbirth) to a clinical experiment.
- -
- Alcohol abuse (defined by a test score for identification of alcohol abuse disorders > 10).
2.2. Electron Microscopy Analysis
2.2.1. Variable Pressure Scanning Electron Microscope
2.2.2. Transmission Electron Microscopy
3. Results and Discussion
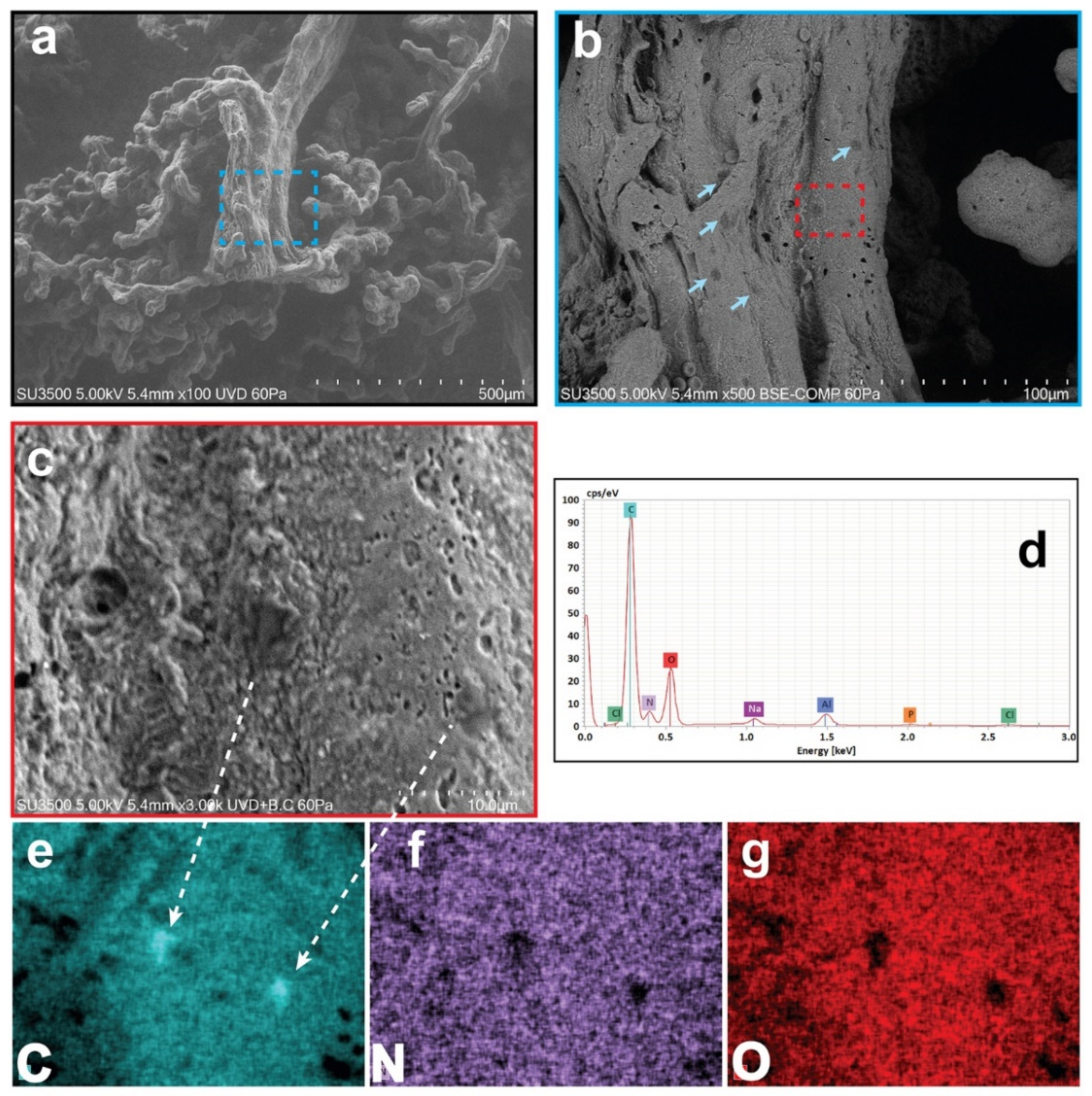
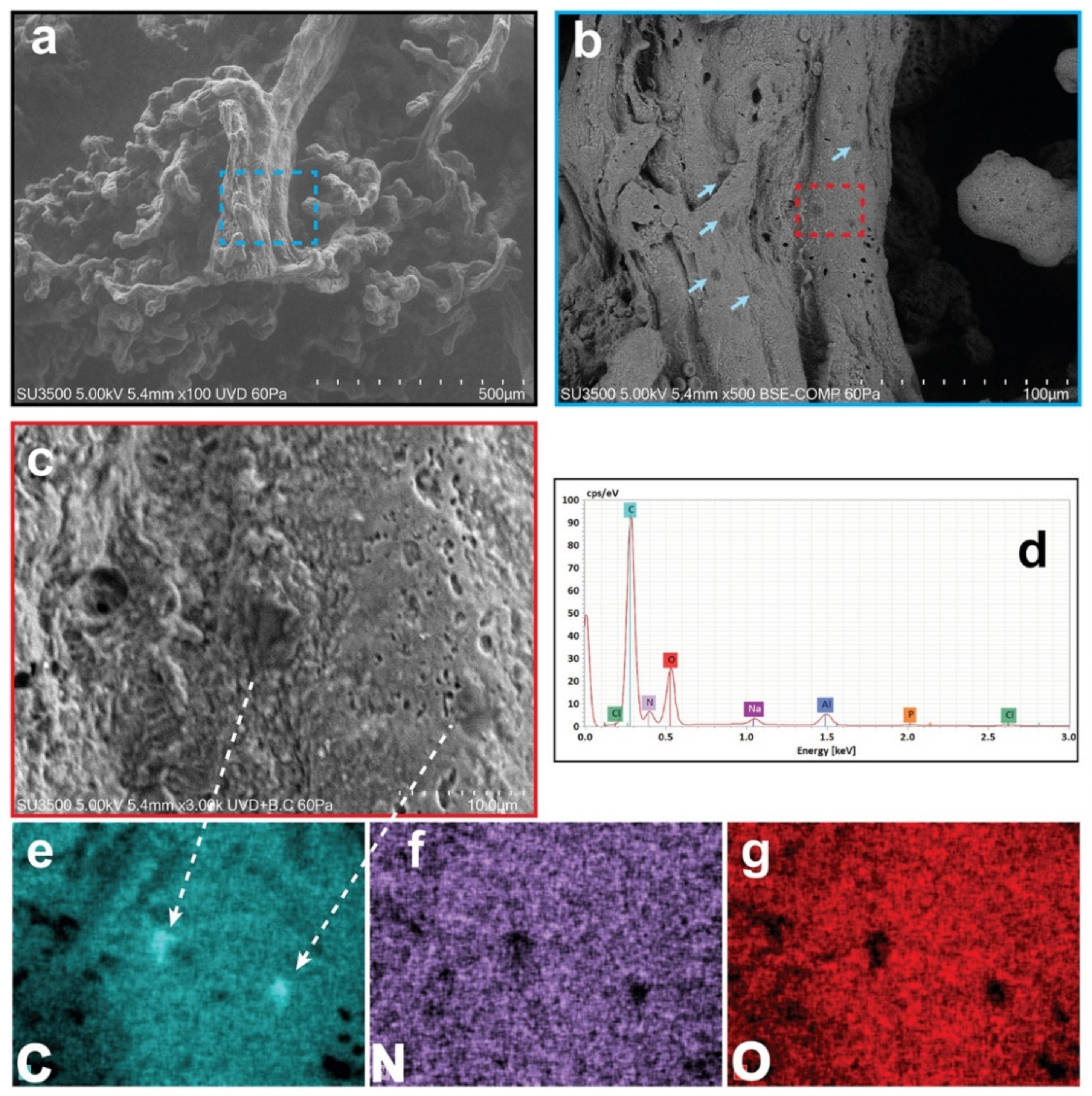
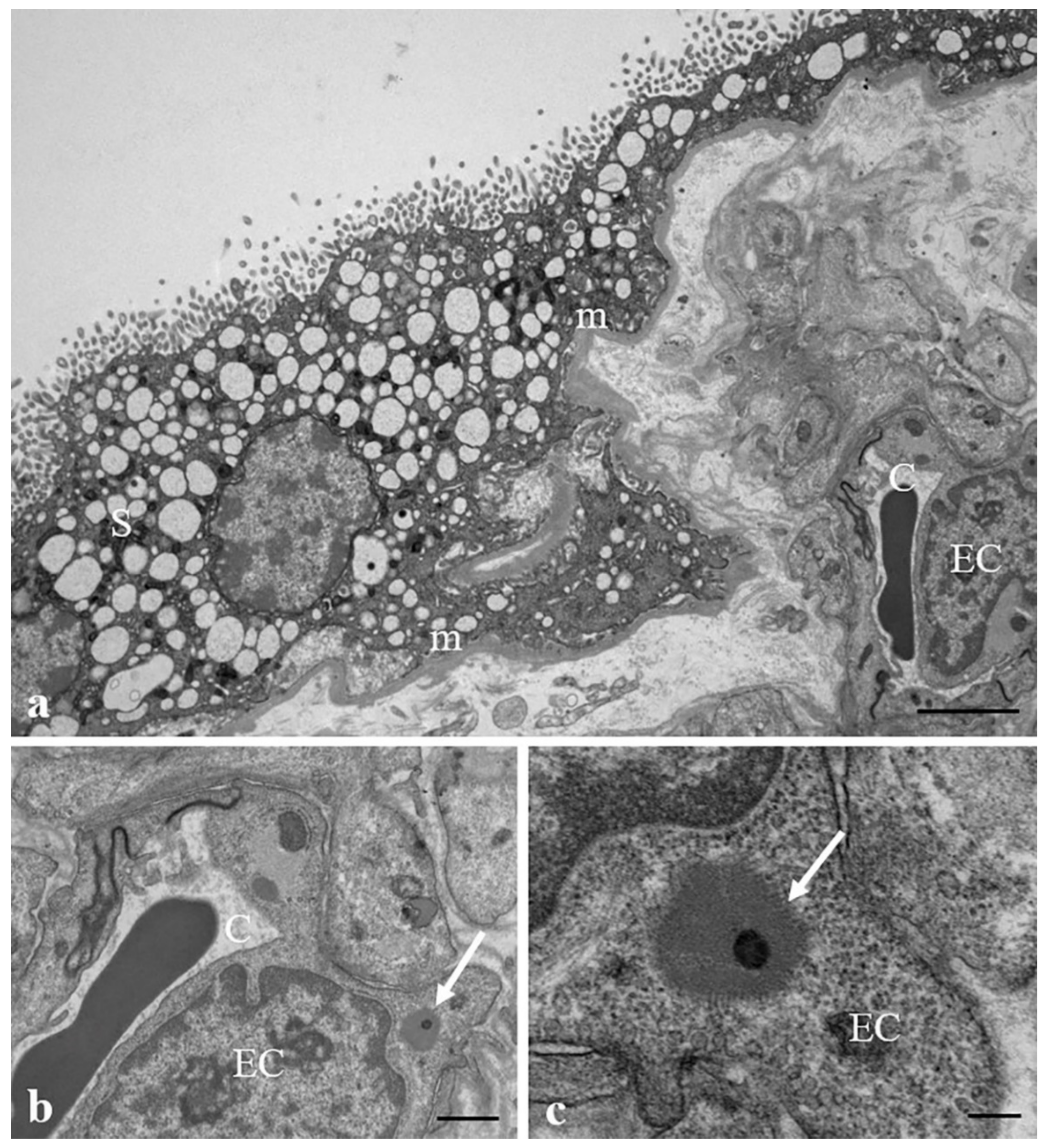
3.1. Results: Morphological and Qualitative Analyses by VP-SEM-dEDS
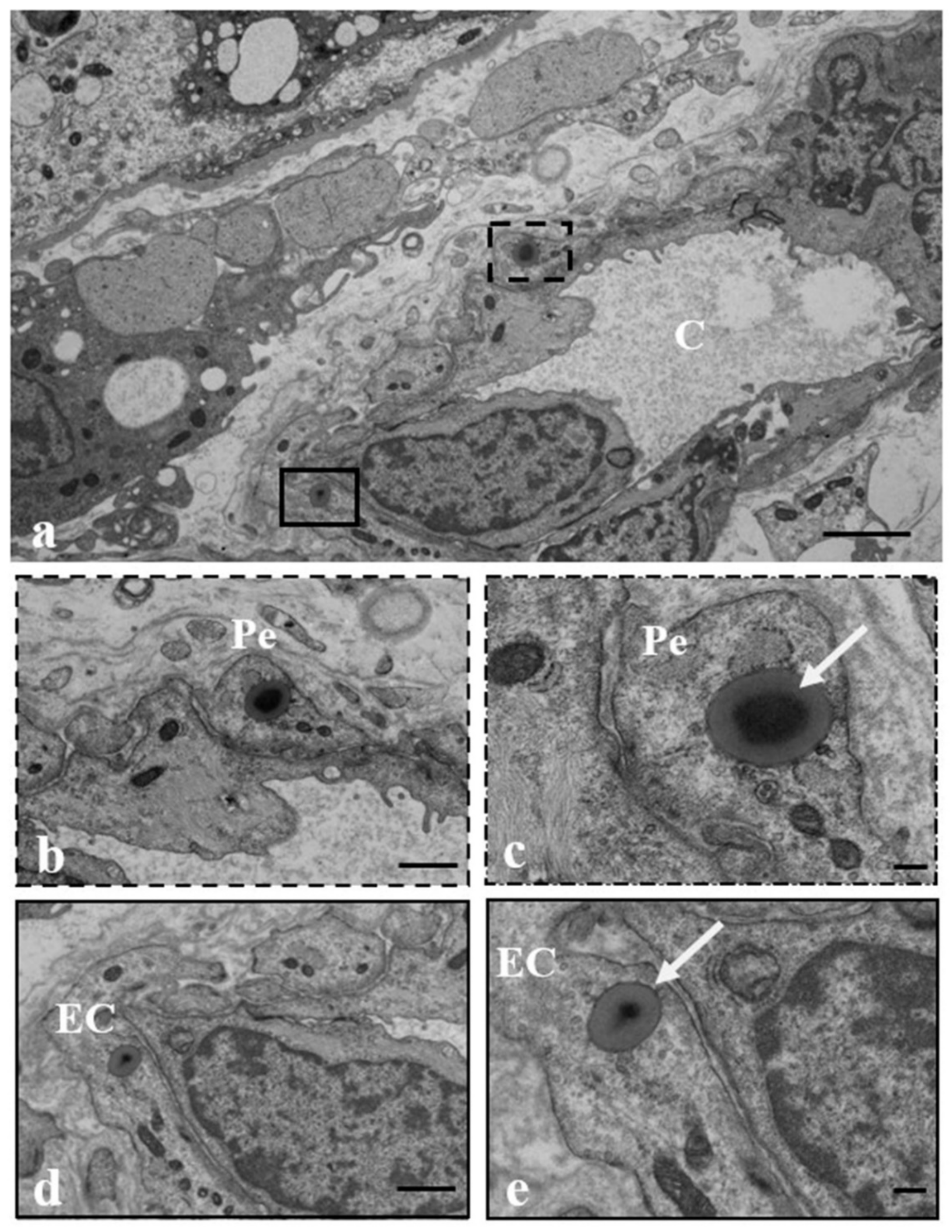
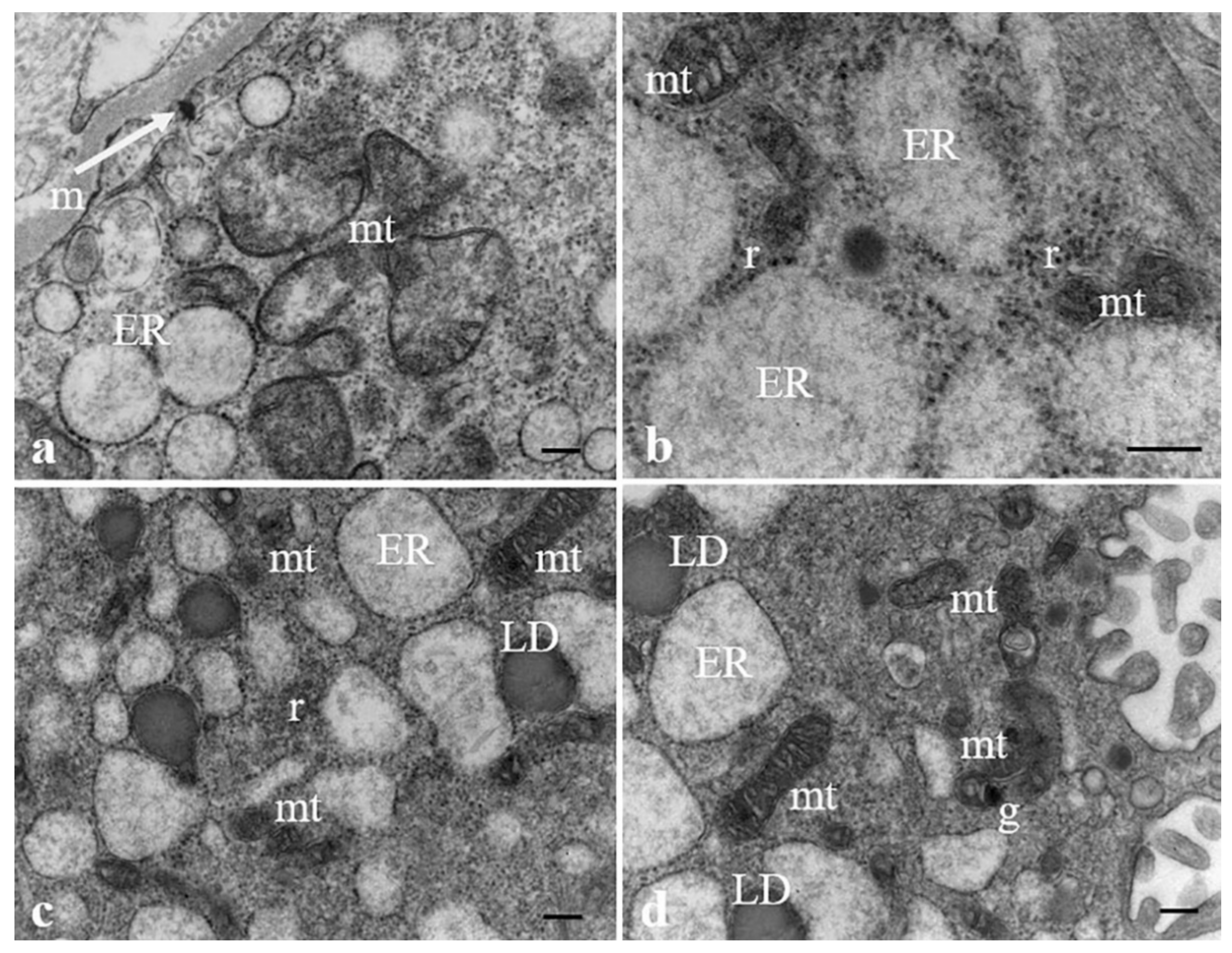
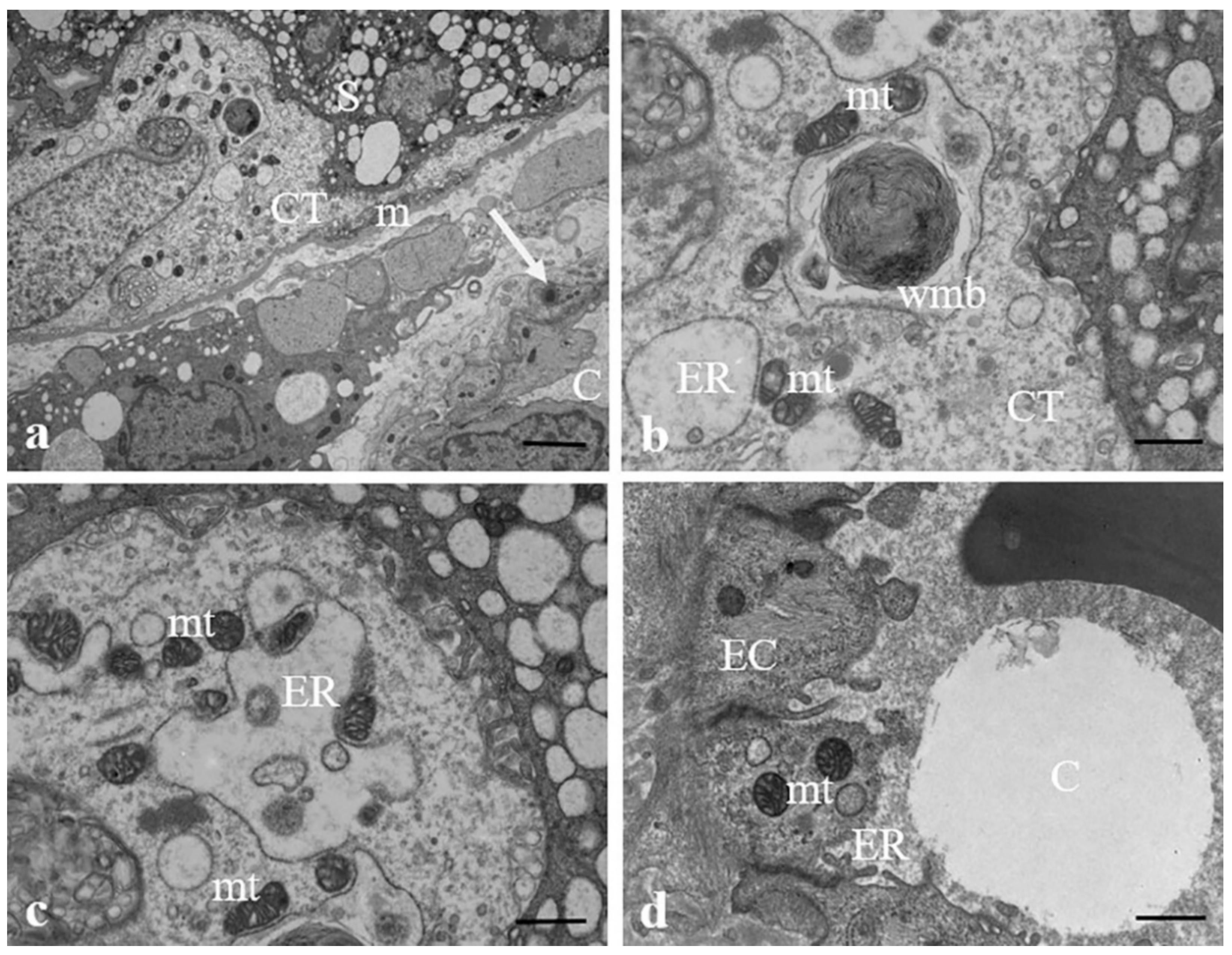
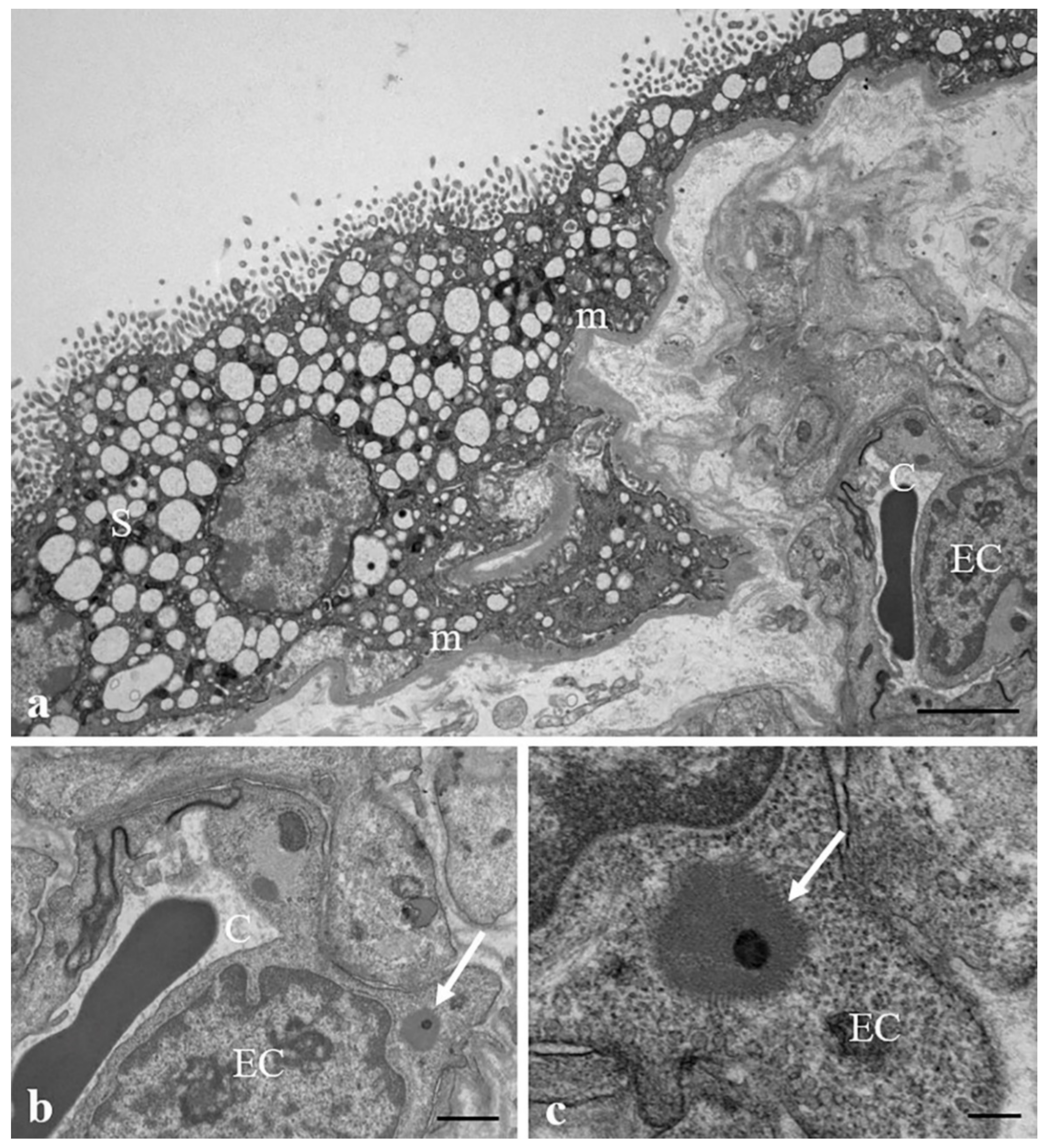
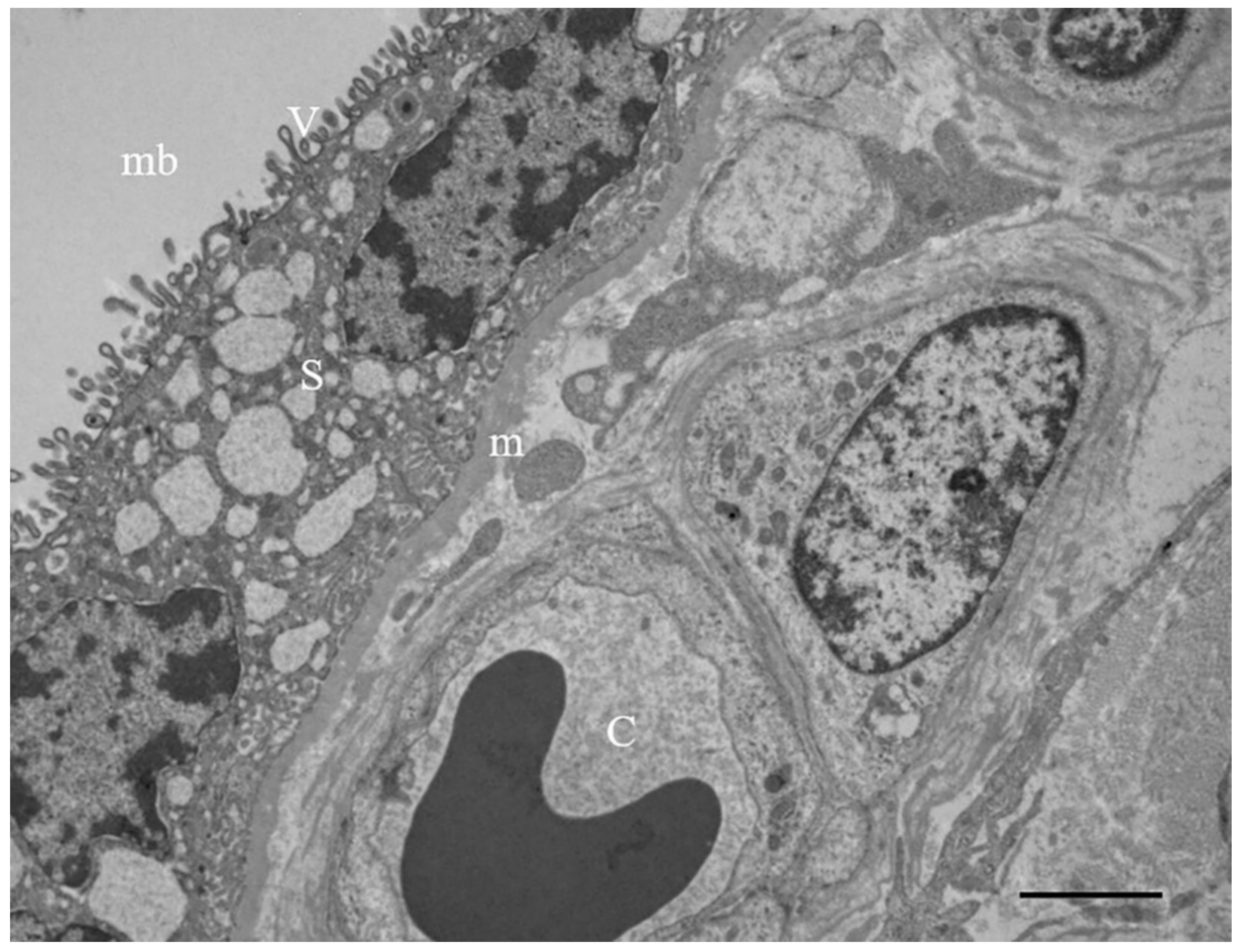
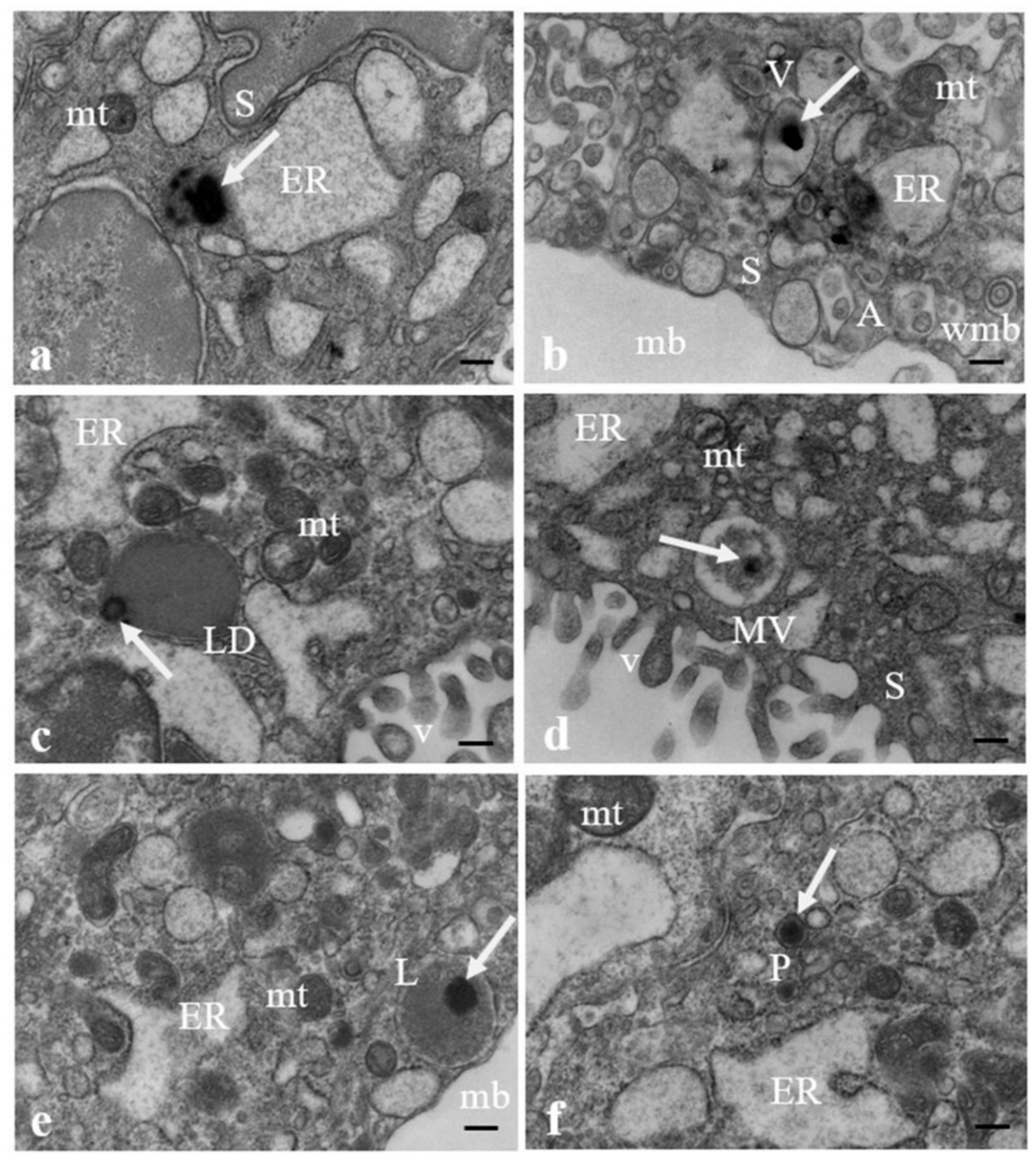
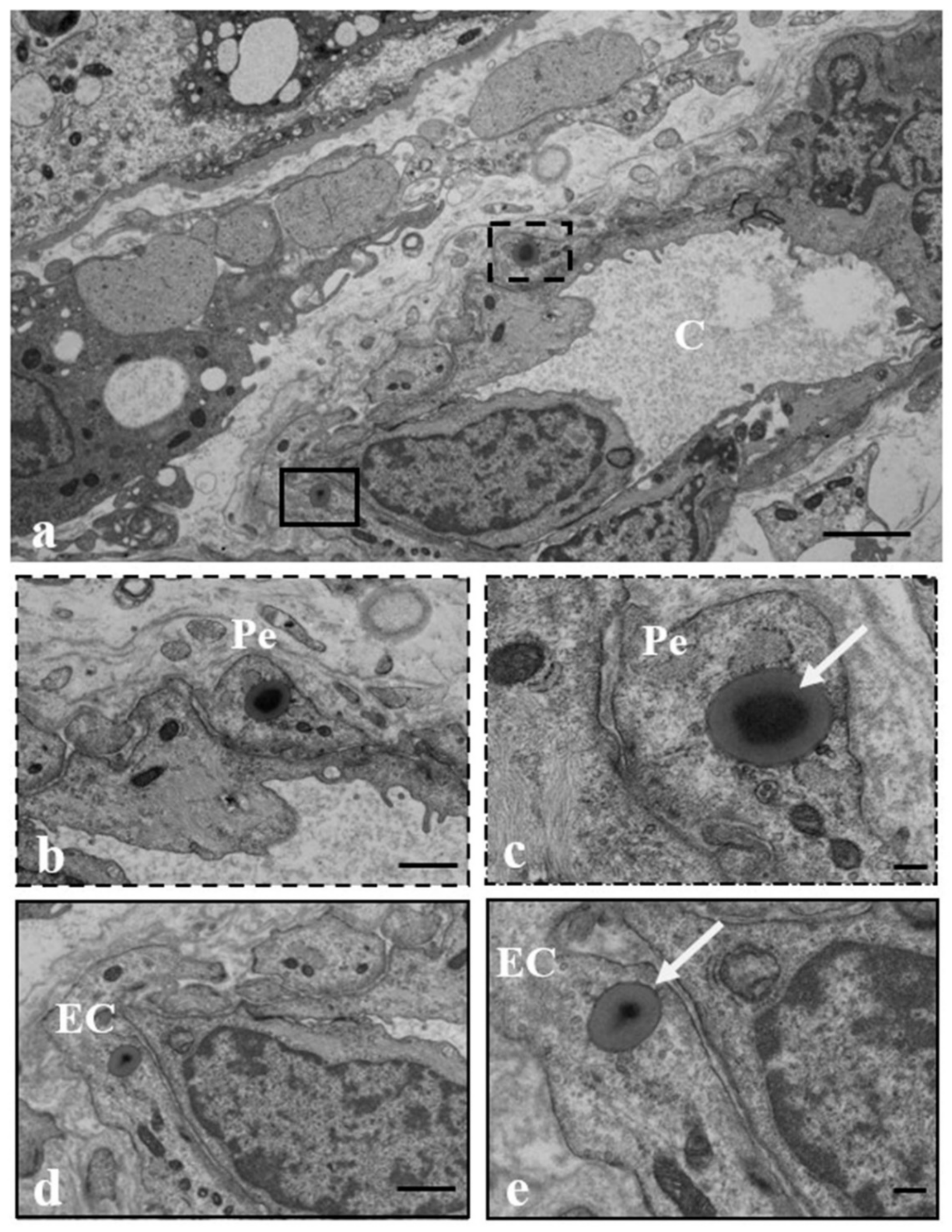
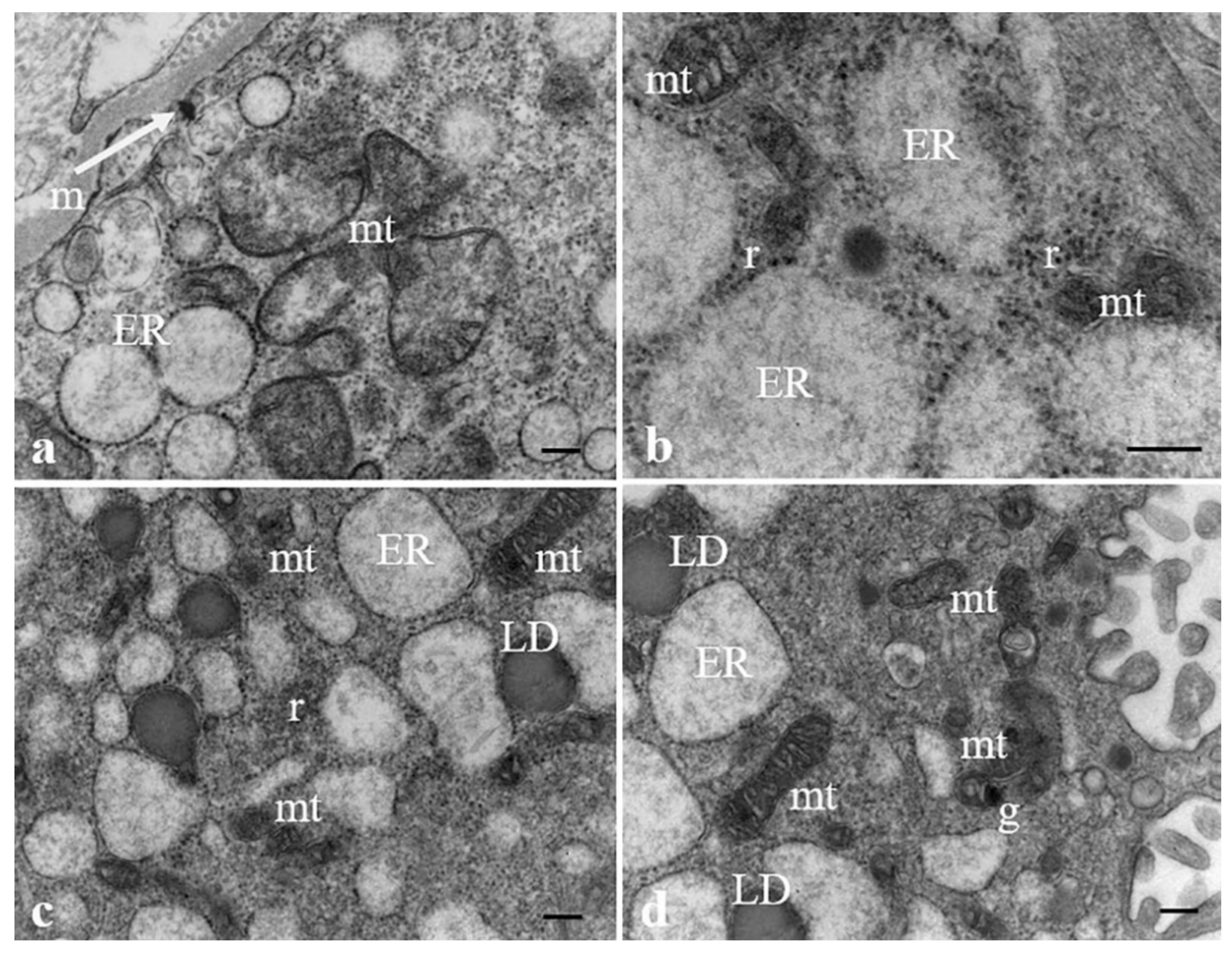
3.2. Results: Observations by TEM
3.3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kershaw, P.J.; Rochman, C.M. (Eds.) Sources, Fate and Effects of Microplastics in the Marine Environment: Part Two of a Global Assessment; GESAMP Report and Studies No. 93, 220; International Maritime Organization: London, UK, 2016; 220p. [Google Scholar]
- EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain). Statement on the presence of microplastics and nanoplastics in food, with particular focus on seafood. EFSA J. 2016, 14, 4501. [Google Scholar] [CrossRef]
- Hartmann, N.B.; Hüffer, T.; Thompson, R.C.; Hassellöv, M.; Verschoor, A.; Daugaard, A.E.; Rist, S.; Karlsson, T.; Brennholt, N.; Cole, M.; et al. Are We Speaking the Same Language? Recommendations for a Definition and Categorization Framework for Plastic Debris. Environ. Sci. Technol. 2019, 53, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Tanentzap, A.J.; Cottingham, S.; Fonvielle, J.; Riley, I.; Walker, L.M.; Woodman, S.G.; Kontou, D.; Pichler, C.M.; Reisner, E.; Lebreton, L. Microplastics and anthropogenic fibre concentrations in lakes reflect surrounding land use. PLoS Biol. 2021, 19, e3001389. [Google Scholar] [CrossRef] [PubMed]
- De Souza Machado, A.A.; Kloas, W.; Zarfl, C.; Hempel, S.; Rillig, M.C. Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 1405–1416. [Google Scholar] [CrossRef]
- Prata, J.C. Airborne microplastics: Consequences to human health? Environ. Pollut. 2018, 234, 115–126. [Google Scholar] [CrossRef]
- Barboza, L.G.A.; Dick Vethaak, A.; Lavorante, B.R.B.O.; Lundebye, A.K.; Guilhermino, L. Marine microplastic debris: An emerging issue for food security, food safety and human health. Mar. Pollut. Bull. 2018, 133, 336–348. [Google Scholar] [CrossRef]
- Cho, Y.; Shim, W.J.; Jang, M.; Han, G.M.; Hong, S.H. Abundance and characteristics of microplastics in market bivalves from South Korea. Environ. Pollut. 2019, 245, 1107–1116. [Google Scholar] [CrossRef]
- Karami, A.; Golieskardi, A.; Choo, C.K.; Larat, V.; Galloway, T.S.; Salamatinia, B. The presence of microplastics in commercial salts from different countries. Sci. Rep. 2017, 7, 46173. [Google Scholar] [CrossRef]
- Van Cauwenberghe, L.; Janssen, C.R. Microplastics in bivalves cultured for human consumption. Environ. Pollut. 2014, 193, 65–70. [Google Scholar] [CrossRef]
- Kosuth, M.; Mason, S.A.; Wattenberg, E.V. Anthropogenic contamination of tap water, beer, and sea salt. PLoS ONE 2018, 13, e0194970. [Google Scholar] [CrossRef]
- Mason, S.A.; Welch, V.G.; Neratko, J. Synthetic Polymer Contamination in Bottled Water. Front. Chem. 2018, 6, 407. [Google Scholar] [CrossRef]
- Schymanski, D.; Goldbeck, C.; Humpf, H.-U.; Fürst, P. Analysis of microplastics in water by micro-Raman spectroscopy: Release of plastic particles from different packaging into mineral water. Water Res. 2018, 129, 154–162. [Google Scholar] [CrossRef]
- De Sá, L.C.; Oliveira, M.; Ribeiro, F.; Rocha, T.L.; Futter, M.N. Studies of the effects of microplastics on aquatic organisms: What do we know and where should we focus our efforts in the future? Sci. Total Environ. 2018, 645, 1029–1039. [Google Scholar] [CrossRef]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Lemos, B.; Ren, H. Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci. Rep. 2017, 7, 46687. [Google Scholar] [CrossRef]
- Reineke, J.J.; Cho, D.Y.; Dingle, Y.T.; Morello, A.P.; Jacobb, J.; Thanos, C.G.; Mathiowitz, E. Unique insights into the intestinal absorption, transit, and subsequent biodistribution of polymer-derived microspheres. Proc. Natl. Acad. Sci. USA 2013, 110, 13803–13808. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hoh, E.; Kurobe, T.; Teh, S.J. Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci. Rep. 2013, 3, 3263. [Google Scholar] [CrossRef]
- Wright, S.L.; Kelly, F.J. Plastic and Human Health: A Micro Issue? Environ. Sci. Technol. 2017, 51, 6634–6647. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef]
- Leslie, H.A.; van Velzen, M.J.M.; Brandsma, S.H.; Vethaak, A.D.; Garcia-Vallejo, J.J.; Lamoree, M.H. Discovery and quantification of plastic particle pollution in human blood. Environ. Int. 2022, 163, 107199. [Google Scholar] [CrossRef]
- Ragusa, A.; Notarstefano, V.; Svelato, A.; Belloni, A.; Gioacchini, G.; Blondeel, C.; Zucchelli, E.; De Luca, C.; D’Avino, S.; Gulotta, A.; et al. Raman Microspectroscopy Detection and Characterisation of Microplastics in Human Breastmilk. Polymers 2022, 14, 2700. [Google Scholar] [CrossRef]
- Braun, T.; Ehrlich, L.; Henrich, W.; Koeppel, S.; Lomako, I.; Schwabl, P.; Liebmann, B. Detection of microplastic in human placenta and meconium in a clinical setting. Pharmaceutics 2021, 13, 921. [Google Scholar] [CrossRef]
- Warner, K.; Linske, E.; Mustain, P.; Valliant, M.; Leavitt, C. Choked, Strangled, Drowned: The Plastics Crisis Unfolding in Our Oceans; Oceana Protecting the World’s Oceans; Oceana: Washington, DC, USA, 2020. [Google Scholar] [CrossRef]
- Rezania, S.; Parka, J.; Md Dinb, M.F.; Mat Taibb, S.; Talaiekhozanic, A.; Yadavd, K.K.; Kamyabe, H. Microplastics pollution in different aquatic environments and biota: A review of recent studies. Mar. Pollut. Bull. 2018, 133, 191–208. [Google Scholar] [CrossRef]
- Huang, D.; Chen, H.; Shen, M.; Tao, J.; Chen, S.; Yin, L.; Zhou, W.; Wang, X.; Ruihao Xiao, R.; Li, R. Recent advances on the transport of microplastics/nanoplastics in abiotic and biotic compartments. J. Haz. Mat. 2022, 438, 129515. [Google Scholar] [CrossRef]
- Yee, M.S.-L.; Hii, L.-W.; Looi, C.K.; Lim, W.-M.; Wong, S.-F.; Kok, Y.-Y.; Tan, B.-K.; Wong, C.-Y.; Leong, C.-O. Impact of Microplastics and Nanoplastics on Human Health. Nanomaterials 2021, 11, 496. [Google Scholar] [CrossRef]
- Campanale, C.; Massarelli, C.; Savino, I.; Locaputo, V.; Uricchio, V.F. A Detailed Review Study on Potential Effects of Microplastics and Additives of Concern on Human Health. Int. J. Environ. Res. Public Health 2020, 17, 1212. [Google Scholar] [CrossRef]
- DeLoid, G.M.; Cao, X.; Bitounis, D.; Singh, D.; Llopis, P.M.; Buckley, B.; Demokritou, P. Toxicity, uptake, and nuclear translocation of ingested micro-nanoplastics in an in vitro model of the small intestinal epithelium. Food Chem. Toxicol. 2021, 158, 112609. [Google Scholar] [CrossRef]
- Hu, L.; Zhao, Y.; Xu, H. Trojan horse in the intestine: A review on the biotoxicity of microplastics combined environmental contaminants. J. Haz. Mat. 2022, 439, 129652. [Google Scholar] [CrossRef]
- Von Moos, N.; Burkhardt-Holm, P.; Köhler, A. Uptake and effects of microplastics on cells and tissue of the blue mussel Mytilus edulis L. after an experimental exposure. Environ. Sci. Technol. 2012, 46, 11327–11335. [Google Scholar] [CrossRef]
- Brown, D.M.; Wilson, M.R.; MacNee, W.; Stone, V.; Donaldson, K. Size- dependent proinflammatory effects of ultrafine polystyrene particles: A role for surface area and oxidative stress in the enhanced activity of ultrafines. Toxicol. Appl. Pharmacol. 2001, 175, 191–199. [Google Scholar] [CrossRef]
- Capriotti, M. Microplastics as Carriers of Endocrine Disruptors. The “Impostors of the Hormones”, Endocrine Disruptors Can Travel in the Environment also Using Microplastics as Media. Earthsize. 2020. Available online: https://earthzine.org/microplastics-as-carriers-of-endocrine-disruptors/ (accessed on 23 April 2020).
- Lim, X.Z. Microplastics are everywhere—But are they harmful? Nature 2021, 593, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Pironti, C.; Ricciardi, M.; Motta, O.; Miele, Y.; Proto, A.; Montano, L. Microplastics in the Environment: Intake through the Food Web, Human Exposure and Toxicological Effects. Toxics 2021, 9, 224. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.; Baek, J.Y.; Koo, J.; Park, S.; Ryu, Y.K.; Kim, K.S.; Zhang, S.; Chung, C.; Dogan, R.; Choi, H.S.; et al. Maternal exposure to polystyrene nanoplastics causes brain abnormalities in progeny. J. Haz. Mat. 2022, 426, 127815. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, E.V.; Valdez, M.C.; Maximillian, E.; Denys, M.E.; Bishay, A.E.; Krum, J.M.; Rabbani, K.M.; Carrillo, V.; Gonzalez, G.M.; Lampel, G.; et al. Persistent autism-relevant behavioral phenotype and social neuropeptide alterations in female mice offspring induced by maternal transfer of PBDE congeners in the commercial mixture DE-71. Arch. Toxicol. 2022, 96, 335–365. [Google Scholar] [CrossRef]
- Fan, X.; Wei, X.; Hu, H.; Zhang, B.; Yang, D.; Du, H.; Zhu, H.; Sun, X.; Oh, Y.; Gu, N. Effects of oral administration of polystyrene nanoplastics on plasma glucose metabolism in mice. Chemosphere 2022, 288, 132607. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Trasande, L.; Kannan, K. Occurrence of Polyethylene Terephthalate and Polycarbonate Microplastics in Infant and Adult Feces. Environ. Sci. Technol. Lett. 2021, 8, 989–994. [Google Scholar] [CrossRef]
- Matassa, R.; Cattaruzza, M.S.; Sandorfi, F.; Battaglione, E.; Relucenti, M.; Familiari, G. Direct imaging evidence of metal inorganic contaminants traced into cigarettes. J. Haz. Mat. 2021, 5, 125092. [Google Scholar] [CrossRef]
- Palmerini, M.G.; Belli, M.; Nottola, S.A.; Miglietta, S.; Bianchi, S.; Bernardi, S.; Antonouli, S.; Cecconi, S.; Familiari, G.; Macchiarelli, G. Mancozeb impairs the ultrastructure of mouse granulosa cells in a dose-dependent manner. J. Reprod. Dev. 2018, 64, 75–82. [Google Scholar] [CrossRef]
- Antonouli, S.; Palmerini, M.G.; Bianchi, S.; Rossi, G.; Cecconi, S.; Belli, M.; Bernardi, S.; Khalili, M.A.; Familiari, G.; Nottola, S.A.; et al. Repeated hyperstimulation affects the ultrastructure of mouse fallopian tube epithelium. J. Reprod. Dev. 2020, 66, 387–397. [Google Scholar] [CrossRef]
- Liu, N.M.; Miyashita, L.; Maher, B.A.; McPhail, G.; Jones, C.J.P.; Barratt, B.; Thangaratinam, S.; Karloukovski, V.; Ahmed, I.A.; Aslam, Z.; et al. Evidence for the presence of air pollution nanoparticles in placental tissue cells. Sci. Total Environ. 2021, 751, 142235. [Google Scholar] [CrossRef]
- Girão, A.V. SEM/EDS and Optical Microscopy Analysis of Microplastics. In Handbook of Microplastics in the Environment; Rocha-Santos, T., Costa, M., Mouneyrac, C., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–22. [Google Scholar] [CrossRef]
- Gniadek, M.; Dąbrowska, A. The marine nano- and microplastics characterisation by SEM-EDX: The potential of the method in comparison with various physical and chemical approaches. Mar. Pollut. Bull. 2019, 148, 210–216. [Google Scholar] [CrossRef]
- Silva, A.B.; Bastos, A.S.; Justino, C.I.L.; da Costa, J.P.; Duarte, A.C.; Rocha-Santos, T.A.P. Microplastics in the environment: Challenges in analytical chemistry—A review. Anal. Chim. Acta 2018, 1017, 1–19. [Google Scholar] [CrossRef]
- Grafmueller, S.; Manser, P.; Diener, L.; Diener, P.A.; Maeder-Althaus, A.; Maurizi, L.; Jochum, W.; Krug, H.F.; Buerki-Thurnherr, T.; von Mandach, U.; et al. Bidirectional Transfer Study of Polystyrene Nanoparticles across the Placental Barrier in an ex Vivo Human Placental Perfusion Model. Environ. Health Perspect. 2015, 123, 12. [Google Scholar] [CrossRef]
- Benirschke, K.; Burton, G.J.; Baergen, R.N. Basic Structure of the Villous Trees. In Pathology of the Human Placenta, 6th ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 55–100. [Google Scholar] [CrossRef]
- Wang, Y.L.; Lee, Y.H.; Hsu, Y.H.; Chiu, I.J.; Huang, C.C.Y.; Chia, Z.C.; Lee, C.P.; Lin, Y.F.; Chiu, H.W. The Kidney-Related Effects of Polystyrene Microplastics on Human Kidney Proximal Tubular Epithelial Cells-HK-2 and Male C57BL/6 Mice. Environ. Health Perspect. 2021, 129, 057003. [Google Scholar] [CrossRef]
- Cheville, N.F. Part II: Organelle Pathology. In Ultrastructural Pathology. The Comparative Cellular Basis of Disease, 2nd ed.; Wiley-Blackwell: Ames, IA, USA, 2009; 138p, ISBN 978-0-813-80330-2. [Google Scholar]
- Johnston, J.A.; Ward, C.L.; Kopito, R.R. Aggresomes: A cellular response to misfolded proteins. J. Cell Biol. 1998, 143, 1883–1898. [Google Scholar] [CrossRef]
- Liu, X.-D.; Ko, S.; Xu, Y.; Fattah, E.A.; Xiang, Q.; Jagannath, C.; Ishii, T.; Komatsu, M.; Tony Eissa, N. Transient aggregation of ubiquitinated proteins is a cytosolic unfolded protein response to inflammation and endoplasmic reticulum stress. J. Biol. Chem. 2012, 287, 19687–19698. [Google Scholar] [CrossRef]
- Nakashima, A.; Shima, T.; Tsuda, S.; Aoki, A.; Kawaguchi, M.; Furuta, A.; Yasuda, I.; Yoneda, S.; Yamaki-Ushijima, A.; Cheng, S.B.; et al. Aggrephagy Deficiency in the Placenta: A New Pathogenesis of Preeclampsia. Int. J. Mol. Sci. 2021, 22, 2432. [Google Scholar] [CrossRef]
- Hollóczki, O. Evidence for protein misfolding in the presence of nanoplastics. Int. J. Quantum Chem. 2021, 121, e26372. [Google Scholar] [CrossRef]
- Burton, G.J.; Yung, H.-W.; Cindrova-Davies, T.; Charnock-Jones, D.S. Placental endoplasmic reticulum stress and oxidative stress in the pathophysiology of unexplained intrauterine growth restriction and early onset preeclampsia. Placenta 2009, 30, 43–48. [Google Scholar] [CrossRef]
- Burton, G.J.; Yung, H.W. Endoplasmic reticulum stress in the pathogenesis of early-onset pre-eclampsia. Pregnancy Hypertens Int. J. Womens Cardiovasc. Health 2011, 1, 72–78. [Google Scholar] [CrossRef]
- Ghemrawi, R.; Battaglia-Hsu, S.F.; Arnold, C. Endoplasmic Reticulum Stress in Metabolic Disorders. Cells 2018, 7, 63. [Google Scholar] [CrossRef]
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative stress, placental ageing-related pathologies and adverse pregnancy outcomes. Am. J. Reprod. Immunol. 2017, 77, e12653. [Google Scholar] [CrossRef]
- Yung, H.W.; Hemberger, M.; Watson, E.D.; Senner, C.E.; Jones, C.P.; Kaufman, R.J.; Charnock-Jones, D.S.; Burton, G.J. Endoplasmic reticulum stress disrupts placental morphogenesis: Implications for human intrauterine growth restriction. J. Pathol. 2012, 228, 554–564. [Google Scholar] [CrossRef]
- Burton, G.J.; Yung, H.W.; Murray, A.J. Mitochondrial—Endoplasmic reticulum interactions in the trophoblast: Stress and senescence. Placenta 2017, 52, 146–155. [Google Scholar] [CrossRef]
- Guzel, E.; Arlier, S.; Guzeloglu-Kayisli, O.; Tabak, M.S.; Ekiz, T.; Semerci, N.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Endoplasmic Reticulum Stress and Homeostasis in Reproductive Physiology and Pathology. Int. J. Mol. Sci. 2017, 18, 792. [Google Scholar] [CrossRef]
- Liu, T.; Hou, B.; Wang, Z.; Yang, Y. Polystyrene microplastics induce mitochondrial damage in mouse GC-2 cells. Ecotoxicol. Environ. Saf. 2022, 237, 113520. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, R.; Li, B.; Du, Y.; Li, J.; Tong, X.; Wu, Y.; Ji, X.; Zhang, Y. Tissue distribution of polystyrene nanoplastics in mice and their entry, transport, and cytotoxicity to GES-1 cells. Environ. Pollut. 2021, 280, 116974. [Google Scholar] [CrossRef]
- Kaasik, A.; Safiulina, D.; Zharkovsky, A.; Veksler, V. Regulation of mitochondrial matrix volume. Am. J. Physiol. Cell. Physiol. 2007, 292, C157–C163. [Google Scholar] [CrossRef]
- Ghadially, F.N. Chapter 3: Mitochondria. In Ultrastructural Pathology of the Cell and Matrix, 3rd ed.; Butterworths: Oxford, UK, 1997; Volume 1, ISBN 0-407-01571-X. [Google Scholar]
- Xavier, V.J.; Martinou, J.C. RNA Granules in the Mitochondria and Their Organization under Mitochondrial Stresses. Int. J. Mol. Sci. 2021, 22, 9502. [Google Scholar] [CrossRef]
- Jarc, E.; Petan, T. Lipid Droplets and the Management of Cellular Stress. Yale J. Biol. Med. 2019, 92, 435–452. [Google Scholar]
- Lee, S.-J.; Zhang, J.; Choi, A.M.K.; Kim, H.P. Mitochondrial dysfunction induces formation of lipid droplets as a generalized response to stress. Oxid. Med. Cell. Longev. 2013, 2013, 327167. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Li, D.; Guo, J.; Chen, J. Mechanistic toxicity assessment of differently sized and charged polystyrene nanoparticles based on human placental cells. Water Res. 2022, 223, 118960. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, P.M.; Twayana, K.S.; Ravanan, P.; Thomas, J.; Mukherjee, A.; Jenkins, D.F.; Chandrasekaran, N. Prospects on the nano-plastic particles internalization and induction of cellular response in human keratinocytes. Part. Fibre Toxicol. 2021, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Cortés, C.; Domenech, J.; Salazar, M.; Pastor, S.; Marcos, R.; Hernádez, A. Nanoplastics as a potential environmental health factor: Effects of polystyrene nanoparticles on human intestinal epithelial Caco-2 82 cells. Environ. Sci. Nano 2020, 7, 272–285. [Google Scholar] [CrossRef]
- Lu, Y.-Y.; Li, H.; Ren, H.; Zhang, X.; Huang, F.; Zhang, D.; Huang, Q.; Zhang, X. Size-dependent effects of polystyrene nanoplastics on autophagy response in human umbilical vein endothelial cells. J. Hazard. Mater. 2022, 421, 126770. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, C.; Yu, S.; Liu, S.; Wang, G.; Lan, H.; Zheng, X.; Li, S. Glycine ameliorates MBP-induced meiotic abnormalities and apoptosis by regulating mitochondrial-endoplasmic reticulum interactions in porcine oocytes. Environ. Pollut. 2022, 309, 119756. [Google Scholar] [CrossRef]
- Gautam, R.; Jo, J.H.; Acharya, M.; Maharjan, A.; Lee, D.; Bahadur, P.K.C.; Kim, C.; Kim, K.; Kim, H.; Heo, Y. Evaluation of potential toxicity of polyethylene microplastics on human derived cell lines. Sci. Total Environ. 2022, 838, 156089. [Google Scholar] [CrossRef]
- Gyllenhammer, L.E.; Entringer, S.; Buss, C.; Wadhwa, P.D. Developmental programming of mitochondrial biology: A conceptual framework and review. Proc. Biol. Sci. 2020, 287, 20192713. [Google Scholar] [CrossRef]
- Kwon, E.J.; Kim, Y.J. What is fetal programming?: A lifetime health is under the control of in utero health. Obstet. Gynecol. Sci. 2017, 60, 506–519. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, Y.; Long, J.; Yang, X.; Bao, L.; Yang, Z.; Wu, B.; Si, R.; Zhao, W.; Peng, C.; et al. Polystyrene microplastic exposure induces insulin resistance in mice via dysbacteriosis and pro-inflammation. Sci. Total Environ. 2022, 838, 155937. [Google Scholar] [CrossRef]
- DeLoid, G.M.; Cao, X.; Coreas, R.; Bitounis, D.; Singh, D.; Zhong, W.; Demokritou, P. Incineration-Generated Polyethylene Micro-Nanoplastics Increase Triglyceride Lipolysis and Absorption in an In Vitro Small Intestinal Epithelium Model. Environ. Sci. Technol. 2022, 56, 12288–12297. [Google Scholar] [CrossRef]
- Naviaux, R.K. Metabolic features of the cell danger response. Mitochondrion 2014, 16, 7–17. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Age | 33 | 25 | 38 | 36 | 28 | 36 | 26 | 42 | 31 | 39 |
| Types of childbirth | Natural | C-section | Natural | Natural | C-section | C-section | Natural | C-section | Natural | C-section |
| Vegetarian | No | No | No | No | No | No | No | No | No | No |
| Seafood in the last 5 days | No | No | No | No | No | No | No | No | No | Yes |
| Pathologies | Gestional Diabetes with insulin | No | No | No | No | No | No | Hypotiroidism | No | No |
| Food in plastic wrapping | Yes | Yes | Yes | No | Yes | Yes | Yes | Yes | Yes | Yes |
| Drinks in plastic containers | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Cosmetics with synthetic polymers | Rilastil, Eucerin | Yes | Scrub Yves Roche | Scub Yves Roche | No | Scrub shower gel | No | No | Bionike | Veralab, Mustela, Dove |
| Toothpastes with syntethic polymers | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Chewing-gum | No | No | Yes, once a month | No | Yes, once a month | No | Yes, once a month | No | No | No |
| Smoke | Ex | 10cg/die | No | No | No | No | No | No | No | No |
| Alcohol | No | No | No | No | No | No | No | No | No | No |
| Patient | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| MPs | ++ | + | ++ | ++ | + | +++ | +++ | ++ | ++ | ++ |
| Villus layer localization | outer/basement membrane | outer | outer | outer | outer | outer/inner | outer/inner | outer/basement membrane | outer | outer/inner |
| Intracellular localization | lysosomes/peroxisomes/lipid droplets | lysosomes | vacuoles | lysosomes/peroxisomes | lysosomes | multivesicular bodies/peroxisomes | multivesicular bodies/lipid droplets | multivesicular bodies | lysosomes/peroxisomes | lysosomes |
| Extracellular localization | stroma | stroma | stroma | stroma | stroma | stroma/sinusoids (pericytes) | stroma/sinusoids (pericytes/endothelial cells) | stroma | stroma | stroma/sinusoids (pericytes) |
| ER dilatation | ++ | + | ++ | ++ | + | +++ | +++ | +++ | ++ | ++ |
| Aggresomes | +++ | - | - | - | - | - | + | + | - | - |
| Mitochondria damaged | ++ (swollen) | + (swollen) | ++ (swollen) | ++ (swollen) | + (swollen) | ++ (swollen/ picnotic) | +++ (swollen/ picnotic) | ++ (swollen/ picnotic) | ++ (swollen) | ++ (swollen/picnotic) |
| Mitochondrial granules | - | - | ++ | ++ | - | ++ | +++ | ++ | ++ | + |
| Whorled membranous bodies | ++ involution of endoplasmic reticulum/mitochondria | + involution of mitochondria/microvilli | +++ involution of endoplasmic reticulum/mitochondria/microvilli | ++ involution of endoplasmic reticulum/mitochondria | + involution of mitochondria/microvilli | ++ involution of endoplasmic reticulum/mitochondria/microvilli | +++ involution of endoplasmic reticulum/mitochondria | ++ involution of endoplasmic reticulum/mitochondria/microvilli | ++ involution of endoplasmic reticulum/mitochondria | ++ involution of endoplasmic reticulum/mitochondria |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragusa, A.; Matta, M.; Cristiano, L.; Matassa, R.; Battaglione, E.; Svelato, A.; De Luca, C.; D’Avino, S.; Gulotta, A.; Rongioletti, M.C.A.; et al. Deeply in Plasticenta: Presence of Microplastics in the Intracellular Compartment of Human Placentas. Int. J. Environ. Res. Public Health 2022, 19, 11593. https://doi.org/10.3390/ijerph191811593
Ragusa A, Matta M, Cristiano L, Matassa R, Battaglione E, Svelato A, De Luca C, D’Avino S, Gulotta A, Rongioletti MCA, et al. Deeply in Plasticenta: Presence of Microplastics in the Intracellular Compartment of Human Placentas. International Journal of Environmental Research and Public Health. 2022; 19(18):11593. https://doi.org/10.3390/ijerph191811593
Chicago/Turabian StyleRagusa, Antonio, Maria Matta, Loredana Cristiano, Roberto Matassa, Ezio Battaglione, Alessandro Svelato, Caterina De Luca, Sara D’Avino, Alessandra Gulotta, Mauro Ciro Antonio Rongioletti, and et al. 2022. "Deeply in Plasticenta: Presence of Microplastics in the Intracellular Compartment of Human Placentas" International Journal of Environmental Research and Public Health 19, no. 18: 11593. https://doi.org/10.3390/ijerph191811593
APA StyleRagusa, A., Matta, M., Cristiano, L., Matassa, R., Battaglione, E., Svelato, A., De Luca, C., D’Avino, S., Gulotta, A., Rongioletti, M. C. A., Catalano, P., Santacroce, C., Notarstefano, V., Carnevali, O., Giorgini, E., Vizza, E., Familiari, G., & Nottola, S. A. (2022). Deeply in Plasticenta: Presence of Microplastics in the Intracellular Compartment of Human Placentas. International Journal of Environmental Research and Public Health, 19(18), 11593. https://doi.org/10.3390/ijerph191811593