The Prevention of Bio-Organic Fertilizer Fermented from Cow Manure Compost by Bacillus sp. XG-1 on Watermelon Continuous Cropping Barrier

Abstract
1. Introduction
2. Materials and Methods
2.1. Media, Chemicals, Compost and Soil
2.2. The Isolation and Identification of Antagonistic Bacillus Strains against Fon
2.3. The Antagonistic Effect against Mycelial Growth and Conidia Germination of Fon
2.4. The Growth Promotion and Investigation of Promotion Substances in XG-1
2.5. The Investigation of Solid State Fermentation Conditions of XG-1
2.6. Pot Experiments Analysis for Preventing the Watermelon Continuous Cropping Barrier
2.7. Statistical Analysis
3. Results
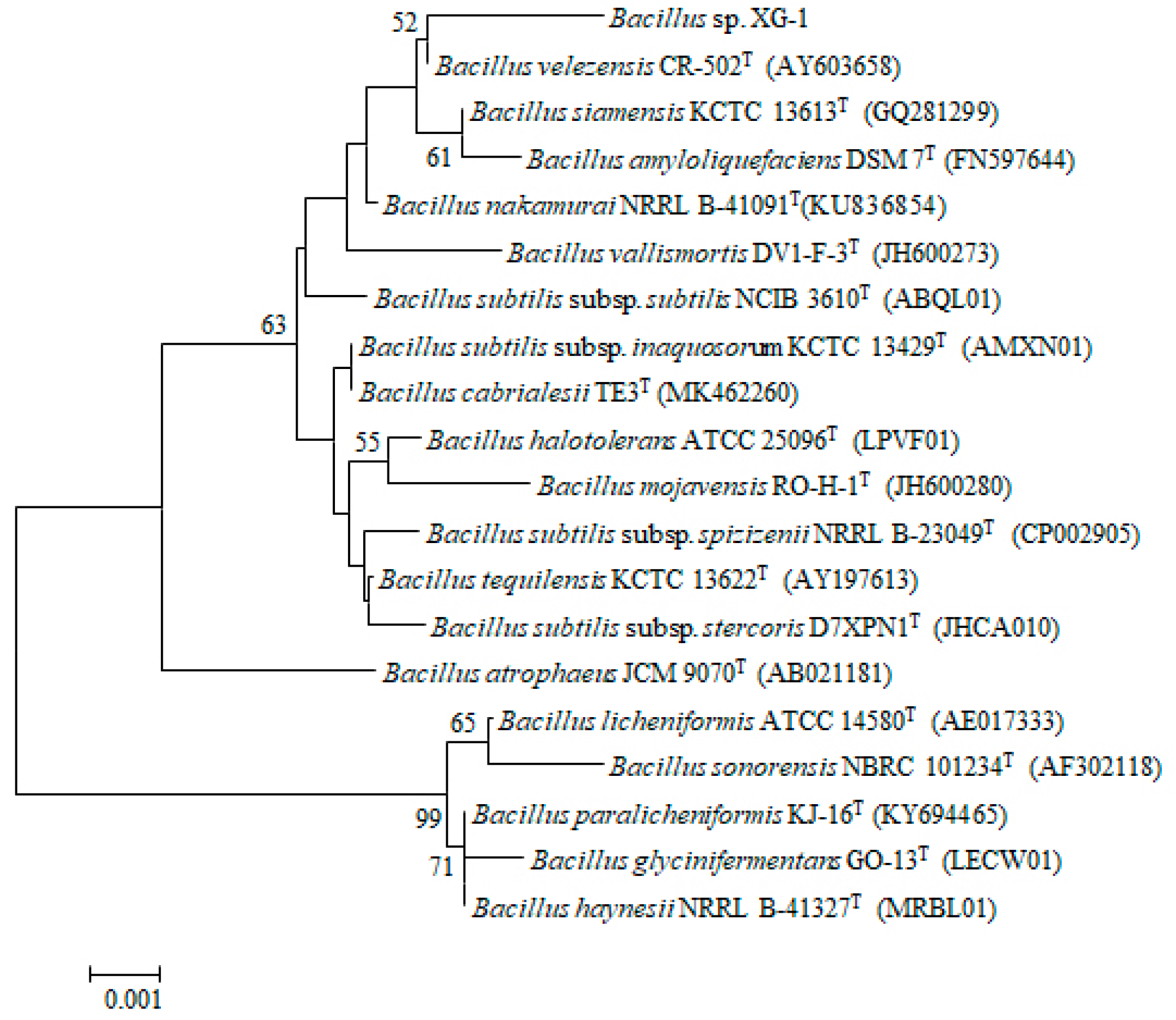
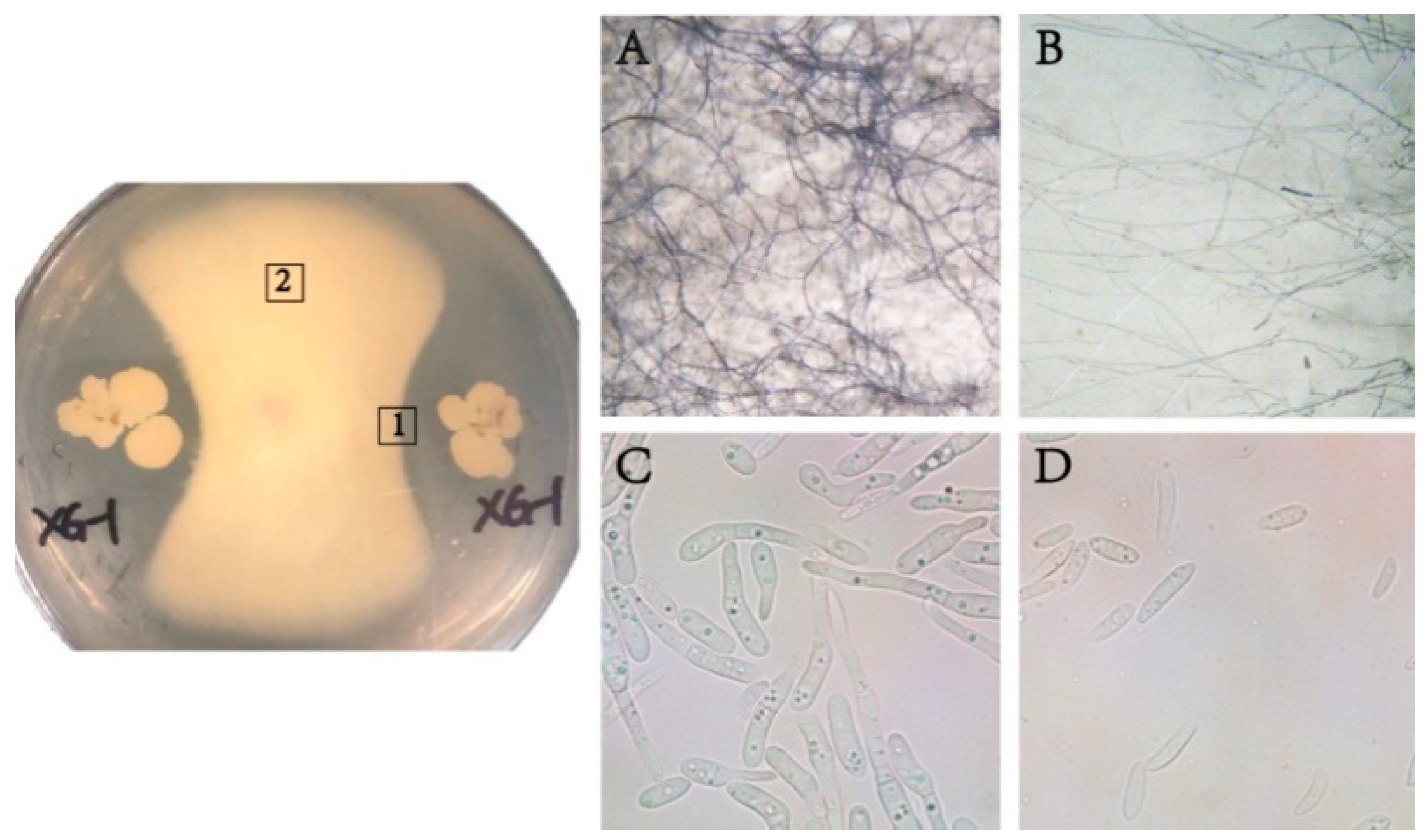
3.1. Strain Isolation and Identification
3.2. The Inhibition of Mycelial Growth and Conidia Germination of Fon by XG-1
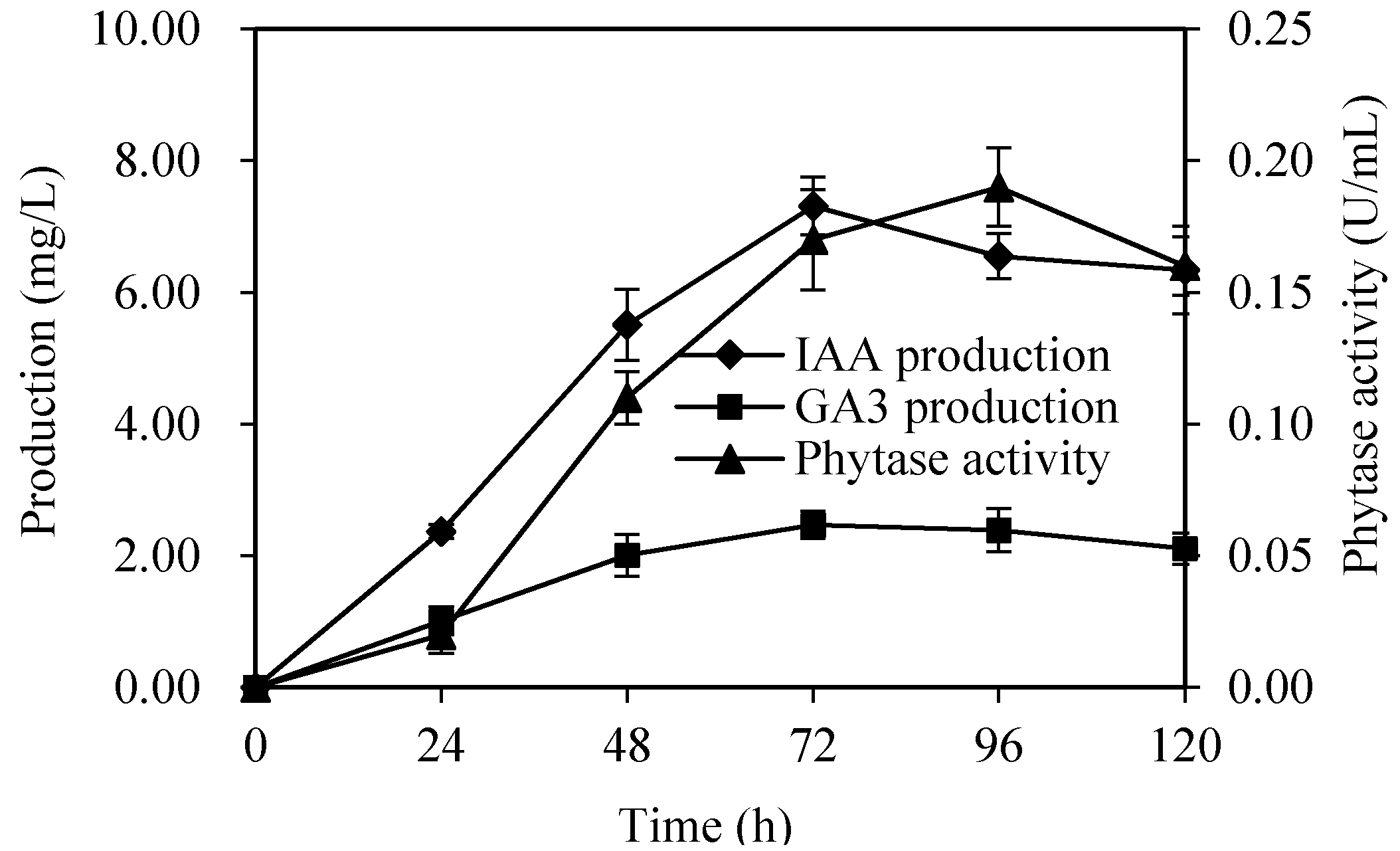
3.3. The Investigation of Growth-Promotion Substances in XG-1
3.4. Preparation of Bio-Organic Fertilizer
3.5. Pot Analysis on the Prevention for the Watermelon Continuous Cropping Barrier by Bio-Organic Fertilizer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wei, X.; Zhao, Y.; Yang, R. Advances on watermelon continuous cropping obstacles and its prevention measures under protective cultivation. China Cucurbits Veg. 2016, 29, 1–5. [Google Scholar]
- Zhang, X.; Huo, Z.; Wu, Z.; Cheng, Z. Analysis on the causes of continuous cropping barrier of watermelon and its control strategies. Agric. Sci. Technol. 2015, 1, 180–185. [Google Scholar]
- Huang, C.; Bu, Y.; Shan, Z.; Dai, C. Research advances in mechanisms of watermelon continuous cropping diseases and its biological control. Chin. J. Ecol. 2016, 35, 1670–1676. [Google Scholar]
- Wang, K. Analysis of obstacles and overcoming measures of continuous watermelon cropping. China Fruit Veg. 2019, 39, 70–72. [Google Scholar]
- Faheem, M.; Raza, W.; Zhong, W.; Nan, Z.; Xu, Y. Evaluation of the biocontrol potential of Streptomyces goshikiensis YCXU against Fusarium oxysporum f. sp. niveum. Biol. Control 2014, 81, 101–110. [Google Scholar] [CrossRef]
- Dickinson, M. Plant fungal pathogens: Methods and Protocols. Plant Pathol. 2012, 61, 1187. [Google Scholar] [CrossRef]
- Wang, B.; Yuan, J.; Zhang, J.; Shen, Z.; Zhang, M.; Li, R.; Ruan, Y.; Shen, Q. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fert. Soils 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, D.; Wang, D.; Miao, Y.; Shao, J.; Zhou, X.; Xu, Z.; Li, Q.; Feng, H.; Li, S.; et al. Whole transcriptomic analysis of the plant-beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 during enhanced biofilm formation regulated by maize root exudates. Bmc Genom. 2015, 16, 685. [Google Scholar] [CrossRef]
- Xie, S.; Jiang, H.; Ding, T.; Xu, Q.; Chai, W.; Cheng, B. Bacillus amyloliquefaciens FZB42 represses plant miR846 to induce systemic resistance via a jasmonic acid-dependent signalling pathway. Mol. Plant Pathol. 2018, 19, 1612–1623. [Google Scholar] [CrossRef]
- Ling, N.; Raza, W.; Ma, J.; Huang, Q.; Shen, Q. Identification and role of organic acids in watermelon root exudates for recruiting Paenibacillus polymyxa SQR-21 in the rhizosphere. Eur. J. Soil Biol. 2011, 47, 374–379. [Google Scholar] [CrossRef]
- Arseneault, T.; Goyer, C.; Filion, M. Phenazine production by Pseudomonas sp. LBUM223 contributes to the biological control of potato common scab. Phytopathology 2013, 103, 995–1000. [Google Scholar] [CrossRef]
- Law, J.W.; Ser, H.; Khan, T.M.; Chuah, L.; Pusparajah, P.; Chan, K.; Goh, B.; Lee, L. The potential of Streptomyces as biocontrol agents against the rice blast Fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, F.; Huang, L.; Zhou, Y. Isolation, identification of biocontrol bacterium and its antagonistic activity against Fusarium oxyspirum f. sp. niveum. Acta Agric. Univ. Jiangxi 2013, 35, 324–328. [Google Scholar]
- Xu, W.; Wang, K.; Wang, H.; Liu, Z. Evaluation of the biocontrol potential of Bacillus sp. WB against Fusarium oxysporum f. sp. niveum. Biol. Control 2020, 147. [Google Scholar] [CrossRef]
- Xu, W.; Wang, H.; Lv, Z.; Shi, Y.; Wang, Z. Antifungal activity and functional components of cell-free supernatant from Bacillus amyloliquefaciens LZN01 inhibit Fusarium oxysporum f. sp. niveum growth. Biotechnol. Biotechnol. Equip. 2019, 33, 1042–1052. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Liang, H.; Huang, J.; Chen, Z.; Nie, Y. The rhizosphere microbial community response to a bio-organic fertilizer: Finding the mechanisms behind the suppression of watermelon Fusarium wilt disease. Acta Physiol. Plant. 2018, 40, 1–14. [Google Scholar] [CrossRef]
- Jia, Z.; Jiang, L.; Hong, L.; Jing, H.; Zhe, C.; Nie, Y.; Wang, C.; Wang, Y.; He, Z. Manipulation of the rhizosphere microbial community through application of a new bio-organic fertilizer improves watermelon quality and health. PLoS ONE 2018, 13, e192967. [Google Scholar]
- Biao, S. Optimization of solid-state fermentation conditions for antagonistic Bacillus subtilis SQR9 producing bio-organic fertilizer. Chin. J. Appl. Environ. Biol. 2013, 19, 90–95. [Google Scholar]
- Huang, Y.; Sun, L.; Zhao, J.; Huang, R.; Li, R.; Shen, Q. Utilization of different waste proteins to create a novel PGPR-containing bio-organic fertilizer. Sci. Rep. 2015, 5, 7766. [Google Scholar] [CrossRef]
- Ye, X.; Li, Z.; Luo, X.; Wang, W.; Li, Y.; Li, R.; Zhang, B.; Qiao, Y.; Zhou, J.; Fan, J.; et al. A predatory myxobacterium controls cucumber Fusarium wilt by regulating the soil microbial community. Microbiome 2020, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.K.; Duan, Y.; Awasthi, S.K.; Liu, T.; Zhang, Z. Effect of biochar and bacterial inoculum additions on cow dung composting. Bioresour. Technol. 2020, 297, 122407. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Yang, X.; Fan, J.; Miao, W.; Ling, N.; Xu, Y.; Huang, Q.; Shen, Q. Suppression of Fusarium wilt of watermelon by a bio-organic fertilizer containing combinations of antagonistic microorganisms. Biocontrol 2009, 54, 287–300. [Google Scholar] [CrossRef]
- Ling, N.; Zhang, W.; Tan, S.; Huang, Q.; Shen, Q. Effect of the nursery application of bioorganic fertilizer on spatial distribution of Fusarium oxysporum f. sp. niveum and its antagonistic bacterium in the rhizosphere of watermelon. Appl. Soil Ecol. 2012, 59, 13–19. [Google Scholar]
- Raza, W.; Yuan, J.; Ling, N.; Huang, Q.; Shen, Q. Production of volatile organic compounds by an antagonistic strain Paenibacillus polymyxa WR-2 in the presence of root exudates and organic fertilizer and their antifungal activity against Fusarium oxysporum f. sp. niveum. Biol. Control 2015, 80, 89–95. [Google Scholar] [CrossRef]
- Landy, M.; Warren, G. Bacillomycin, an antibiotic from Bacillus subtilis, active against pathogenic fungi. Proc Soc Exp Biol Med. P. Soc. Exp. Biol. Med. 1948, 67, 539–541. [Google Scholar] [CrossRef]
- Whitman, W. Systematic Bacteriology. Asian Austral. J. Anim. Sci. 2013, 26, 1237–1246. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.S.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Shao, J.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Contribution of indole-3-acetic acid in the plant growth promotion by the rhizospheric strain Bacillus amyloliquefaciens SQR9. Biol. Fertil. Soils 2015, 51, 321–330. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Van Themaat, E.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Wang, Y.; Zheng, X. Molecular detection of Fusarium oxysporum f. sp. niveum and Mycosphaerella melonis in infected plant tissues and soil. Fems Microbiol. Lett. 2005, 249, 39–47. [Google Scholar]
- Zhang, H.; Chen, F.; Zhao, H.Z.; Lu, J.S.; Zhao, M.J.; Hong, Q.; Huang, X. Colonization on cucumber root and enhancement of chlorimuron-ethyl degradation in rhizosphere by Hansschlegelia zhihuaiae S113 and root exudates. J. Agric. Food Chem. 2018, 66, 4584–4591. [Google Scholar] [CrossRef]
- Weert, S.; Vermeiren, H.; Mulders, I.; Kuiper, I.; Lugtenberg, B. Flagella-driven chemotaxis towards exudate components is an important trait for tomato root colonization by Pseudomonas fluorescens. Mol. Plant Microbe Interact. 2002, 15, 1173–1180. [Google Scholar] [CrossRef]
- Kim, O.; Cho, Y.; Lee, K.; Yoon, S.; Kim, M.; Na, H.; Park, S.; Jeon, Y.; Lee, J.; Yi, H. Introducing EzTaxon-e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef]
- Huang, L.; Song, L.; Xia, X.; Mao, W. Plant-soil feedbacks and soil sickness: From mechanisms to application in agriculture. J. Chem. Ecol. 2013, 39, 232–242. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, B.; Lin, S.; Li, X.; Ye, X. Accumulation of cinnamic acid and vanillin in eggplant root exudates and the relationship with continuous cropping obstacle. Afr. J. Biotechnol. 2011, 10, 2659–2665. [Google Scholar]
- Xie, X.; Chen, Y.; Bu, Y.; Dai, C.D. A review of allelopathic researches on phenolic acids. Acta Ecol. Sin. 2014, 34, 6417–6428. [Google Scholar]
- Hao, W.; Ren, L.; Ran, W.; Shen, Q. Allelopathic effects of root exudates from watermelon and rice plants on Fusarium oxysporum f.sp. niveum. Plant Soil 2010, 336, 485–497. [Google Scholar] [CrossRef]
- Lv, W.; Yang, G.; Li, S.; Zhang, C.; Zhu, Y.; Zheng, X.; Shen, Q. Effects of phenolic acids on watermelon seed germination, seedling growth, and physiological and biochemical properties. Acta Agric. Shanghai 2013, 29, 1–4. [Google Scholar]
- Naima, B.; Ibtissem, B.; Mahmoud, M. Evaluation of the efficiency of Trichoderma, Penicillium, and Aspergillus species as biological control agents against four soil-borne fungi of melon and watermelon. Egypt. J. Biol. Pest Control 2018, 28, 25–37. [Google Scholar]
- Wang, X.; Zhang, G.; Li, R.; Lu, Y.; Ran, W.; Shen, Q. Control of watermelon Fusarium wilt by using antagonist-enhanced biological organic fertilizers. Plant. Nutr. Fert. Sci. 2013, 19, 223–231. [Google Scholar]
- Bashan, Y.; Holguin, G.; Bashan, D. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances (1997–2003). Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Faheem, M.; Yousaf, S.; Rajer, F.; Yamin, M. Volatile and non-volatile antifungal compounds produced by Trichoderma harzianum SQR-T037 suppressed the growth of Fusarium oxysporum f. sp. niveum. Sci. Lett. 2013, 1, 21–24. [Google Scholar]
- Chen, X.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.; Reva, O. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Ning, L.; Huang, Q.; Guo, S.; Shen, Q. Paenibacillus polymyxa SQR-21 systemically affects root exudates of watermelon to decrease the conidial germination of Fusarium oxysporum f.sp. niveum. Plant. Soil 2011, 341, 485–493. [Google Scholar]
- Xu, Z.; Shao, J.; Li, B.; Yan, X.; Shen, Q.; Zhang, R. Contribution of Bacillomycin D in Bacillus amyloliquefaciens SQR9 to antifungal activity and biofilm formation. Appl. Environ. Microb. 2013, 79, 808–815. [Google Scholar] [CrossRef]
- Zhao, M.; Li, M.; Wang, M.; Wang, Y.; Zhang, X. Effects of watermelon replanting on main microflora of rhizosphere and activities of soil enzymes. Microbiology 2008, 35, 1251–1254. [Google Scholar]
- Zhang, M.; Li, R.; Cao, L.; Shi, J.; Liu, H.; Huang, Y.; Shen, Q. Algal sludge from Taihu Lake can be utilized to create novel PGPR-containing bio-organic fertilizers. J. Environ. Manag. 2014, 132, 230–236. [Google Scholar] [CrossRef]
- Chen, H.; Awasthi, S.K.; Liu, T.; Duan, Y.; Ren, X.; Zhang, Z.; Pandey, A.; Awasthi, M.K. Effects of microbial culture and chicken manure biochar on compost maturity and greenhouse gas emissions during chicken manure composting. J. Hazard. Mater. 2020, 389, 121908. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Factor | ||
|---|---|---|---|
| A: Temperature (°C) | B: Water Content (%) | C: Number of Turning (/d) | |
| 1 | 25 | 45 | 0 |
| 2 | 30 | 50 | 1 |
| 3 | 37 | 55 | 2 |
| Project | Factor | Yi | ||
|---|---|---|---|---|
| A | B | C | ||
| 1 | 1 | 1 | 1 | 1.99 |
| 2 | 1 | 2 | 2 | 2.91 |
| 3 | 1 | 3 | 3 | 2.55 |
| 4 | 2 | 1 | 2 | 4.18 |
| 5 | 2 | 2 | 3 | 4.62 |
| 6 | 2 | 3 | 1 | 3.24 |
| 7 | 3 | 1 | 3 | 3.99 |
| 8 | 3 | 2 | 1 | 3.9 |
| 9 | 3 | 3 | 2 | 3.78 |
| kj1 | 7.45 | 10.16 | 9.13 | Yi |
| kj2 | 12.04 | 11.43 | 10.87 | |
| kj3 | 11.67 | 9.57 | 11.16 | |
| Kj1 | 2.48 | 3.39 | 3.04 | |
| Kj2 | 4.01 | 3.81 | 3.62 | |
| Kj3 | 3.89 | 3.19 | 3.72 | |
| Rj | 1.80 | 0.74 | 1.01 | |
| Optimal level | A2 | B2 | C2 | |
| Optimal project | A2B2C3 | |||
| Treatment | Disease Incidence (%) | Strain XG-1 (50 d) | Fon (50 d) | |||
|---|---|---|---|---|---|---|
| 25 d | 50 d | Rhizosphere Soil (105 CFU/g) | Bulk Soil (105 CFU/g) | Rhizosphere Soil (Log10 copies g−1 soil) | Bulk Soil (Log10 copies g−1 soil) | |
| S1 | 37.5 ± 3.1 b | 79.2 ± 4.9 b | - | - | 5.06 ± 0.22 b | 4.97 ± 0.24 b |
| S2 | - | - | - | - | - | - |
| S1 + BOF | 9.2 ± 2.1 a | 10.8 ± 1.5 a | 9.79 ± 0.22 a | 4.97 ± 0.15 a | 3.11 ± 0.12 a | 3.93 ± 0.21 a |
| S2 + BOF | - | - | 9.91 ± 0.18 a | 4.75 ± 0.11 a | - | - |
| S1 + CMC | 35.0 ± 2.9 b | 77.5 ± 5.1 b | - | - | 4.88 ± 0.20 b | 4.93 ± 0.17 b |
| S2 + CMC | - | - | - | - | - | - |
| Treatment | Seedling Height (cm) | Root Length (cm) | Fresh Weight of Seedling (g) | Fresh Weight of Root (g) |
|---|---|---|---|---|
| S1 | 24.0 ± 3.1 a | 11.5 ± 3.0 a | 8.31 ± 0.48 a | 0.56 ± 0.11 a |
| S2 | 38.3 ± 5.7 c | 14.3 ± 3.9 b | 13.47 ± 0.39 b | 0.81 ± 0.12 b |
| S1 + BOF | 42.9 ± 4.3c d | 19.8 ± 5.1 c | 15.34 ± 0.62 bc | 1.10 ± 0.27 c |
| S2 + BOF | 44.1 ± 5.1 d | 21.0 ± 3.3 c | 17.11 ± 1.41 c | 1.19 ± 0.37 c |
| S1 + CMC | 29.9 ± 3.3 b | 12.9 ± 2.2 ab | 10.02 ± 1.06 a | 0.67 ± 0.12 a |
| S2 + CMC | 40.2 ± 3.2 c | 18.9 ± 2.6 c | 16.37 ± 1.21 c | 1.05 ± 0.36 bc |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Hua, Z.-W.; Liang, W.-Z.; Niu, Q.-H.; Wang, X. The Prevention of Bio-Organic Fertilizer Fermented from Cow Manure Compost by Bacillus sp. XG-1 on Watermelon Continuous Cropping Barrier. Int. J. Environ. Res. Public Health 2020, 17, 5714. https://doi.org/10.3390/ijerph17165714
Zhang H, Hua Z-W, Liang W-Z, Niu Q-H, Wang X. The Prevention of Bio-Organic Fertilizer Fermented from Cow Manure Compost by Bacillus sp. XG-1 on Watermelon Continuous Cropping Barrier. International Journal of Environmental Research and Public Health. 2020; 17(16):5714. https://doi.org/10.3390/ijerph17165714
Chicago/Turabian StyleZhang, Hao, Zi-Wei Hua, Wen-Zhi Liang, Qiu-Hong Niu, and Xiang Wang. 2020. "The Prevention of Bio-Organic Fertilizer Fermented from Cow Manure Compost by Bacillus sp. XG-1 on Watermelon Continuous Cropping Barrier" International Journal of Environmental Research and Public Health 17, no. 16: 5714. https://doi.org/10.3390/ijerph17165714
APA StyleZhang, H., Hua, Z.-W., Liang, W.-Z., Niu, Q.-H., & Wang, X. (2020). The Prevention of Bio-Organic Fertilizer Fermented from Cow Manure Compost by Bacillus sp. XG-1 on Watermelon Continuous Cropping Barrier. International Journal of Environmental Research and Public Health, 17(16), 5714. https://doi.org/10.3390/ijerph17165714