
Differential Expression of the Apolipoprotein AI Gene in Spotnape Ponyfish (Nuchequula nuchalis) Inhabiting Different Salinity Ranges at the Top of the Estuary and in the Deep-Bay Area of Gwangyang Bay, South Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
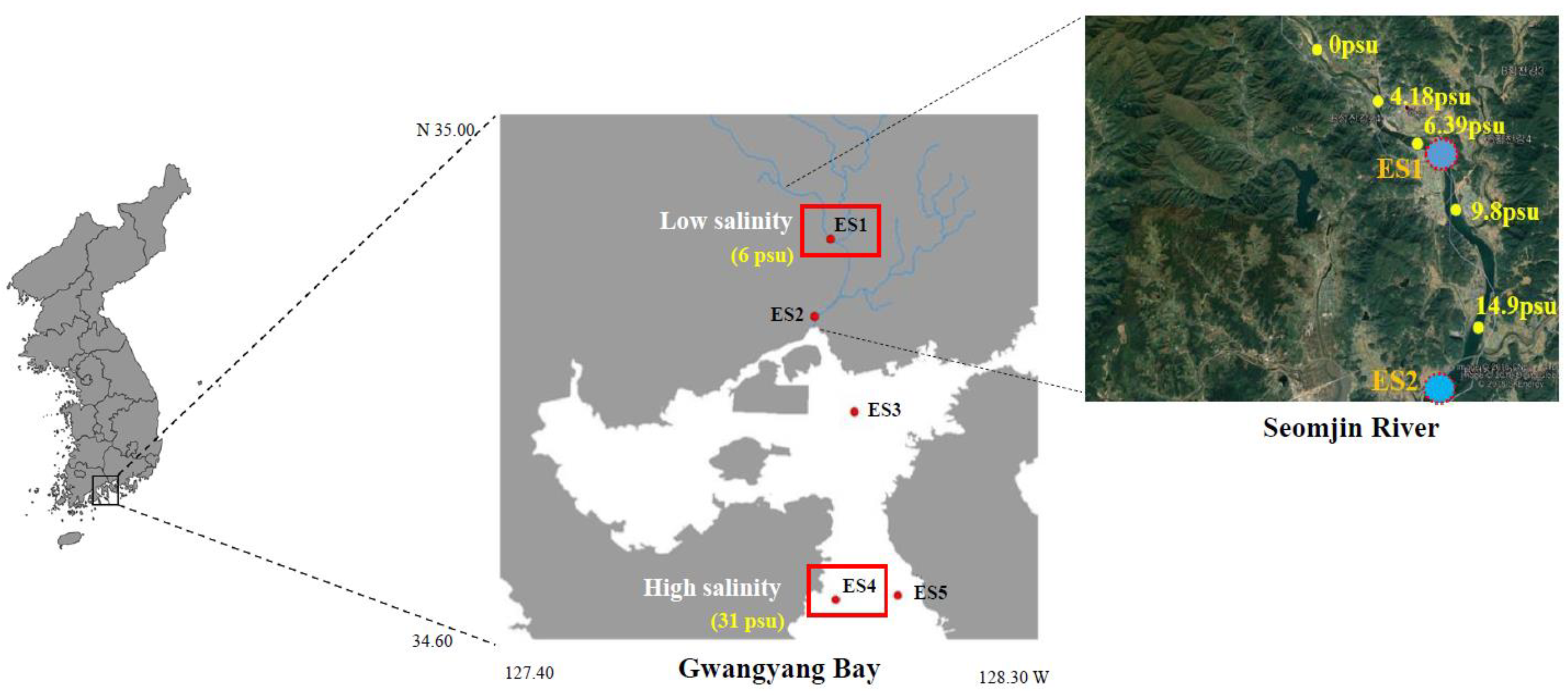
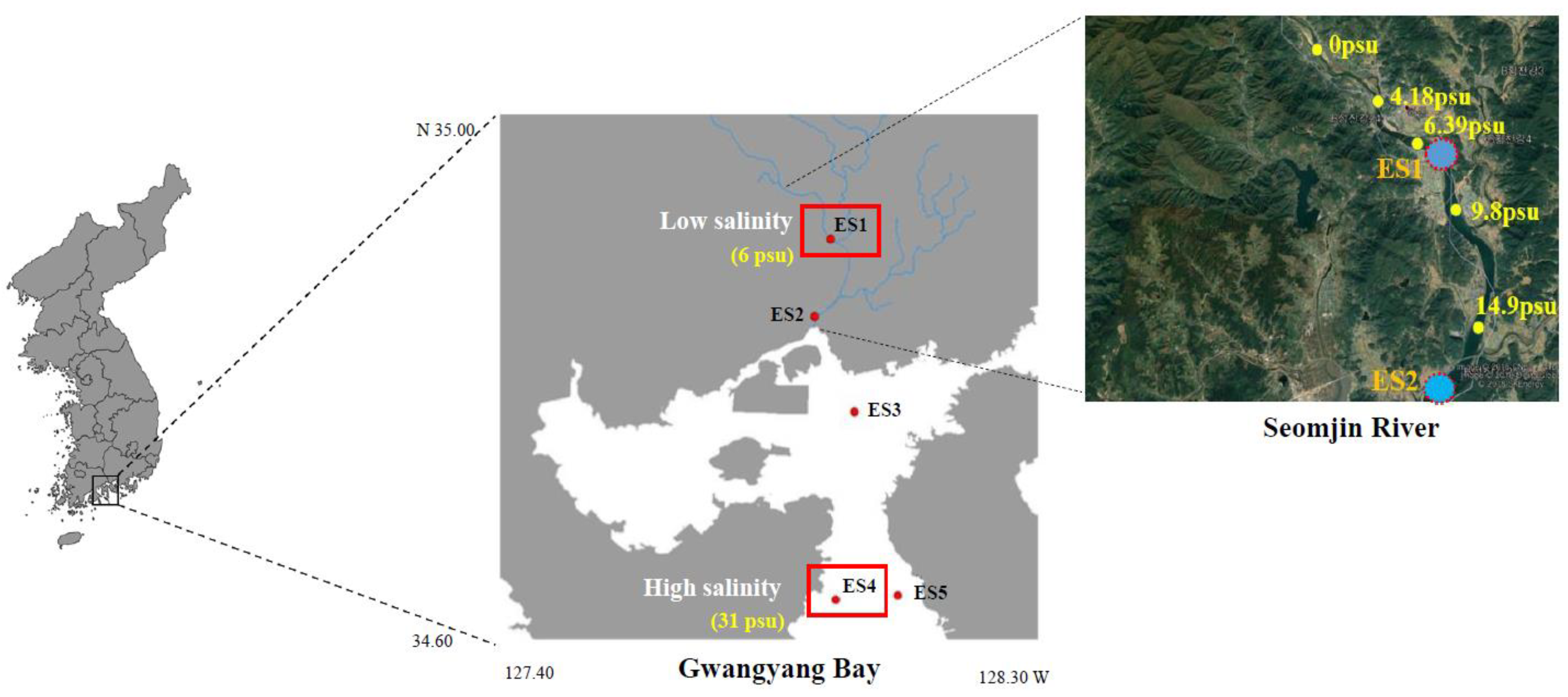
2.2. Sample Collection and Environmental Factor Measurements
2.3. Stable Isotope Analysis of the Bulk Fish Tissue
2.4. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
2.5. Data Analysis
3. Results
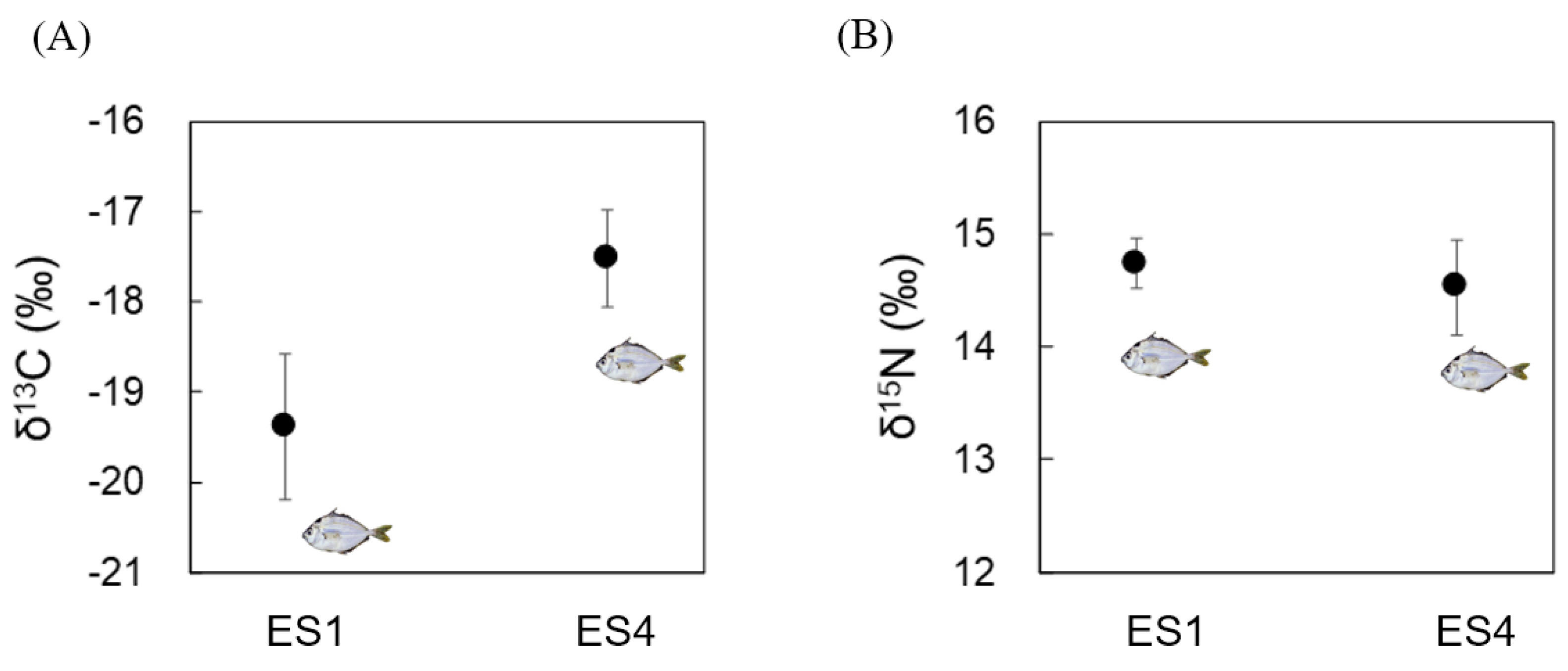
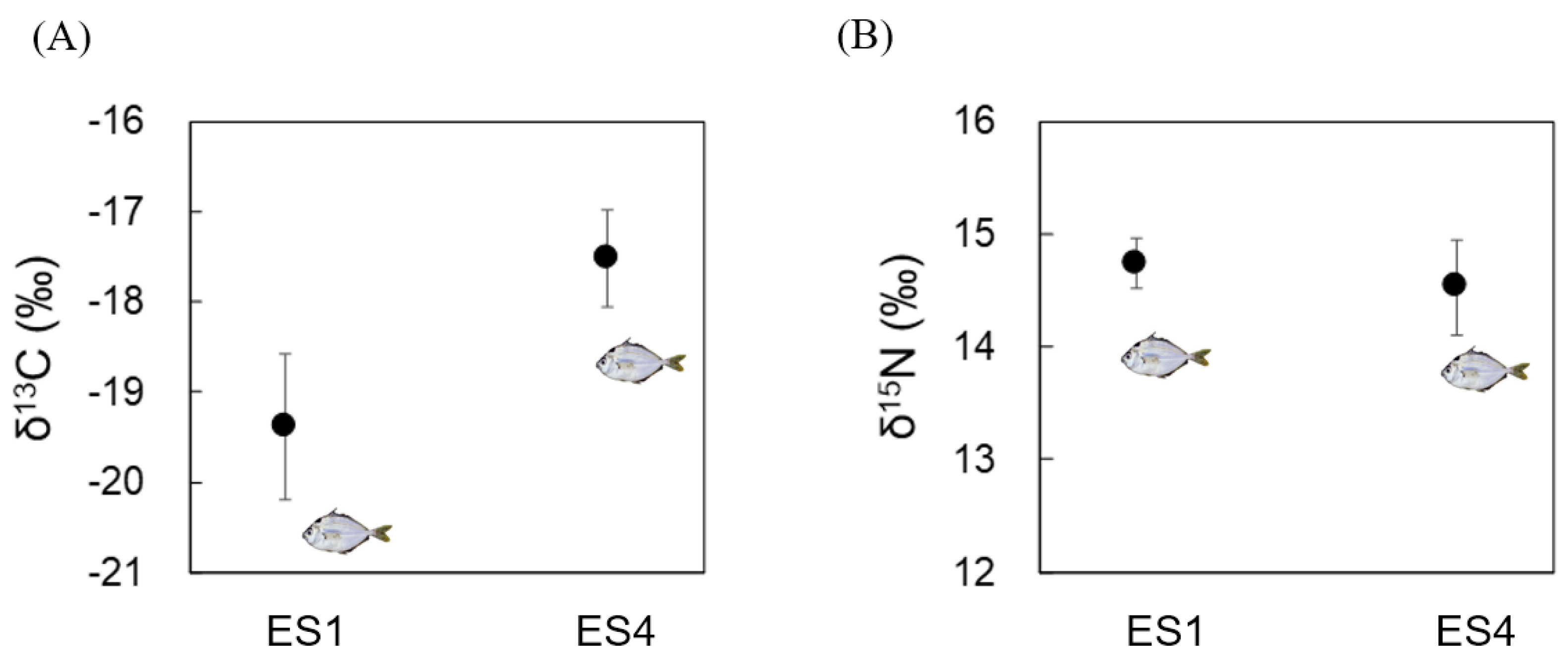
3.1. Analyses of δ13C and δ15N in Bulk Tissue from N. nuchalis
3.2. Characterization and Phylogenetic Analysis of the N. nuchalis ApoAI Gene
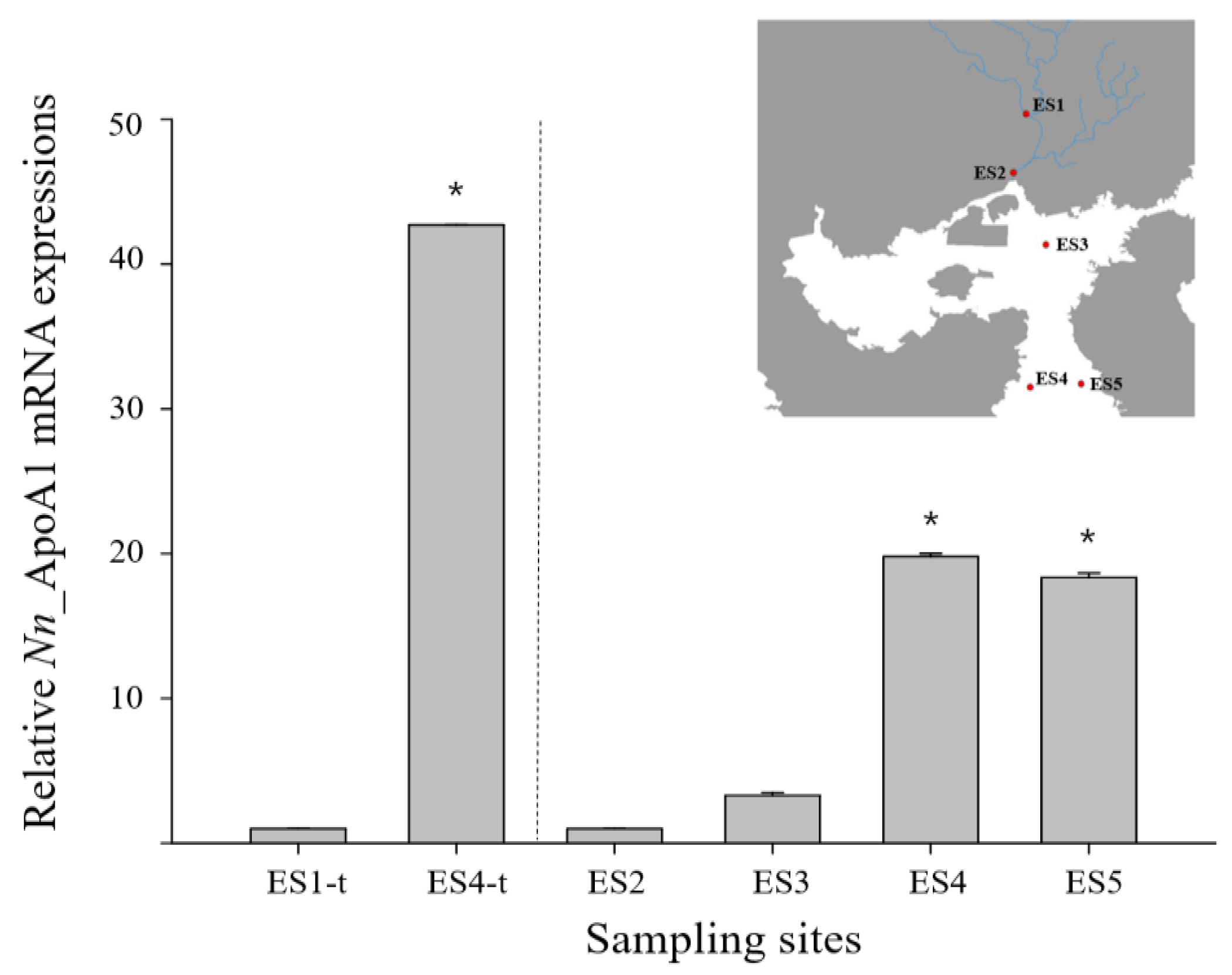
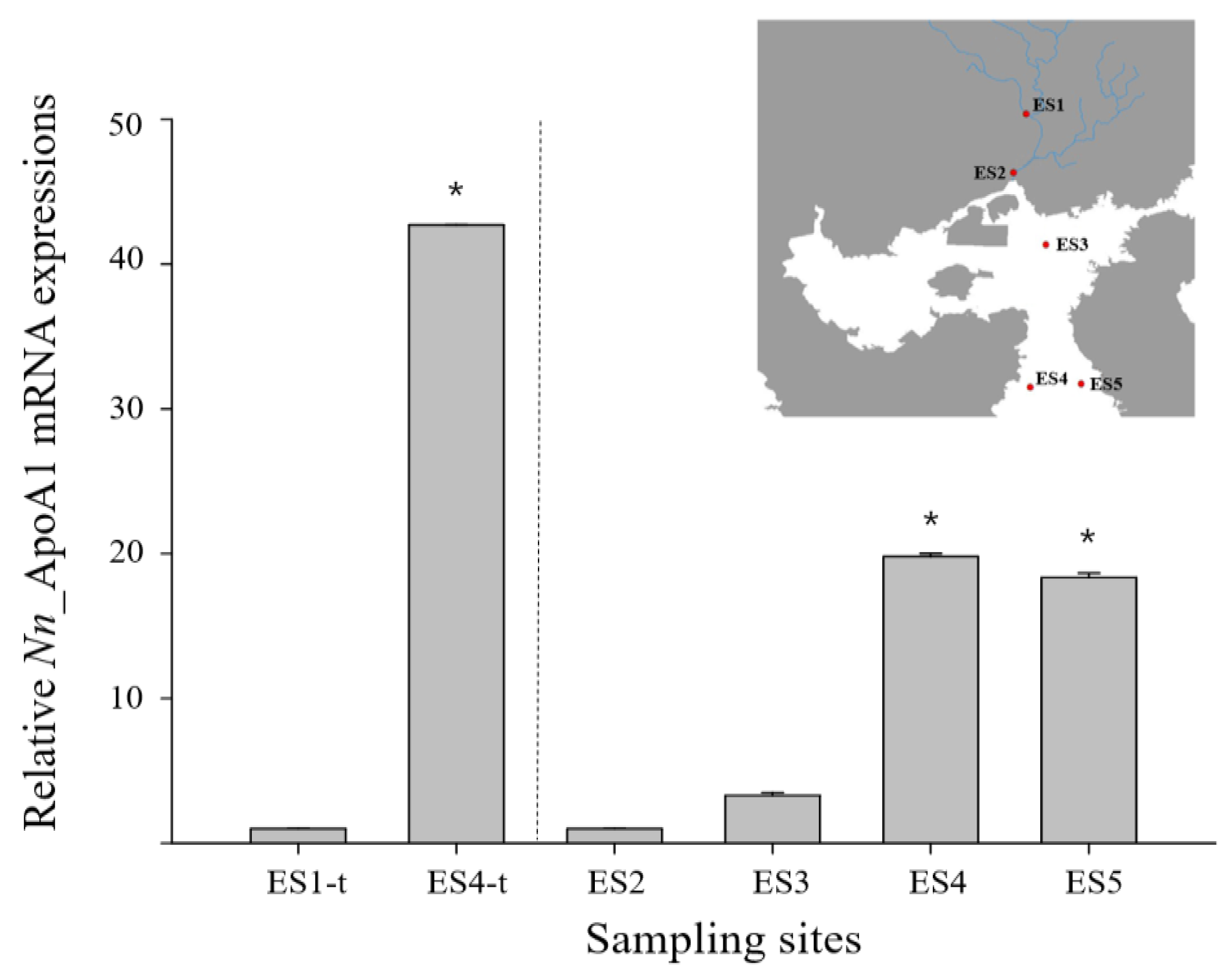
3.3. ApoAI Gene Expressions in N. nuchalis Collected from Different Salinity Environments
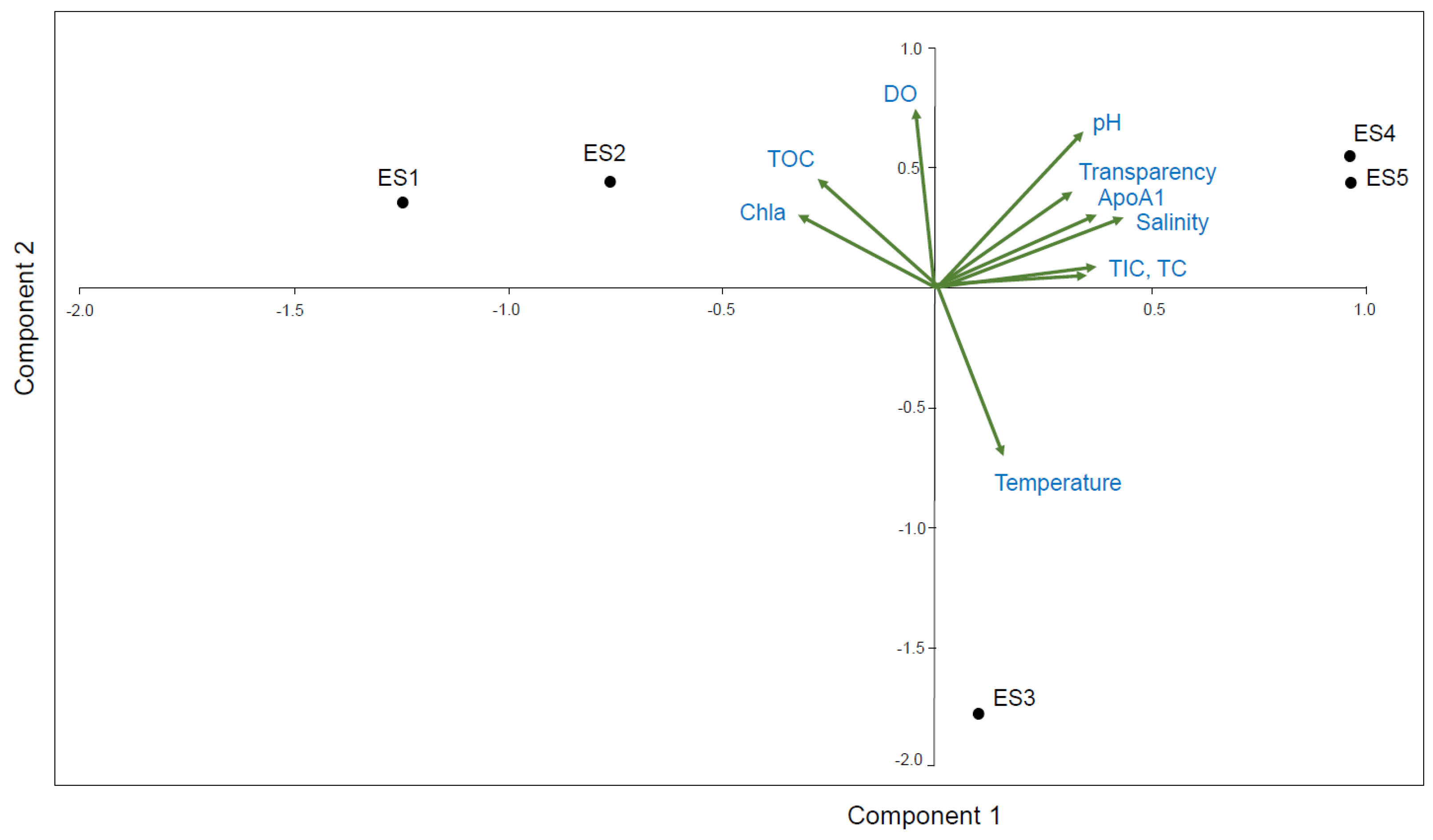
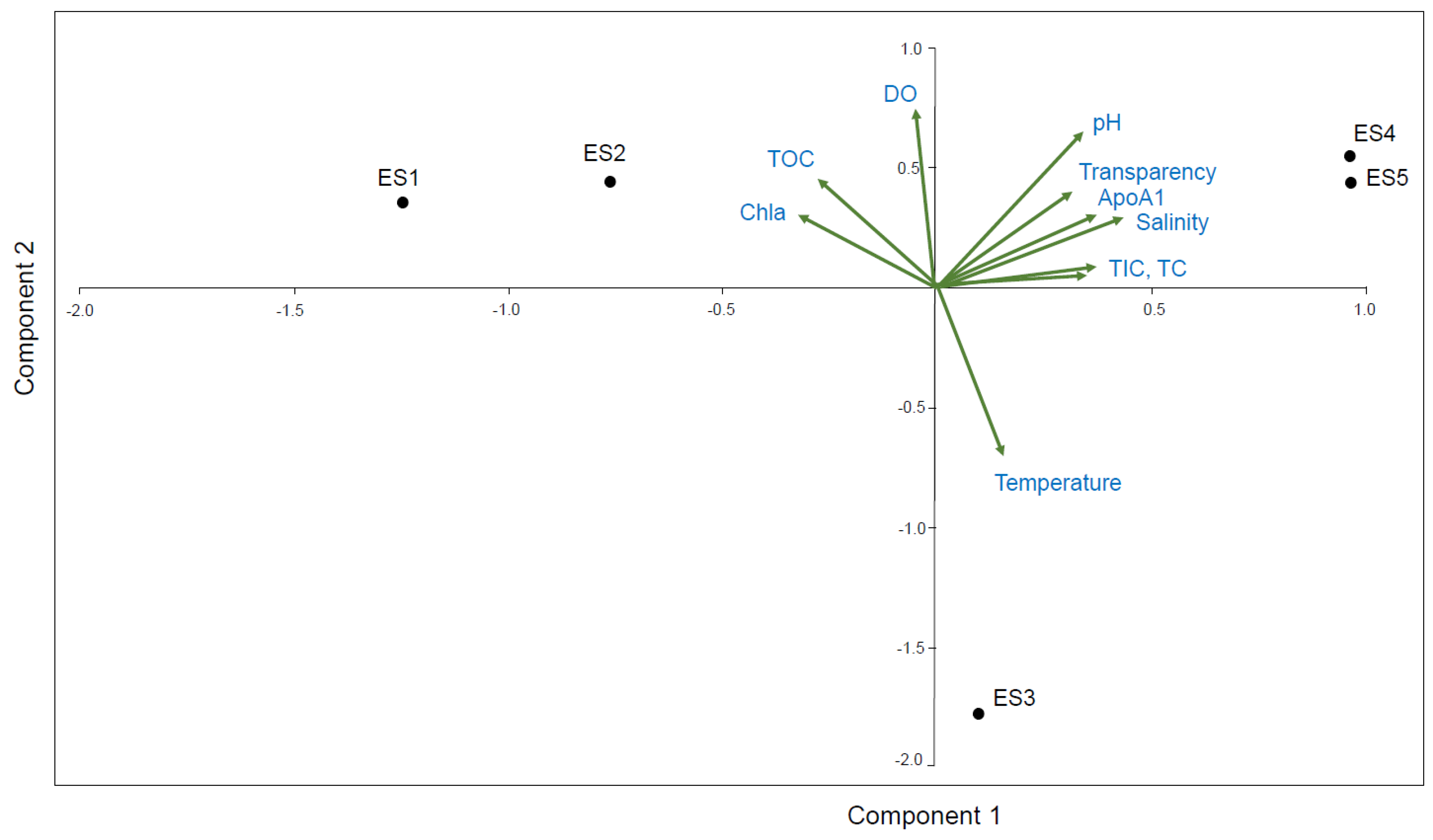
3.4. PCA Correlation between the ApoAI Transcript Expression and Nine Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Lee, Y.S.; Kang, C.K. Causes of COD increases in Gwangyang Bay, South Korea. J. Environ. Monit. 2010, 12, 1537–1546. [Google Scholar] [CrossRef]
- Kang, C.K.; Kim, J.B.; Lee, K.S.; Kim, J.B.; Lee, P.Y.; Hong, J.S. Trophic importance of benthic microalgae to macrozoobenthos in coastal bay systems in Korea: Dual stable C and N isotope analyses. Mar. Ecol. Prog. Ser. 2003, 259, 79–92. [Google Scholar] [CrossRef]
- Kim, D.K.; Park, K.; Jo, H.; Kwak, I.S. Comparison of water sampling between environmental DNA metabarcoding and conventional microscopic identification: A case study in Gwangyang bay, South Korea. Appl. Sci. 2019, 9, 3272. [Google Scholar] [CrossRef]
- Jo, H.; Kim, D.K.; Park, K.; Kwak, I.S. Discrimination of spatial distribution of aquatic organisms in a coastal ecosystem using eDNA. Appl. Sci. 2019, 9, 3450. [Google Scholar] [CrossRef]
- Kim, D.K.; Jo, H.; Han, I.; Kwak, I.S. Explicit characterization of spatial heterogeneity based on water quality, sediment contamination, and ichthyofauna in a riverine-to-coastal zone. Int. J. Environ. Res. Public Health 2019, 16, 409. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Tsutsui, S.; Tasumi, S.; Suetake, H.; Kikuchi, K.; Suzuki, Y. Tandem repeat L-rhamnose-binding lectin from the skin mucus of ponyfish, Leiognathus nuchalis. Biochem. Biophys. Res. Commun. 2005, 333, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Chyung, M.K. The Fishes of Korea; Iljisa: Seoul, Korea, 1977; 727p. [Google Scholar]
- Lee, J.S.; Huh, S.H. Reproductive biology of the slimy, Leiognathus nuchalis (Teleostei: Leiognathidae). Korean J. Ichthyol. 2000, 12, 192–202. [Google Scholar]
- Choi, B.; Jo, H.; Park, K.; Kwak, I.S. Isotopic evidence for ontogenetic shift in food resource utilization during the migration of the slipmouth Leiognathus nuchalis in Gwangyang bay, Korea. Korean J. Ichthyol. 2020, 32, 84–90. [Google Scholar] [CrossRef]
- Huh, S.H.; Kwak, S.N. Feeding habits of Leiognathus nuchalis in eelgrass (Zostera marina) bed in Kwangyang Bay. Korean J. Ichthyol. 1997, 9, 221–227. [Google Scholar]
- Shi, Z.; Huang, X.; Fu, R.; Wang, H.; Luo, H.; Bo, C. Salinity stress on embryos and early larval stages of the pomfret Pampus punctatissimus. Aquaculture 2008, 275, 306–310. [Google Scholar] [CrossRef]
- Cui, Q.; Qiu, L.; Yang, X.; Shang, S.; Yang, B.; Chen, M.; Liu, X.; Chen, B.; Fu, X.; Wang, W.; et al. Transcriptome profiling of the low-salinity stress responses in the gills of the juvenile Pseudopleuronectes yokohamae. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 32, 100612. [Google Scholar] [CrossRef]
- Torres, G.; Charmantier-Daures, M.; Chifflet, S.; Anger, K. Effects of long-term exposure to different salinities on the location and activity of Na+/K+-ATPase in the gills of juvenile mitten crab, Eriocheir sinensis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Wu, X.; Zhao, L.; Ye, H.; Cheng, Y.; Zeng, C. Effects of salinity on gonadal development, osmoregulation and metabolism of adult male Chinese mitten crab, Eriocheir sinensis. PLoS ONE 2017, 12, e0179036. [Google Scholar]
- Li, E.; Wang, S.; Li, C.; Wang, X.; Chen, K.; Chen, L. Transcriptome sequencing revealed the genes and pathways involved in salinity stress of Chinese mitten crab, Eriocheir sinensis. Physiol. Genom. 2014, 46, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhou, J.; Wei, B.; Cheng, Y.; Zhang, L.; Zhen, X. Comparative transcriptome analysis reveals osmotic-regulated genes in the gill of Chinese mitten crab (Eriocheir Sinensis). PLoS ONE 2019, 14, e0210469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, J.; Qi, T.; Ge, B.; Liu, Q.; Jiang, S.; Zhang, H.; Wang, Z.; Ding, G.; Tang, B. Comparative transcriptome analysis of Eriocheir japonica sinensis response to environmental salinity. PLoS ONE 2018, 13, e0203280. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, Y.H.; Hu, H.Q.; Niu, H.; Li, M.Y. Apolipoprotein A-I, a hyperosmotic adaptation-related protein in ayu (Plecoglossus altivelis). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 196–201. [Google Scholar]
- Mohapatra, A.; Karan, S.; Kar, B.; Garg, L.C.; Dixit, A.; Sahoo, P.K. Apolipoprotein A-I in Labeo rohita: Cloning and functional characterisation reveal its broad spectrum antimicrobial property, and indicate significant role during ectoparasitic infection. Fish Shellfish Immunol. 2016, 55, 717–728. [Google Scholar] [CrossRef]
- Andreeva, A.M.; Serebryakova, M.V.; Lamash, N.E. Oligomeric protein complexes of apolipoproteins stabilize the internal fluid environment of organism in redfins of the Tribolodon genus [Pisces; Cypriniformes, Cyprinidae]. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 90–97. [Google Scholar] [CrossRef]
- Tian, Y.; Wen, H.; Qi, X.; Mao, X.; Shi, Z.; Li, J.; He, F.; Yang, W.; Zhang, X.; Li, Y. Analysis of apolipoprotein multigene family in spotted sea bass (Lateolabrax maculatus) and their expression profiles in response to Vibrio harveyi infection. Fish Shellfish Immunol. 2019, 92, 111–118. [Google Scholar] [CrossRef]
- Andreeva, A.M.; Vasiliev, A.S.; Toropygin, I.Y.; Garina, D.V.; Lamash, N.; Filippova, A. Involvement of apolipoprotein A in maintaining tissue fluid balance in goldfish Carassius auratus. Fish Physiol. Biochem. 2019, 45, 1717–1730. [Google Scholar] [CrossRef] [PubMed]
- Pridgeon, J.W.; Klesius, P.H. Apolipoprotein A1 in channel catfish: Transcriptional analysis, antimicrobial activity, and efficacy as plasmid DNA immunostimulant against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2013, 35, 1129–1137. [Google Scholar] [CrossRef]
- Wei, J.; Gao, P.; Zhang, P.; Guo, M.; Xu, M.; Wei, S.; Yan, Y.; Qin, Q. Isolation and function analysis of apolipoprotein A-I gene response to virus infection in grouper. Fish Shellfish Immunol. 2015, 43, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of protein hydrolysates from anchovy (Engraulis encrasicolus) waste for twelve weeks decreases metabolic dysfunction-associated fatty liver disease severity in ApoE–/–mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef] [PubMed]
- Abbate, J.M.; Macrì, F.; Arfuso, F.; Iaria, C.; Capparucci, F.; Anfuso, C.; Ieni, A.; Cicero, L.; Briguglio, G.; Lanteri, G. Anti-atherogenic effect of 10% supplementation of anchovy (Engraulis encrasicolus) waste protein hydrolysates in ApoE-deficient mice. Nutrients 2021, 13, 2137. [Google Scholar] [CrossRef]
- Choi, B.; Ha, S.Y.; Lee, J.S.; Chikaraishi, Y.; Ohkouchi, N.; Shin, K.H. Trophic interaction among organisms in a seagrass meadow ecosystem as revealed by bulk δ13C and amino acid δ15N analyses. Limnol. Oceanogr. 2017, 62, 1426–1435. [Google Scholar] [CrossRef]
- Villarroel, F.; Bastías, A.; Casado, A.; Amthauer, R.; Concha, M.I. Apolipoprotein A-I, an antimicrobial protein in Oncorhynchus mykiss: Evaluation of its expression in primary defence barriers and plasma levels in sick and healthy fish. Fish Shellfish Immunol. 2007, 23, 197–209. [Google Scholar] [CrossRef]
- Sato, H.; Feix, J.B. Peptide–membrane interactions and mechanisms of membrane destruction by amphipathic α-helical antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1245–1256. [Google Scholar] [CrossRef]
- Watanabe, T. Lipid nutrition in fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1982, 73, 3–15. [Google Scholar] [CrossRef]
- Kondo, H.; Morinaga, K.; Misaki, R.; Nakaya, M.; Watabe, S. Characterization of the pufferfish Takifugu rubripes apolipoprotein multigene family. Gene 2005, 346, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Dodds, A.W.; Gudmundsdóttir, S.; Bambir, S.H.; Magnadóttir, B. The ontogenic transcription of complement component C3 and apolipoprotein A-I tRNA in Atlantic cod (Gadus morhua L.) —A role in development and homeostasis? Dev. Comp. Immunol. 2005, 29, 1065–1077. [Google Scholar] [CrossRef]
- Fang, X.; Wei, Y.; Liu, Y.; Wang, J.; Dai, J. The identification of apolipoprotein genes in rare minnow (Gobiocypris rarus) and their expression following perfluorooctanoic acid exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Román-Padilla, J.; Rodríguez-Rúa, A.; Manchado, M.; Hachero-Cruzado, I. Molecular characterization and developmental expression patterns of apolipoprotein A-I in Senegalese sole (Solea senegalensis Kaup). Gene Expr. Patterns 2016, 21, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, T.; Pellizzeri, V.; Benameur, Q.; Gervasi, C.; Santini, A.; Cicero, N.; Dugo, G. Valorization of raw materials from agricultural industry for astaxanthin and β-carotene production by Xanthophyllomyces dendrorhous. Nat. Prod. Res. 2018, 32, 1554–1561. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity (psu) | Temperature (°C) | Transparency (cm) | DO (mg L−1) | pH | TOC (mg L−1) | TIC (mg L−1) | TC (mg L−1) | Chl-a (μg L−1) | |
|---|---|---|---|---|---|---|---|---|---|
| ES1 | 6.39 | 20 | 135 | 7.99 | 7.6 | 3.617 | 7.546 | 11.163 | 4.93 |
| ES2 | 14.9 | 20.6 | 175 | 7.8 | 7.83 | 3.961 | 9.32 | 13.281 | 4.64 |
| ES3 | 27.3 | 22.7 | 200 | 6.68 | 7.43 | 2.562 | 10.612 | 13.174 | 1.3 |
| ES4 | 31.22 | 21.3 | 360 | 8.05 | 8.19 | 2.779 | 12.572 | 15.35 | 0.87 |
| ES5 | 31.23 | 20.7 | 350 | 7.66 | 8.14 | 2.932 | 14.084 | 17.015 | 2.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.; Kim, W.-S.; Choi, B.; Kwak, I.-S. Differential Expression of the Apolipoprotein AI Gene in Spotnape Ponyfish (Nuchequula nuchalis) Inhabiting Different Salinity Ranges at the Top of the Estuary and in the Deep-Bay Area of Gwangyang Bay, South Korea. Int. J. Environ. Res. Public Health 2021, 18, 10960. https://doi.org/10.3390/ijerph182010960
Park K, Kim W-S, Choi B, Kwak I-S. Differential Expression of the Apolipoprotein AI Gene in Spotnape Ponyfish (Nuchequula nuchalis) Inhabiting Different Salinity Ranges at the Top of the Estuary and in the Deep-Bay Area of Gwangyang Bay, South Korea. International Journal of Environmental Research and Public Health. 2021; 18(20):10960. https://doi.org/10.3390/ijerph182010960
Chicago/Turabian StylePark, Kiyun, Won-Seok Kim, Bohyung Choi, and Ihn-Sil Kwak. 2021. "Differential Expression of the Apolipoprotein AI Gene in Spotnape Ponyfish (Nuchequula nuchalis) Inhabiting Different Salinity Ranges at the Top of the Estuary and in the Deep-Bay Area of Gwangyang Bay, South Korea" International Journal of Environmental Research and Public Health 18, no. 20: 10960. https://doi.org/10.3390/ijerph182010960
APA StylePark, K., Kim, W.-S., Choi, B., & Kwak, I.-S. (2021). Differential Expression of the Apolipoprotein AI Gene in Spotnape Ponyfish (Nuchequula nuchalis) Inhabiting Different Salinity Ranges at the Top of the Estuary and in the Deep-Bay Area of Gwangyang Bay, South Korea. International Journal of Environmental Research and Public Health, 18(20), 10960. https://doi.org/10.3390/ijerph182010960