Female Fertility and Environmental Pollution


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. How Environmental Pollution Affects Female Fertility
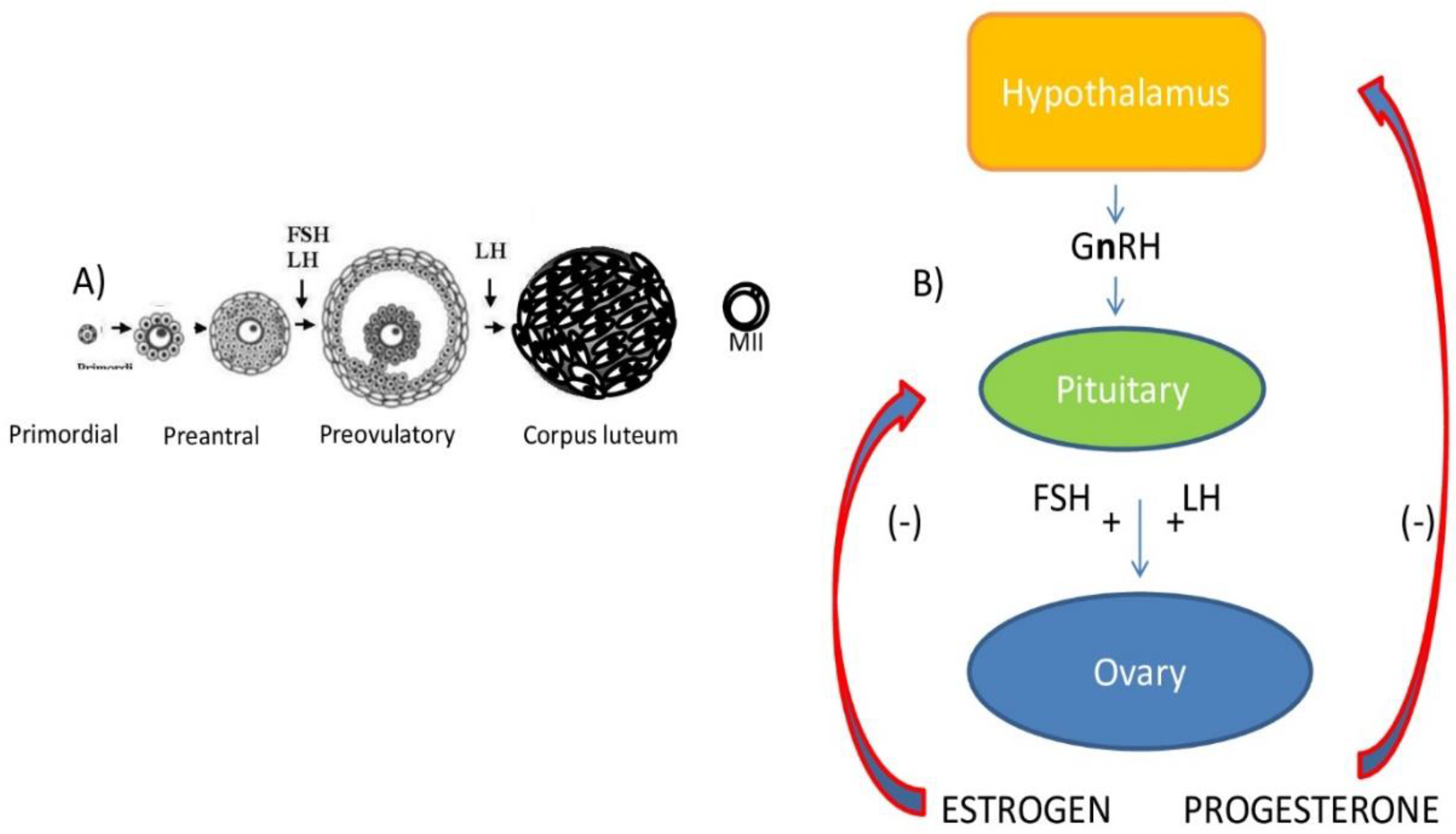
3. Overview of Mammalian Oogenesis
3.1. Toxicity of Heavy Metals (HMs)
3.1.1. Lead (Pb)
3.1.2. Cadmium (Cd)
3.1.3. Mercury (Hg)
3.1.4. Zinc (Zn), Cobalt (Co) and Nickel (Ni)
4. Air Pollution
5. EDs: Phthalates and Bisphenol A (BPA)
6. Thermal Stress
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Muralikrishna, I.V.; Manickam, V. Science and Engineering for Industry. Environmental Management; Butterworth-Heinemann: Waltham, MA, USA, 2017; pp. 1–4. [Google Scholar]
- Rai, P.K. Particulate Matter and Its Size Fractionation. In Biomagnetic Monitoring of Particulate Matter, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–13. [Google Scholar]
- Borght, M.V.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Di Nisio, V.; Macchiarelli, G.; A Nottola, S.; Halvaei, I.; De Santis, L.; Cecconi, S. Technologies for the Production of Fertilizable Mammalian Oocytes. Appl. Sci. 2019, 9, 1536. [Google Scholar] [CrossRef]
- Quaas, A.; Dokras, A. Diagnosis and treatment of unexplained infertility. Rev. Obstet. Gynecol. 2008, 1, 69–76. [Google Scholar] [PubMed]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Kahn, L.G.; Philippat, C.; Nakayama, S.F.; Slama, R.; Trasande, L. Endocrine-disrupting chemicals: Implications for human health. Lancet Diabetes Endocrinol. 2020, 8, 703–718. [Google Scholar] [CrossRef]
- Hipwell, A.E.; Kahn, L.G.; Factor-Litvak, P.; Porucznik, C.A.; Siegel, E.L.; Fichorova, R.N.; Hamman, R.F.; Klein-Fedyshin, M.; Harley, K.G. Program collaborators for Environmental influences on Child Health Outcomes Exposure to non-persistent chemicals in consumer products and fecundability: A systematic review. Hum. Reprod. Updat. 2018, 25, 51–71. [Google Scholar] [CrossRef]
- Skakkebaek, N.E. Testicular Dysgenesis Syndrome. Horm. Res. 2003, 60, 49. [Google Scholar] [CrossRef]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Hunt, P.A.; Hassold, T.J. Human female meiosis: What makes a good egg go bad? Trends Genet. 2008, 24, 86–93. [Google Scholar] [CrossRef]
- Canipari, R.; Cellini, V.; Cecconi, S. The ovary feels fine when paracrine and autocrine networks cooperate with gonadotropins in the regulation of folliculogenesis. Curr. Pharm. Des. 2012, 18, 245–255. [Google Scholar] [PubMed]
- Jamnongjit, M.; Hammes, S.R. Ovarian Steroids: The Good, the Bad, and the Signals that Raise Them. Cell Cycle 2006, 5, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Zuccotti, M.; Merico, V.; Cecconi, S.; Redi, C.A.; Garagna, S. What does it take to make a developmentally competent mammalian egg? Hum. Reprod. Update 2011, 17, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Ciccarelli, C.; Barberi, M.; Macchiarelli, G.; Canipari, R. Granulosa cell-oocyte interactions. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 115, S19–S22. [Google Scholar] [CrossRef] [PubMed]
- Pangas, S.A.; Rajkovic, A. Ovary as a Biomarker of Health and Longevity: Insights from Genetics. Semin. Reprod. Med. 2017, 35, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Rapino, C.; Di Nisio, V.; Rossi, G.; Maccarrone, M. The (endo)cannabinoid signaling in female reproduction: What are the latest advances? Prog. Lipid Res. 2020, 77, 101019. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Rossi, G.; Oddi, S.; Di Nisio, V.; Maccarrone, M. Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation. Int. J. Mol. Sci. 2019, 20, 2866. [Google Scholar] [CrossRef]
- Liu, Y.-X. Regulation of follicular development and differentiation by intra-ovarian factors and endocrine hormones. Front. Biosci. 2019, 24, 983–993. [Google Scholar] [CrossRef]
- Edson, M.A.; Nagaraja, A.K.; Matzuk, M.M. The Mammalian Ovary from Genesis to Revelation. Endocr. Rev. 2009, 30, 624–712. [Google Scholar] [CrossRef]
- Fritz, R.; Jindal, S. Reproductive aging and elective fertility preservation. J. Ovarian Res. 2018, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.V.; Nithila, P.; Reddy, S.J. Estimation of heavy metals in drinking water and development of heavy metal pollution index. J. Environ. Sci. Health Part A Environ. Sci. Eng. Toxicol. 1996, 31, 283–289. [Google Scholar] [CrossRef]
- Chowdhury, S.; Mazumder, M.J.; Al-Attas, O.; Husain, T. Heavy metals in drinking water: Occurrences, implications, and future needs in developing countries. Sci. Total Environ. 2016, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xun, P.; Kaufman, J.D.; Hayden, K.M.; Espeland, M.A.; Whitsel, E.A.; Serre, M.L.; Vizuete, W.; Orchard, T.; Harris, W.S.; et al. Erythrocyte omega-3 index, ambient fine particle exposure and brain aging. Neurology 2020, 95, e995–e1007. [Google Scholar] [CrossRef] [PubMed]
- Oken, E.; Belfort, M.B. Fish, fish oil, and pregnancy. JAMA 2010, 304, 1717–1718. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.M.; Egeland, G.M. Fish consumption, mercury exposure, and heart diseases. Nutr. Rev. 2004, 62, 68–72. [Google Scholar]
- Bloom, M.S.; Parsons, P.J.; Steuerwald, A.J.; Schisterman, E.F.; Browne, R.W.; Kim, K.; Coccaro, G.A.; Conti, G.C.; Narayan, N.; Fujimoto, V.Y. Toxic trace metals and human oocytes during in vitro fertilization (IVF). Reprod. Toxicol. 2010, 29, 298–305. [Google Scholar] [CrossRef]
- Nandi, P.; Banerjee, S.; Basu, S.; Das, M.; Dasgupta, P.; Varghese, A. A preliminary study on heavy metals (Lead, Cadmium and Arsenic) in follicular fluid and its effect on oocyte, embryo quality and outcome of IVF treatment. Fertil. Steril. 2011, 96, S155. [Google Scholar] [CrossRef]
- Aquino, N.B.; Sevigny, M.B.; Sabangan, J.; Louie, M.C. The Role of Cadmium and Nickel in Estrogen Receptor Signaling and Breast Cancer: Metalloestrogens or Not? J. Environ. Sci. Health Part C 2012, 30, 189–224. [Google Scholar] [CrossRef]
- Rzymski, P.; Tomczyk, K.; Rzymski, P.; Poniedziałek, B.; Opala, T.; Wilczak, M. Impact of heavy metals on the female reproductive system. Ann. Agric. Environ. Med. 2015, 22, 259–264. [Google Scholar] [CrossRef]
- Singh, L.; Anand, M.; Singh, S.; Taneja, A. Environmental toxic metals in placenta and their effects on preterm delivery-current opinion. Drug Chem. Toxicol. 2018, 43, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Myers, R.P.; Wei, T.; Bind, E.; Kassim, P.; Wang, G.; Ji, Y.; Hong, X.; Caruso, D.; Bartell, T.; et al. Placental transfer and concentrations of cadmium, mercury, lead, and selenium in mothers, newborns, and young children. J. Expo. Sci. Environ. Epidemiol. 2014, 24, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Wijnen, H.; Vader, H.; Kooistra, L.; Pop, V. Maternal selenium status during early gestation and risk for preterm birth. CMAJ 2011, 183, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Falcon, M.; Vinas, P.; Luna, A.; Romero, M.F. Placental lead and outcome of pregnancy. Toxicology 2003, 185, 59–66. [Google Scholar] [CrossRef]
- Llanos, M.N.; Ronco, A.M. Fetal growth restriction is related to placental levels of cadmium, lead and arsenic but not with antioxidant activities. Reprod. Toxicol. 2009, 27, 88–92. [Google Scholar] [CrossRef]
- Iyengar, G.V.; Rapp, A. Human placenta as a ‘dual’ biomarker for monitoring fetal and maternal environment with special reference to potentially toxic trace elements. Part 3: Toxic trace elements in placenta and placenta as a biomarker for these elements. Sci. Total Environ. 2001, 280, 221–238. [Google Scholar] [CrossRef]
- Kot, K.; Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Malinowski, W.; Szymański, S.; Mularczyk, M.; Tomska, N.; Rotter, I. Interactions between 14 Elements in the Human Placenta, Fetal Membrane and Umbilical Cord. Int. J. Environ. Res. Public Health 2019, 16, 1615. [Google Scholar] [CrossRef]
- Gilfillan, S.C. Lead Poisoning and The Fall of Rome. J. Occup. Med. Off. Publ. Ind. Med. Assoc. 1965, 7, 53–60. [Google Scholar]
- Paul, C. Etude sur l’intoxication lente par les preparations de plomb, de son influence sur le produit de la conception. Arch. General. Med. 1860, 15, 511–533. [Google Scholar]
- Foster, W.G. Reproductive toxicity of chronic lead exposure in the female cynomolgus monkey. Reprod. Toxicol. 1992, 6, 123–131. [Google Scholar] [CrossRef]
- Wide, M.; Wide, L. Estradiol receptor activity in uteri of pregnant mice given lead before implantation. Fertil. Steril. 1980, 34, 503–508. [Google Scholar] [CrossRef]
- Garg, S.K.; Sharma, A.; Choudhury, S.; Yadav, R.S.; Gupta, K.; Sood, N.; Nakade, U.P. Lead-induced adverse effects on the reproductive system of rats with particular reference to histopathological changes in uterus. Indian J. Pharmacol. 2015, 47, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Wide, M. Lead exposure on critical days of fetal life affects fertility in the female mouse. Teratology 1985, 32, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Wide, M.; D’Argy, R. Effect of inorganic lead on the primordial germ cells in the mouse embryo. Teratology 1986, 34, 207–212. [Google Scholar] [CrossRef]
- Davis, J.M.; Svendsgaard, D.J. Lead and child development. Nature 1987, 329, 297–300. [Google Scholar] [CrossRef]
- Hertz-Picciotto, I. The evidence that lead increases the risk for spontaneous abortion. Am. J. Ind. Med. 2000, 38, 300–309. [Google Scholar] [CrossRef]
- Faikoğlu, R.; Savan, K.; Utku, Ç.; Takar, N.; Zebitay, A.G. Significance of Maternal Plasma Lead Level in Early Pregnancy Loss. J. Environ. Sci. Health Part A 2006, 41, 501–506. [Google Scholar] [CrossRef]
- Valentino, M.; Coppa, G.; Ruschioni, A. Pregnancy in a worker exposed to lead. Med. Lav. 1984, 75, 296–299. [Google Scholar]
- Hu, H. Knowledge of diagnosis and reproductive history among survivors of childhood plumbism. Am. J. Public Health 1991, 81, 1070–1072. [Google Scholar] [CrossRef]
- Nordberg, G.F.; Mahaffey, K.R.; A Fowler, B. Introduction and summary. International workshop on lead in bone: Implications for dosimetry and toxicology. Environ. Health Perspect. 1991, 91, 3–7. [Google Scholar] [CrossRef]
- Gulson, B.L.; Mizon, K.J.; Korsch, M.J.; Palmer, J.M.; Donnelly, J.B. Mobilization of lead from human bone tissue during pregnancy and lactation—A summary of long-term research. Sci. Total Environ. 2003, 303, 79–104. [Google Scholar] [CrossRef]
- Weinberg, C.R.; Wilcox, A.J.; Baird, D.D. Reduced fecundability in women with prenatal exposure to cigarette smoking. Am. J. Epidemiol. 1989, 129, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Rahimzadeh, M.R.; Rahimzadeh, M.R.; Kazemi, S.; Moghadamnia, A.A. Cadmium toxicity and treatment: An update. Casp. J. Intern. Med. 2017, 8, 135–145. [Google Scholar]
- Akesson, A.; Berglund, M.; Schutz, A.; Bjellerup, P.; Bremme, K.; Vahter, M. Cadmium exposure in pregnancy and lactation in relation to iron status. Am. J. Public Health 2002, 92, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Berglund, M.; Akesson, A.; Nermell, B.; Vahter, M. Intestinal absorption of dietary cadmium in women depends on body iron stores and fiber intake. Environ. Health Perspect. 1994, 102, 1058–1066. [Google Scholar] [CrossRef]
- Bæcklund, M.; Pedersen, N.L.; Björkman, L.; Vahter, M. Variation in Blood Concentrations of Cadmium and Lead in the Elderly. Environ. Res. 1999, 80, 222–230. [Google Scholar] [CrossRef]
- Vahter, M.; Berglund, M.; Åkesson, A. Toxic metals and the menopause. Br. Menopause Soc. J. 2004, 10, 60–64. [Google Scholar] [CrossRef]
- Kawai, M.; Swan, K.F.; Green, A.E.; Edwards, D.E.; Anderson, M.B.; Henson, M.C. Placental endocrine disruption induced by cadmium: Effects on P450 cholesterol side-chain cleavage and 3beta-hydroxysteroid dehydrogenase enzymes in cultured human trophoblasts. Biol. Reprod. 2002, 67, 178–183. [Google Scholar] [CrossRef]
- Stoica, A.; Katzenellenbogen, B.S.; Martin, M.B. Activation of estrogen receptor-alpha by the heavy metal cadmium. Mol. Endocrinol. 2000, 14, 545–553. [Google Scholar]
- Debby, O.T. Effect of Cadmium on Female Reproduction and Treatment Options. Res. J. Obstet. Gynecol. 2018, 11, 41–48. [Google Scholar] [CrossRef]
- Henson, M.C.; Chedrese, P.J. Endocrine Disruption by Cadmium, a Common Environmental Toxicant with Paradoxical Effects on Reproduction. Exp. Biol. Med. 2004, 229, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Chirumbolo, S.; Dadar, M.; Pivina, L.; Lindh, U.; Butnariu, M.; Aaseth, J. Mercury exposure and its effects on fertility and pregnancy outcome. Basic Clin. Pharmacol. Toxicol. 2019, 125, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Kis, M.; Sipka, G.; Maróti, P. Stoichiometry and kinetics of mercury uptake by photosynthetic bacteria. Photosynth. Res. 2017, 132, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Baldi, F.; Filippelli, M.; Olson, G.J. Biotransformation of mercury by bacteria isolated from a river collecting cinnabar mine waters. Microb. Ecol. 1989, 17, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.-Y.; Kim, S.-J.; Kim, H.-G.; Lee, J.; Choi, Y.; Lee, H.; Kim, Y. Evaluation of estrogenicity of major heavy metals. Sci. Total Environ. 2003, 312, 15–21. [Google Scholar] [CrossRef]
- Verma, R.; Vijayalakshmy, K.; Chaudhiry, V. Detrimental impacts of heavy metals on animal reproduction: A review. J. Entomol. Zoo. Stud. 2018, 6, 27–30. [Google Scholar]
- Hurley, L.S.; Swenerton, H. Congenital malformations resulting from zinc deficiency in rats. Proc. Soc. Exp. Biol. Med. 1966, 123, 692–696. [Google Scholar] [CrossRef]
- Chow, B.F.; Sherwin, R.W.; Hsueh, A.M.; Blackwell, B.N.; Blackwell, R.Q. Growth and Development of Rats in Relation to the Maternal Diet: A Review1,2. Forum. Nutr. 1969, 11, 45–56. [Google Scholar] [CrossRef]
- Mashhadi, M.A.; Bakhshipour, A.; Zakeri, Z.; Moghadam, A.A. Reference Range for Zinc Level in Young Healthy Population in Southeast of Iran. Health Scope 2016, 6. [Google Scholar] [CrossRef]
- Beaver, L.M.; Truong, L.; Barton, C.L.; Chase, T.T.; Gonnerman, G.D.; Wong, C.P.; Tanguay, R.L.; Ho, E. Combinatorial effects of zinc deficiency and arsenic exposure on zebrafish (Danio rerio) development. PLoS ONE 2017, 12, e0183831. [Google Scholar] [CrossRef]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elements Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Durán, C.; Weisstaub, G. Zinc Supplementation and Growth of the Fetus and Low Birth Weight Infant. J. Nutr. 2003, 133, 1494S–1497S. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Diaz, F.J. Zinc Depletion Causes Multiple Defects in Ovarian Function during the Periovulatory Period in Mice. Endocrinology 2012, 153, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, L.O.; Harbak, H.; Bennekou, P. Cobalt metabolism and toxicology—A brief update. Sci. Total Environ. 2012, 432, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Lauwerys, R.; Lison, D. Health risks associated with cobalt exposure—An overview. Sci. Total Environ. 1994, 150, 1–6. [Google Scholar] [CrossRef]
- Philippe, J. Fertility and irradiation: A preconceptional investigation in teratology. Am. J. Obstet. Gynecol. 1975, 123, 714–718. [Google Scholar] [CrossRef]
- Bucher, J.R.; Elwell, M.R.; Thompson, M.B.; Chou, B.J.; Renne, R.; Ragan, H.A. Inhalation toxicity studies of cobalt sulfate in F344/N rats and B6C3F1 mice. Fundam. Appl. Toxicol. 1990, 15, 357–372. [Google Scholar] [CrossRef]
- Sengupta, P.; Banerjee, R.; Nath, S.; Das, S. Metals and female reproductive toxicity. Hum. Exp. Toxicol. 2014, 34, 679–697. [Google Scholar] [CrossRef]
- Forgacs, Z.; Massanyi, P.; Lukac, N.; Somosy, Z. Reproductive toxicology of nickel—Review. J. Environ. Sci. Health A Tox. Hazard Subst. Environ. Eng. 2012, 47, 1249–1260. [Google Scholar] [CrossRef]
- Forgacs, Z.; Paksy, K.; Varga, B.; Lazar, P.; Tatrai, E. Effects of NiSO4 on the ovarian function in rats. CEJOEM 1997, 3, 48–57. [Google Scholar]
- Carré, J.; Gatimel, N.; Moreau, J.; Parinaud, J.; Léandri, R. Does air pollution play a role in infertility? A systematic review. Environ. Health 2017, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Conforti, A.; Mascia, M.; Cioffi, G.; De Angelis, C.; Coppola, G.; De Rosa, P.; Pivonello, R.; Alviggi, C.; De Placido, G. Air pollution and female fertility: A systematic review of literature. Reprod. Biol. Endocrinol. 2018, 16, 117. [Google Scholar] [CrossRef] [PubMed]
- De Coster, S.; Van Larebeke, N. Endocrine-Disrupting Chemicals: Associated Disorders and Mechanisms of Action. J. Environ. Public Health 2012, 2012, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Palmerini, M.G.; Zhurabekova, G.; Balmagambetova, A.; Nottola, S.A.; Miglietta, S.; Belli, M.; Bianchi, S.; Cecconi, S.; Di Nisio, V.; Familiari, G.; et al. The pesticide Lindane induces dose-dependent damage to granulosa cells in an in vitro culture. Reprod. Biol. 2017, 17, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Iorio, R.; Castellucci, A.; Ventriglia, G.; Teoli, F.; Cellini, V.; Macchiarelli, G.; Cecconi, S. Ovarian toxicity: From environmental exposure to chemotherapy. Curr. Pharm. Des. 2014, 20, 5388–5397. [Google Scholar] [CrossRef] [PubMed]
- Paro, R.; Tiboni, G.M.; Buccione, R.; Rossi, G.; Cellini, V.; Canipari, R.; Cecconi, S. The fungicide mancozeb induces toxic effects on mammalian granulosa cells. Toxicol. Appl. Pharmacol. 2012, 260, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Zhang, Q. Associating ambient exposure to fine particles and human fertility rates in China. Environ. Pollut. 2018, 235, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Gaskins, A.J.; Mínguez-Alarcón, L.; Fong, K.C.; Abdelmessih, S.; Coull, B.A.; Chavarro, J.E.; Schwartz, J.; Kloog, I.; Souter, I.; Hauser, R.; et al. Exposure to Fine Particulate Matter and Ovarian Reserve Among Women from a Fertility Clinic. Epidemiology 2019, 30, 486–491. [Google Scholar] [CrossRef]
- Santi, D.; La Marca, A.; Michelangeli, M.; Casonati, A.; Grassi, R.; Baraldi, E.; Simoni, M. Ovarian reserve and exposure to environmental pollutants (ORExPo study). In Proceedings of the 21st European Congress of Endocrinology, Lyon, France, 18–21 May 2019. [Google Scholar]
- Maluf, M.; Perin, P.M.; Januário, D.A.N.F.; Saldiva, P.H.N. In vitro fertilization, embryo development, and cell lineage segregation after pre- and/or postnatal exposure of female mice to ambient fine particulate matter. Fertil. Steril. 2009, 92, 1725–1735. [Google Scholar] [CrossRef]
- Perin, P.M.; Maluf, M.; Czeresnia, C.E.; Januário, D.A.N.F.; Saldiva, P.H.N. Impact of short-term preconceptional exposure to particulate air pollution on treatment outcome in couples undergoing in vitro fertilization and embryo transfer (IVF/ET). J. Assist. Reprod. Genet. 2010, 27, 371–382. [Google Scholar] [CrossRef]
- Slama, R.; Bottagisi, S.; Solansky, I.; Lepeule, J.; Giorgis-Allemand, L.; Sram, R. Short-Term Impact of Atmospheric Pollution on Fecundability. Epidemiology 2013, 24, 871–879. [Google Scholar] [CrossRef]
- Nieuwenhuijsen, M.J.; Basagaña, X.; Dadvand, P.; Martinez, D.; Cirach, M.; Beelen, R.; Jacquemin, B. Air pollution and human fertility rates. Environ. Int. 2014, 70, 9–14. [Google Scholar] [CrossRef]
- Mahalingaiah, S.; Hart, J.; Laden, F.; Farland, L.; Hewlett, M.; Chavarro, J.; Aschengrau, A.; Missmer, S. Adult air pollution exposure and risk of infertility in the Nurses’ Health Study II. Hum. Reprod. 2016, 31, 638–647. [Google Scholar] [CrossRef]
- Yang, S.; Tan, Y.; Mei, H.; Wang, F.; Li, N.; Zhao, J.; Zhang, Y.; Qian, Z.; Chang, J.J.; Syberg, K.M.; et al. Ambient air pollution the risk of stillbirth: A prospective birth cohort study in Wuhan, China. Int. J. Hyg. Environ. Health 2018, 221, 502–509. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Q.; He, S.; Wu, K.; Ren, M.; Dong, H.; Di, J.; Yu, Z.; Huang, C. Ambient air pollution and gestational diabetes mellitus: A review of evidence from biological mechanisms to population epidemiology. Sci. Total. Environ. 2020, 719. [Google Scholar] [CrossRef]
- Kaufmann, P.; Mayhew, T.M.; Charnock-Jones, D.S. Aspects of human fetoplacental vasculogenesis and angiogenesis. II. Changes during normal pregnancy. Placenta 2004, 25, 114–126. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Development of the Human Placenta and Fetal Heart: Synergic or Independent? Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Kingdom, J.C.; Huppertz, B.; Seaward, G.; Kaufmann, P. Development of the placental villous tree and its consequences for fetal growth. Eur. J. Obstet. Gynecol. Reprod. Biol. 2000, 92, 35–43. [Google Scholar] [CrossRef]
- Rutland, C.; Mukhopadhyay, M.; Underwood, S.; Clyde, N.; Mayhew, T.M.; Mitchell, C.A. Induction of Intrauterine Growth Restriction by Reducing Placental Vascular Growth with the Angioinhibin TNP-470. Biol. Reprod. 2005, 73, 1164–1173. [Google Scholar] [CrossRef][Green Version]
- Veras, M.; Damaceno-Rodrigues, N.R.; Caldini, E.G.; Ribeiro, A.A.C.M.; Mayhew, T.M.; Saldiva, P.H.N.; Dolhnikoff, M. Particulate Urban Air Pollution Affects the Functional Morphology of Mouse Placenta1. Biol. Reprod. 2008, 79, 578–584. [Google Scholar] [CrossRef]
- Segal, T.R.; Giudice, L.C. Before the beginning: Environmental exposures and reproductive and obstetrical outcomes. Fertil. Steril. 2019, 112, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Practice Committee of the American Society for Reproductive Medicine. Smoking and infertility: A committee opinion. Fertil. Steril. 2018, 110, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Health Effects of Secondhand Smoke. Available online: www.lung.org/quit-smoking/smoking-facts/health-effects/secondhand-smoke (accessed on 13 July 2020).
- Harmful Chemicals in Tobacco Products. Available online: https://www.cancer.org/cancer/cancer-causes/tobacco-and-cancer/carcinogens-found-in-tobacco-products.html (accessed on 13 July 2020).
- Alviggi, C.; Guadagni, R.; Conforti, A.; Coppola, G.; Picarelli, S.; De Rosa, P.; Vallone, R.; Strina, I.; Pagano, T.; Mollo, A.; et al. Association between intrafollicular concentration of benzene and outcome of controlled ovarian stimulation in IVF/ICSI cycles: A pilot study. J. Ovarian Res. 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Mattison, D.R.; Donald, R.; Plowchalk, D.R.; David, R.; Meadows, M.J.; Miller, M.M.; Malek, A.; London, S. The Effect of Smoking on Oogenesis, Fertilization, and Implantation. Seminar. Reprodu. Med. 1989, 7, 291–304. [Google Scholar] [CrossRef]
- Furlong, H.C.; Stämpfli, M.R.; Gannon, A.; Foster, W. Identification of microRNAs as potential markers of ovarian toxicity. J. Appl. Toxicol. 2018, 38, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signaling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [PubMed]
- Hong, L.; Wang, Y.; Chen, W.; Yang, S. MicroRNA-508 suppresses epithelial-mesenchymal transition, migration, and invasion of ovarian cancer cells through the MAPK1/ERK signaling pathway. J. Cell. Biochem. 2018, 119, 7431–7440. [Google Scholar] [CrossRef] [PubMed]
- Damalas, C.A.; Koutroubas, S.D. Farmers’ Exposure to Pesticides: Toxicity Types and Ways of Prevention. Toxics 2016, 4, 1. [Google Scholar] [CrossRef]
- Rossi, G.; Palmerini, M.G.; Macchiarelli, G.; Buccione, R.; Cecconi, S. Mancozeb adversely affects meiotic spindle organization and fertilization in mouse oocytes. Reprod. Toxicol. 2006, 22, 51–55. [Google Scholar] [CrossRef]
- Cecconi, S.; Paro, R.; Rossi, G.; Macchiarelli, G. The Effects of the Endocrine Disruptors Dithiocarbamates on the Mammalian Ovary with Particular Regard to Mancozeb. Curr. Pharm. Des. 2007, 13, 2989–3004. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Buccione, R.; Baldassarre, M.; Macchiarelli, G.; Palmerini, M.G.; Cecconi, S. Mancozeb exposure in vivo impairs mouse oocyte fertilizability. Reprod. Toxicol. 2006, 21, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Palmerini, M.G.; Belli, M.; Nottola, S.A.; Miglietta, S.; Bianchi, S.; Bernardi, S.; Antonouli, S.; Cecconi, S.; Familiari, G.; Macchiarelli, G. Mancozeb impairs the ultrastructure of mouse granulosa cells in a dose-dependent manner. J. Reprod. Dev. 2018, 64, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Rossi, G.; Carta, G.; Di Luigi, G.; Cellini, V.; Canipari, R.U.C.; Buccione, R. Effects of trifluralin on the mouse ovary. Environ. Toxicol. 2011, 28, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Warner, G.R.; Li, Z.; Houde, M.L.; Atkinson, C.E.; Meling, D.D.; Chiang, C.; Flaws, J.A. Ovarian Metabolism of an Environmentally Relevant Phthalate Mixture. Toxicol. Sci. 2019, 169, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Ventrice, P.; Ventrice, D.; Russo, E.; De Sarro, G. Phthalates: European regulation, chemistry, pharmacokinetic and related toxicity. Environ. Toxicol. Pharmacol. 2013, 36, 88–96. [Google Scholar] [CrossRef]
- Krieg, S.A.; Shahine, L.K.; Lathi, R.B. Environmental exposure to endocrine-disrupting chemicals and miscarriage. Fertil. Steril. 2016, 106, 941–947. [Google Scholar] [CrossRef]
- Heudorf, U.; Mersch-Sundermann, V.; Angerer, J. Phthalates: Toxicology and exposure. Int. J. Hyg. Environ. Health 2007, 210, 623–634. [Google Scholar] [CrossRef]
- Lovekamp-Swan, T.; Davis, B.J. Mechanisms of phthalate ester toxicity in the female reproductive system. Environ. Health Perspect. 2003, 111, 139–145. [Google Scholar] [CrossRef]
- Thayer, K.A.; Doerge, D.R.; Hunt, D.; Schurman, S.H.; Twaddle, N.C.; Churchwell, M.I.; Garantziotis, S.; Kissling, G.E.; Easterling, M.R.; Bucher, J.R.; et al. Pharmacokinetics of bisphenol A in humans following a single oral administration. Environ. Int. 2015, 83, 107–115. [Google Scholar] [CrossRef]
- Zhang, T.; Shen, W.; De Felici, M.; Zhang, X.-F. Di(2-ethylhexyl)phthalate: Adverse effects on folliculogenesis that cannot be neglected. Environ. Mol. Mutagen. 2016, 57, 579–588. [Google Scholar] [CrossRef]
- Patiño-García, D.; Cruz-Fernandes, L.; Buñay, J.; Palomino, J.; Moreno, R.D. Reproductive Alterations in Chronically Exposed Female Mice to Environmentally Relevant Doses of a Mixture of Phthalates and Alkylphenols. Endocrinology 2017, 159, 1050–1061. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Zhou, C.; Rattan, S.; Flaws, J.A. Effects of Endocrine-Disrupting Chemicals on the Ovary. Biol. Reprod. 2015, 93, 20. [Google Scholar] [CrossRef] [PubMed]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: Effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Pallotti, F.; Pelloni, M.; Gianfrilli, D.; Lenzi, A.; Lombardo, F.; Paoli, D. Mechanisms of Testicular Disruption from Exposure to Bisphenol A and Phtalates. J. Clin. Med. 2020, 9, 471. [Google Scholar] [CrossRef] [PubMed]
- Craig, Z.R.; Wang, W.; A Flaws, J. Endocrine-disrupting chemicals in ovarian function: Effects on steroidogenesis, metabolism and nuclear receptor signaling. Reproduction 2011, 142, 633–646. [Google Scholar] [CrossRef]
- Krotz, S.P.; Carson, S.A.; Tomey, C.; Buster, J.E. Phthalates and bisphenol do not accumulate in human follicular fluid. J. Assist. Reprod. Genet. 2012, 29, 773–777. [Google Scholar] [CrossRef]
- Ikezuki, Y.; Tsutsumi, O.; Takai, Y.; Kamei, Y.; Taketani, Y. Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. 2002, 17, 2839–2841. [Google Scholar] [CrossRef]
- Palioura, E.; Diamanti-Kandarakis, E. Polycystic ovary syndrome (PCOS) and endocrine disrupting chemicals (EDCs). Rev. Endocr. Metab. Disord. 2015, 16, 365–371. [Google Scholar] [CrossRef]
- Hu, Y.; Wen, S.; Yuan, D.; Peng, L.; Zeng, R.; Yang, Z.; Liu, Q.; Xu, L.; Kang, D. The association between the environmental endocrine disruptor bisphenol A and polycystic ovary syndrome: A systematic review and meta-analysis. Gynecol. Endocrinol. 2018, 34, 370–377. [Google Scholar] [CrossRef]
- Soave, I.; Occhiali, T.; Assorgi, C.; Marci, R.; Caserta, D. Environmental toxin exposure in polycystic ovary syndrome women and possible ovarian neoplastic repercussion. Curr. Med. Res. Opin. 2020, 36, 693–703. [Google Scholar] [CrossRef]
- Paoli, D.; Pallotti, F.; Dima, A.P.; Albani, E.; Alviggi, C.; Causio, F.; Dioguardi, C.C.; Conforti, A.; Ciriminna, R.; Fabozzi, G.; et al. Phthalates and Bisphenol A: Presence in Blood Serum and Follicular Fluid of Italian Women Undergoing Assisted Reproduction Techniques. Toxics 2020, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Sumner, R.N.; Tomlinson, M.; Craigon, J.; England, G.C.W.; Lea, R.G. Independent and combined effects of diethylhexyl phthalate and polychlorinated biphenyl 153 on sperm quality in the human and dog. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Effects of heat stress on mammalian reproduction. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3341–3350. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M. Heat stress on reproductive function and fertility in mammals. Reprod. Med. Biol. 2012, 11, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Z.; Sui, H.-S.; Miao, D.-Q.; Liu, N.; Zhou, P.; Ge, L.; Tan, J.-H. Effects of heat stress during in vitro maturation on cytoplasmic versus nuclear components of mouse oocytes. Reproduction 2009, 137, 181–189. [Google Scholar] [CrossRef]
- Zheng, H.-Y.; Yang, C.-Y.; Yu, N.-Q.; Huang, J.-X.; Zheng, W.; Abdelnour, S.A.; Shang, J.-H. Effect of season on the in-vitro maturation and developmental competence of buffalo oocytes after somatic cell nuclear transfer. Environ. Sci. Pollut. Res. 2020, 27, 7729–7735. [Google Scholar] [CrossRef]
- Hozyen, H.F.; Ahmed, H.H.; Shalaby, S.I.A.; Essawy, G.E.S. Seasonal Heat Stress Effect on Cholesterol, Estradiol and Progesterone during Follicular Development in Egyptian Buffalo. Int. J. Biol. Life Agric. Sci. 2016, 9. [Google Scholar]
- Matsuzuka, T.; Ozawa, M.; Nakamura, A.; Ushitani, A.; Hirabayashi, M.; Kanai, Y. Effects of Heat Stress on the Redox Status in the Oviduct and Early Embryonic Development in Mice. J. Reprod. Dev. 2005, 51, 281–287. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Bauer, M. Heat shock proteins in porcine ovary: Synthesis, accumulation and regulation by stress and hormones. Cell Stress Chaperon 2010, 16, 379–387. [Google Scholar] [CrossRef]
- Li, B.; Weng, Q.; Liu, Z.; Shen, M.; Zhang, J.; Wu, W.; Liu, H. Selection of antioxidants against ovarian oxidative stress in mouse model. J. Biochem. Mol. Toxicol. 2017, 31, e21997. [Google Scholar] [CrossRef]
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G. National, Regional, and Global Trends in Infertility Prevalence Since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.; Qiu, X.; Jin, L.; Ma, J.; Li, Z.; Zhang, L.; Zhu, H.; Finnell, R.H.; Zhu, T. Association of selected persistent organic pollutants in the placenta with the risk of neural tube defects. Proc. Natl. Acad. Sci. USA 2011, 108, 12770–12775. [Google Scholar] [CrossRef] [PubMed]
- Zona, A.; Iavarone, I.; Buzzoni, C.; Conti, S.; Santoro, M.; Fazzo, L.; Pasetto, R.; Pirastu, R.; Bruno, C.; Ancona, C.; et al. SENTIERI: Epidemiological Study of Residents in National Priority Contaminated Sites. Fifth Report. Epidemiol. Prev. 2019, 43, 1–208. [Google Scholar] [PubMed]
- Bellingham, M.; Fowler, P.A.; Macdonald, E.S.; Mandon-Pepin, B.; Cotinot, C.; Rhind, S.; Sharpe, R.M.; Evans, N.P. Timing of Maternal Exposure and Foetal Sex Determine the Effects of Low-level Chemical Mixture Exposure on the Foetal Neuroendocrine System in Sheep. J. Neuroendocr. 2016, 28, 12. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canipari, R.; De Santis, L.; Cecconi, S. Female Fertility and Environmental Pollution. Int. J. Environ. Res. Public Health 2020, 17, 8802. https://doi.org/10.3390/ijerph17238802
Canipari R, De Santis L, Cecconi S. Female Fertility and Environmental Pollution. International Journal of Environmental Research and Public Health. 2020; 17(23):8802. https://doi.org/10.3390/ijerph17238802
Chicago/Turabian StyleCanipari, Rita, Lucia De Santis, and Sandra Cecconi. 2020. "Female Fertility and Environmental Pollution" International Journal of Environmental Research and Public Health 17, no. 23: 8802. https://doi.org/10.3390/ijerph17238802
APA StyleCanipari, R., De Santis, L., & Cecconi, S. (2020). Female Fertility and Environmental Pollution. International Journal of Environmental Research and Public Health, 17(23), 8802. https://doi.org/10.3390/ijerph17238802