
Exe-Muscle: An Exercised Human Skeletal Muscle Gene Expression Database

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Data Processing and Website Construction
2.3. Short Time-Series Expression Miner (STEM) Trend Analysis
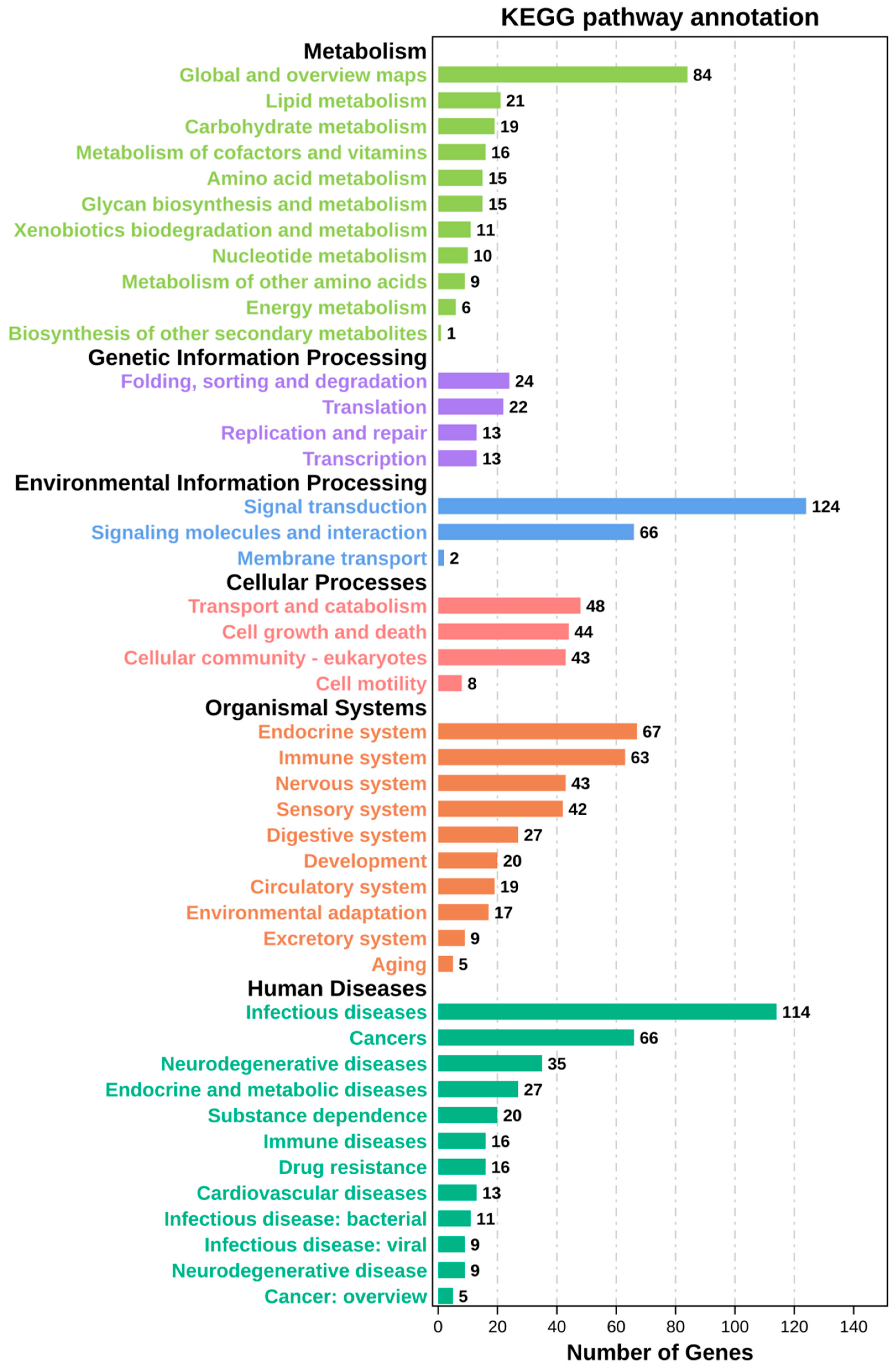
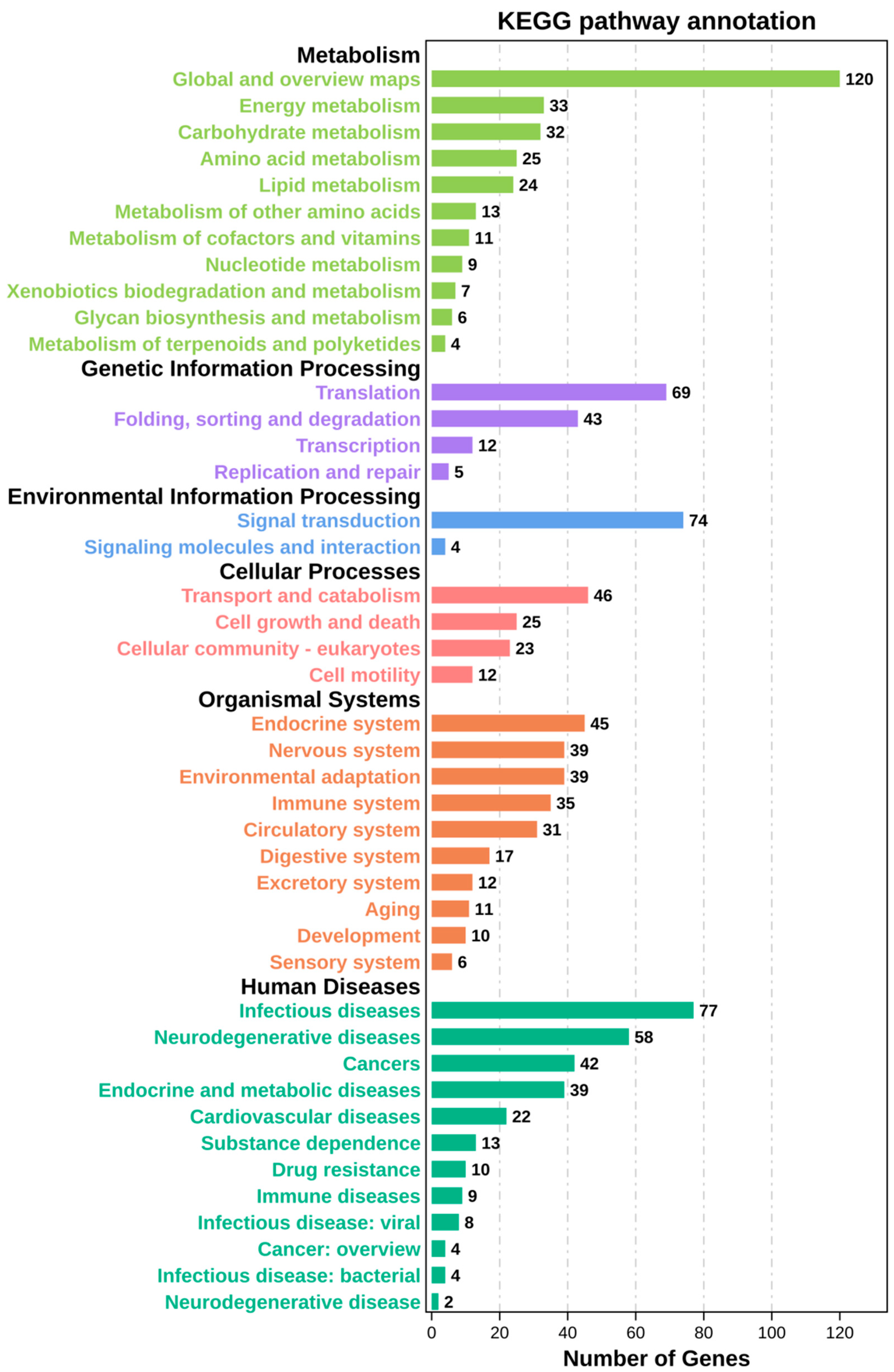
2.4. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis of Time-Series Genes
3. Results
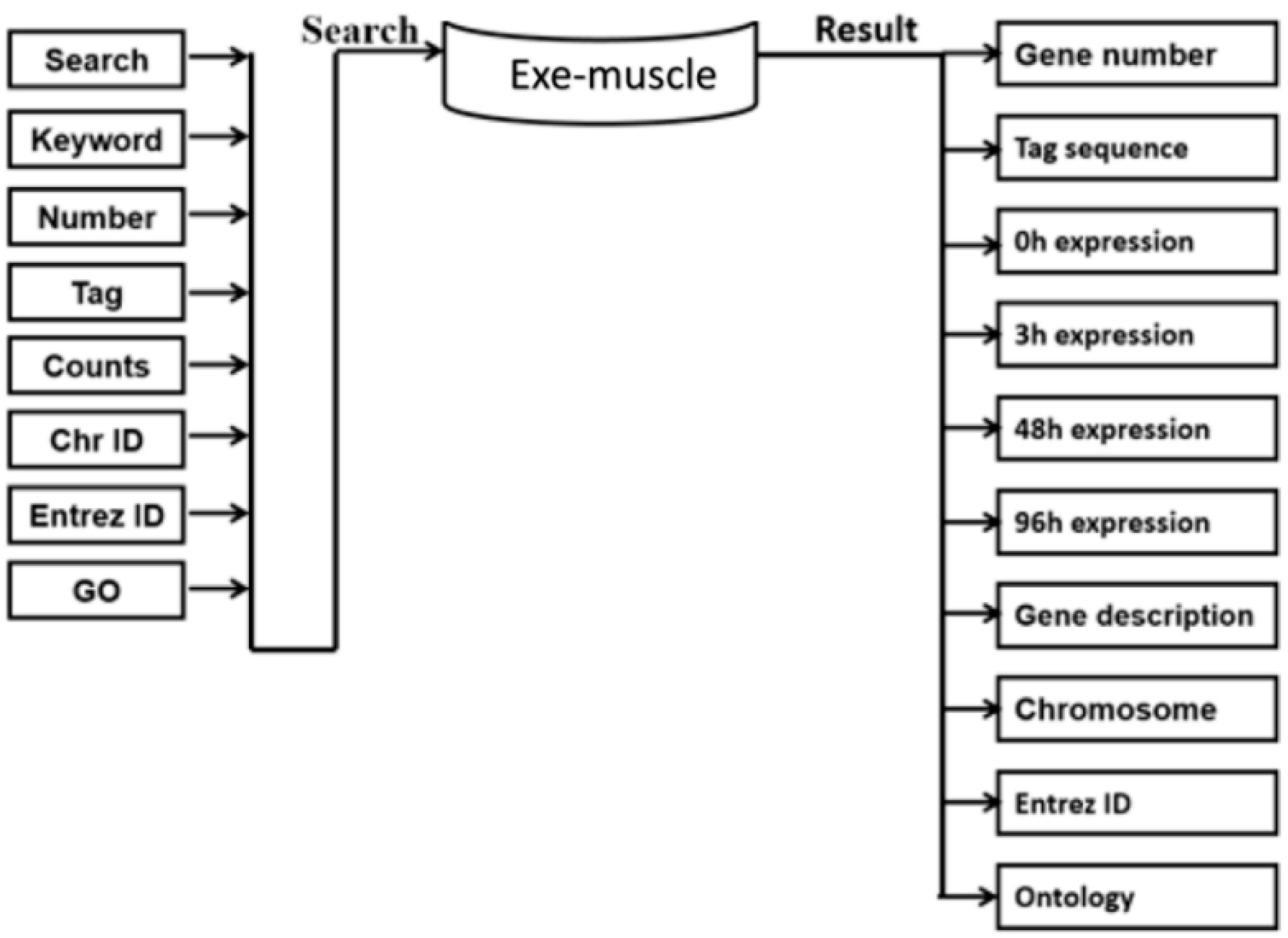
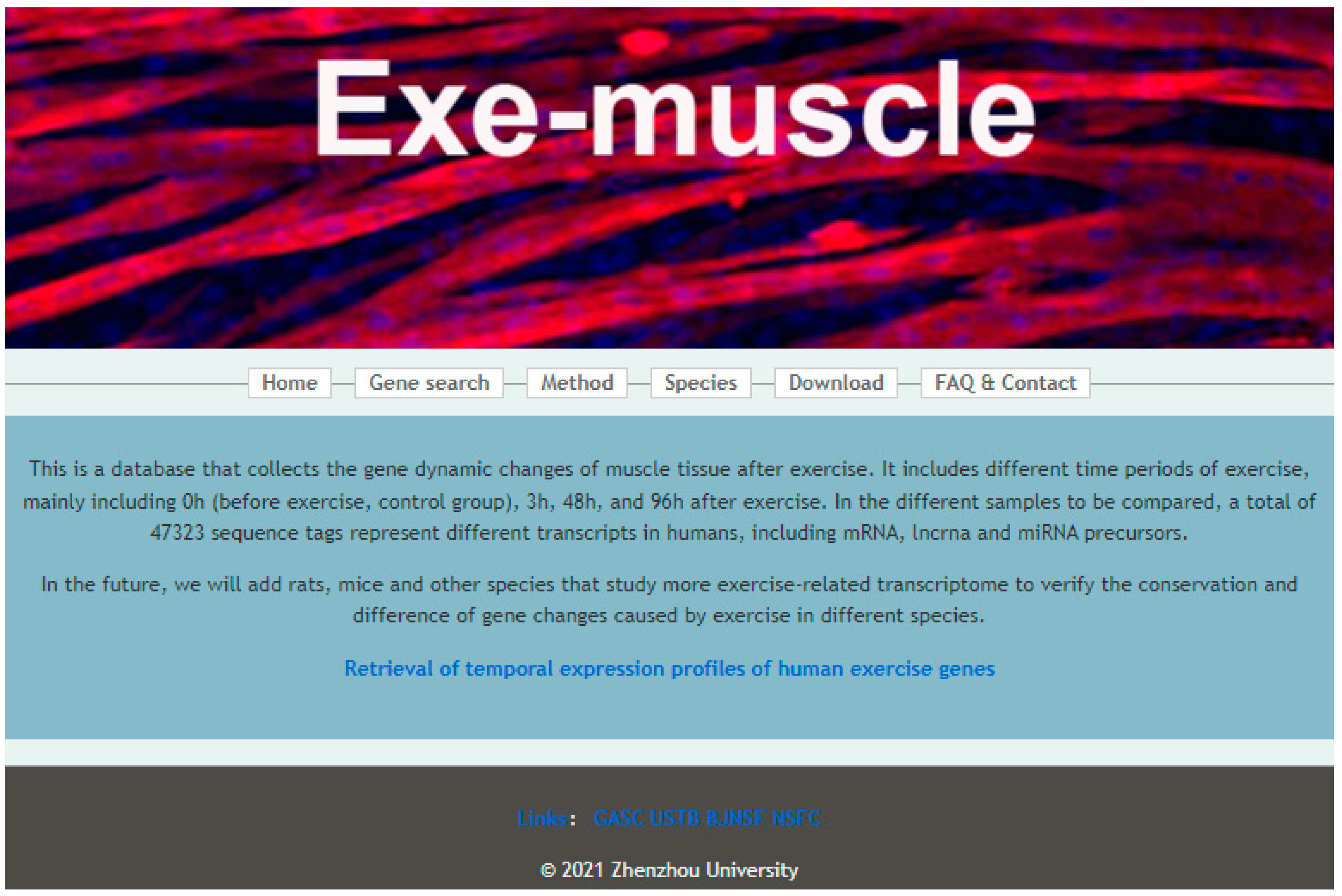
3.1. Construction of the Exe-Muscle Database System
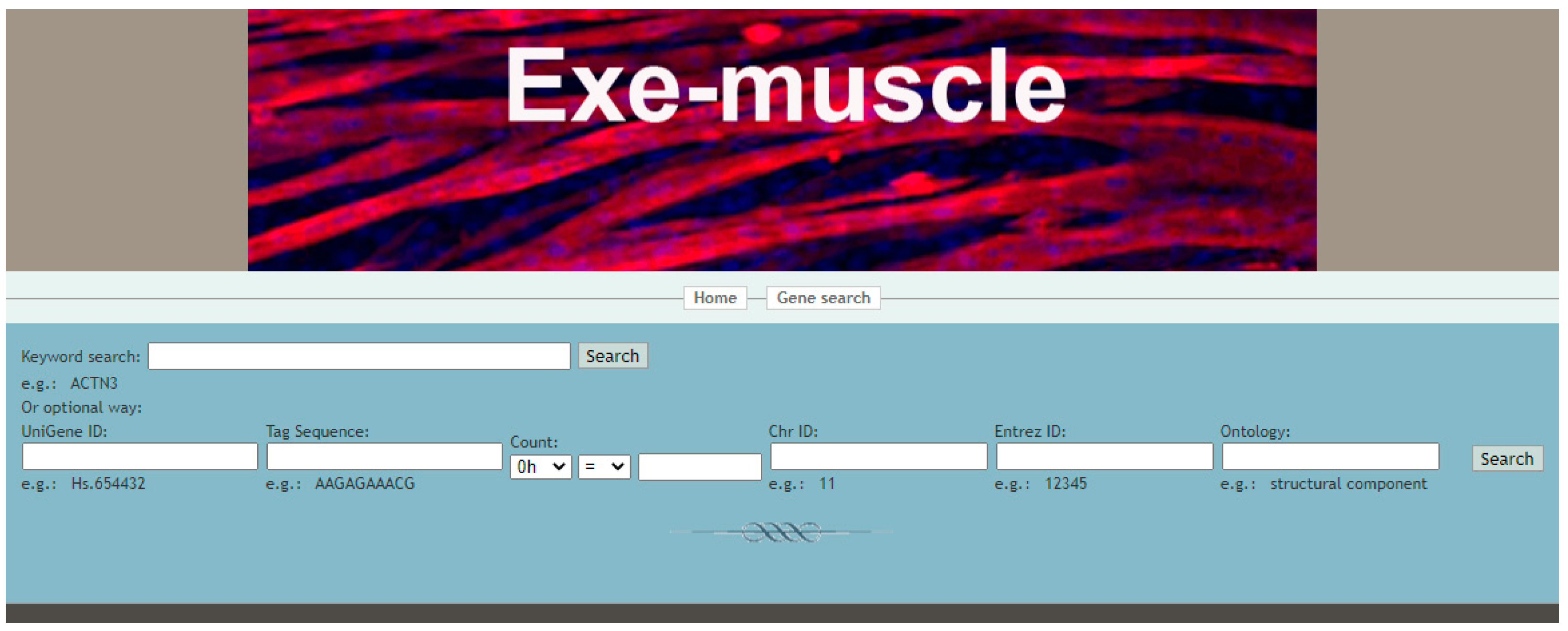
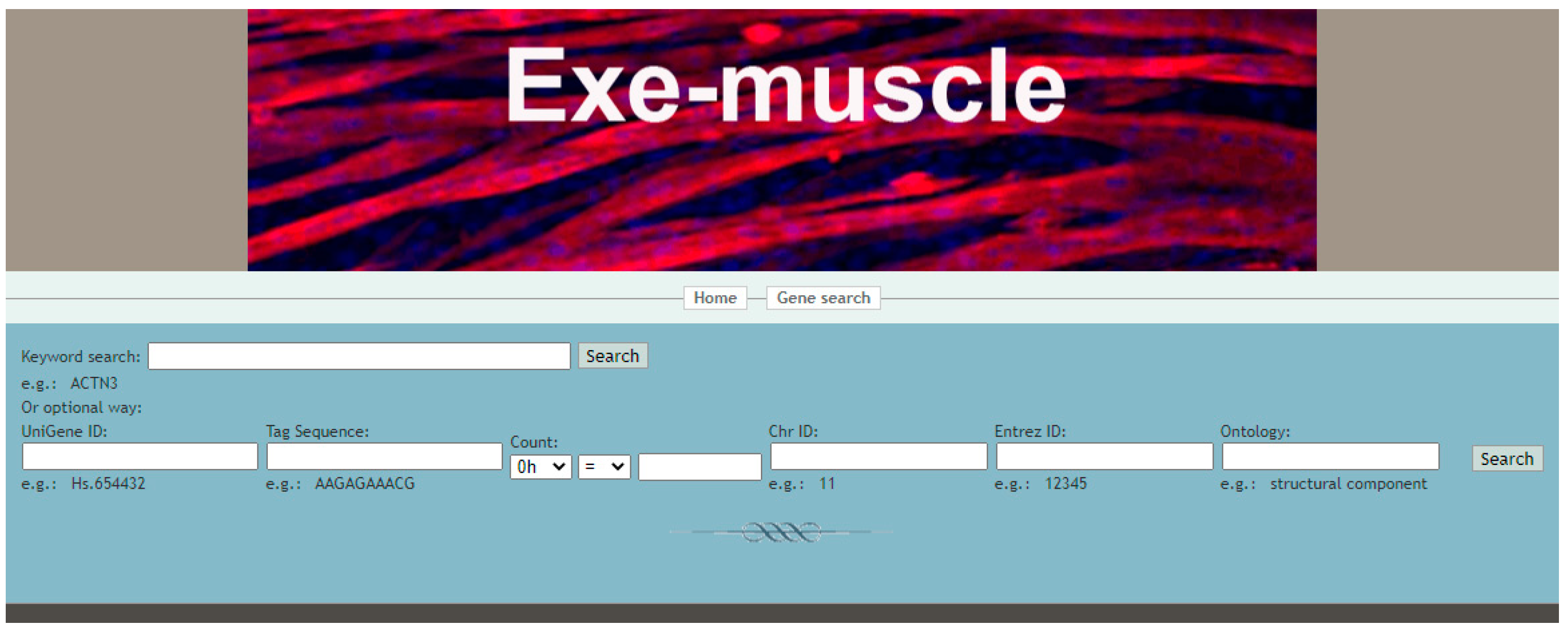
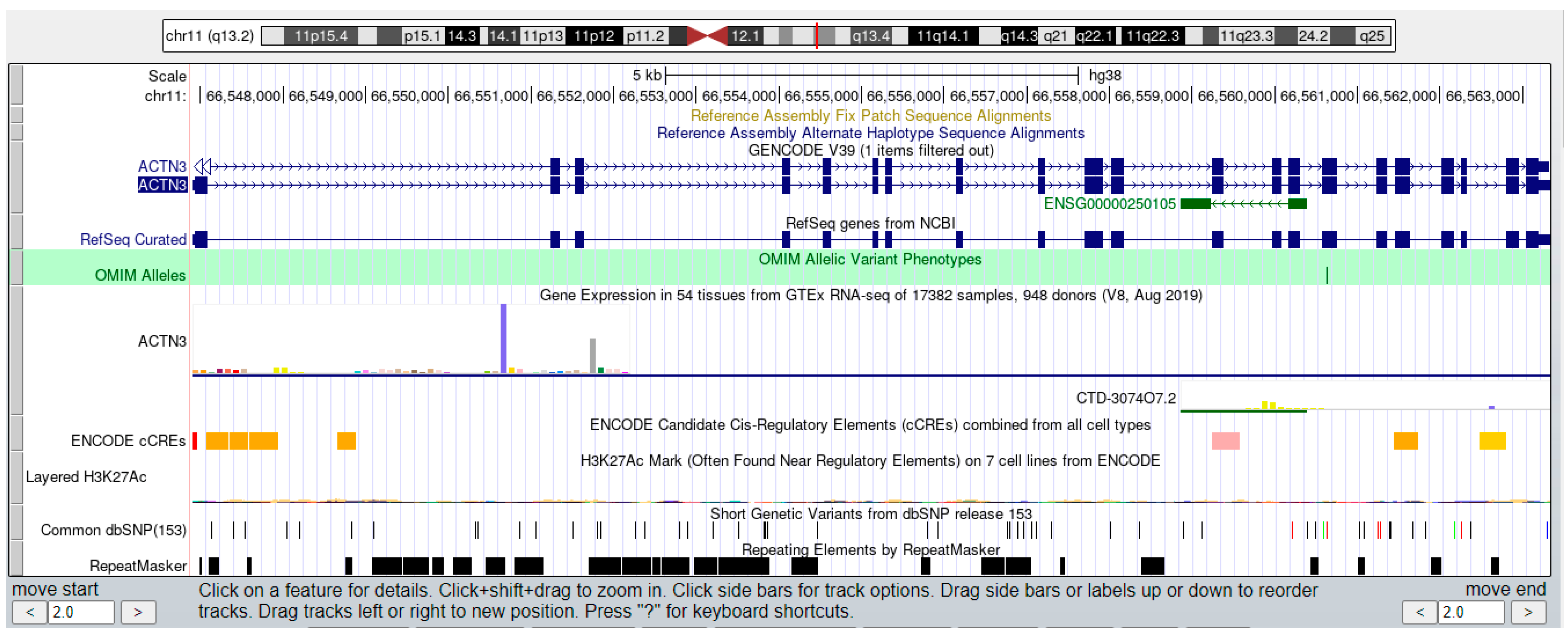
3.2. Using the Exe-Muscle Database to Query the Changes of Related Genes during Exercise
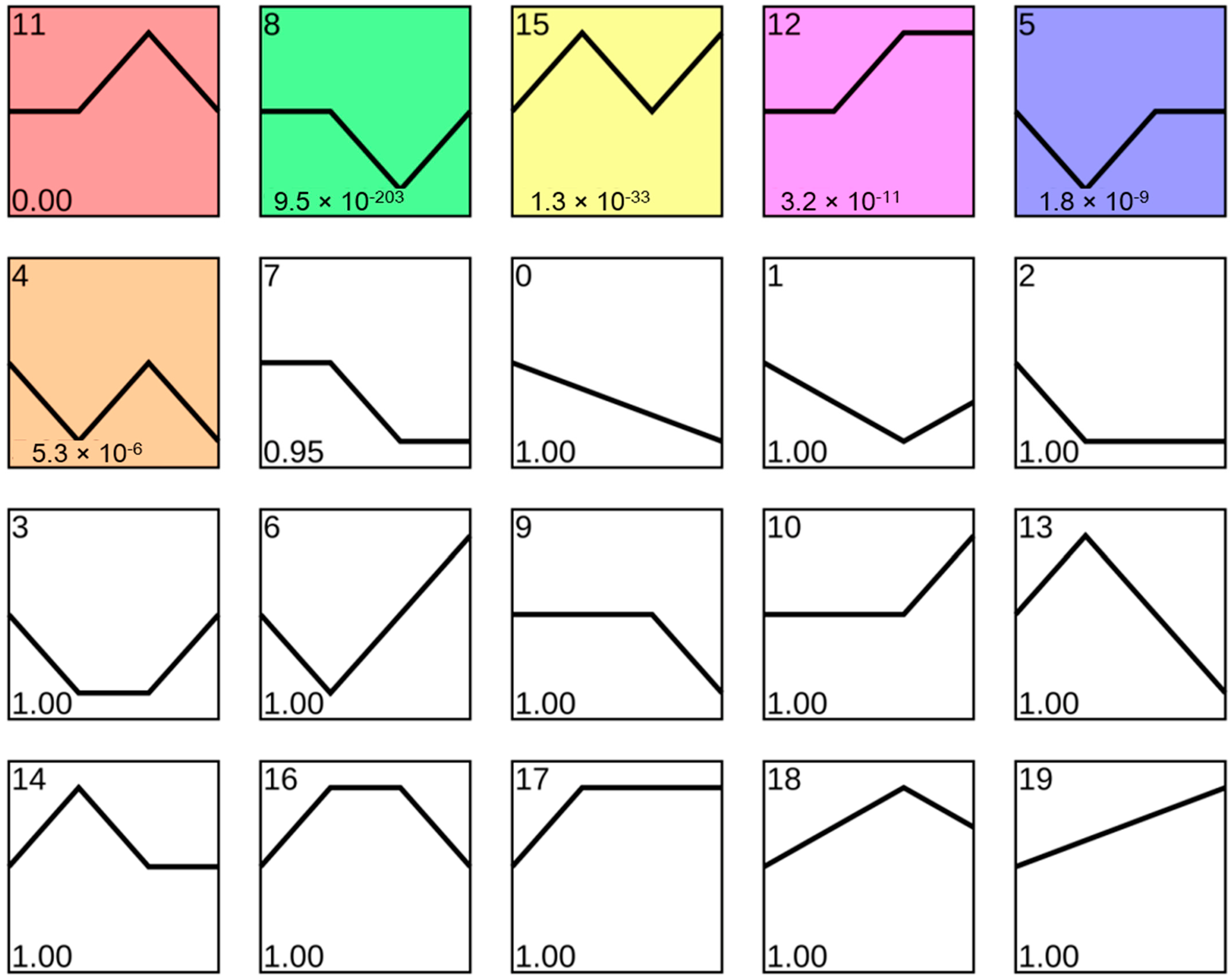
3.3. Using Exe-Muscle to Explore the Change Trend of Genes in Muscle during Exercise
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sartori, F.M.O.; Buzby, C.; Plavskin, Y.; Siegal, M.L. High-Throughput Live Imaging of Microcolonies to Measure Heterogeneity in Growth and Gene Expression. J. Vis. Exp. 2021, 170, e62038. [Google Scholar] [CrossRef]
- Ishii, S.; Fukui, K.; Yokoshima, S.; Kumagai, K.; Beniyama, Y.; Kodama, T.; Fukuyama, T.; Okabe, T.; Nagano, T.; Kojima, H.; et al. High-throughput Screening of Small Molecule Inhibitors of the Streptococcus Quorum-sensing Signal Pathway. Sci. Rep. 2017, 7, 4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagura-Ikeda, M.; Imai, K.; Tabata, S.; Miyoshi, K.; Murahara, N.; Mizuno, T.; Horiuchi, M.; Kato, K.; Imoto, Y.; Iwata, M.; et al. Clinical Evaluation of Self-Collected Saliva by Quantitative Reverse Transcription-PCR (RT-qPCR), Direct RT-qPCR, Reverse Transcription-Loop-Mediated Isothermal Amplification, and a Rapid Antigen Test to Diagnose COVID-19. J. Clin. Microbiol. 2020, 58, e01438-20. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Cho, J.W.; Lee, S.; Yun, A.; Kim, H.; Bae, D.; Yang, S.; Kim, C.Y.; Lee, M.; Kim, E.; et al. TRRUST v2: An expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018, 46, D380–D386. [Google Scholar] [CrossRef]
- Liang, W.; Zhao, Y.; Huang, W.; Gao, Y.; Xu, W.; Tao, J.; Yang, M.; Li, L.; Ping, W.; Shen, H.; et al. Non-invasive diagnosis of early-stage lung cancer using high-throughput targeted DNA methylation sequencing of circulating tumor DNA (ctDNA). Theranostics 2019, 9, 2056–2070. [Google Scholar] [CrossRef]
- Midkiff, D.; San-Miguel, A. Microfluidic Technologies for High Throughput Screening Through Sorting and On-Chip Culture of C. elegans. Molecules 2019, 24, 4292. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Feng, Y.C.; Mou, H.M.; Yue, T. Analysis of screening coronary heart disease markers and bioinformatics based on GEO database chip. Chin. J. Integr. Med. Cardio-Cerebrovasc. Dis. 2021, 19, 3460–3464. [Google Scholar]
- Raue, U.; Slivka, D.; Jemiolo, B.; Hollon, C.; Trappe, S. Proteolytic gene expression differs at rest and after resistance exercise between young and old women. J. Gerontol. 2007, 62, 1407–1412. [Google Scholar] [CrossRef] [Green Version]
- McGlory, C.; Devries, M.C.; Phillips, S.M. Skeletal muscle and resistance exercise training: The role of protein synthesis in recovery and remodeling. J. Appl. Physiol. 2017, 122, 541–548. [Google Scholar] [CrossRef]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [Green Version]
- Colberg, S.R.; Sigal, R.J.; Fernhall, B.; Regensteiner, J.G.; Blissmer, B.J.; Rubin, R.R.; Chasan-Taber, L.; Albright, A.L.; Braun, B. Exercise and type 2 diabetes: American College of Sports Medicine and the American Diabetes Association: Joint position statement. Exercise and type 2 diabetes. Med. Sci. Sports Exerc. 2010, 42, 2282–2303. [Google Scholar]
- Kataoka, R.; Vasenina, E.; Hammert, W.B.; Ibrahim, A.H.; Dankel, S.J.; Buckner, S.L. Is there Evidence for the Suggestion that Fatigue Accumulates Following Resistance Exercise? Sports Med. 2022, 52, 25–36. [Google Scholar] [CrossRef]
- Louis, E.; Raue, U.; Yang, Y.; Jemiolo, B.; Trappe, S. Time course of proteolytic, cytokine, and myostatin gene expression after acute exercise in human skeletal muscle. J. Appl. Physiol. 2007, 103, 1744–1751. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, L.; Sun, X. Effect of aerobic exercise on gene expression in human skeletal muscle. J. China Med. Univ. 2020, 49, 556–560. [Google Scholar]
- Liao, B.; Li, Y. Effects of Exercise Combined with Chronic Rapamycin on mRNA Expression of Protein Involved in Energy Metabolism in Rats Fed a High–fat Diet. Chin. J. Sports Med. 2020, 39, 39–46. [Google Scholar]
- Shirvani, H.; Mirnejad, R.; Soleimani, M.; Arabzadeh, E. Swimming Exercise Improves Gene Expression of PPAR-γ and Downregulates the Overexpression of TLR4, MyD88, IL-6, and TNF-α after High-Fat Diet in Rat Skeletal Muscle Cells. Gene 2021, 775, 145441. [Google Scholar] [CrossRef]
- Lundberg, T.R.; Fernandez-Gonzalo, R.; Gustafsson, T.; Tesch, P.A. Aerobic exercise does not compromise muscle hypertrophy response to short-term resistance training. J. Appl. Physiol. 2013, 114, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Cho, H.S.; Yoon, K.J.; Lee, W.; Moon, H.Y. Exercise-induced changes of gene expression in the cerebellum of aged mice. Biochem. Biophys. Res. Commun. 2020, 521, 952–956. [Google Scholar] [CrossRef]
- Chung, H.C.; Keiller, D.R.; Roberts, J.D.; Gordon, D.A. Do exercise-associated genes explain phenotypic variance in the three components of fitness? a systematic review & meta-analysis. PLoS ONE 2021, 16, e0249501. [Google Scholar]
- Stipp, D. Gene Chip Breakthrough. Fortune 1997, 135, 56. [Google Scholar]
- Detlev, G.; Klaus, R. Gene Chip Technology. In Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Zambon, A.C.; Mcdearmon, E.L.; Salomonis, N.; Vranizan, K.M.; Johansen, K.L.; Adey, D.; Takahashi, J.S.; Schambelan, M.; Conklin, B.R. Time-and exercise-dependent gene regulation in human skeletal muscle. Genome Biol. 2003, 4, R61. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Creer, A.; Jemiolo, B.; Trappe, S. Time course of myogenic and metabolic gene expression in response to acute exercise in human skeletal muscle. J. Appl. Physiol. 2005, 98, 1745–1752. [Google Scholar] [CrossRef] [Green Version]
- Mustian, K.M.; Janelsins, M.C.; Mohile, S.G.; Kamen, C.S.; Peppone, L.J.; Flannery, M.A.; Tejany, M.A.; Peoples, A.R.; Heckler, C.E.; Voss, J. Exercise-induced changes in gene expression, muscular strength, and cancer-related fatigue in older prostate cancer patients. J. Clin. Oncol. 2014, 22, 51–56. [Google Scholar] [CrossRef]
- Schmutz, S.; Däpp, C.; Wittwer, M.; Vogt, M.; Hoppeler, H.; Flück, M. Endurance training modulates the muscular transcriptome response to acute exercise. Pflügers Arch. Eur. J. Physiol. 2006, 451, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Rao, W.; Cheng, P. The Interactive Query Techniques of Database Based on Web. Microcomput. Dev. 1999, 4, 23–25. [Google Scholar]
- Zhang, Y. To Access Information Classification and Code Online—An Example of Web Database Development Based on PHP+Apache+MySQL. Inf. Sci. 2004, 22, 510–512. [Google Scholar]
- Neubauer, O.; Sabapathy, S.; Lazarus, R.; Jowett, J.B.M.; Desbrow, B.; Peake, J.M.; Cameron-Smith, D.; Haseler, L.J.; Wagner, K.-H.; Bulmer, A.C. Transcriptome analysis of neutrophils after endurance exercise reveals novel signaling mechanisms in the immune response to physiological stress. J. Appl. Physiol. 2013, 114, 1677–1688. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, T.; Li, M. Construction of Development Environment of Linux-based Database-motivated Websites. J. Guangxi Acad. Sci. 2002, 18, 214–217. [Google Scholar]
- Niemi, A.K.; Majamaa, K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. Eur. J. Hum. Genet. 2005, 13, 965–969. [Google Scholar] [CrossRef]
- MacArthur, D.G.; Seto, J.T.; Raftery, J.M.; Quinlan, K.G.; Huttley, G.A.; Hook, J.W.; Lemckert, F.A.; Kee, A.J.; Edwards, M.R.; Berman, Y.; et al. Loss of ACTN3 gene function alters mouse muscle metabolism and shows evidence of positive selection in humans. Nat. Genet. 2007, 39, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, W.; Ma, Z.; Hu, Y.; Chen, H. Tartary buckwheat database (TBD): An integrative platform for gene analysis of and biological information on Tartary buckwheat. J. Zhejiang Univ. Sci. B 2021, 22, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M. Exercise and Gene Expression. Prog. Mol. Biol. Transl. Sci. 2015, 135, 457–469. [Google Scholar] [PubMed]
- Turner, D.C.; Gorski, P.P.; Seaborne, R.A.; Viggars, M.; Murphy, M.; Jarvis, J.C.; Martin, N.R.W.; Stewart, C.E.; Sharples, A.P. Mechanical loading of bioengineered skeletal muscle in vitro recapitulates gene expression signatures of resistance exercise in vivo. J. Cell. Physiol. 2021, 236, 6534–6547. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, K.; Song, J.; Kong, W.; Deng, Z. Exe-Muscle: An Exercised Human Skeletal Muscle Gene Expression Database. Int. J. Environ. Res. Public Health 2022, 19, 8806. https://doi.org/10.3390/ijerph19148806
Huang K, Song J, Kong W, Deng Z. Exe-Muscle: An Exercised Human Skeletal Muscle Gene Expression Database. International Journal of Environmental Research and Public Health. 2022; 19(14):8806. https://doi.org/10.3390/ijerph19148806
Chicago/Turabian StyleHuang, Kaiyuan, Jingwen Song, Weishuai Kong, and Zhongyuan Deng. 2022. "Exe-Muscle: An Exercised Human Skeletal Muscle Gene Expression Database" International Journal of Environmental Research and Public Health 19, no. 14: 8806. https://doi.org/10.3390/ijerph19148806
APA StyleHuang, K., Song, J., Kong, W., & Deng, Z. (2022). Exe-Muscle: An Exercised Human Skeletal Muscle Gene Expression Database. International Journal of Environmental Research and Public Health, 19(14), 8806. https://doi.org/10.3390/ijerph19148806