
Mercury Levels in Feathers of Penguins from the Antarctic Peninsula Area: Geographical and Inter-Specific Differences

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Material and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mason, R.P.; Choi, A.L.; Fitzgerald, W.F.; Hammerschmidt, C.R.; Lamborg, C.H.; Soerensen, A.L.; Sunderland, E.M. Mercury biogeochemical cycling in the ocean and policy implications. Environ. Res. 2012, 119, 101–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamborg, C.H.; Hammerschmidt, C.R.; Bowman, K.L.; Swarr, G.J.; Munson, K.M.; Ohnemus, D.C.; Lam, P.J.; Heimbürger, L.E.; Rijkenberg, M.J.A.; Saito, M.A. A global ocean inventory of anthropogenic mercury based on water column measurements. Nature 2014, 512, 65–68. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a global pollutant: Sources, pathways, and effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef]
- Evers, D. The Effects of Methylmercury on Wildlife: A Comprehensive Review and Approach for Interpretation. Encycl. Anthr. 2018, 5, 181–194. [Google Scholar] [CrossRef]
- Weiner, J.G.; Krabbemhoft, D.P.; Heinz, G.H.; Scheuhammer, A.M. Ecotoxicology of mercury. In Handbook of Ecotoxicology, 2nd ed.; Hoffman, D.J., Rattner, B.A., Burton, G.A., Jr., Cairns, J., Jr., Eds.; CRC Press: New York, NY, USA, 2003; pp. 409–463. [Google Scholar]
- Jackson, T.A. Long-range atmospheric transport of mercury to ecosystems, and the importance of anthropogenic emissions—A critical review and evaluation of the published evidence. Environ. Rev. 1997, 5, 99–120. [Google Scholar] [CrossRef]
- Smichowski, P.; Vodopivez, C.; Muñoz-Olivas, R.; Gutierrez, A.M. Monitoring trace elements in selected organs of Antarctic penguin (Pygoscelis adeliae) by plasma-based techniques. Microchem. J. 2006, 82, 1–7. [Google Scholar] [CrossRef]
- Spalding, M.G.; Frederick, P.; McGill, H.C.; Bouton, S.; McDowell, L.R. Methylmercury accumulation in tissues and its effects on growth and appetite in captive great egrets. J. Wildl. Dis. 2000, 36, 411–422. [Google Scholar] [CrossRef]
- UNEP, United Nations Environmental Programme. Global Mercury Assessment; UNEP Chemicals: Geneva, Switzerland, 2002. [Google Scholar]
- Bargagli, R.; Agnorelli, C.; Borghini, F.; Monaci, F. Enhanced deposition and bioaccumulation of mercury in Antarctic terrestrial ecosystems facing a coastal polynya. Environ. Sci. Technol. 2005, 39, 8150–8155. [Google Scholar] [CrossRef] [PubMed]
- Bargagli, R.; Monaci, F.; Bucci, C. Environmental biogeochemistry of mercury in Antarctic ecosystems. Soil. Biol. Biochem. 2007, 39, 352–360. [Google Scholar] [CrossRef]
- Metcheva, R.; Dimitrova Yurukova, L.; Teodorova, S.E.; Nikolova, E. The penguin feathers as bioindicator of Antarctica environmental state. Sci. Total Env. 2006, 362, 259–265. [Google Scholar] [CrossRef]
- Jerez, S.; Motas, M.; Palacios, M.J.; Valera, F.; Cuervo, J.J.; Barbosa, A. Concentration of trace elements in feathers of three Antarctic penguins: Geographical and interspecific differences. Env. Pollut. 2011, 159, 2412–2419. [Google Scholar] [CrossRef]
- Esteban, M.; Castaño, A. Non-invasive matrices in human biomonitoring: A review. Env. Int. 2009, 35, 438–449. [Google Scholar] [CrossRef]
- Dauwe, T.; Bervoets, L.; Blust, R.; Pinxten, R.; Eens, M. Can excrement and feathers of nestling songbirds be used as biomonitors for heavy metal pollution. Arch Env. Contam. Toxicol. 2000, 39, 541–546. [Google Scholar] [CrossRef]
- Adout, A.; Hawlena, D.; Maman, R.; Paz-Tal, O.; Karpas, Z. Determination of trace elements in pigeon and raven feathers by ICP-MS. Int. J. Mass. Spectrom. 2007, 267, 109–116. [Google Scholar] [CrossRef]
- Altmeyer, M.; Dittmen, J.; Dmowski, K.; Wagner, G.; Müller, P. Distribution of elements in flight feathers of a white-tailed eagle. Sci. Total Env. 1991, 105, 157–164. [Google Scholar] [CrossRef]
- Palacios, M.J.; Valera, F.; Colominas-Ciuró, R.; Barbosa, A. Cellular and humoral immunity in two highly demanding energetic life stages: Reproduction and moulting in the Chinstrap Penguin. J. Ornithol. 2018, 159, 283–290. [Google Scholar] [CrossRef]
- Brasso, R.L.; Drummond, B.E.; Borret, S.R.; Chiaradia, A.; Polito, M.J.; Raya Rey, A. Unique pattern of molt leads to low intraindividual variation in feather mercury concentrations in penguins. Env. Toxicol. Chem. 2013, 32, 2331–2334. [Google Scholar] [CrossRef]
- Bond, A.L.; Diamond, A.W. Total and methyl mercury concentrations in seabird feathers and eggs. Arch Env. Contam. Toxicol. 2009, 56, 286–291. [Google Scholar] [CrossRef]
- Pertierra, L.R.; Hughes, K.A.; Vega, G.C.; Olalla-Tárraga, M.Á. High Resolution Spatial Mapping of Human Footprint across Antarctica and Its Implications for the Strategic Conservation of Avifauna. PLoS ONE 2017, 12, e0168280. [Google Scholar] [CrossRef] [Green Version]
- Bargagli, R.; Monaci, F.; Sanchez-Hernandez, J.; Cateni, D. Biomagnification of mercury in an Antarctic marine coastal food web. Mar. Ecol. Prog. Ser. 1998, 169, 65–76. [Google Scholar] [CrossRef]
- Dimitrijević, D.; Paiva, V.H.; Ramos, J.A.; Seco, J.; Ceia, F.; Chipev, N.; Valente, T.; Barbosa, A.; Xavier, J. Isotopic niches of sympatric Gentoo and Chinstrap Penguins: Evidence of competition for Antarctic krill? Polar Biol. 2018, 41, 1655–1669. [Google Scholar] [CrossRef]
- Colominas-Ciuró, R.; Bertellotti, M.; D’Amico, V.L.; Carabajal, E.; Benzal, J.; Vidal, V.; Motas, M.; Santos, M.; Coria, N.; Barbosa, A. Diet, antioxidants and oxidative status in pygoscelid penguins. Mar. Ecol. Prog. Ser. 2021, 665, 201–216. [Google Scholar] [CrossRef]
- Smith, P.N.; Cobb, G.P.; Godard-Codding, C.; Hoff, D.; McMurry, S.T.; Rainwater, T.R.; Reynolds, K.D. Contaminant exposure in terrestrial vertebrates. Environ. Pollut. 2007, 150, 41–64. [Google Scholar] [CrossRef]
- Brasso, R.L.; Polito, M.J.; Lynch, H.J.; Naveen, R.; Emslie, S.D. Penguin eggshell membranes reflect homogeneity of mercury in the marine food web surrounding the Antarctic Peninsula. Sci. Total Env. 2012, 439, 165–171. [Google Scholar] [CrossRef]
- Masello, J.F.; Barbosa, A.; Kato, A.; Mattern, T.; Medeiros, R.; Stockdale, J.E.; Kümmel, M.N.; Bustamante, P.; Belliure, J.; Benzal, J.; et al. How animals distribute themselves in space: Energy landscapes of Antarctic avian predators. Mov. Ecol. 2021, 9, 24. [Google Scholar] [CrossRef]
- Brooks, S.; Lindberg, S.; Southworth, G.; Arimoto, R. Springtime atmospheric mercury speciation in the McMurdo, Antarctica coastal region. Atmos. Env. 2008, 42, 2885–2893. [Google Scholar] [CrossRef]
- Sheppard, D.S.; Patterson, J.E.; McAdam, M.K. Mercury content of Antarctic ice and snow: Further results. Atmos. Env. 1991, 25, 1657–1660. [Google Scholar] [CrossRef]
- Calle, P.; Alvarado, O.; Monserrate, L.; Cevallos, J.M.; Calle, N.; Alava, J.J. Mercury accumulation in sediments and seabird feathers from the Antarctic Peninsula. Mar. Pollut. Bull. 2015, 91, 410–417. [Google Scholar] [CrossRef]
- Bargagli, R. Antarctic Ecosystems: Environmental Contamination, Climate Change, and Human Impact; Springer: Berlin, Germany, 2005. [Google Scholar]
- dos Santos, I.R.; Silva-Filho, E.V.; Schaefer, C.; Sella, S.M.; Silva, C.A.; Gomes, V.; Passos, M.J.A.C.R.; Ngan, P.V. Baseline mercury and zinc concentrations in terrestrial and coastal organisms of Admiralty Bay, Antarctica. Env. Pollut. 2006, 140, 304–311. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M.; Jeitner, C.; Burke, S.; Volz, C.D.; Snigaroff, R.; Snigaroff, D.; Shukla, S. Mercury and other metals in eggs and feathers of glaucous-winged gulls (Larus glaucescens) in the Aleutians. Env. Monit. Assess. 2009, 152, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polito, M.J.; Brasso, R.L.; Trivelpiece, W.Z.; Karnovsky, N.; Patterson, W.P.; Emslie, S.D. Differing foraging strategies influence mercury (Hg) exposure in an Antarctic penguin community. Environ. Pollut. 2016, 218, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Frias, J.E.; Gil, M.N.; Esteves, J.L.; Garcia Borboroglu, P.; Kane, O.J.; Smith, J.R.; Boersma, P.D. Mercury levels in feathers of Magellanic penguins. Mar. Pollut. Bull. 2012, 64, 1265–1269. [Google Scholar] [CrossRef]
- Goutner, V.; Becker, P.H.; Liordos, V.; Tsachalidis, E.P. Mercury in white stork (Ciconia ciconia) chick feathers from Northeastern Mediterranean areas in relation to age, brood size, and hatching order. Arch. Environ. Contam. Toxicol. 2011, 61, 327–336. [Google Scholar] [CrossRef]
- Kojadinovic, J.; Bustamante, P.; Churlaud, C.; Cosson, R.P.; Le Corre, M. Mercury in seabird feathers: Insight on dietary habits and evidence for exposure levels in the western Indian Ocean. Sci. Total Environ. 2007, 384, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Ebinghaus, R.; Kock, H.H.; Temme, C.; Einax, J.W.; Löwe, A.G.; Richter, A.; Burrows, J.P.; Schroeder, W.H. Antarctic springtime depletion of atmospheric mercury. Environ. Sci. Technol. 2002, 36, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Pfaffhuber, K.A.; Berg, T.; Hirdma, D.; Stohl, A. Atmospheric mercury observations from Antarctica: Seasonal variation and source and sink region calculations. Atmos. Chem. Phys. 2011, 11, 29139–29177. [Google Scholar] [CrossRef] [Green Version]
- Stern, G.A.; Macdonald, R.W.; Outridge, P.M.; Wilson, S.; Chetelat, J.; Cole, A.; Hintelmann, H.; Loseto, L.L.; Steffen, A.; Wang, F.; et al. How does climate change influence arctic mercury? Sci. Total Environ. 2012, 414, 22–42. [Google Scholar] [CrossRef] [PubMed]
- Krabbenhoft, D.P.; Sunderland, E.M. Global change and mercury. Science 2013, 341, 1457–1458. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, B.D.; Driscoll, C.T.; Maxwell, J.A.; Holsen, T.M. Changing climate alters inputs and pathways of mercury deposition to forested ecosystems. Biogeochemistry 2014, 119, 215–228. [Google Scholar] [CrossRef] [Green Version]
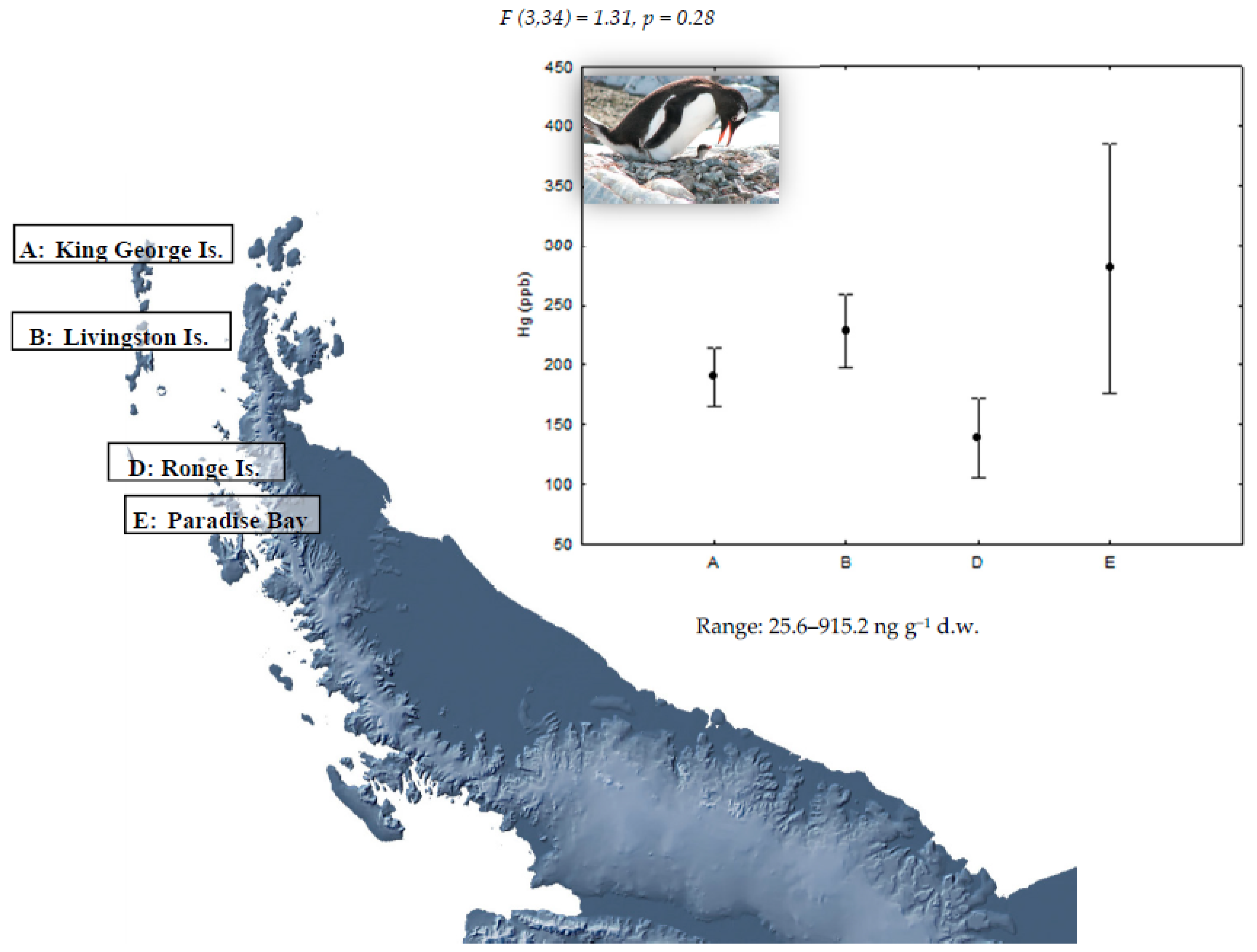
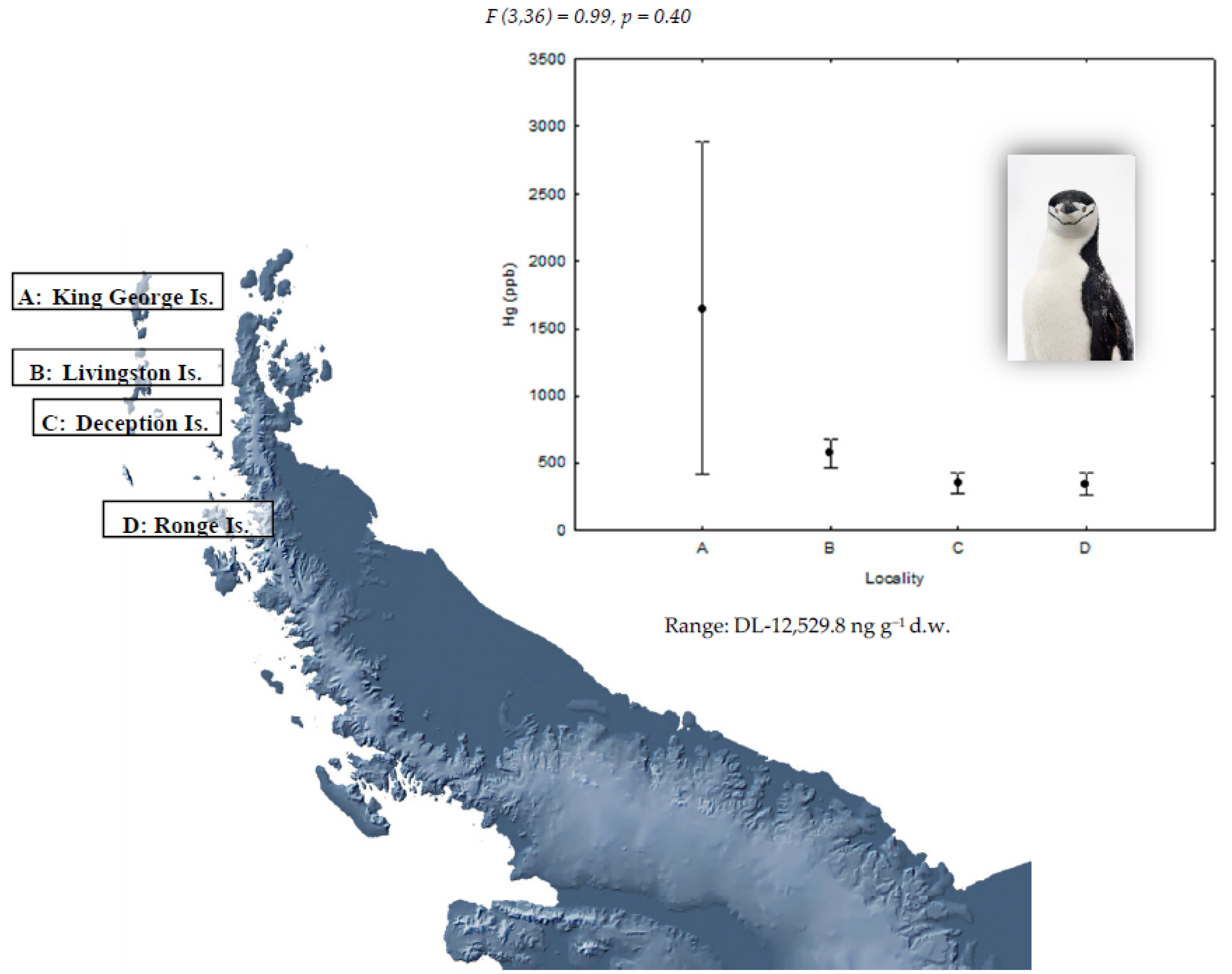
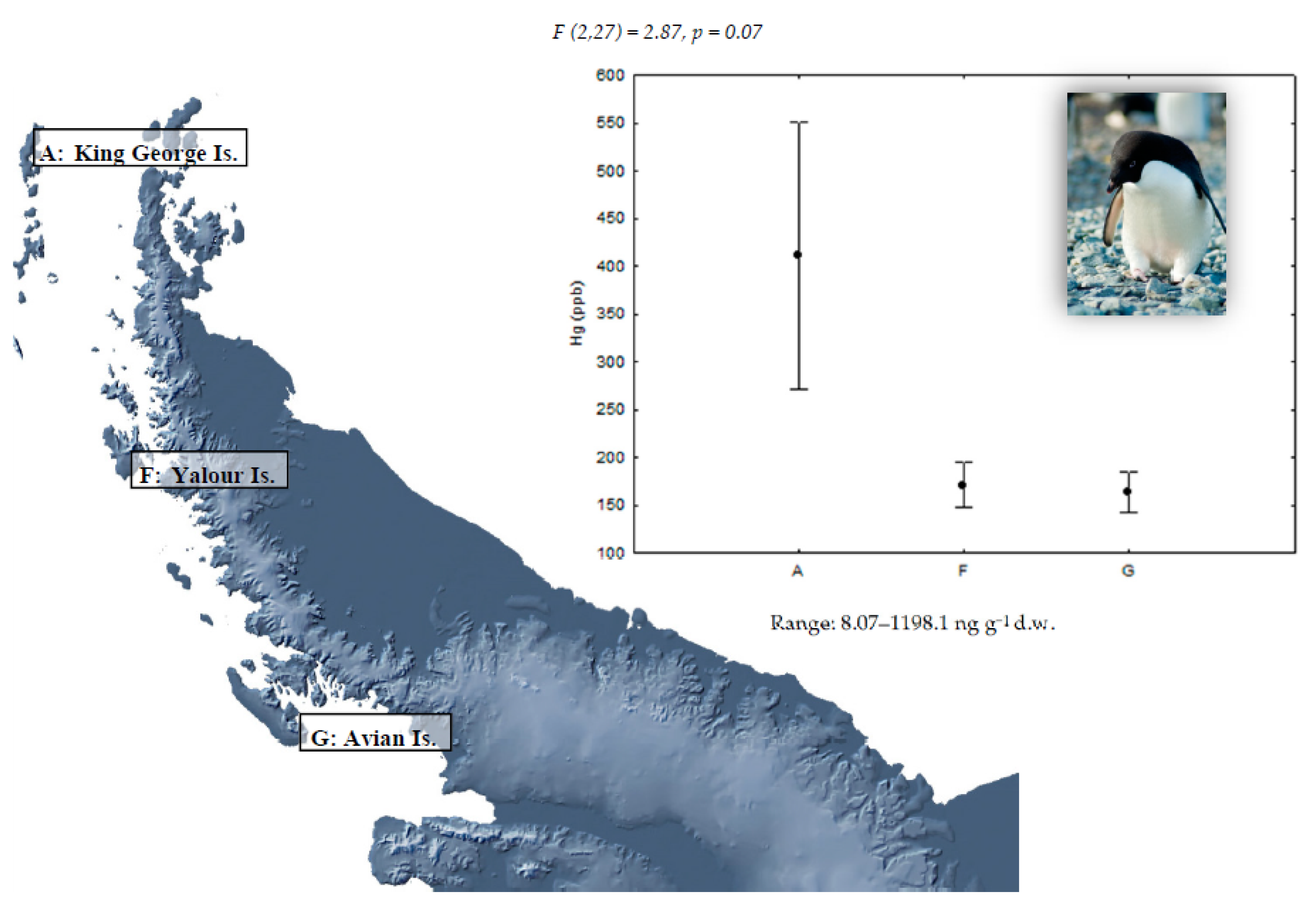
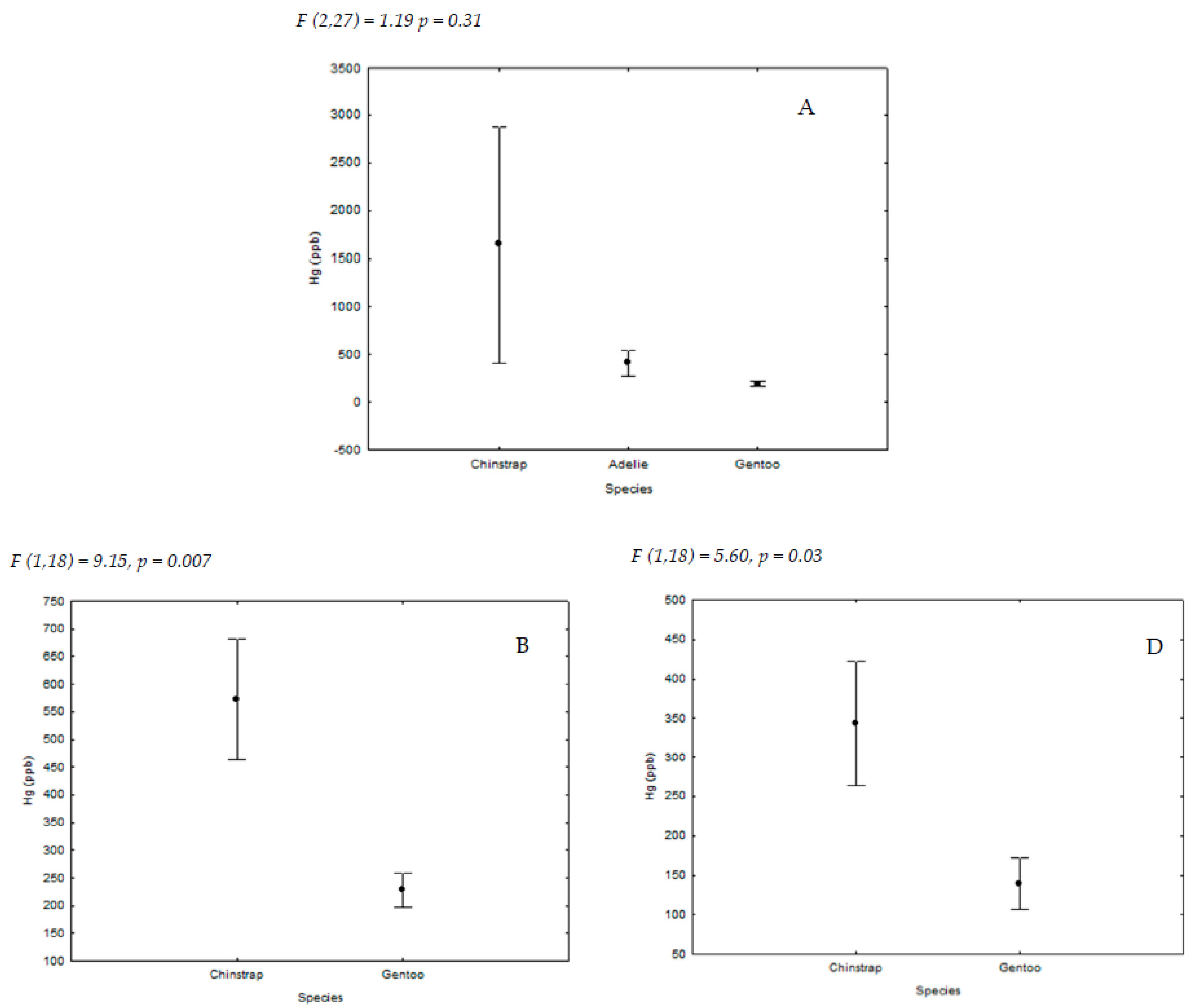

{kind=link}
{kind=link}
{kind=link}
{kind=link}
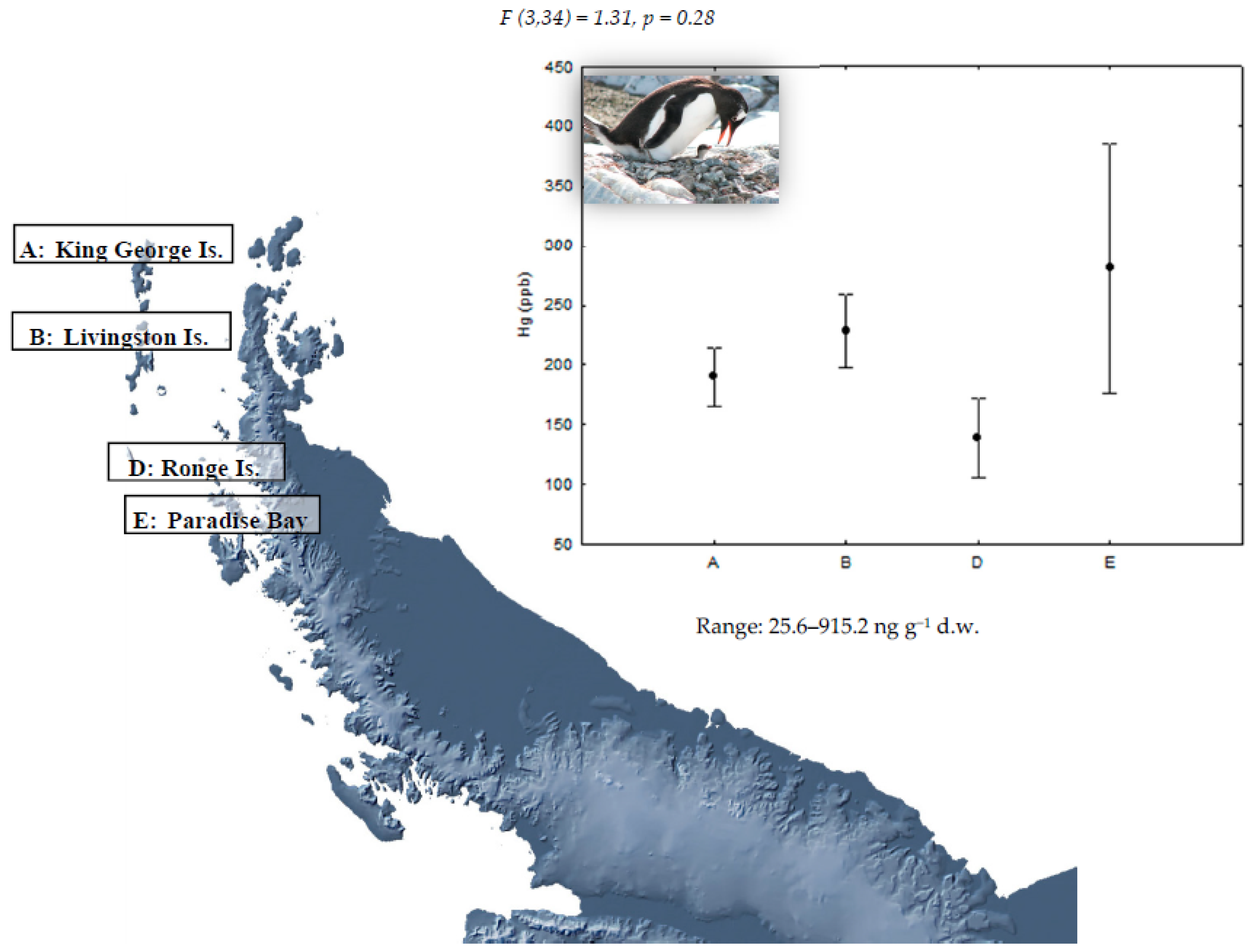
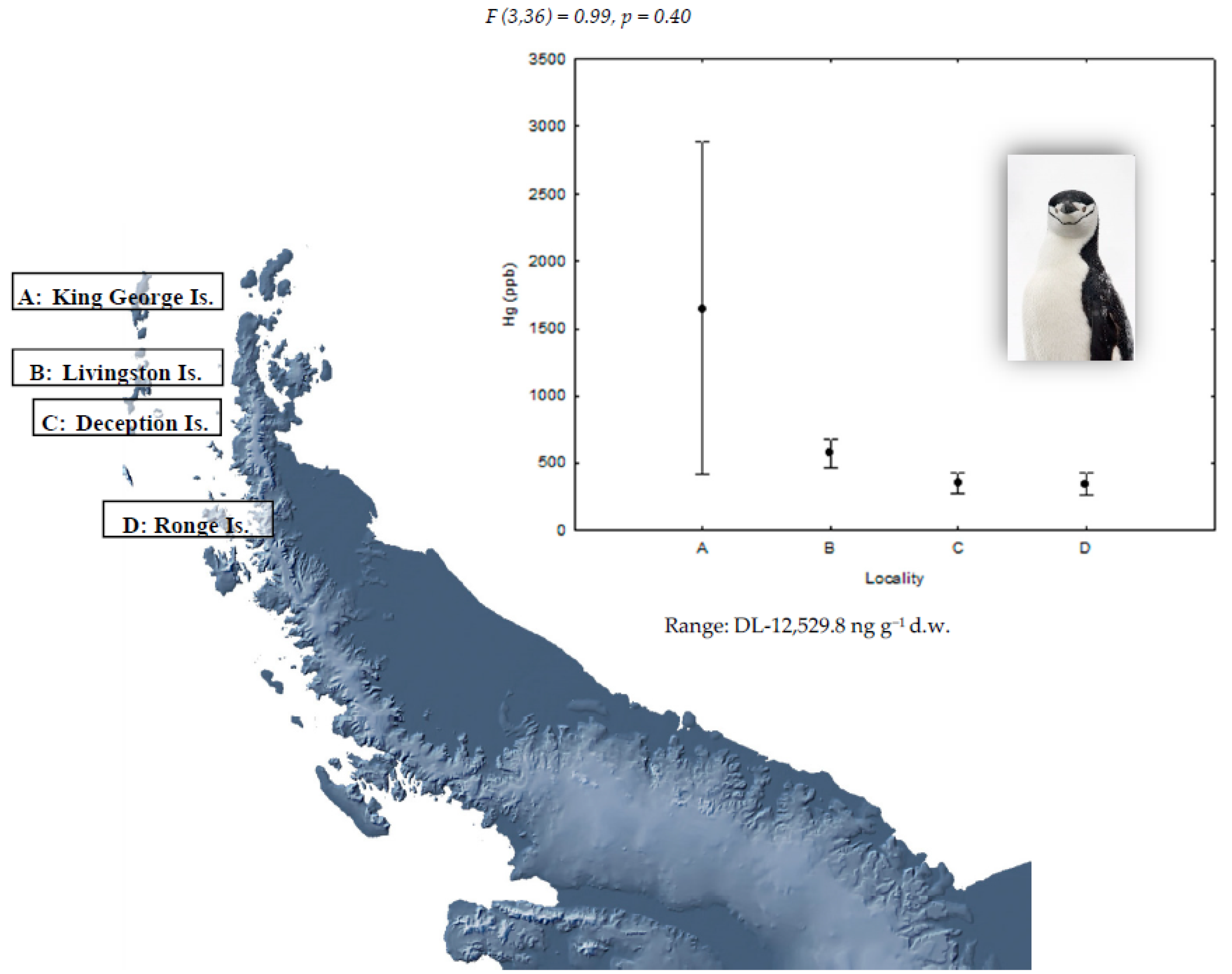
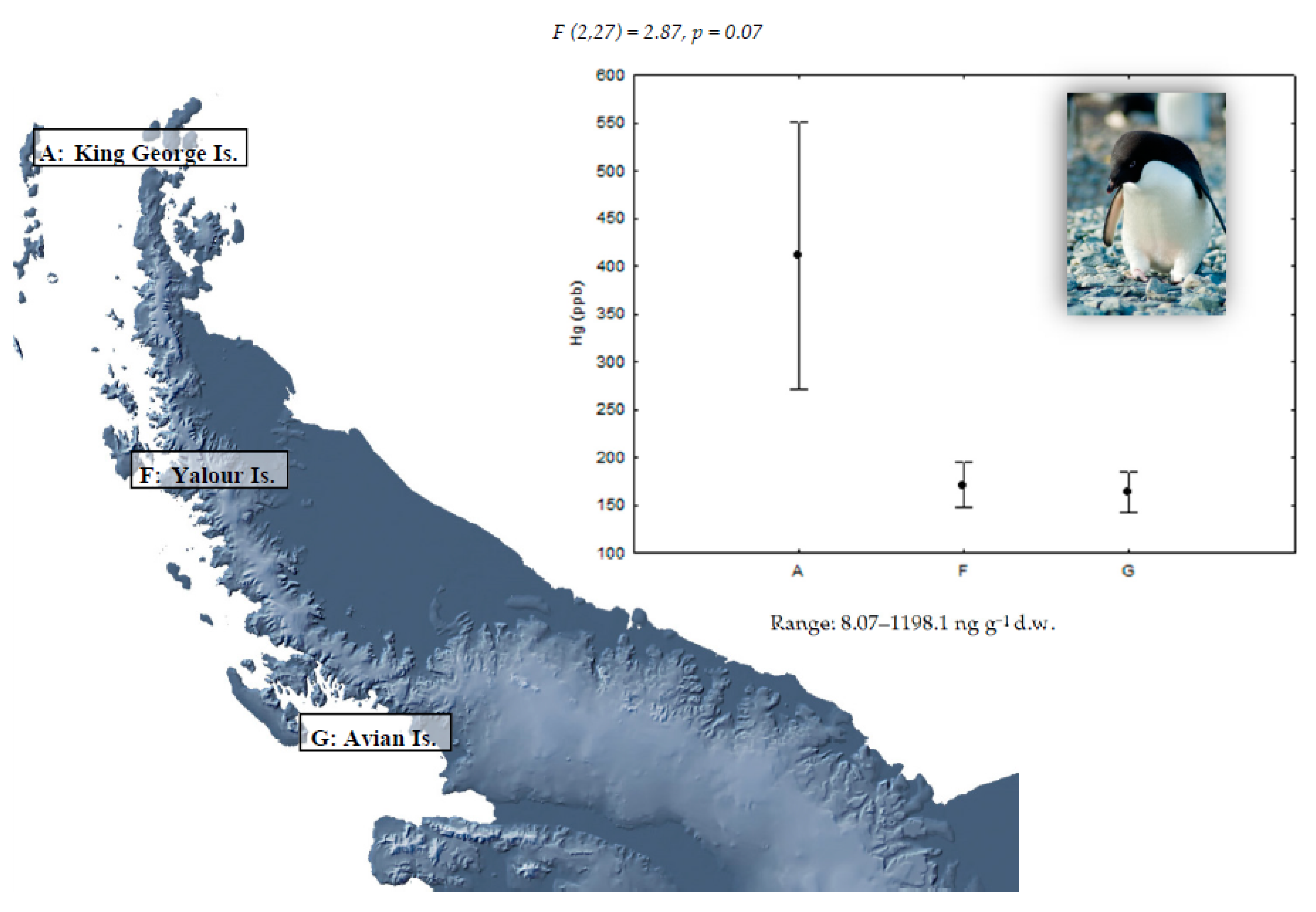
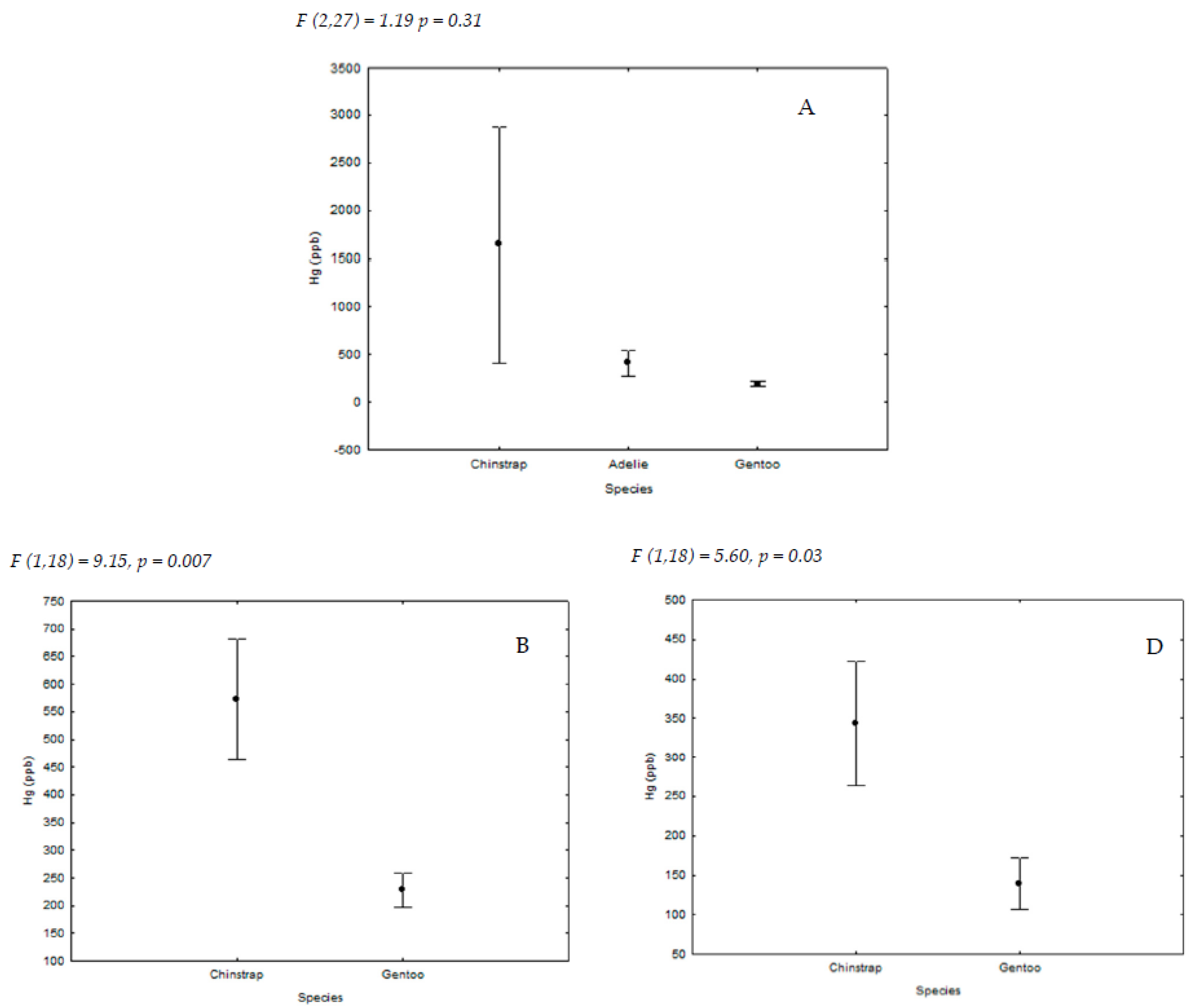
| Locations | Species | ||
|---|---|---|---|
| P. papua | P. antarctica | P. adeliae | |
| King George Island (62°15′ S, 58°37′ W) A | 10 | 10 | 10 |
| Livingston Island (62°39′ S, 60°36′ W) B | 10 | 10 | - |
| Deception Island (63°00′ S, 60°40′ W) C | - | 10 | - |
| Ronge Island (64°40′ S, 62°40′ W) D | 10 | 10 | - |
| Paradise Bay (64°53′ S, 62°53′ W) E | 8 | - | - |
| Yalour Island (65°15′ S, 64°11′ W) F | - | - | 10 |
| Avian Island (67°46′ S, 68°64′ W) G | - | - | 10 |
| Element | Detection Limit | DORM-2 | % Recovery | DOLT-2 | % Recovery |
|---|---|---|---|---|---|
| Hg202 | 0.18 | 4.64 ± 0.26 | 89.2 | 2.14 ± 0.28 | 91.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motas, M.; Jerez, S.; Esteban, M.; Valera, F.; Cuervo, J.J.; Barbosa, A. Mercury Levels in Feathers of Penguins from the Antarctic Peninsula Area: Geographical and Inter-Specific Differences. Int. J. Environ. Res. Public Health 2021, 18, 9918. https://doi.org/10.3390/ijerph18189918
Motas M, Jerez S, Esteban M, Valera F, Cuervo JJ, Barbosa A. Mercury Levels in Feathers of Penguins from the Antarctic Peninsula Area: Geographical and Inter-Specific Differences. International Journal of Environmental Research and Public Health. 2021; 18(18):9918. https://doi.org/10.3390/ijerph18189918
Chicago/Turabian StyleMotas, Miguel, Silvia Jerez, Marta Esteban, Francisco Valera, José Javier Cuervo, and Andrés Barbosa. 2021. "Mercury Levels in Feathers of Penguins from the Antarctic Peninsula Area: Geographical and Inter-Specific Differences" International Journal of Environmental Research and Public Health 18, no. 18: 9918. https://doi.org/10.3390/ijerph18189918
APA StyleMotas, M., Jerez, S., Esteban, M., Valera, F., Cuervo, J. J., & Barbosa, A. (2021). Mercury Levels in Feathers of Penguins from the Antarctic Peninsula Area: Geographical and Inter-Specific Differences. International Journal of Environmental Research and Public Health, 18(18), 9918. https://doi.org/10.3390/ijerph18189918