
Screening of Eurasian Tundra Reindeer for Viral Sequences by Next-Generation Sequencing

 ,
, 
 ,
,  , , ,
, , ,
Abstract
1. Introduction

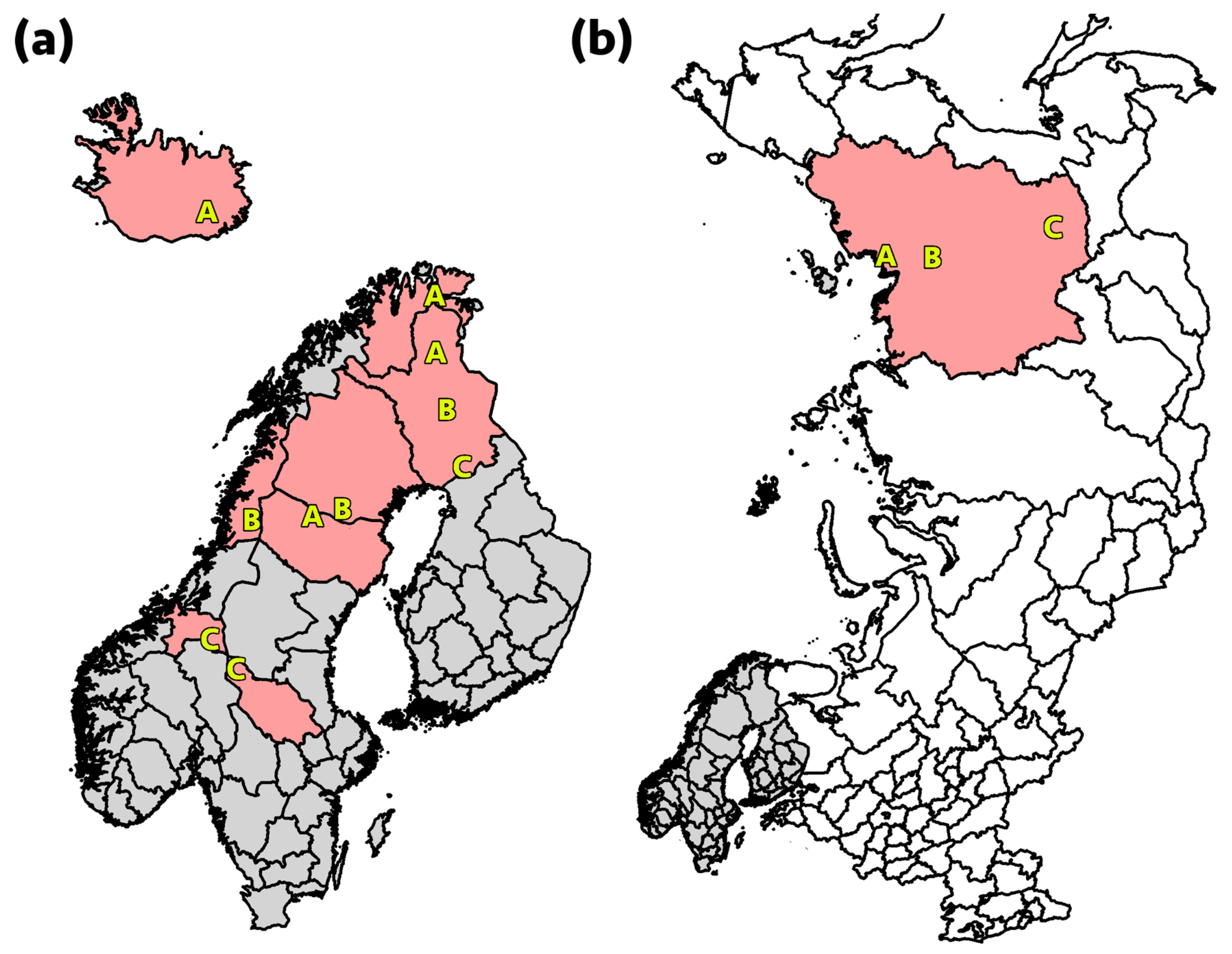{kind=link}
{kind=link}
| Virus | Information | References |
|---|---|---|
| Flaviviridae | Serological studies have reported pestivirus antibodies in reindeer from Finland, Norway, Sweden, and Iceland, as well as in caribou from Canada. The clinical relevance of pestivirus infections in reindeer is unknown. It may, however, be reasonable to assume that also reindeer may be persistently infected, with abortion, stillbirth, and the birth of persistently virus shedding offspring (i.e., persistently infected animals), as seen for many other host species. West Nile virus has also been demonstrated to infect reindeer, causing clinical disease. | [11,12,13,14,15] |
| Herpesviridae Alphaherpesvirinae | Cervid herpesvirus 2 (CvHV2) is enzootic in the Fennoscandian reindeer populations and antibodies against alphaherpesvirus have also been detected in caribou in Alaska (USA) and Canada. CvHV2 has been shown to act as the primary cause of infectious keratoconjunctivitis in reindeer during outbreaks and after experimental ocular inoculation, although many types of bacteria may contribute to the disease. CvHV2 may also cause respiratory infections in reindeer, and possibly abortion and weak-borne calves. | [16,17,18,19,20,21,22] |
| Herpesviridae Gammaherpesvirinae Genus Macavirus | The subfamily Gammaherpesvirinae contains several closely related virus species that are associated with malignant catarrhal fever (MCF). Sheep and goats are healthy carriers of ovine herpesvirus 2 and caprine herpesvirus 2, respectively, but may transmit the virus to susceptible domestic and wild ruminants. One clinical case of MCF in reindeer has been reported. The recorded symptoms were hair loss and thickening of the skin, with crusts in the axillary region, distal parts of the feet, and on the muzzle. Further, the animal had swollen eyelids, opaque cornea and fibrinopurulent eye discharge. | [23,24,25] |
| Papillomaviridae | Papillomaviruses cause mostly benign processes in the skin (papillomas, fibropapillomas or warts) or mucous membranes (condylomas) in many animal species, including reindeer. The clinical outcome may be serious for the individual. Papilloma viruses are considered species-specific, but several virus species may circulate in the same host species. The prevalence of papilloma viruses in reindeer is scarce. Generalized papillomatosis has been reported, affecting the skin in coalescing warts all over the body. | [25,26,27] |
| Poxviridae Genus Parapoxvirus | Orf virus (ORFV) and pseudocowpoxvirus (PCPV) have small ruminants and cattle as their main reservoirs. ORFV cause contagious ecthyma in and around the mouth in sheep and goats, and a similar disease has been reported in reindeer in Sweden, Finland, and Norway. Early outbreaks in Finland were caused by ORFV, whereas later outbreaks, from 1999–2000, have been associated with PCPV, with a milder clinical appearance as compared to ORFV. | [25,28,29,30,31] |
2. Materials and Methods
2.1. Ethical Statement
2.2. Sample Collection
2.3. Nucleic Acid Extraction
2.4. NGS Library Preparation and Sequencing
2.5. Bioinformatics
3. Results
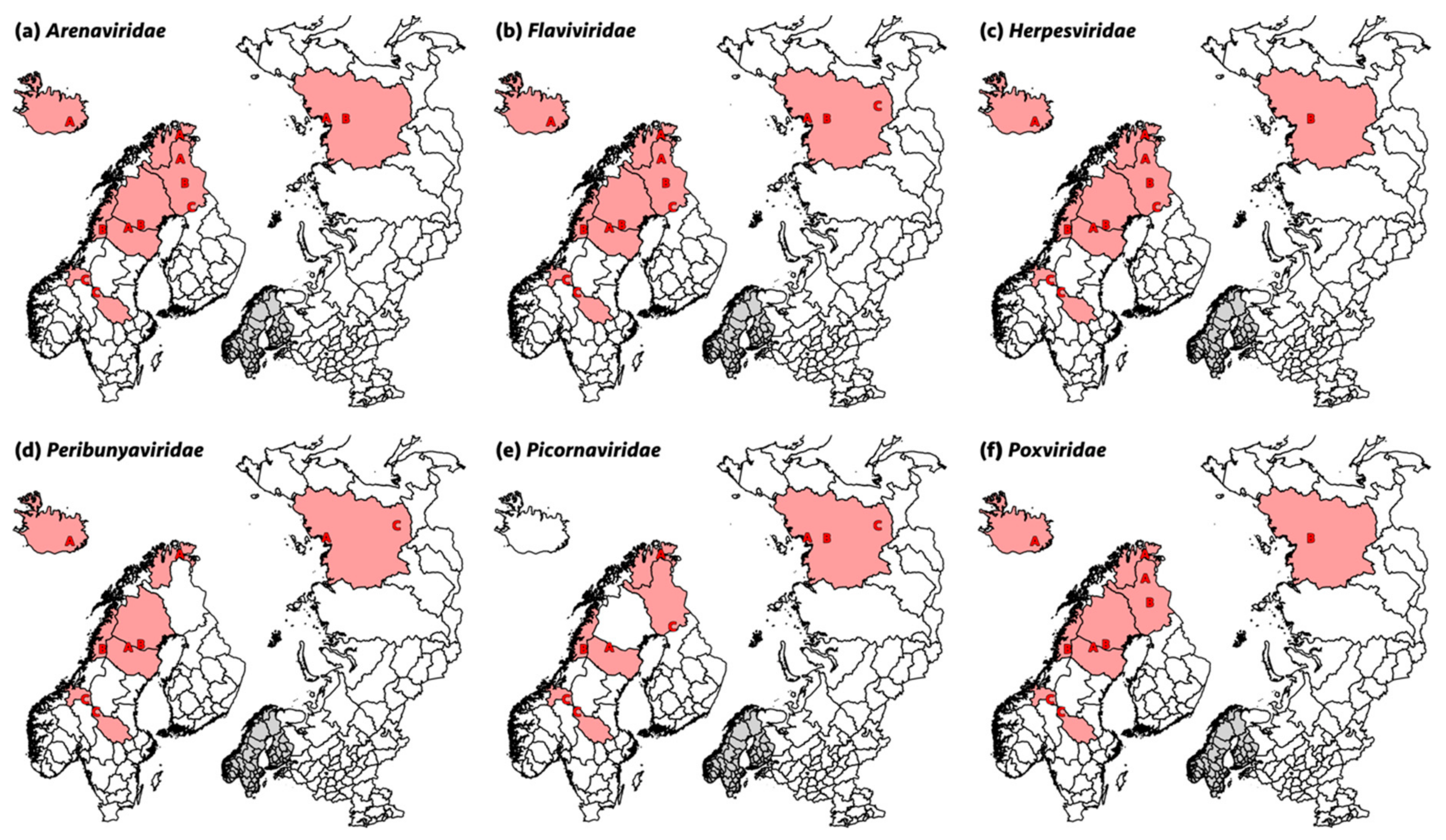
3.1. Arenaviridae
3.2. Flaviviridae
3.3. Herpesviridae
3.4. Papillomaviridae
3.5. Paramyxoviridae
3.6. Parvoviridae
3.7. Peribunyaviridae
3.8. Picobirnaviridae
3.9. Picornaviridae
3.10. Poxviridae
3.11. Small Circular DNA Viruses
3.12. Other Viruses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zell, R. Global climate change and the emergence/re-emergence of infectious diseases. Int. J. Med. Microbiol. Suppl. 2004, 293, 16–26. [Google Scholar] [CrossRef]
- Xu, L.; Myneni, R.; Chapin, F.S., III; Callaghan, T.V.; Pinzon, J.E.; Tucker, C.J.; Zhu, Z.; Bi, J.; Ciais, P.; Tømmervik, H.; et al. Temperature and vegetation seasonality diminishment over northern lands. Nat. Clim. Chang. 2013, 3, 581–586. [Google Scholar] [CrossRef]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Jones, K.; Patel, N.G.; Levy, M.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Thórisson, S. The history of reindeer in Iceland and reindeer study 1979–1981. Rangifer 1984, 4, 22–38. [Google Scholar] [CrossRef][Green Version]
- Forbes, B.C.; Kumpula, T.; Meschtyb, N.; Laptander, R.; Macias-Fauria, M.; Zetterberg, P.; Verdonen, M.; Skarin, A.; Kim, K.-Y.; Boisvert, L.N.; et al. Sea ice, rain-on-snow and tundra reindeer nomadism in Arctic Russia. Biol. Lett. 2016, 12, 20160466. [Google Scholar] [CrossRef]
- Risvoll, C.; Hovelsrud, G.K. Pasture access and adaptive capacity in reindeer herding districts in Nordland, Northern Norway. Polar J. 2016, 6, 87–111. [Google Scholar] [CrossRef]
- Sánchez Romano, J.; Leijon, M.; Hagström, Å.; Jinnerot, T.; Rockström, U.K.; Tryland, M. Chlamydia pecorum associated with an outbreak of infectious keratoconjunctivitis in semi-domesticated reindeer in Sweden. Front. Vet. Sci. 2019, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Tryland, M.; Nymo, I.H.; Sánchez Romano, J.; Mørk, T.; Klein, J.; Rockström, U. Infectious disease outbreak associated with supplementary feeding of semi-domesticated reindeer. Front. Vet. Sci. 2019, 6, 126. [Google Scholar] [CrossRef]
- Dudley, J.P.; Hoberg, E.P.; Jenkins, E.J.; Parkinson, A.J. Climate change in the North American Arctic: A one health perspective. EcoHealth 2015, 12, 713–725. [Google Scholar] [CrossRef]
- Macdonald, E.; Handeland, K.; Blystad, H.; Bergsaker, M.; Fladberg, M.; Gjerset, B.; Nilsen, O.; Os, H.; Sandbu, S.; Stokke, E.; et al. Public health implications of an outbreak of rabies in arctic foxes and reindeer in the Svalbard archipelago, Norway, September 2011. Eurosurveillance 2011, 16, 19985. [Google Scholar] [CrossRef]
- Palmer, M.V.; Stoffregen, W.C.; Rogers, U.G.; Hamir, A.N.; Richt, J.A.; Pedersen, D.D.; Waters, W.R. West Nile virus infection in reindeer (Rangifer Tarandus). J. Vet. Diagn. Investig. 2004, 16, 219–222. [Google Scholar] [CrossRef]
- Larska, M. Pestivirus infection in reindeer (Rangifer Tarandus). Front. Microbiol. 2015, 6, 1187. [Google Scholar] [CrossRef]
- Omazic, A.; Aurosell, C.; Fedorov, V.; Hagström, Å.; Kantanen, J.; Leijon, M.; Mørk, T.; Nordtun, C.S.; Nymo, I.H.; Þórisson, S.G.; et al. Seroprevalence of pestivirus in Eurasian tundra reindeer in Finland, Sweden, Norway, Iceland and Russian Federation. Infect. Ecol. Epidemiol. 2019, 9, 1682223. [Google Scholar] [CrossRef]
- Das Neves, C.G.; Wensman, J.J.; Nymo, I.H.; Skjerve, E.; Alenius, S.; Tryland, M. Pestivirus infections in semi-domesticated Eurasian tundra reindeer (Rangifer tarandus tarandus): A retrospective cross-sectional serological study in Finnmark county, Norway. Viruses 2019, 12, 29. [Google Scholar] [CrossRef]
- das Neves, C.G.; Roth, S.; Rimstad, E.; Thiry, E.; Tryland, M. Cervid herpesvirus 2 infection in reindeer: A review. Vet. Microbiol. 2010, 143, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.L.; das Neves, C.G.; Finstad, G.F.; Beckmen, K.B.; Skjerve, E.; Nymo, I.H.; Tryland, M. Evidence of alphaherpesvirus infections in Alaskan caribou and reindeer. BMC Vet. Res. 2012, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, A.M.; Curry, P.; Elkin, B.; Russell, D.; Veitch, A.; Branigan, M.; Campbell, M.; Croft, B.; Cuyler, C.; Côté, S.D.; et al. Multi-pathogen serological survey of migratory caribou herds: A snapshot in time. PLoS ONE 2019, 14, e0219838. [Google Scholar] [CrossRef]
- Tryland, M.; Das Neves, C.G.; Sunde, M.; Mørk, T. Cervid herpesvirus 2, the primary agent in an outbreak of infectious keratoconjunctivitis in semidomesticated reindeer. J. Clin. Microbiol. 2009, 47, 3707–3713. [Google Scholar] [CrossRef] [PubMed]
- Tryland, M.; Romano, J.S.; Marcin, N.; Nymo, I.H.; Josefsen, T.D.; Sørensen, K.K.; Mørk, T. Cervid herpesvirus 2 and not Moraxella bovoculi caused keratoconjunctivitis in experimentally inoculated semi-domesticated Eurasian tundra reindeer. Acta Vet. Scand. 2017, 59, 23. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Romano, J.; Mørk, T.; Laaksonen, S.; Ågren, E.; Nymo, I.H.; Sunde, M.; Tryland, M. Infectious keratoconjunctivitis in semi-domesticated Eurasian tundra reindeer (Rangifer tarandus tarandus): Microbiological study of clinically affected and unaffected animals with special reference to cervid herpesvirus 2. BMC Vet. Res. 2018, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Romano, J.; Sørensen, K.K.; Larsen, A.K.; Mørk, T.; Tryland, M. Ocular histopathological findings in semi-domesticated Eurasian tundra reindeer (Rangifer tarandus tarandus) with infectious keratoconjunctivitis after experimental inoculation with cervid herpesvirus 2. Viruses 2020, 12, 1007. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2008, 154, 171–177. [Google Scholar] [CrossRef]
- Li, H.; McGuire, T.C.; Müller-Doblies, U.U.; Crawford, T.B. A simpler, more sensitive competitive inhibition enzyme-linked immunosorbent assay for detection of antibody to malignant catarrhal fever viruses. J. Vet. Diagn. Investig. 2001, 13, 361–364. [Google Scholar] [CrossRef]
- Tryland, M.; Das Neves, C.G.; Klein, J.; Mørk, T.; Hautaniemi, M.; Wensman, J.J. Viral infections and diseases. In Reindeer and Caribou—Health and Disease; Tryland, M., Kutz, S.J., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 273–3034. [Google Scholar]
- Moreno-López, J.; Mörner, T.; Pettersson, U. Papillomavirus DNA associated with pulmonary fibromatosis in European elks. J. Virol. 1986, 57, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.L.; Schapendonk, C.M.E.; Van Leeuwen, M.; Kuiken, T.; Bodewes, R.; Raj, V.S.; Haagmans, B.L.; Das Neves, C.G.; Tryland, M.; Osterhaus, A. Identification and characterization of two novel viruses in ocular infections in reindeer. PLoS ONE 2013, 8, e69711. [Google Scholar] [CrossRef] [PubMed]
- Büttner, M.; Von Einem, C.; McInnes, C.; Oksanen, A. Clinical findings and diagnosis of a severs parapoxvirus epidemic in Finnish reindeer. Tierarztl. Prax. 1995, 23, 614–618. [Google Scholar]
- Tryland, M.; Oksanen, A.; Aschfalk, A.; Josefsen, T.D. Parapoxvirus infection in Norwegian semi-domesticated reindeer (Rangifer tarandus tarandus). Vet. Rec. 2001, 149, 394–395. [Google Scholar] [CrossRef]
- Kummeneje, K.; Krogsrud, J. Contagious ecthyma (orf) in reindeer (Rangifer tarandus). Vet. Rec. 1979, 105, 60–61. [Google Scholar] [CrossRef]
- Nordkvist, M. Munvårtsjuka-En ny rensjukdom? Rennäringsnytt 1973, 8, 6–8. [Google Scholar]
- Jore, S.; Vanwambeke, S.O.; Viljugrein, H.; Isaksen, K.; Kristoffersen, A.B.; Woldehiwet, Z.; Johansen, B.; Brun, E.; Brun-Hansen, H.; Westermann, S.; et al. Climate and environmental change drives Ixodes ricinus geographical expansion at the northern range margin. Parasites Vectors 2014, 7, 11. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Jaenson, D.G.; Eisen, L.; Petersson, E.; Lindgren, E. Changes in the geographical distribution and abundance of the tick Ixodes ricinus during the past 30 years in Sweden. Parasites Vectors 2012, 5, 8. [Google Scholar] [CrossRef]
- Ogryzkov, S.E. The pathology of foot and mouth disease in reindeer. Trudy II Vses. Konf. Patol. Anat. Zhivotnykh Mosk. Vet Akad. 1964, 420–425. [Google Scholar]
- Rehbinder, C.; Belák, S.; Nordkvist, M. A serological, retrospective study in reindeer on five different viruses. Rangifer 1992, 12, 191. [Google Scholar] [CrossRef]
- Schürch, A.C.; Schipper, D.; Bijl, M.A.; Dau, J.; Beckmen, K.B.; Schapendonk, C.M.E.; Raj, V.S.; Osterhaus, A.D.M.E.; Haagmans, B.L.; Tryland, M.; et al. Metagenomic survey for viruses in Western Arctic Caribou, Alaska, through iterative assembly of taxonomic units. PLoS ONE 2014, 9, e105227. [Google Scholar] [CrossRef]
- Johansson, Ö.; Ullman, K.; Lkhagvajav, P.; Wiseman, M.; Malmsten, J.; Leijon, M. Detection and genetic characterization of viruses present in free-ranging snow leopards using next-generation sequencing. Front. Vet. Sci. 2020, 7, 645. [Google Scholar] [CrossRef]
- Blomström, A.-L.; Widén, F.; Hammer, A.-S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [PubMed]
- HTStream. 2020. Available online: https://s4hts.github.io/HTStream/ (accessed on 30 March 2021).
- National Center for Biotechnology Information (NCBI). 1988. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 30 March 2021).
- Li, L.; Kapoor, A.; Slikas, B.; Bamidele, O.S.; Wang, C.; Shaukat, S.; Alam Masroor, M.; Wilson, M.L.; Ndjango, J.-B.N.; Peeters, M.; et al. Multiple diverse circoviruses infect farm animals and are commonly found in human and chimpanzee feces. J. Virol. 2010, 84, 1674–1682. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Romano, J.; Grund, L.; Obiegala, A.; Nymo, I.H.; Ancin-Murguzur, F.J.; Li, H.; Król, N.; Pfeffer, M.; Tryland, M. A multi-pathogen screening of captive reindeer (Rangifer tarandus) in Germany based on serological and molecular assays. Front. Vet. Sci. 2019, 6, 461. [Google Scholar] [CrossRef]
- Slukinova, O.S.; Kyuregyan, K.K.; Karlsen, A.A.; Potemkin, I.A.; Kichatova, V.S.; Semenov, S.I.; Stepanov, K.M.; Rumyantseva, T.D.; Mikhailov, M.I. Serological evidence of Hepatitis E virus circulation among reindeer and reindeer herders. Vector Borne Zoonotic Dis. 2021. [Google Scholar] [CrossRef]
- Das Neves, C.G.; Ihlebæk, H.M.; Skjerve, E.; Hemmingsen, W.; Li, H.; Tryland, M. Gammaherpesvirus infection in semidomesticated reindeer (Rangifer Tarandus Tarandus): A cross-sectional, serologic study in Northern Norway. J. Wildl. Dis. 2013, 49, 261–269. [Google Scholar] [CrossRef]
- das Neves, C.G.; Sacristán, C.; Madslien, K.; Tryland, M. Gammaherpesvirus in cervid species from Norway: Characterization of a new virus in wild and semi-domesticated Eurasian tundra reindeer (Rangifer tarandus tarandus). Viruses 2020, 12, 876. [Google Scholar] [CrossRef] [PubMed]
- Box, J.; Colgan, W.T.; Christensen, T.R.; Schmidt, N.M.; Lund, M.; Parmentier, F.-J.W.; Brown, R.; Bhatt, U.S.; Euskirchen, E.S.; Romanovsky, V.E.; et al. Key indicators of Arctic climate change: 1971–2017. Environ. Res. Lett. 2019, 14, 045010. [Google Scholar] [CrossRef]
- Hoberg, E.P.; Brooks, D.R. Evolution in action: Climate change, biodiversity dynamics and emerging infectious disease. Philos. Trans. R. Soc. B. Biol. Sci. 2015, 370, 20130553. [Google Scholar] [CrossRef]
- Pauchard, A.; Milbau, A.; Albihn, A.; Alexander, J.M.; Burgess, T.; Daehler, C.C.; Englund, G.; Essl, F.; Evengård, B.; Greenwood, G.B.; et al. Non-native and native organisms moving into high elevation and high latitude ecosystems in an era of climate change: New challenges for ecology and conservation. Biol. Invasions 2016, 18, 345–353. [Google Scholar] [CrossRef]
- Omazic, A.; Bylund, H.; Boqvist, S.; Högberg, A.; Björkman, C.; Tryland, M.; Evengård, B.; Koch, A.; Berggren, C.; Malogolovkin, A.; et al. Identifying climate-sensitive infectious diseases in animals and humans in Northern regions. Acta Vet. Scand. 2019, 61, 1–12. [Google Scholar] [CrossRef]
- Jaenson, T.G.T.; Eisen, L.; Comstedt, P.; Mejlon, H.A.; Lindgren, E.; Bergström, S.; Olsen, B. Risk indicators for the tick Ixodes ricinus and Borrelia burgdorferi sensu lato in Sweden. Med. Vet. Entomol. 2009, 23, 226–237. [Google Scholar] [CrossRef]
- Hugot, J.; Gonzalez, J.-P.; Denys, C. Evolution of the old world Arenaviridae and their rodent hosts: Generalized host-transfer or association by descent? Infect. Genet. Evol. 2001, 1, 13–20. [Google Scholar] [CrossRef]
- Desfarges, S.; Ciuffi, A. Viral integration and consequences on host gene expression. In Viruses: Essential Agents of Life; Springer: Berlin, Germany, 2012; pp. 147–175. [Google Scholar]
- Charrel, R.; de Lamballerie, X. Zoonotic aspects of arenavirus infections. Vet. Microbiol. 2010, 140, 213–220. [Google Scholar] [CrossRef]
- Root, J.J.; Bosco-Lauth, A.M. West Nile virus associations in wild mammals: An update. Viruses 2019, 11, 459. [Google Scholar] [CrossRef]
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, migration and emerging zoonoses: West Nile virus, Lyme disease, Influenza A and Enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef]
- Bakonyi, T.; Haussig, J.M. West Nile virus keeps on moving up in Europe. Eurosurveillance 2020, 25, 2001938. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. BioMed Res. Int. 2015, 2015, 1–20. [Google Scholar] [CrossRef]
- Fyodorova, M.V.; Savage, H.M.; Lopatina, J.V.; Bulgakova, T.A.; Ivanitsky, A.V.; Platonova, O.V.; Platonov, A.E. Evaluation of potential West Nile virus vectors in Volgograd region, Russia, 2003 (Diptera: Culicidae): Species composition, bloodmeal host utilization, and virus infection rates of mosquitoes. J. Med. Entomol. 2006, 43, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Lindström, A.; Lilja, T. First finding of the West Nile virus vector Culex modestus Ficalbi 1889 (Diptera; Culicidae) in Sweden. J. Eur. Mosq. Control. Assoc. 2018, 36, 1–2. [Google Scholar]
- Jourdain, E.; Olsen, B.; Lundkvist, A.; Hubálek, Z.; Šikutová, S.; Waldenström, J.; Karlsson, M.; Wahlström, M.; Jozan, M.; Falk, K.I. Surveillance for West Nile virus in wild birds from northern Europe. Vector Borne Zoonotic Dis. 2011, 11, 77–79. [Google Scholar] [CrossRef]
- OIE. Classical Swine Fever. 2020. Available online: https://www.oie.int/en/disease/classical-swine-fever/ (accessed on 30 March 2021).
- Titov, I.; Tsybanov, S.; Malogolovkin, A. Genotyping of classical swine fever virus using high-resolution melt analysis. J. Virol. Methods 2015, 224, 53–57. [Google Scholar] [CrossRef]
- Lindberg, A.L.; Alenius, S. Principles for eradication of bovine viral diarrhoea virus (BVDV) infections in cattle populations. Vet. Microbiol. 1999, 64, 197–222. [Google Scholar] [CrossRef]
- Becher, P.; Orlich, M.; Kosmidou, A.; König, M.; Baroth, M.; Thiel, H.-J. Genetic diversity of pestiviruses: Identification of novel groups and implications for classification. Virology 1999, 262, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Das Neves, C.G.; Thiry, J.; Skjerve, E.; Yoccoz, N.G.; Rimstad, E.; Thiry, E.; Tryland, M. Alphaherpesvirus infections in semidomesticated reindeer: A cross-sectional serological study. Vet. Microbiol. 2009, 139, 262–269. [Google Scholar] [CrossRef]
- Kautto, A.H.; Alenius, S.; Mossing, T.; Becher, P.; Belák, S.; Larska, M. Pestivirus and alphaherpesvirus infections in Swedish reindeer (Rangifer tarandus tarandus L.). Vet. Microbiol. 2012, 156, 64–71. [Google Scholar] [CrossRef]
- Thiry, J.; Keuser, V.; Muylkens, B.; Meurens, F.; Gogev, S.; Vanderplasschen, A.; Thiry, E. Ruminant alphaherpesviruses related to bovine herpesvirus 1. Vet. Res. 2006, 37, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, S.; Kong, X. Characterization of the genes encoding UL24, TK and gH proteins from duck enteritis virus (DEV): A proof for the classification of DEV. Virus Genes 2006, 33, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.R.; Adkins, S.; Alkhovskiy, S.; Beer, M.; Blair, C.; Calisher, C.H.; Drebot, M.; Lambert, A.J.; De Souza, W.M.; Marklewitz, M.; et al. ICTV virus taxonomy profile: Peribunyaviridae. J. Gen. Virol. 2020, 101, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Chenais, E.; Stahl, K.; Frössling, J.; Blomqvist, G.; Näslund, K.; Svensson, L.; Renström, L.; Mieziewska, K.; Elvander, M.; Valarcher, J.F. Schmallenberg virus beyond latitude 65° N. Transbound. Emerg. Dis. 2013, 62, e11–e18. [Google Scholar] [CrossRef]
- Malmsten, A.; Malmsten, J.; Blomqvist, G.; Näslund, K.; Vernersson, C.; Hägglund, S.; Dalin, A.-M.; Ågren, E.O.; Valarcher, J.-F. Serological testing of Schmallenberg virus in Swedish wild cervids from 2012 to 2016. BMC Vet. Res. 2017, 13, 84. [Google Scholar] [CrossRef]
- European Food Safety Authorities. “Schmallenberg” virus: Analysis of the epidemiological data and assessment of impact. EFSA J. 2012, 10, 2768. [Google Scholar]
- Clima Temps. Sweden Climate Graph. Available online: http://www.sweden.climatemps.com/#brief (accessed on 29 March 2021).
- Rima, B.; Balkema-Buschmann, A.; Dundon, W.G.; Duprex, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.; Lee, B.; Rota, P.; et al. ICTV virus taxonomy profile: Paramyxoviridae. J. Gen. Virol. 2019, 100, 1593–1594. [Google Scholar] [CrossRef]
- Berns, K.I.; Parrish, C.R. Parvoviridae. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 2437–2477. [Google Scholar]
- Delmas, B.; Attoui, H.; Ghosh, S.; Malik, Y.S.; Mundt, E.; Vakharia, V.N. ICTV report consortium ICTV virus taxonomy profile: Picobirnaviridae. J. Gen. Virol. 2019, 100, 133–134. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Di Felice, E.; Robetto, S.; Guidetti, C.; Orusa, R.; Martella, V.; Marsilio, F. Molecular detection of kobuviruses in European roe deer (Capreolus capreolus) in Italy. Arch. Virol. 2015, 160, 2083–2086. [Google Scholar] [CrossRef]
- FAO. Foot and Mouth Disease Situation—March 2019; European Commission for the Control of Foot and Mouth Disease: Rome, Italy, 2019. [Google Scholar]
- EFSA Panel on Animal Health and Welfare. Scientific opinion on foot and mouth disease in Thrace. EFSA J. 2012, 10, 2635. [Google Scholar]
- Rahman, A.-U.; Dhama, K.; Ali, Q.; Raza, M.A.; Chaudhry, U.; Shabbir, M.Z. Foot and mouth disease in a wide range of wild hosts: A potential constraint in disease control efforts worldwide particularly in disease-endemic settings. Acta Trop. 2020, 210, 105567. [Google Scholar] [CrossRef]
- Tryland, M. Asymptomatic parapoxvirus infections in semi-domesticated reindeer (Rangifer tarandus tarandus). In Proceedings of the XIVth International Poxvirus and Iridovirus Workshop, Lake Placid, NY, USA, 20–25 September 2002; pp. 20–25. [Google Scholar]
- Tryland, M.; Beckmen, K.B.; Burek-Huntington, K.A.; Breines, E.M.; Klein, J. Orf virus infection in Alaskan mountain goats, Dall’s sheep, muskoxen, caribou and Sitka black-tailed deer. Acta Vet. Scand. 2018, 60, 1–11. [Google Scholar] [CrossRef]
- Haig, D.M. Orf virus infection and host immunity. Curr. Opin. Infect. Dis. 2006, 19, 127–131. [Google Scholar] [CrossRef]
- Fleming, S.B.; McCaughan, C.A.; Andrews, A.E.; Nash, A.D.; Mercer, A.A. A homolog of interleukin-10 is encoded by the poxvirus Orf virus. J. Virol. 1997, 71, 4857–4861. [Google Scholar] [CrossRef]
- Danilov, P.I.; Panchenko, D.V.; Tirronen, K.F. The European roe deer (Capreolus capreolus L.) at the northern boundary of its range in Eastern Fennoscandia. Russ. J. Ecol. 2017, 48, 459–465. [Google Scholar] [CrossRef]
- Rosvold, J.; Andersen, R. Wild boar in Norway—Is climate a limiting factor? NTNU Vitensk. Rapp. Zool. 2008, 1, 1–23. [Google Scholar]
- Rossi, S.; Viarouge, C.; Faure, E.; Gilot-Fromont, E.; Gache, K.; Gibert, P.; Verheyden, H.; Hars, J.; Klein, F.; Maillard, D.; et al. Exposure of wildlife to the Schmallenberg virus in France (2011–2014): Higher, faster, stronger (than bluetongue). Transbound. Emerg. Dis. 2015, 64, 354–363. [Google Scholar] [CrossRef]
- Ruiz-Fons, F.; Segalés, J.; Gortázar, C. A review of viral diseases of the European wild boar: Effects of population dynamics and reservoir rôle. Vet. J. 2008, 176, 158–169. [Google Scholar] [CrossRef]


| Sampling Site | Sampling 1 | Sampling 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Sampling | Total no. of Reindeer | No. of Calves | No. of Adults | Time of Sampling | Total no. of Reindeer | No. of Calves | No. of Adults | |
| Finland, A | December 2016 | 19 1 | 10 | 9 | November 2017 | 22 | 13 | 9 |
| Finland, B | January 2017 | 20 1 | 14 | 6 | October 2017 | 20 | 10 | 10 |
| Finland, C | February 2017 | 21 1 | 10 | 11 | October 2017 | 20 | 10 | 10 |
| Norway, A | November 2016 | 20 1 | 10 | 10 | November 2017 | 20 | 11 | 9 |
| Norway, B | January 2017 | 20 1 | 10 | 10 | April 2018 | 21 | 11 | 10 |
| Norway, C | January 2017 | 20 1 | 10 | 10 | January 2018 | 20 | 10 | 10 |
| Sweden, A | December 2016 | 20 | 10 | 10 | December 2017 | 20 | 10 | 10 |
| Sweden, B | November 2016 | 33 2 | 14 | 9 | December 2017 | 20 | 10 | 10 |
| Sweden, C | November 2016 | 19 1 | 9 | 10 | November 2017 | 20 | 10 | 10 |
| Iceland | August 2017 | 25 | 2 | 23 | September 2018 | 24 | 0 | 24 |
| Russia, A | December 2017 | 20 | 4 | 16 | n/a | n/a | n/a | n/a |
| Russia, B | n/a | n/a | n/a | n/a | Nov 2019 | 20 | 0 | 20 |
| Russia, C | n/a | n/a | n/a | n/a | Nov 2019 | 20 | 0 | 20 |
| Total | 237 | 103 | 124 | 247 | 95 | 152 | ||
| Sampling Site | Sampling 1 | Sampling 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Sampling | Total no. of Reindeer | No. of Calves | No. of Adults | Time of Sampling | Total no. of Reindeer | No. of Calves | No. of Adults | |
| Finland, A | December 2016 | 19 | 10 | 9 | November 2017 | 21 | 13 | 8 |
| Finland, B | January 2017 | 21 | 14 | 7 | October 2017 | 20 | 10 | 10 |
| Finland, C | February 2017 | 21 | 10 | 11 | October 2017 | 20 | 10 | 10 |
| Norway, A | November 2016 | 20 | 10 | 10 | November 2017 | 19 | 11 | 8 |
| Norway, B | January 2017 | 20 | 10 | 10 | April 2018 | 20 | 10 | 10 |
| Norway, C | January 2017 | 20 | 10 | 10 | January 2018 | 19 | 9 | 10 |
| Sweden, A | December 2016 | 20 | 10 | 10 | December 2017 | 20 | 10 | 10 |
| Sweden, B | November 2016 | 30 1 | 13 | 7 | December 2017 | 20 | 10 | 10 |
| Sweden, C | November 2016 | 20 | 10 | 10 | November 2017 | 20 | 10 | 10 |
| Iceland | August 2017 | 25 | 1 | 24 | September 2018 | 22 | 0 | 22 |
| Russia, A | December 2017 | 20 | 4 | 16 | n/a | n/a | n/a | n/a |
| Russia, B | n/a | n/a | n/a | n/a | Nov 2019 | 20 | 0 | 20 |
| Russia, C | n/a | n/a | n/a | n/a | Nov 2019 | 20 | 0 | 20 |
| Total | 236 | 102 | 124 | 241 | 93 | 148 | ||
| Virus Family | Sweden | Norway | Finland | Iceland | Russia | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling 1 | Sampling 2 | Sampling 1 | Sampling 2 | Sampling 1 | Sampling 2 | Sampling 1 | Sampling 2 | Sampling 1 | Sampling 2 | ||||||||||||||
| A | B | C | A | B | C | A | B | C | A | B | C | A | B | C | A | B | C | A | A | A | B | C | |
| Adenoviridae | X | X | X | X | |||||||||||||||||||
| Arenaviridae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||
| Astroviridae | X | X | X | X | |||||||||||||||||||
| Caliciviridae | X | ||||||||||||||||||||||
| Flaviviridae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||
| Herpesviridae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||
| Papillomaviridae | X | X | X | X | X | X | X | X | X | X | X | ||||||||||||
| Paramyxoviridae | X | X | X | X | X | X | X | X | X | X | |||||||||||||
| Parvoviridae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||
| Peribunyaviridae | X | X | X | X | X | X | X | X | X | X | X | X | |||||||||||
| Picobirnaviridae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||||
| Picornaviridae | X | X | X | X | X | X | X | X | X | X | |||||||||||||
| Poxviridae | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||||||||
| Small circular DNA viruses | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez Romano, J.; Omazic, A.; Leijon, M.; Hagström, Å.; Tryland, M.; Kantanen, J.; Reilas, T.; Rockström, U.; Fedorov, V.; Albihn, A. Screening of Eurasian Tundra Reindeer for Viral Sequences by Next-Generation Sequencing. Int. J. Environ. Res. Public Health 2021, 18, 6561. https://doi.org/10.3390/ijerph18126561
Sánchez Romano J, Omazic A, Leijon M, Hagström Å, Tryland M, Kantanen J, Reilas T, Rockström U, Fedorov V, Albihn A. Screening of Eurasian Tundra Reindeer for Viral Sequences by Next-Generation Sequencing. International Journal of Environmental Research and Public Health. 2021; 18(12):6561. https://doi.org/10.3390/ijerph18126561
Chicago/Turabian StyleSánchez Romano, Javier, Anna Omazic, Mikael Leijon, Åsa Hagström, Morten Tryland, Juha Kantanen, Tiina Reilas, Ulrika Rockström, Valery Fedorov, and Ann Albihn. 2021. "Screening of Eurasian Tundra Reindeer for Viral Sequences by Next-Generation Sequencing" International Journal of Environmental Research and Public Health 18, no. 12: 6561. https://doi.org/10.3390/ijerph18126561
APA StyleSánchez Romano, J., Omazic, A., Leijon, M., Hagström, Å., Tryland, M., Kantanen, J., Reilas, T., Rockström, U., Fedorov, V., & Albihn, A. (2021). Screening of Eurasian Tundra Reindeer for Viral Sequences by Next-Generation Sequencing. International Journal of Environmental Research and Public Health, 18(12), 6561. https://doi.org/10.3390/ijerph18126561