
Perspectives of Nanoparticles in Male Infertility: Evidence for Induced Abnormalities in Sperm Production

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Male Reproductive System and Potential
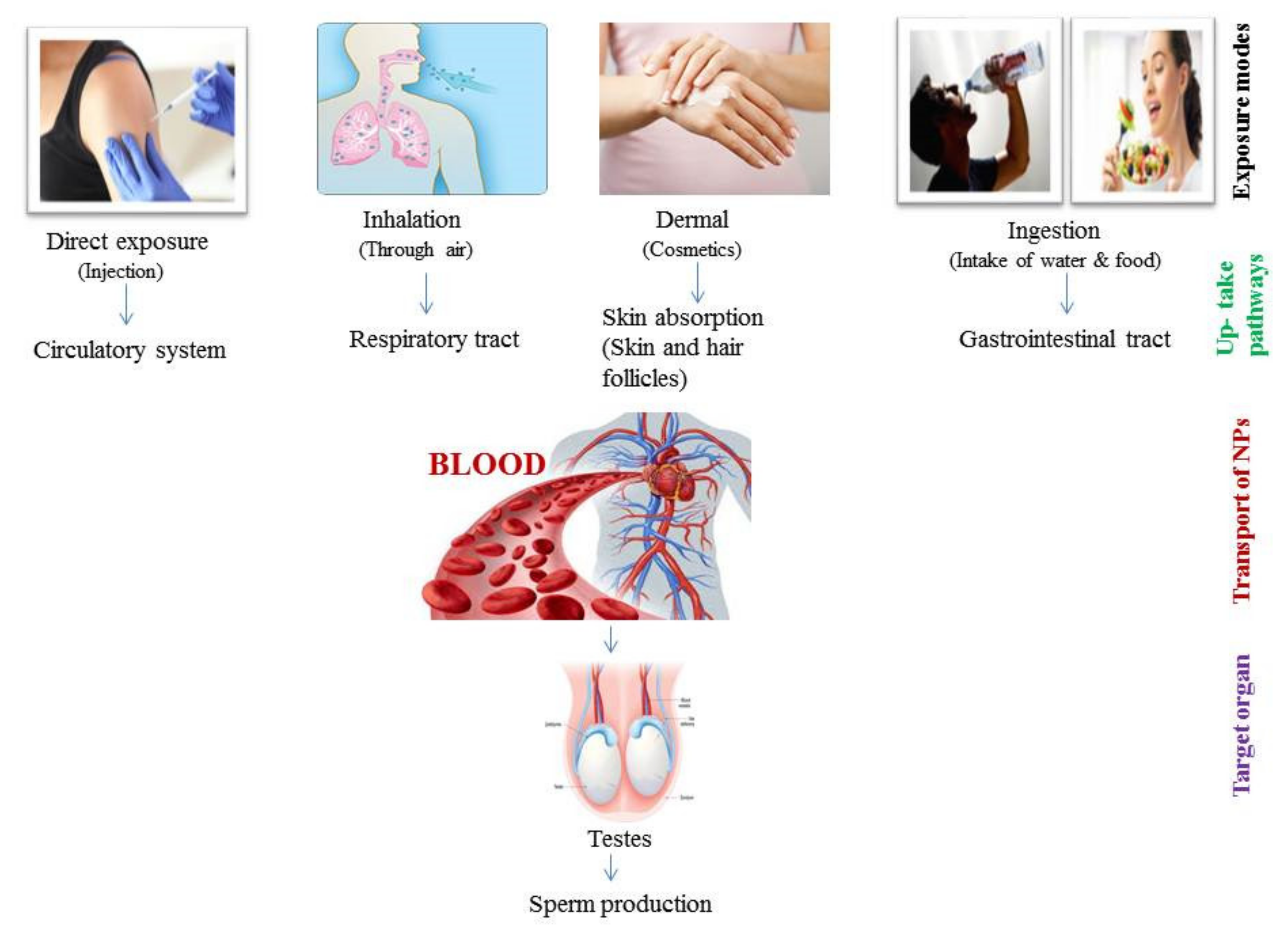
3. Path of NPs into the Reproductive System
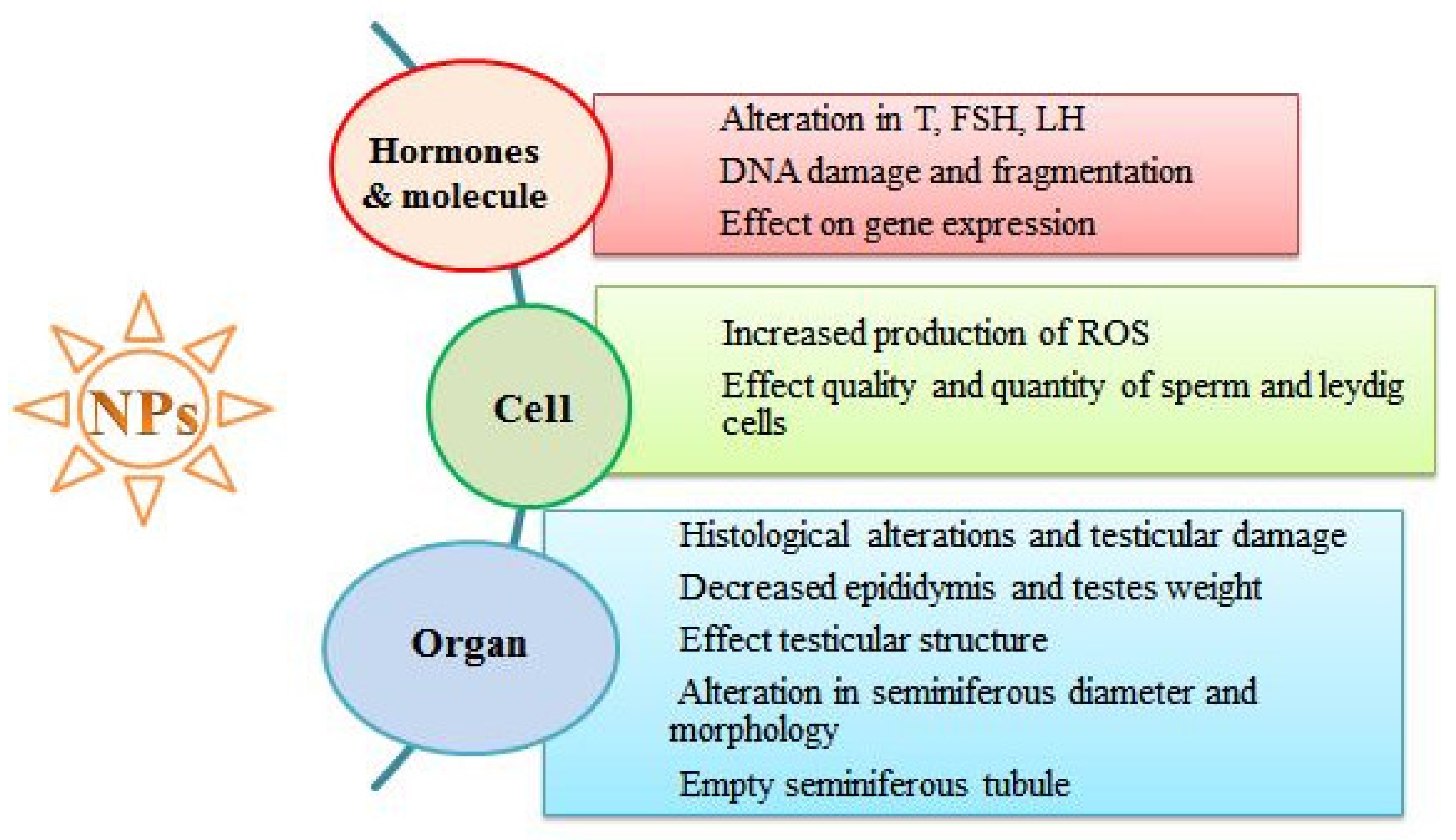
4. Adverse Effects of NPs on the Reproductive System
4.1. Adverse Effects of NPs on Reproductive Organ Weight
4.2. Adverse Effects of NPs on Seminiferous Tubules
4.3. Adverse Effects of NPs on Sperm Cells
4.4. Adverse Effects of NPs on Hormones Involved in Sperm Production
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Tiwari, J.N.; Tiwari, R.N.; Kim, K.S. Zero-dimensional, one-dimensional, two-dimensional and three-dimensional nanostructured materials for advanced electrochemical energy devices. Prog. Mater. Sci. 2012, 57, 724–803. [Google Scholar] [CrossRef]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella Jr, M.F.; Rejeski, D.; Hull, M.S. Nanotechnology in the real world: Redeveloping the nanomaterial consumer products inventory. Beilstein J. Nanotech. 2015, 6, 1769–1780. [Google Scholar] [CrossRef]
- Ema, M.; Kobayashi, N.; Naya, M.; Hanai, S.; Nakanishi, J. Reproductive and developmental toxicity studies of manufactured nanomaterials. Reprod. Toxicol. 2010, 30, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Oberdorster, G.; Oberdorster, E.; Oberdorster, J. Nanotoxicology: An emerging discipline evolution from studies of ultrafine particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Maisano, M.; Cappello, T.; Catanese, E.; Vitale, V.; Natalotto, A.; Giannetto, A.; Barreca, D.; Brunelli, E.; Mauceri, A.; Fasulo, S. Developmental abnormalities and neurotoxicological effects of CuO NPs on the black sea urchin Arbacia lixula by embryotoxicity assay. Mar. Environ. Res. 2015, 111, 121–127. [Google Scholar] [CrossRef]
- Cappello, T.; Vitale, V.; Oliva, S.; Villari, V.; Mauceri, A.; Fasulo, S.; Maisano, M. Alteration of neurotransmission and skeletogenesis in sea urchin Arbacia lixula embryos exposed to copper oxide nanoparticles. Comp. Biochem. Physiol. C 2017, 199, 20–27. [Google Scholar] [CrossRef]
- Tsuji, J.S.; Maynard, A.D.; Howard, P.C.; James, J.T.; Lam, C.W.; Warheit, D.B.; Santamaria, A.B. Research strategies for safety evaluation of nanomaterials, part IV: Risk assessment of nanoparticles. Toxicol. Sci. 2006, 89, 42–50. [Google Scholar] [CrossRef]
- Jamuna Bai, A.; Ravishankar Rai, V. Environmental risk, human health, and toxic effects of nanoparticles. Nanomater. Environ. Protect. 2014, 523. [Google Scholar]
- Yah, C.S. The toxicity of Gold Nanoparticles in relation to their physiochemical properties. Biomed. Res. 2013, 24, 400–413. [Google Scholar]
- Wu, T.; Tang, M. Review of the effects of manufactured nanoparticles on mammalian target organs. J. Appl. Toxicol. 2018, 38, 25–40. [Google Scholar] [CrossRef]
- Sajid, M.; Ilyas, M.; Basheer, C.; Tariq, M.; Daud, M.; Baig, N.; Shehzad, F. Impact of nanoparticles on human and environment: Review of toxicity factors, exposures, control strategies, and future prospects. Environ. Sci. Pollut. Res. 2015, 22, 4122–4143. [Google Scholar] [CrossRef]
- Joris, F.; Manshian, B.B.; Peynshaert, K.; De Smedt, S.C.; Braeckmans, K.; Soenen, S.J. Assessing nanoparticle toxicity in cell-based assays: Influence of cell culture parameters and optimized models for bridging the in vitro–in vivo gap. Chem. Soc. Rev. 2013, 42, 8339–8359. [Google Scholar] [CrossRef]
- Verma, A.; Stellacci, F. Effect of surface properties on nanoparticle–cell interactions. Small 2010, 6, 12–21. [Google Scholar] [CrossRef]
- Yildirimer, L.; Thanh, N.T.; Loizidou, M.; Seifalian, A.M. Toxicology and clinical potential of nanoparticles. Nano Today 2011, 6, 585–607. [Google Scholar] [CrossRef]
- Mu, Q.; Jiang, G.; Chen, L.; Zhou, H.; Fourches, D.; Tropsha, A.; Yan, B. Chemical basis of interactions between engineered nanoparticles and biological systems. Chem. Rev. 2014, 114, 7740–7781. [Google Scholar] [CrossRef]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular uptake of nanoparticles: Journey inside the cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef]
- Wisniewski, P.; Romano, R.M.; Kizys, M.M.L.; Oliveira, K.; Kasamatsu, T.; Giannocco, G.; Chiamolera, M.I.; Dias-da-Silva, M.; Romano, M.A. Adult exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of the hypothalamic-pituitary-testicular axis. Toxicology 2015, 329, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hougaard, K.S.; Campagnolo, L.; Chavatte-Palmer, P.; Tarrade, A.; Rousseau-Ralliard, D.; Valentino, S.; Park, M.V.D.Z.; de Jong, W.H.; Wolterink, G.; Piersma, A.H.; et al. A perspective on the developmental toxicity of inhaled nanoparticles. Reprod. Toxicol. 2015, 56, 118–140. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Choi, Y.J.; Song, H.; Kim, J.H. Potential toxicity of engineered nanoparticles in mammalian germ cells and developing embryos: Treatment strategies and anticipated applications of nanoparticles in gene delivery. Hum. Reprod. Update 2016, 22, 588–619. [Google Scholar] [CrossRef] [PubMed]
- Azenabor, A.; Ekun, A.O.; Akinloye, O. Impact of inflammation on male reproductive tract. J. Reprod. Infertil. 2015, 16, 123. [Google Scholar]
- Walczak–Jedrzejowska, R.; Wolski, J.K.; Slowikowska–Hilczer, J. The role of oxidative stress and antioxidants in male fertility. Cent. Eur. J. Urol. 2013, 66, 60. [Google Scholar] [CrossRef]
- Ren, C.; Hu, X.; Zhou, Q. Graphene oxide quantum dots reduce oxidative stress and inhibit neurotoxicity in vitro and in vivo through catalase-like activity and metabolic regulation. Adv. Sci. 2018, 5, 1700595. [Google Scholar] [CrossRef] [PubMed]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Han, J.W.; Jeong, J.K.; Gurunathan, S.; Choi, Y.J.; Das, J.; Kwon, D.N.; Cho, S.G.; Park, C.; Seo, H.G.; Park, J.K.; et al. Male-and female-derived somatic and germ cell-specific toxicity of silver nanoparticles in mouse. Nanotoxicology 2016, 10, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Ema, M.; Hougaard, K.S.; Kishimoto, A.; Honda, K. Reproductive and developmental toxicity of carbon-based nanomaterials: A literature review. Nanotoxicology 2016, 10, 391–412. [Google Scholar] [CrossRef]
- Opris, R.; Toma, V.; Olteanu, D.; Baldea, I.; Baciu, A.M.; Lucaci, F.I.; Berghian-Sevastre, A.; Tatomir, C.; Moldovan, B.; Clichici, S.; et al. Effects of silver nanoparticles functionalized with Cornus mas L. extract on architecture and apoptosis in rat testicle. Nanomedicine 2019, 14, 275–299. [Google Scholar] [CrossRef]
- Ogunsuyi, O.M.; Ogunsuyi, O.I.; Akanni, O.; Alabi, O.A.; Alimba, C.G.; Adaramoye, O.A.; Cambier, S.; Eswara, S.; Gutleb, A.C.; Bakare, A.A. Alteration of sperm parameters and reproductive hormones in Swiss mice via oxidative stress after co-exposure to titanium dioxide and zinc oxide nanoparticles. Andrologia 2020, 52, e13758. [Google Scholar] [CrossRef] [PubMed]
- Al-Bairuty, G.A.; Taha, M.N. Effects of copper nanoparticles on reproductive organs of male albino rats. Int. J. Sci. Technol. 2016, 143, 201617. [Google Scholar] [CrossRef]
- Kong, L.; Hu, W.; Lu, C.; Cheng, K.; Tang, M. Mechanisms underlying nickel nanoparticle induced reproductive toxicity and chemo-protective effects of vitamin C in male rats. Chemosphere 2019, 218, 259–265. [Google Scholar] [CrossRef]
- Deonandan, R.; Jaleel, M. Global decline in semen quality: Ignoring the developing world introduces selection bias. Int. J. Gen. Med. 2012, 5, 303. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Li, B.; Xu, K.; Liu, D.; Hu, J.; Yang, Y.; Nie, H.; Fan, L.; Zhu, W. Decline in semen quality among 30,636 young Chinese men from 2001 to 2015. Fertil. Steril. 2017, 107, 83–88. [Google Scholar] [CrossRef]
- Auger, J.; Kunstmann, J.M.; Czyglik, F.; Jouannet, P. Decline in semen quality among fertile men in Paris during the past 20 years. New Eng. J. Med. 1995, 332, 281–285. [Google Scholar] [CrossRef]
- Lan, Z.; Yang, W.X. Nanoparticles and spermatogenesis: How do nanoparticles affect spermatogenesis and penetrate the blood–testis barrier. Nanomedicine 2012, 7, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.G.; Yang, R.J.; Yue, W.B.; Xun, W.J.; Zhang, C.X.; Ren, Y.S.; Shi, L.; Lei, F.L. Effect of elemental nano-selenium on semen quality, glutathione peroxidase activity, and testis ultrastructure in male Boer goats. Anim. Reprod. Sci. 2010, 118, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Bae, E.; Yi, J.; Kim, Y.; Choi, K.; Lee, S.H.; Yoon, J.; Lee, B.C.; Park, K. Repeated-dose toxicity and inflammatory responses in mice by oral administration of silver nanoparticles. Environ. Toxicol. Pharmacol. 2010, 30, 162–168. [Google Scholar] [CrossRef]
- Shittu, O.K.; Aaron, S.Y.; Oladuntoye, M.D.; Lawal, B. Diminazene aceturate modified nanocomposite for improved efficacy in acute trypanosome infection. J. Acute Dis. 2018, 7, 36. [Google Scholar]
- Singh, R.; Singh, K. (Eds.) Male Infertility: Understanding, Causes and Treatment; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Jimenez-Reina, L.; Maartens, P.J.; Jimena-Medina, I.; Agarwal, A.; du Plessis, S.S. Overview of the Male Reproductive System. In Exercise and Human Reproduction; Springer: New York, NY, USA, 2016; pp. 1–17. [Google Scholar]
- Cooper, T.G.; Noonan, E.; Von Eckardstein, S.; Auger, J.; Baker, H.W.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization reference values for human semen characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Wang, C.; Swerdloff, R.S. Limitations of semen analysis as a test of male fertility and anticipated needs from newer tests. Fertil. Steril. 2014, 102, 1502–1507. [Google Scholar] [CrossRef]
- Skakkebaek, N.E.; Rajpert-De Meyts, E.; Buck Louis, G.M.; Toppari, J.; Andersson, A.M.; Eisenberg, M.L.; Jensen, T.K.; Jorgensen, N.; Swan, S.H.; Sapra, K.J.; et al. Male reproductive disorders and fertility trends: Influences of environment and genetic susceptibility. Physiol. Rev. 2016, 96, 55–97. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Gaskins, A.J.; Williams, P.L.; Mendiola, J.; Jørgensen, N.; Levine, H.; Hauser, R.; Swan, S.H.; Chavarro, J.E. Intake of fruits and vegetables with low-to-moderate pesticide residues is positively associated with semen-quality parameters among young healthy men. J. Nutr. 2016, 146, 1084–1092. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s second scientific statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Jensen, T.K.; Heitmann, B.L.; Jensen, M.B.; Halldorsson, T.I.; Andersson, A.M.; Skakkebæk, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Dalgard, C.; et al. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2013, 97, 411–418. [Google Scholar] [CrossRef]
- Nordkap, L.; Jensen, T.K.; Hansen, Å.M.; Lassen, T.H.; Bang, A.K.; Joensen, U.N.; Jensen, M.B.; Skakkebaek, N.E.; Jørgensen, N. Psychological stress and testicular function: A cross-sectional study of 1,215 Danish men. Fertil. Steril. 2016, 105, 174–187. [Google Scholar] [CrossRef]
- Sharma, R.; Harlev, A.; Agarwal, A.; Esteves, S.C. Cigarette smoking and semen quality: A new meta-analysis examining the effect of the 2010 World Health Organization laboratory methods for the examination of human semen. European Urol. 2016, 70, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Nordkap, L.; Joensen, U.N.; Jensen, M.B.; Jørgensen, N. Regional differences and temporal trends in male reproductive health disorders: Semen quality may be a sensitive marker of environmental exposures. Mol. Cell. Endocrinol. 2012, 355, 221–230. [Google Scholar] [CrossRef]
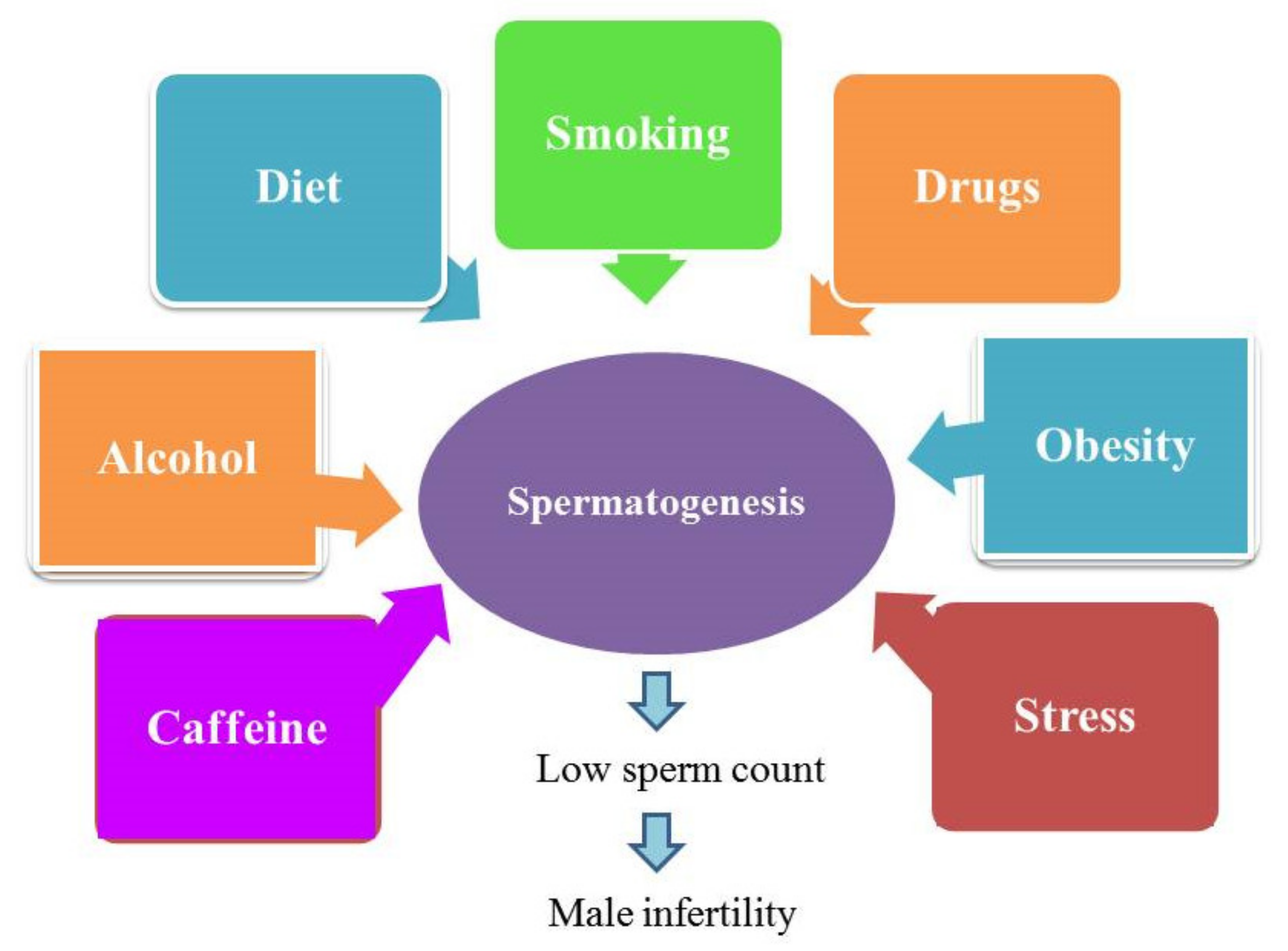
- Durairajanayagam, D. Lifestyle causes of male infertility. Arab J. Urol. 2018, 16, 10–20. [Google Scholar] [CrossRef]
- Coder, P.S.; Sloter, E.D.; Stump, D.G.; Nemec, M.D.; Bowman, C.J. Evaluation of a male reproductive toxicant. Comprehen. Toxicol. 2010, 11, 61–86. [Google Scholar]
- Lipskind, S.T.; Gargiulo, A.R. Computer-assisted laparoscopy in fertility preservation and reproductive surgery. J. Minim. Invasive Gynecol. 2013, 20, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Barkalina, N.; Charalambous, C.; Jones, C.; Coward, K. Nanotechnology in reproductive medicine: Emerging applications of nanomaterials. Nanomed. Nanotech. Biol. Med. 2014, 10, e921–e938. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Z.; Bagalkot, V.; Vasilliou, C.C.; Gu, F.; Alexis, F.; Zhang, L.; Shaikh, M.; Yuet, K.; Langer, R.; Kantoff, P.W.; et al. Superparamagnetic iron oxide nanoparticle–aptamer bioconjugates for combined prostate cancer imaging and therapy. Chem. Med. Chem. 2008, 3, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Thoeny, H.C.; Triantafyllou, M.; Birkhaeuser, F.D.; Froehlich, J.M.; Tshering, D.W.; Binser, T.; Fleischmann, A.; Vermathen, P.; Studer, U.E. Combined ultrasmall superparamagnetic particles of iron oxide–enhanced and diffusion-weighted magnetic resonance imaging reliably detect pelvic lymph node metastases in normal-sized nodes of bladder and prostate cancer patients. European Urol. 2009, 55, 761–769. [Google Scholar] [CrossRef]
- Kim, D.; Jeong, Y.Y.; Jon, S. A drug-loaded aptamer−gold nanoparticle bioconjugate for combined CT imaging and therapy of prostate cancer. ACS Nano 2010, 4, 3689–3696. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.F.; de Leon, P.M.M.; Komninou, E.R.; Dellagostin, O.A.; Deschamps, J.C.; Seixas, F.K.; Collares, T. NanoSMGT: Transgene transmission into bovine embryos using halloysite clay nanotubes or nanopolymer to improve transfection efficiency. Theriogenology 2011, 76, 1552–1560. [Google Scholar] [CrossRef]
- Yang, P.T.; Hoang, L.; Jia, W.W.; Skarsgard, E.D. In utero gene delivery using chitosan-DNA nanoparticles in mice. J. Surg. Res. 2011, 171, 691–699. [Google Scholar] [CrossRef]
- Kunzmann, A.; Andersson, B.; Thurnherr, T.; Krug, H.; Scheynius, A.; Fadeel, B. Toxicology of engineered nanomaterials: Focus on biocompatibility, biodistribution and biodegradation. Biochim. Biophys. Acta (BBA) Gen. Subj. 2011, 1810, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Taylor, U.A.W.E.; Barchanski, A.; Garrels, W.; Klein, S.; Kues, W.; Barcikowski, S.; Rath, D. Toxicity of gold nanoparticles on somatic and reproductive cells. In Nano-Biotechnology for Biomedical and Diagnostic Research; Springer: Dordrecht, The Netherlands, 2012; pp. 125–133. [Google Scholar]
- Taylor, U.; Petersen, S.; Barchanski, A.; Mittag, A.; Barcikowski, S.; Rath, D. Influence of gold nanoparticles on vitality parameters of bovine spermatozoa. In Reproduction in Domestic Animals; Wiley-Blackwell Publishing, Inc.: Hoboken, NJ, USA, 2010; Volume 45, p. 60. [Google Scholar]
- Wiwanitkit, V.; Sereemaspun, A.; Rojanathanes, R. Effect of gold nanoparticles on spermatozoa: The first world report. Fertil. Steril. 2009, 91, e7–e8. [Google Scholar] [CrossRef]
- Skuridin, S.G.; Dubinskaya, V.A.; Rudoy, V.M.; Dement’eva, O.V.; Zakhidov, S.T.; Marshak, T.L.; Evdokimov, Y.M. Effect of gold nanoparticles on DNA package in model systems. In Doklady Biochemistry and Biophysics; Springer Nature AG: Cham, Switzerland, 2010; Volume 432, p. 141. [Google Scholar]
- Niska, K.; Zielinska, E.; Radomski, M.W.; Inkielewicz-Stepniak, I. Metal nanoparticles in dermatology and cosmetology: Interactions with human skin cells. Chem. Biol. Interact. 2018, 295, 38–51. [Google Scholar] [CrossRef]
- Olugbodi, J.O.; David, O.; Oketa, E.N.; Lawal, B.; Okoli, B.J.; Mtunzi, F. Silver nanoparticles stimulates spermatogenesis impairments and hematological alterations in testis and epididymis of male rats. Molecules 2020, 25, 1063. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, R.F.; Singh, T.R.R.; Morrow, D.I.; Woolfson, A.D. Microneedle-Mediated Transdermal and Intradermal Drug Delivery; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 1–216. [Google Scholar]
- Filon, F.L.; Mauro, M.; Adami, G.; Bovenzi, M.; Crosera, M. Nanoparticles skin absorption: New aspects for a safety profile evaluation. Regul. Toxicol. Pharmacol. 2015, 72, 310–322. [Google Scholar] [CrossRef]
- Rancan, F.; Gao, Q.; Graf, C.; Troppens, S.; Hadam, S.; Hackbarth, S.; Kembuan, C.; Blume-Peytavi, U.; Ruhl, E.; Lademann, J.; et al. Skin penetration and cellular uptake of amorphous silica nanoparticles with variable size, surface functionalization, and colloidal stability. ACS Nano 2012, 6, 6829–6842. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 2012, 15, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Bergin, I.L.; Witzmann, F.A. Nanoparticle toxicity by the gastrointestinal route: Evidence and knowledge gaps. Int. J. Biomed. Nanosci. Nanotechnol. 2013, 3, 163–210. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Wang, Y.; Zhou, Y.; Zhang, Q.; Ge, Y.; Chen, M.; Hong, J.; Wang, L. Exposure to TiO2 nanoparticles induces immunological dysfunction in mouse testitis. J. Agric. Food Chem. 2016, 64, 346–355. [Google Scholar] [CrossRef]
- Miresmaeili, S.M.; Halvaei, I.; Fesahat, F.; Fallah, A.; Nikonahad, N.; Taherinejad, M. Evaluating the role of silver nanoparticles on acrosomal reaction and spermatogenic cells in rat. Iran. J. Reprod. Med. 2013, 11, 423. [Google Scholar]
- Sleiman, H.K.; Romano, R.M.; Oliveira, C.A.D.; Romano, M.A. Effects of prepubertal exposure to silver nanoparticles on reproductive parameters in adult male Wistar rats. J. Toxicol. Environ. Health A 2013, 76, 1023–1032. [Google Scholar] [CrossRef]
- Dekkers, S.; Krystek, P.; Peters, R.J.; Lankveld, D.P.; Bokkers, B.G.; van Hoeven-Arentzen, P.H.; Bouwmeester, H.; Oomen, A.G. Presence and risks of nanosilica in food products. Nanotoxicology 2011, 5, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Winkler, H.C.; Suter, M.; Naegeli, H. Critical review of the safety assessment of nano-structured silica additives in food. J. Nanobiotechnol. 2016, 14, 44. [Google Scholar] [CrossRef]
- Rincker, M.J.; Hill, G.M.; Link, J.E.; Meyer, A.M.; Rowntree, J.E. Effects of dietary zinc and iron supplementation on mineral excretion, body composition, and mineral status of nursery pigs. J. Anim. Sci. 2005, 83, 2762–2774. [Google Scholar] [CrossRef]
- Rim, K.T.; Song, S.W.; Kim, H.Y. Oxidative DNA damage from nanoparticle exposure and its application to workers’ health: A literature review. Saf. Health Work 2013, 4, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.; Halappanavar, S.; Hougaard, K.S.; Williams, A.; Madsen, A.M.; Lamson, J.S.; Andersen, O.; Yauk, C.; Wallin, K.; Vogel, U. Maternal inhalation of surface-coated nanosized titanium dioxide (UV-Titan) in C57BL/6 mice: Effects in prenatally exposed offspring on hepatic DNA damage and gene expression. Nanotoxicology 2013, 7, 85–96. [Google Scholar] [CrossRef]
- Bakand, S.; Hayes, A. Toxicological considerations, toxicity assessment, and risk management of inhaled nanoparticles. Int. J. Mol. Sci. 2016, 17, 929. [Google Scholar] [CrossRef]
- Meier, M.J.; O’Brien, J.M.; Beal, M.A.; Allan, B.; Yauk, C.L.; Marchetti, F. In utero exposure to benzo [a] pyrene increases mutation burden in the soma and sperm of adult mice. Environ. Health Perspect. 2017, 125, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Taneda, S.; Taya, K.; Watanabe, G.; Li, X.; Fujitani, Y.; Ito, Y.; Nakajima, T.; Suzuki, A.K. Effects of inhaled nanoparticle-rich diesel exhaust on regulation of testicular function in adult male rats. Inhal. Toxicol. 2009, 21, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Boisen, A.M.Z.; Shipley, T.; Jackson, P.; Wallin, H.; Nellemann, C.; Vogel, U.; Yauk, C.L.; Hougaard, K.S. In utero exposure to nanosized carbon black (Printex90) does not induce tandem repeat mutations in female murine germ cells. Reprod. Toxicol. 2013, 41, 45–48. [Google Scholar] [CrossRef]
- Skovmand, A.; Jensen, A.C.; Maurice, C.; Marchetti, F.; Lauvås, A.J.; Koponen, I.K.; Jensen, K.A.; Goericke-Pesch, S.; Vogel, U.B.; Hougaard, K.S. Effects of maternal inhalation of carbon black nanoparticles on reproductive and fertility parameters in a four-generation study of male mice. Part. Fibre Toxicol. 2019, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Mostafalou, S.; Mohammadi, H.; Ramazani, A.; Abdollahi, M. Different biokinetics of nanomedicines linking to their toxicity; an overview. Daru J. Pharm. Sci. 2013, 21, 14. [Google Scholar] [CrossRef]
- Wang, R.; Song, B.; Wu, J.; Zhang, Y.; Chen, A.; Shao, L. Potential adverse effects of nanoparticles on the reproductive system. Int. J. Nanomed. 2018, 13, 8487–8506. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Suzuki, A.K.; Zhang, Y.; Fujitani, Y.; Nagaoka, K.; Watanabe, G.; Taya, K. Effects of exposure to nanoparticle-rich diesel exhaust on pregnancy in rats. J. Reprod. Dev. 2013, 59, 14–150. [Google Scholar] [CrossRef]
- Rollerova, E.; Jurcovicova, J.; Mlynarcikova, A.; Sadlonova, I.; Bilanicova, D.; Wsolova, L.; Kiss, A.; Kovriznych, J.; Kronek, J.; Ciampor, F.; et al. Delayed adverse effects of neonatal exposure to polymeric nanoparticle poly (ethylene glycol)-block-polylactide methyl ether on hypothalamic–pituitary–ovarian axis development and function in Wistar rats. Reprod. Toxicol. 2015, 57, 165–175. [Google Scholar] [CrossRef]
- Hussein, M.M.; Ali, H.A.; Saadeldin, I.M.; Ahmed, M.M. Querectin alleviates zinc oxide nanoreprotoxicity in male albino rats. J. Biochem. Mol. Toxicol. 2016, 30, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Gu, W.; Ye, L.; Yang, H. Biodistribution of PAMAM dendrimer conjugated magnetic nanoparticles in mice. J. Mater. Sci. Mater. Med. 2014, 25, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Sundarraj, K.; Manickam, V.; Raghunath, A.; Periyasamy, M.; Viswanathan, M.P.; Perumal, E. Repeated exposure to iron oxide nanoparticles causes testicular toxicity in mice. Environ. Toxicol. 2017, 32, 594–608. [Google Scholar] [CrossRef]
- Kielbik, P.; Kaszewski, J.; Dabrowski, S.; Faundez, R.; Witkowski, B.S.; Wachnicki, L.; Zhydachevskyy, Y.; Sapierzynski, R.; Gajewski, Z.; Godlewski, M.M. Transfer of orally administered ZnO: Eu nanoparticles through the blood–testis barrier: The effect on kinetic sperm parameters and apoptosis in mice testes. Nanotechnology 2019, 30, 455101. [Google Scholar] [CrossRef]
- Gao, G.; Ze, Y.; Zhao, X.; Sang, X.; Zheng, L.; Ze, X.; Gui, S.; Sheng, L.; Sun, Q.; Hong, J.; et al. Titanium dioxide nanoparticle-induced testicular damage, spermatogenesis suppression, and gene expression alterations in male mice. J. Haz. Mater. 2013, 258, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, L.; Klein, J.P.; Forest, V.; Boudard, D.; Martini, M.; Pourchez, J.; Blanchin, M.G.; Cottier, M. Testicular biodistribution of silica-gold nanoparticles after intramuscular injection in mice. Biomed. Microdev. 2015, 17, 66. [Google Scholar] [CrossRef]
- Miura, N.; Ohtani, K.; Hasegawa, T.; Yoshioka, H.; Hwang, G.W. High sensitivity of testicular function to titanium nanoparticles. J. Toxicol. Sci. 2017, 42, 359–366. [Google Scholar] [CrossRef]
- Wang, Z.; Qu, G.; Su, L.; Wang, L.; Yang, Z.; Jiang, J.; Liu, S.; Jiang, G. Evaluation of the biological fate and the transport through biological barriers of nanosilver in mice. Curr. Pharm. Des. 2013, 19, 6691–6697. [Google Scholar] [CrossRef]
- Thakur, M.; Gupta, H.; Singh, D.; Mohanty, I.R.; Maheswari, U.; Vanage, G.; Joshi, D.S. Histopathological and ultra structural effects of nanoparticles on rat testis following 90 days (chronic study) of repeated oral administration. J. Nanobiotechnol. 2014, 12, 42. [Google Scholar] [CrossRef]
- Zhou, Q.; Yue, Z.; Li, Q.; Zhou, R.; Liu, L. Exposure to PbSe nanoparticles and male reproductive damage in a rat model. Environ. Sci. Technol. 2019, 53, 13408–13416. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Orekoya, B. Lipid profile and oxidative stress markers in rats following oral and repeated exposure to Fijk herbal mixture. J. Toxicol. 2014, 2014, 876035. [Google Scholar] [CrossRef]
- Kong, L.; Tang, M.; Zhang, T.; Wang, D.; Hu, K.; Lu, W.; Wei, C.; Liang, G.; Pu, Y. Nickel nanoparticles exposure and reproductive toxicity in healthy adult rats. Int. J. Mol. Sci. 2014, 15, 21253–21269. [Google Scholar] [CrossRef] [PubMed]
- Gromadzka-Ostrowska, J.; Dziendzikowska, K.; Lankoff, A.; Dobrzyńska, M.; Instanes, C.; Brunborg, G.; Gajowik, A.; Radzikowska, J.; Wojewodzka, M.; Kruszewski, M. Silver nanoparticles effects on epididymal sperm in rats. Toxicol. Lett. 2012, 214, 251–258. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, B.; Hong, W.; Chen, L.; Yao, L.; Zhao, Y.; Aguilar, Z.P.; Xu, H. ZnO nanoparticles induced male reproductive toxicity based on the effects on the endoplasmic reticulum stress signaling pathway. Int. J. Nanomed. 2019, 14, 9563. [Google Scholar] [CrossRef]
- Mahfouz, R.; Sharma, R.; Thiyagarajan, A.; Kale, V.; Gupta, S.; Sabanegh, E.; Agarwal, A. Semen characteristics and sperm DNA fragmentation in infertile men with low and high levels of seminal reactive oxygen species. Fertil. Steril. 2010, 94, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.H.M.; Ibrahim, I.A.; Shehata, M.A.; Mohammed, H.O. Histopathological and genetic study on the protective role of β-carotene on testicular tissue of adult male albino rats treated with titanium dioxide nanoparticles. Afr. J. Pharm. Pharmacol. 2020, 14, 9–19. [Google Scholar]
- Yousef, M.I. Reproductive toxicity of aluminum oxide nanoparticles and zinc oxide nanoparticles in male rats. Nanoparticle 2019, 1, 3. [Google Scholar]
- Ahmed, S.M.; Abdelrahman, S.A.; Shalaby, S.M. Evaluating the effect of silver nanoparticles on testes of adult albino rats (histological, immunohistochemical and biochemical study). J. Mol. Histol. 2017, 48, 9–27. [Google Scholar] [CrossRef] [PubMed]
- El-Azab, N.E.E.; Elmahalaway, A.M. A Histological and Immunohistochemical Study on Testicular Changes Induced by Sliver Nanoparticles in Adult Rats and the Possible Protective Role of Camel Milk. Egypt. J. Histol. 2020, 42, 1044–1058. [Google Scholar] [CrossRef]
- Mesallam, D.I.; Deraz, R.H.; Aal, S.M.A.; Ahmed, S.M. Toxicity of Subacute Oral Zinc Oxide Nanoparticles on Testes and Prostate of Adult Albino Rats and Role of Recovery. J. Histol. Histopathol. 2019, 6, 1–11. [Google Scholar] [CrossRef]
- Hu, W.; Yu, Z.; Gao, X.; Wu, Y.; Tang, M.; Kong, L. Study on the damage of sperm induced by nickel nanoparticle exposure. Environ. Geochem. Health 2020, 42, 1715–1724. [Google Scholar] [CrossRef]
- Qin, F.; Shen, T.; Li, J.; Qian, J.; Zhang, J.; Zhou, G.; Tong, J. SF-1 mediates reproductive toxicity induced by Cerium oxide nanoparticles in male mice. J. Nanobiotechnol. 2019, 17, 41. [Google Scholar] [CrossRef]
- Iyiola, O.; Olafimihan, T.F.; Sulaiman, F.A.; Anifowoshe, A.T. Genotoxicity and histopathological assessment of silver nanoparticles in Swiss albino mice. Cuad. Investig. UNED 2018, 10, 102–109. [Google Scholar] [CrossRef][Green Version]
- Lafuente, D.; Garcia, T.; Blanco, J.; Sánchez, D.J.; Sirvent, J.J.; Domingo, J.L.; Gómez, M. Effects of oral exposure to silver nanoparticles on the sperm of rats. Reprod. Toxicol. 2016, 60, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, Z.; Khorsandi, L.; Nejad-Dehbashi, F. Protective effect of zingerone against mouse testicular damage induced by zinc oxide nanoparticles. Environ. Sci. Pollut. Res. 2019, 26, 25814–25824. [Google Scholar] [CrossRef]
- Bakare, A.A.; Udoakang, A.J.; Anifowoshe, A.T.; Fadoju, O.M.; Ogunsuyi, O.I.; Alabi, O.A.; Alimba, C.G.; Oyeyemi, I.T. Genotoxicity of titanium dioxide nanoparticles using the mouse bone marrow micronucleus and sperm morphology assays. J. Pollut. Eff. Cont. 2016, 4, 41. [Google Scholar]
- Hamdi, H. Testicular dysfunction induced by aluminum oxide nanoparticle administration in albino rats and the possible protective role of the pumpkin seed oil. J. Basic Appl. Zool. 2020, 81, 42. [Google Scholar] [CrossRef]
- Smith, M.A.; Michael, R.; Aravindan, R.G.; Dash, S.; Shah, S.I.; Galileo, D.S.; Martin-DeLeon, P.A. Anatase titanium dioxide nanoparticles in mice: Evidence for induced structural and functional sperm defects after short-, but not long-, term exposure. As. J. Androl. 2015, 17, 261. [Google Scholar]
- Karimi, S.; Khorsandi, L.; Nejaddehbashi, F. Protective effects of curcumin on testicular toxicity induced by titanium dioxide nanoparticles in mice. JBRA Assist. Reprod. 2019, 23, 344. [Google Scholar] [CrossRef]
- Song, G.; Lin, L.; Liu, L.; Wang, K.; Ding, Y.; Niu, Q.; Mu, L.; Wang, H.; Shen, H.; Guo, S. Toxic effects of anatase titanium dioxide nanoparticles on spermatogenesis and testicles in male mice. Pol. J. Environ. Stud. 2017, 26, 2739–2745. [Google Scholar] [CrossRef]
- Yoshida, S.; Hiyoshi, K.; Ichinose, T.; Takano, H.; Oshio, S.; Sugawara, I.; Takeda, K.; Shibamoto, T. Effect of nanoparticles on the male reproductive system of mice. Int. J. Androl. 2009, 32, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Meena, R.; Kajal, K.; Paulraj, R. Cytotoxic and genotoxic effects of titanium dioxide nanoparticles in testicular cells of male Wistar rat. Appl. Biochem. Biotechnol. 2015, 175, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.M.; Ibrahim, M.A.; Noshy, P.A. Reproductive toxicity provoked by titanium dioxide nanoparticles and the ameliorative role of Tiron in adult male rats. Biochem. Biophys. Res. Commun. 2017, 486, 595–600. [Google Scholar] [CrossRef]
- Miura, N.; Ohtani, K.; Hasegawa, T.; Hojo, R.; Yanagiba, Y.; Suzuki, T.; Suda, M.; Wang, R.S. Hazardous effects of titanium dioxide nanoparticles on testicular function in mice. Fund. Toxicol. Sci. 2014, 1, 81–85. [Google Scholar] [CrossRef]
- Abdulla, I.T. Histological effects of titanium dioxide nanoparticles size 10 nm in mice testes. Sci. J. Univ. Zakho 2017, 5, 158–161. [Google Scholar] [CrossRef][Green Version]
- Mohammadi Fartkhooni, F.; Noori, A.; Momayez, M.; Sadeghi, L.; Shirani, K.; Yousefi Babadi, V. The effects of nano titanium dioxide (TiO2) in spermatogenesis in wistar rat. Euro J. Exp. Bio. 2013, 3, 145–149. [Google Scholar]
- Orazizadeh, M.; Khorsandi, L.; Absalan, F.; Hashemitabar, M.; Daneshi, E. Effect of beta-carotene on titanium oxide nanoparticles-induced testicular toxicity in mice. J. Assist. Reprod. Genet. 2014, 31, 561–568. [Google Scholar] [CrossRef]
- Miura, N.; Ohtani, K.; Hasegawa, T.; Yoshioka, H.; Hwang, G.W. Biphasic adverse effect of titanium nanoparticles on testicular function in mice. Sci. Rep. 2019, 9, 14373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Gurunathan, S.; Kim, J.H. Effects of silver nanoparticles on neonatal testis development in mice. Int. J. Nanomed. 2015, 10, 6243. [Google Scholar]
- Baki, M.E.; Miresmaili, S.M.; Pourentezari, M.; Amraii, E.; Yousefi, V.; Spenani, H.R.; Talebi, A.R.; Anvari, M.; Fazilati, M.; Fallah, A.S.; et al. Effects of silver nano-particles on sperm parameters, number of Leydig cells and sex hormones in rats. Iran. J. Reprod. Med. 2014, 12, 139–144. [Google Scholar] [PubMed]
- Garcia, T.X.; Costa, G.M.; França, L.R.; Hofmann, M.C. Sub-acute intravenous administration of silver nanoparticles in male mice alters Leydig cell function and testosterone levels. Reprod. Toxicol. 2014, 45, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.R.H. Studies on the genotoxicity behavior of silver nanoparticles in the presence of heavy metal cadmium chloride in mice. J. Nanomater. 2016, 2016, 5283162. [Google Scholar] [CrossRef]
- Mathias, F.T.; Romano, R.M.; Kizys, M.M.; Kasamatsu, T.; Giannocco, G.; Chiamolera, M.I.; Dias-da-Silva, M.R.; Romano, M.A. Daily exposure to silver nanoparticles during prepubertal development decreases adult sperm and reproductive parameters. Nanotoxicology 2015, 9, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Fathi, N.; Hoseinipanah, S.M.; Alizadeh, Z.; Assari, M.J.; Moghimbeigi, A.; Mortazavi, M.; Hosseini, H.; Bahmanzadeh, M. The effect of silver nanoparticles on the reproductive system of adult male rats: A morphological, histological and DNA integrity study. Adv. Clin. Experim. Med. 2019, 28, 299–305. [Google Scholar] [CrossRef]
- Salman, R.A. The influence of ZnO NPs on reproductive system tissues of albino male mice. Histopathological study. Int. J. Sci. Res. 2017, 6, 2021–2025. [Google Scholar]
- Talebi, A.R.; Khorsandi, L.; Moridian, M. The effect of zinc oxide nanoparticles on mouse spermatogenesis. J. Assist. Reprod. Genet. 2013, 30, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Abbasalipourkabir, R.; Moradi, H.; Zarei, S.; Asadi, S.; Salehzadeh, A.; Ghafourikhosroshahi, A.; Mortazavi, M.; Ziamajidi, N. Toxicity of zinc oxide nanoparticles on adult male Wistar rats. Food Chem. Toxicol. 2015, 84, 154–160. [Google Scholar] [CrossRef]
- Mozaffari, Z.; Parivar, K.; Roodbari, N.H.; Irani, S. Histopathological evaluation of the toxic effects of zinc oxide (ZnO) nanoparticles on testicular tissue of NMRI adult mice. Adv. Stud. Biol. 2015, 7, 275–291. [Google Scholar] [CrossRef]
- Yousefalizadegan, N.; Mousavi, Z.; Rastegar, T.; Razavi, Y.; Najafizadeh, P. Reproductive toxicity of manganese dioxide in forms of micro-and nanoparticles in male rats. Int. J. Reprod. BioMed. 2019, 17, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, N.K.; Awasthi, K.K.; John, P.J. Effects of nano-graphene oxide on testis, epididymis and fertility of Wistar rats. Basic Clin. Pharmacol. Toxicol. 2017, 121, 202–210. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, D.; Li, Q.; Deng, H.; Shen, J.; Zheng, G.; Sun, M. Rat testis damage caused by lead sulfide nanoparticles after oral exposure. J. Nanosci. Nanotechnol. 2016, 16, 2378–2383. [Google Scholar] [CrossRef] [PubMed]
- Nazar, M.; Talebi, A.R.; Sharifabad, M.H.; Abbasi, A.; Khoradmehr, A.; Danafar, A.H. Acute and chronic effects of gold nanoparticles on sperm parameters and chromatin structure in Mice. Int. J. Reprod. BioMed. 2016, 14, 637. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, O.A.; Akinloye, O.; Adaramoye, O.A. Cerium oxide nanoparticle elicits oxidative stress, endocrine imbalance and lowers sperm characteristics in testes of balb/c mice. Andrologia 2018, 50, e12920. [Google Scholar] [CrossRef]
- Lauvas, A.J.; Skovmand, A.; Poulsen, M.S.; Kyjovska, Z.O.; Roursgaard, M.; Goericke-Pesch, S.; Vogel, U.; Hougaard, K.S. Airway exposure to TiO2 nanoparticles and quartz and effects on sperm counts and testosterone levels in male mice. Reprod. Toxicol. 2019, 90, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Behnammorshedi, M.; Nazem, H.; Moghadam, M.S. The effect of gold nanoparticle on luteinizing hormone, follicle stimulating hormone, testosterone and testis in male rat. Biomed. Res. 2015, 26, 348–352. [Google Scholar]
- Khorsandi, L.; Orazizadeh, M.; Moradi-Gharibvand, N.; Hemadi, M.; Mansouri, E. Beneficial effects of quercetin on titanium dioxide nanoparticles induced spermatogenesis defects in mice. Environ. Sci. Pollut. Res. 2017, 24, 5595–5606. [Google Scholar] [CrossRef]
- Pinho, A.R.; Rebelo, S.; Pereira, M.L. The impact of zinc oxide nanoparticles on male (in)fertility. Materials 2020, 13, 849. [Google Scholar] [CrossRef]
- Santonastaso, M.; Mottola, F.; Colacurci, N.; Iovine, C.; Pacifico, S.; Cammarota, M.; Cesaroni, F.; Rocco, L. In vitro genotoxic effects of titanium dioxide nanoparticles (n-TiO2) in human sperm cells. Mol. Reprod. Develop. 2019, 86, 1369–1377. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Values | 95% CI |
|---|---|---|
| Volume | 1.5 mL | 1.4–1.7 |
| Sperm concentration | 15 million spermatozoa/mL | 12–16 |
| Total number of sperm per ejaculation | 39 million spermatozoa | 33–46 |
| Sperm Morphology | 4% normal forms | 3–4 |
| Sperm vitality | 58% live | 55–63 |
| Progressive motility | 32% | 31–34 |
| Total (progressive and nonprogressive sperm motility) | 40% | 38–42 |
| Test Material | Histological Evaluation | References | |
|---|---|---|---|
| Nanoparticles | Sperm Morphology | Seminiferous Tubules | |
| Nickel nanoparticles | Increased number of abnormal sperms in epididymis, cell apoptosis, no proper arrangement of germinal cells, large gap in lumen of seminiferous tubules | [108] | |
| Silver nanoparticles | Different sperm cell abnormalities including double head, long tail, No hook or wrong hook attachment | Adverse hypertrophic seminiferous tubules | [110] |
| Development of abnormal spermatids | Atrophy in seminiferous tubules, necrosis and degradation of spermatogenic cells, and in spermatogonia and Sertoli cells, ultra-structural alterations | [96] | |
| Shrunken seminiferous tubules, loss of sperms in seminiferous tubules, presence of multinucleated giant cells | [105] | ||
| Sperm with coiled, bent and headless tail, detached head | Increased desquamation in the lumen | [111] | |
| Zinc oxide nanoparticles | Detachment (D), sloughing (S), vacuoles (V) in seminiferous tubules, loss of spermatids, disorganization of germ cells, vacuolization in germinal epithelium | [112] | |
| Titanium dioxide nanoparticles | Amorphous head, double tails of sperm, double head with fused tails, short and knobbed hook | Depletion and necrosis in spermatogenic cells, vacuolation | [113] |
| Zinc oxide nanoparticles and titanium dioxide nanoparticles | Massive head, double hook, double tail with pin head, folded spermatozoa | Seminiferous tubules with variation in size, depletion in spermatogenic cells, necrosis in spermatogenic cells, increased luminal width, congestion in interstitial blood vessels | [28] |
| Cerium oxide nanoparticles | Necrosis in seminiferous tubules, apoptosis in interstitial tissues, loss of spermatozoa, decline in the number of Sertoli cells, Leydig cells, and spermatids | [109] | |
| Silica-gold nanoparticles | Empty seminiferous tubules | [93] | |
| Aluminum oxide nanoparticle | Vacuolization, edema in interstitial cells and congestion in blood vessels, necrosis in spermatogenic cells | [114] | |
| Anatase titanium dioxide nanoparticles | Coil and folded sperm with missing cap | [115] | |
| Test Material | Experimental Model | Age | Exposure of NPs | Findings | Reference | |||
|---|---|---|---|---|---|---|---|---|
| Size | Route | Doses | Days | |||||
| Carbon black nanoparticles | ICR male mice | 14, 56, 95 nm | Intratracheally | 0.1 mg | 10 times/week | T level increase (14, 56 nm) DSP decreased | [118] | |
| Titanium dioxide nanoparticles | Male Wistar rats | 21 nm | Intravenously | TSH decreased, Sperm count reduced | [119] | |||
| Adult male albino rats | 180–200 g | Orally | 100 mg/kg/day | 8 weeks | Increased sperm abnormalities, motility, curved sperm tail, decreased sperm viability, sperm count and T level | [120] | ||
| C57BL/6Jgpt delta mice | 8 weeks | Intravenously | 0, 2, 10 mg/kg/BW/week | 4 weeks Kill 9 days after last injection | Number of sperm head reduced in testes and cauda epididymis, toxic effect on sperm quality | [121] | ||
| Albino male mice | 4 months | 10 nm | Intraperitoneally | 5, 10, 50, 100, 150 mg/kg/BW/day | 14 days | [122] | ||
| Adult male Wistar rats | 2 months 150–250 g | 18 nm | Intraperitoneally | 1 mL of 30, 50 mg/kg/dos | Alternate days for 21 days | Decreased T level, increased LH, no change in FSH | [123] | |
| NMRI mice | 6–8 weeks 25–30 g | 300 mg/kg | 35 days | T level decreased, sperm motility and number changed | [124] | |||
| Adult Swiss male mice | 11–15 weeks 28–32 g | <25 nm | Intraperitoneally | TiO2NPs: 9.38, 18.75, 37.5, 75 mg/kg b.w | Daily for 35 days | Changes in sperm motility, number, increased abnormalities, reduced LH level | [28] | |
| Titanium nanoparticles | MaleC57BL/6J | 8 weeks | Intravenously | 10, 50 mg/kg (single injection) | 1 3 9 days | Decreased sperm motility, sperm number not reduced in cauda epididymis and testes, blockage of blood vessel at higher dose (50 mg) After 3 days oral administration: Sperm motility reduced, no change in spermatozoa in cauda epididymis | [125] | |
| Orally | 20, 100 mg/kg (single dose) | |||||||
| Silver nanoparticles | Male Wistar rats | Postnatal day PND 23 | 60 nm | Orally | 15, 50 µg/kg BW | Once/day PND 53 PND 90 | PND 90: Reduction in sperm count and TSH PND 53 and 90: sperm reserve in epididymis | [73] |
| Male Wistar rats | 200–250 g 45–50 days | 70 nm | Orally | 25, 50, 100, 200 mg/kg/day | Every 12 h in 48 days | Decrease in number of spermatogenic cells at 200 mg/kg dose, primary spermatocytes, spermatids and spermatozoa reduced at dose of 50, 100, 200 mg/kg | [72] | |
| Male Wistar rats | 14 weeks | 20 nm, 200 nm | intravenously | 5, 10 mg/kg body mass (20 nm) or 5 mg/kg (200 nm) | 24 h, 7 days, 28 days | Reduction in sperm count, increased abnormal spermatozoa after 1–4 weeks than 24 h | [100] | |
| Male ICR mice | 8 days old | 15 nm | Sub-cutaneously | 1, 5 mg/kg/dose (5 doses for every 3 days) | PND 28, PND 42, PND 63, PND 100 | On PND 42 and 63: Sperm abnormalities increased at 5 mg On PND 100: Epididymal level of sperm decreased at 5 mg | [126] | |
| Male rats | 100–150 g | 100 nm | Sub-dermally | 0, 10, 50 mg/kg BW | 7 days, 28 days | Dose and time dependent changes in T, LH and FSH Decrease in sperm velocity parameters | [65] | |
| Male Wistar rats | 70 nm | Orally | 25, 50, 100, 200 mg/kg/day | 45 days | Reduction in number of Leydig cells, T level, FSH, sperm motility, number of immotile sperms increased in a dose dependent manner | [127] | ||
| Male CD1 mice | 4–5 weeks | 10 nm | Intravenously | 1 mg/kg 5 times (once every 3 days) | 15, 60 and 120 days | No change in sperm count, motility and fertility indices, T level increased on day 15 | [128] | |
| Male Swiss Webster mice | 30–35 g | 56.67 ± 9.77 nm | Intraperitoneally | 20, 41, 82 mg/kg | After 24 h from injection | Decreased concentration of sperm, increased number of abnormal sperms at all doses | [129] | |
| Male Wistar rats | 10–12 weeks | 5-20 nm | Orally | 20 µg/kg/day | 90 days | No changes | [96] | |
| Male Wistar rats | 60 nm | Orally | 15, 30 µg/kg/day | PND 23, PND 58 | At both doses: Sperm abnormalities increased, no changes in T, LH and FSH | [130] | ||
| Male adult Wistar rats | 200–250 g | 60–80 nm | Intraperitoneally | 30, 125, 300 mg/kg | Single dose | Sperm count and vitality reduced, decreased spermatogonia, Leydig and Sertoli cells | [131] | |
| Polyvinil pyrrolidone coated silver nanoparticles | Adult male Sprague Dawley rats | 240–280 g | 20–30 nm | Orally | 50, 100, 200 mg/kg/day | 90 days | No significant changes on sperm motility At 100 mg: Number of abnormal sperms increased | [111] |
| Zinc oxide nanoparticles | Adult albino mice | 3 weeks | 80 nm | Orally | 0, 150, 350 mg/kg/BW | 15 days | [132] | |
| Adult male NMRI mice | Orally | 0, 5, 50, 300 mg/kg/day | 35 days | At 50 and 300 mg: Significant alterations in sperm number, sperm motility and abnormality of epididymal sperms | [133] | |||
| Adult male Wistar rats | 6–8 weeks 180–220 g | 10–30 nm | Intraperitoneally | 0, 50, 100, 150, 200 mg/kg/BW/day | 10 days | More than 50 mg: Viability and number of sperms reduced More than 100: Quality of sperms decreased | [134] | |
| Mature NMRI mice | 28–32 g | 20 nm | Intraperitoneally | 0, 250, 500, 700 mg/kg/day | 1 week after single injection | No change | [135] | |
| Wistar Male albino rats | 4–5 months 160–170 g | 100 nm | Orally | 100 mg/kgBW/day | 75 days | Number of sperms and motility decreased, reduced T while LH and FSH increased | [104] | |
| Adult Swiss male mice | 11–15 weeks 28–32 g | <100 nm | Intraperitoneally | 9.38, 18.75, 37.5, 75 mg/kg b.w/day | 35 days | TSH level increased | [28] | |
| Manganese dioxide nanoparticles | Male Wistar rats | 180 ± 5 g 8–10 weeks | <5 mm 25–85 nm | Sub-cutaenously | 100 mg/kg | Once a week for 4 weeks | At both doses: Increased number of immotile sperms, decreased number of epididymal sperm, spermatogonia and spermatocytes, no change in T and FSH | [136] |
| Graphene Oxide nanoparticles | Adult Wistar rats | 10–12 weeks | 0.8–2 nm | Intraperitonealy | 0.4, 2 and 10 mg/kg BW | Repeated exposure for 15 and 30 days | Reduced epididymal sperm count, motility, increased sperm abnormality | [137] |
| Nickel nanoparticle | Sprague Dawley rats | 80–100 g | 90 nm | Orally | 0, 5, 15, 45 mg/kg | 10 weeks | Decreased FSH, T but LH increased, sperm motility reduced | [99] |
| Lead sulphide nanoparticles | Sprague Dawley rats | 180–240 g) 12–14 weeks | 30 nm | Orally | 0, 25, 50, 100 mg/kg | 5 days/week for 6 weeks | Decreased sperm count and rate of sperm survival | [138] |
| Lead selenide nanoparticles | Male specific pathogen free Sprague Dawley rats | 170–200 g 6–7 weeks | 8 nm, 30 nm, 70 nm | Intraperitoneally | 10 mg/kg/week | 60 days | At 8 nm and 30 nm: Increased sperm abnormalities, poor quality and quantity of sperms, T and FSH more reduced in 8 nm as compared to 30 nm and 70 nm in a size dependent manner | [97] |
| Gold nanoparticles | Male bulb –c mice | 5 weeks | 10–30 nm | Intraperitoneally | 40 and 200 µg/kg/day | 7 and 35 days | Increased number of abnormal spermatozoa, reduced sperm motility and altered sperm morphology | [139] |
| Aluminium oxide nanoparticles | Wistar Male albino rats | 4–5 months 160–170 g | 50 nm, | Orally | 70 mg/kgBW/day | 75 days | Decreased semen characteristics: number of sperms and its motility, TSH, T, increased FSH, LH | [104] |
| Cerium oxide nanoparticles | Male bulb –c mice | 8 weeks 30–35 g | Less than 10 nm | Intraperitoneally | 100, 200 and 300 μg/kg body weight | Three times/week for 5 weeks | Decreased T, FSH and LH, reduced sperm count and motility, increased sperm abnormality | [140] |
| Titanium dioxide nanoparticles and α-quartz particles | Male mice C57BL/6J | 9 weeks | Intratracheally | 63 µg | Once weekly for seven weeks | No effect on sperm count and T level | [141] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iftikhar, M.; Noureen, A.; Uzair, M.; Jabeen, F.; Abdel Daim, M.; Cappello, T. Perspectives of Nanoparticles in Male Infertility: Evidence for Induced Abnormalities in Sperm Production. Int. J. Environ. Res. Public Health 2021, 18, 1758. https://doi.org/10.3390/ijerph18041758
Iftikhar M, Noureen A, Uzair M, Jabeen F, Abdel Daim M, Cappello T. Perspectives of Nanoparticles in Male Infertility: Evidence for Induced Abnormalities in Sperm Production. International Journal of Environmental Research and Public Health. 2021; 18(4):1758. https://doi.org/10.3390/ijerph18041758
Chicago/Turabian StyleIftikhar, Mehwish, Aasma Noureen, Muhammad Uzair, Farhat Jabeen, Mohamed Abdel Daim, and Tiziana Cappello. 2021. "Perspectives of Nanoparticles in Male Infertility: Evidence for Induced Abnormalities in Sperm Production" International Journal of Environmental Research and Public Health 18, no. 4: 1758. https://doi.org/10.3390/ijerph18041758
APA StyleIftikhar, M., Noureen, A., Uzair, M., Jabeen, F., Abdel Daim, M., & Cappello, T. (2021). Perspectives of Nanoparticles in Male Infertility: Evidence for Induced Abnormalities in Sperm Production. International Journal of Environmental Research and Public Health, 18(4), 1758. https://doi.org/10.3390/ijerph18041758